1. Introduction
The global agricultural sector faces unprecedented challenges in the 21st century, including the need to ensure food security for a growing population while minimizing environmental impacts and adapting to climate change [
1,
2]. Conventional monoculture farming systems, characterized by high input dependency and simplified biodiversity, have been associated with soil degradation, nutrient leaching, and reduced ecosystem services [
3,
4]. In response to these challenges, there is a growing interest in ecological intensification approaches that harness natural processes and ecosystem services to enhance agricultural productivity and sustainability [
5,
6].
Integrated farming systems, which combine multiple agricultural activities within a single production unit, have emerged as a promising strategy for ecological intensification [
7,
8]. These systems aim to optimize resource use efficiency, enhance biodiversity, and improve overall system resilience [
9]. Among various integrated approaches, the rice-fish co-culture system has garnered significant attention, particularly in Asia, due to its potential to increase food production, improve farmer livelihoods, and enhance ecosystem services [
10,
11].
Building upon traditional rice-fish systems, an innovative rice-loach-shrimp (RLS) integrated farming model has been developed in Shaoxing, Zhejiang Province, China [
12]. This system integrates rice cultivation with the production of pond loach (
Misgurnus anguillicaudatus) and freshwater shrimp (
Macrobrachium nipponense), two high-value aquatic species well-adapted to rice field environments. The RLS system operates on the principle of ecological synergy, where the aquatic animals contribute to nutrient cycling and pest control, while the rice provides shelter and supplementary food for the animals. This multi-trophic integration aims to create a more balanced and resilient agroecosystem, potentially offering enhanced ecological and economic benefits compared to conventional rice monoculture (RM) systems [
13,
14].
However, the complex interactions between soil properties, nutrient dynamics, and microbial communities in such multi-trophic integrated systems remain poorly understood. Soil microorganisms play crucial roles in nutrient cycling, organic matter decomposition, and the maintenance of soil structure, directly influencing ecosystem functioning and crop productivity [
15,
16]. The introduction of aquatic animals into rice fields may significantly alter soil physicochemical properties and microbial habitats, potentially leading to shifts in microbial community structure and function [
17,
18].
Understanding these intricate soil-microbial interactions is essential for optimizing the management of integrated farming systems and maximizing their ecological benefits. Traditional univariate approaches often fail to capture the full complexity of these relationships, necessitating the application of advanced multivariate statistical techniques [
19,
20]. Multivariate analyses offer promising tools for exploring the multifaceted nature of soil-microbial interactions in integrated farming systems. These methods may help reveal patterns and relationships that are less apparent when variables are examined in isolation, potentially providing a more comprehensive view of the soil ecosystem dynamics.
In this context, our study aims to investigate the complex soil-microbial interactions in the RLS integrated farming system compared to conventional RM using a comprehensive multivariate analysis approach. Specifically, we seek to: (1) Compare the correlations among soil physicochemical properties, enzyme activities, and microbial populations in RLS and RM systems. (2) Identify the key factors driving soil microbial community structure and abundance using Principal Component Analysis (PCA) and Random Forest modeling. (3) Develop and compare structural equation models (SEM) to quantify the direct and indirect relationships among soil properties, nutrient dynamics, enzyme activities, and microbial populations in both farming systems.
Our study builds upon a five-year field experiment conducted in Shaoxing, China, comparing the RLS integrated system with conventional RM. This long-term dataset provides a unique opportunity to examine the temporal dynamics of soil-microbial interactions and assess the stability of potential benefits associated with the integrated farming approach. By employing advanced statistical techniques such as correlation analysis, Principal Component Analysis, Random Forest modeling, and structural equation modeling, we aim to provide a holistic understanding of the soil ecosystem functioning in the RLS integrated system. This knowledge will not only contribute to the optimization of the RLS model but also inform the development of sustainable agricultural practices that enhance soil health, ecosystem services, and food production.
2. Materials and Methods
2.1. Study Area and Experimental Design
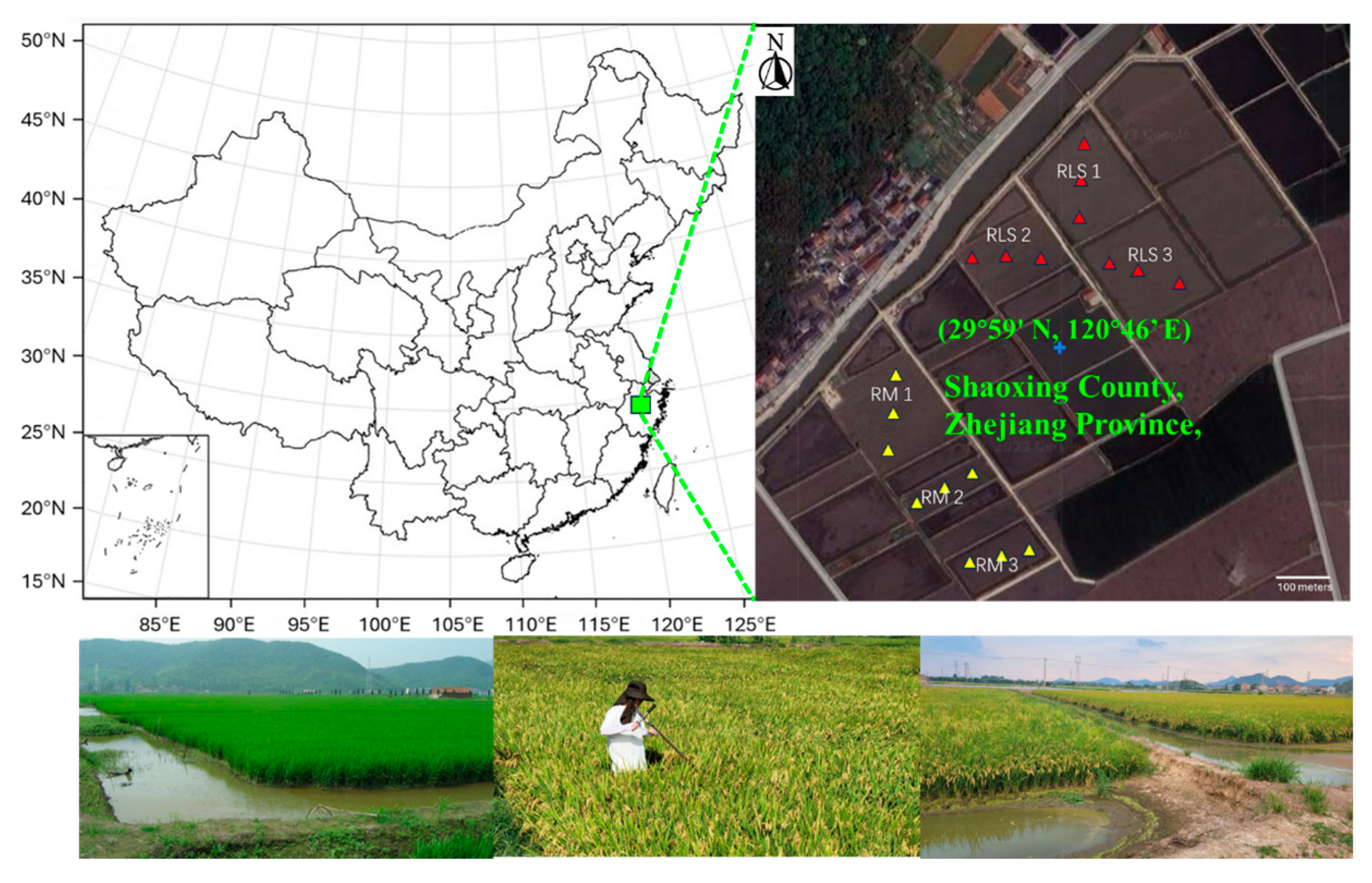
The study was conducted from 2017 to 2021 in paddy fields located at the rice-fish ecological co-culture demonstration area in Shaoxing County, Zhejiang Province, China (29°59′ N, 120°46′ E;
Figure 1). The area is characterized by a subtropical monsoon climate with a mean annual temperature of 16.4 °C and average annual precipitation of 1400 mm. The soil is classified as a silty clay loam; soil texture and fertility are representative of the paddy soils in the region.
We employed a randomized complete block design with two treatments: Rice-Loach-Shrimp integrated farming system (RLS) and conventional Rice Monoculture (RM). Each treatment was replicated in three fields, ranging from 0.2 to 0.7 ha in size. The RLS system integrated the cultivation of rice (Oryza sativa) with pond loach and freshwater shrimp aquaculture, while the RM system consisted of conventional double-cropping rice cultivation.
In the RLS fields, two adjacent peripheral trenches (2 m wide and 0.5 m deep) were excavated along the field edges to accommodate aquatic animals. The agricultural cycle began with rice seeding in early April, followed by the introduction of juvenile loaches (5–8 cm in length, 75,000 individuals ha−1) in mid-May. After the rice harvest in late July, the paddies were inundated, and shrimp post-larvae (1.5–2 cm in length, 600,000 individuals ha−1) were then released into these water-filled fields. Feeding commences one-week post-shrimp introduction, utilizing a commercial feed (30% crude protein, 5% crude lipid) administered twice daily at 0630 and 1600 h. Feed rations, adjusted to 3–5% of estimated aquatic biomass, are modulated based on environmental conditions, growth rates, and feed consumption observed on feeding trays after a 1-h period. Selective harvesting of market-size shrimp and loach initiates in early October using 1.5 cm mesh trap nets, with sub-marketable individuals retained to sustain the system until the following March.
The RM fields adhered to local conventional practices throughout the growing seasons, without the integration of aquatic animals.
During the early rice season, both RLS and RM systems employed identical practices for rice seeding, fertilizer application, and herbicide use. Insecticides were not applied in either system specifically during the early rice stage.
2.2. Soil Sampling and Analysis
Soil samples were collected annually in mid-July from 2017 to 2021, one week before the early rice harvest. In each field, three composite samples were obtained from the rice-growing area along a diagonal transect. Each composite sample consisted of 6–9 subsamples collected from the 0–20 cm depth using a 5 cm diameter soil auger. Samples were sealed in plastic bags and stored at 4 °C until analysis.
Soil chemical properties were analyzed following standard methods [
21]. For soil chemical analysis, air-dried soil sample (0.5 kg) was ground and passed through a 0.15 mm sieve. Soil pH was measured in a 1:2.5 soil-water suspension. Soil organic matter (SOM) was determined by the potassium dichromate wet oxidation method. Total nitrogen (TN) was measured using the Kjeldahl method, while total phosphorus (TP) was quantified by molybdenum blue colorimetry following H₂SO₄-HClO₄ digestion. Available nitrogen (Na) was determined by the alkali diffusion method, and available phosphorus (Pa) was extracted using 0.5 M NaHCO₃ and measured by molybdenum blue colorimetry.
Soil enzyme activities were assayed on fresh soil samples. Soil moisture content was determined by oven-drying subsamples at 105 °C for 48 h, with all results expressed on an oven-dry weight basis. Urease (UR) activity was determined by the indophenol blue colorimetric method, acid phosphatase (ACP) activity by the p-nitrophenol colorimetric method, and catalase (CA) activity by titration with potassium permanganate [
22]. Soil respiration (Resp) was quantified by measuring CO₂ evolution using the alkali absorption method [
23].
Soil microbial populations were assessed using the dilution plate count method [
24]. Culturable bacteria (Bact), fungi (Fung), and actinomycetes (Actino) were isolated using beef-peptone agar, Czapek’s agar, and starch-casein agar, respectively. The numbers of these were counted by the dilution–plate method [
25].
2.3. Statistical Analysis
To unravel the complex soil-microbial interactions in the RLS and RM systems, we employed a suite of multivariate statistical techniques. All statistical analyses were performed using R version 4.4.0 [
26]. Differences were considered significant at
p < 0.05.
2.3.1. Correlation Analysis
Pearson correlation coefficients were calculated between all pairs of soil physicochemical properties, enzyme activities, and microbial populations for both RLS and RM systems. The correlation matrices were visualized using heatmaps generated with the ‘corrplot’ package (version 0.92) [
27].
2.3.2. Principal Component Analysis (PCA)
PCA was performed to explore the relationships among soil properties, nutrient dynamics, and microbial communities in both farming systems. The analysis was conducted using the ‘FactoMineR’ package (version 2.11) [
28]. Variables were standardized prior to analysis, and biplots were generated to visualize the relationships between variables and observations.
2.3.3. Random Forest Modeling
Random forest models were constructed to identify the key factors influencing microbial populations in both systems. Separate models were built for bacterial, fungal, actinobacterial, and total microbial populations using the ‘randomForest’ package (version 4.7-1.1) [
29]. The importance of predictor variables was assessed using the percent increase in mean squared error (%IncMSE) when each variable was permuted.
2.3.4. Structural Equation Modeling (SEM)
SEM was employed to quantify the direct and indirect relationships among soil properties, nutrient dynamics, enzyme activities, and microbial populations in both systems. The models were constructed using the ‘lavaan’ package (version 0.6-18) [
30]. Model fit was assessed using multiple indices, including the Comparative Fit Index (CFI), Tucker-Lewis Index (TLI), Root Mean Square Error of Approximation (RMSEA), and Standardized Root Mean Square Residual (SRMR).
3. Results
3.1. Soil Health Indicators and Their Correlations in RLS and RM Systems
Descriptive statistics and correlation analysis revealed both similarities and differences in soil health indicators between the Rice-Loach-Shrimp (RLS) integrated farming system and the Rice Monoculture (RM) system (
Table 1,
Figure 2).
Soil nutrient indicators showed strong positive correlations in both systems, with Total Nitrogen (TN) strongly correlated with Total Phosphorus (TP) in RLS (r = 0.91, p < 0.001) and RM (r = 0.86, p < 0.001). However, the RLS system exhibited higher mean values for TN (0.871 vs. 0.842 g/kg), Available Nitrogen (Na) (50.856 vs. 46.924 mg/kg soil), and Available Phosphorus (Pa) (24.015 vs. 17.786 mg/kg soil) compared to RM.
Soil Organic Matter (SOM) demonstrated significant positive correlations with most soil health indicators in both systems. The RLS system showed higher mean SOM content (38.951 vs. 32.956 g/kg) and stronger correlations with enzyme activities, particularly Urease (UR) (r = 0.89, p < 0.001) and Catalase (CA) (r = 0.91, p < 0.001).
Enzyme activities and microbial populations were generally higher in the RLS system. Mean values for UR (4.255 vs. 3.625 mg/g), CA (21.724 vs. 19.439 mL/g), and Acid Phosphatase (ACP) (34.298 vs. 22.067 mg/g dried soil) were notably higher in RLS. Similarly, bacterial (28.123 vs. 19.699 × 105 cells/g dried soil), fungal (2.699 vs. 2.048 × 102 cells/g dried soil), and actinomycete (2.167 vs. 0.915 × 105 cells/g dried soil) populations were more abundant in RLS.
Soil pH showed weak correlations with most indicators in both systems. However, the RLS system had a slightly higher mean pH (6.264 vs. 5.976) and exhibited weaker correlations between pH and other indicators compared to RM. Soil respiration (Resp) was higher in RLS (mean 2.052 vs. 1.658 mg CO2/g dried soil), indicating potentially higher microbial activity. Resp showed moderate correlations with most soil health indicators in both systems.
These results suggest that while both RLS and RM systems maintain similar correlation patterns among soil health indicators, the RLS system generally exhibits higher values for nutrient content, organic matter, enzyme activities, and microbial populations. This may indicate enhanced soil fertility and biological activity in the integrated farming system, possibly due to the influence of aquatic organisms on soil processes.
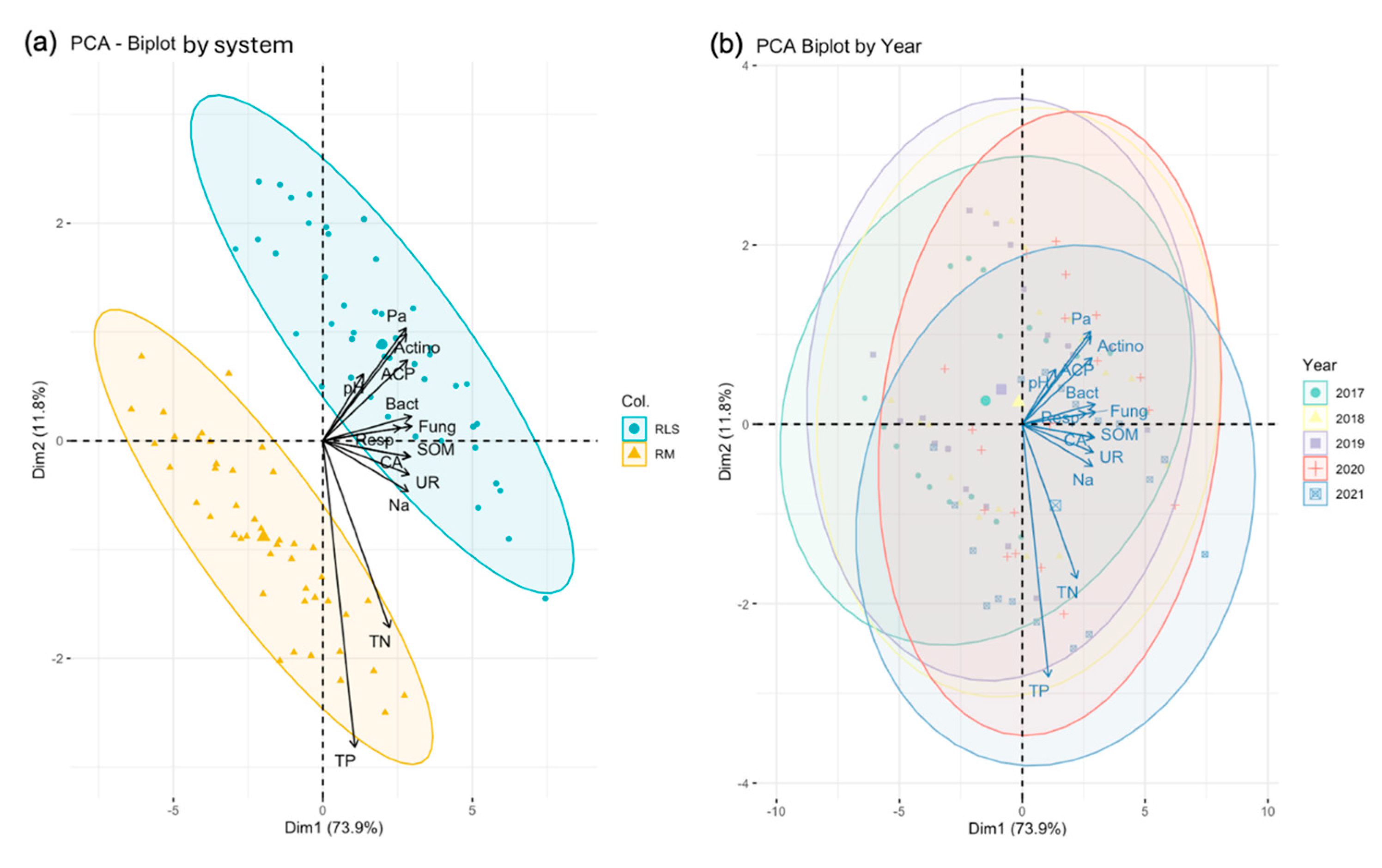
3.2. Principal Component Analysis of Soil-Microbial Interactions
Principal Component Analysis (PCA) revealed distinct patterns of soil-microbial interactions between the RLS and RM systems (
Figure 3). The first two principal components explained 85.8% of the total variance (PC1: 73.9%, PC2: 11.9%), effectively capturing the majority of the variation in the measured soil physicochemical properties and microbial indicators.
The RLS and RM systems formed clearly separated clusters along the first principal component (Dim1), indicating significant differences in soil properties and microbial communities between the two systems (
Figure 3a). The RLS system was positively associated with available phosphorus (Pa), soil organic matter (SOM), enzyme activities (UR, CA, ACP), and microbial populations (Bact, Fung, Actino). In contrast, the RM system showed stronger association with total nitrogen (TN) and total phosphorus (TP).
Temporal analysis (
Figure 3b) revealed relatively consistent patterns of soil-microbial interactions from 2017 to 2021, with overlapping confidence ellipses across years. This suggests that the differences between RLS and RM systems were stable and persistent throughout the five-year study period.
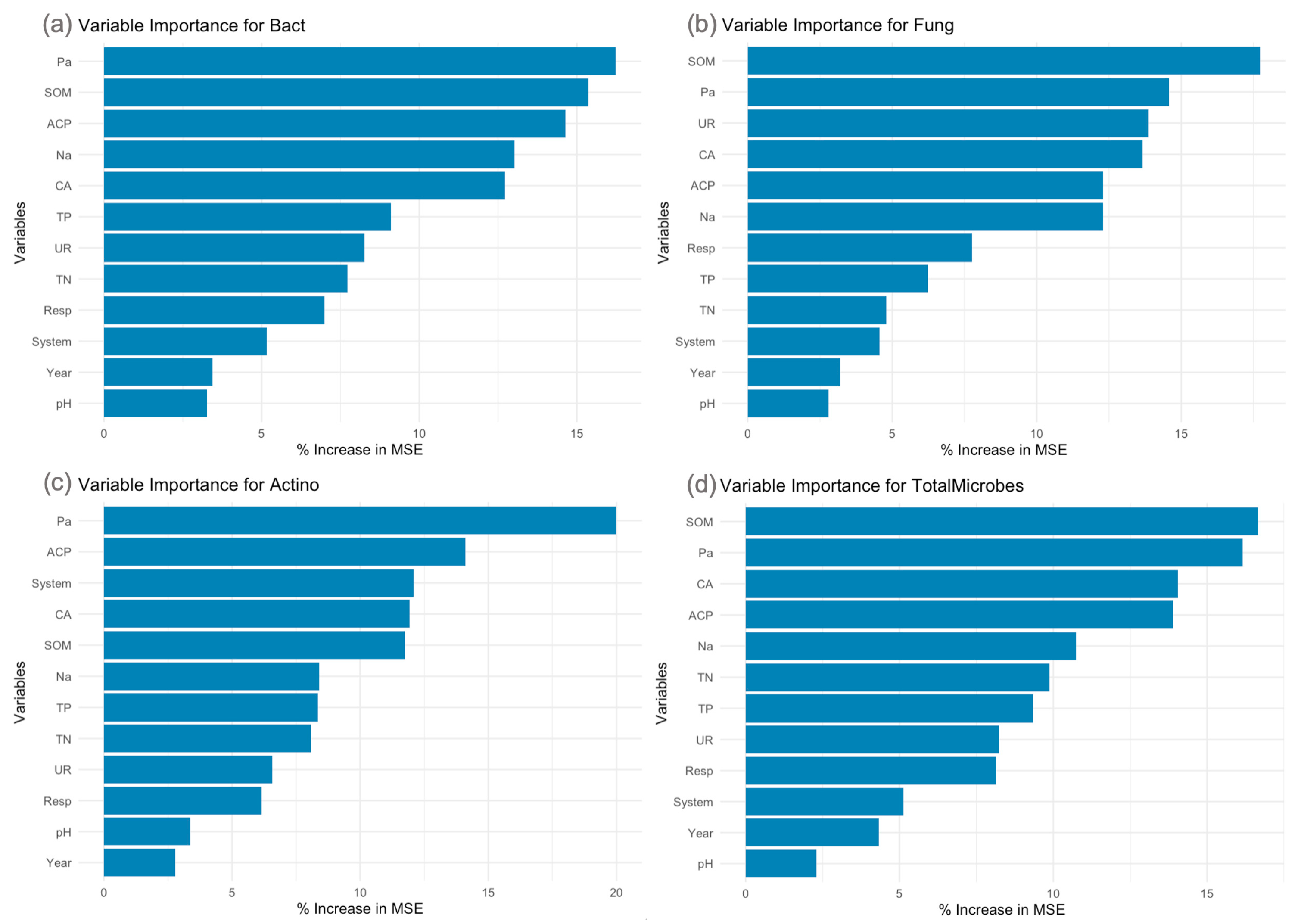
3.3. Random Forest Analysis of Soil-Microbial Interactions
Random forest models demonstrated high predictive power for microbial populations, with R-squared values of 0.972 for bacteria, 0.955 for fungi, 0.879 for actinobacteria, and 0.977 for total microbial population (
Figure 4).
For bacterial populations, available phosphorus (Pa) and soil organic matter (SOM) emerged as the most critical predictors, followed by acid phosphatase activity (ACP) and available nitrogen (Na). Fungal populations were primarily influenced by SOM and Pa, followed by urease (UR) and catalase (CA) activities. Actinobacterial populations were overwhelmingly predicted by available phosphorus (Pa), followed by ACP activity and the farming system type.
The total microbial population showed a balanced influence from various factors, with SOM, Pa, CA activity, and ACP activity all contributing significantly. Notably, the farming system type consistently appeared as a moderately important factor across all microbial groups.
Comparison of the two farming systems revealed higher levels of available nutrients, soil organic matter, and enzyme activities in the RLS system. This was accompanied by substantially higher microbial populations in the RLS system, resulting in a 46.9% increase in total microbial biomass compared to the RM system.
3.4. Structural Equation Modeling of Soil-Microbial Interactions
Structural equation modeling (SEM) revealed complex interactions among soil properties, nutrient dynamics, enzyme activities, and microbial populations in the two systems (
Figure 5). Both models demonstrated moderate fit, with similar fit indices for RLS (CFI = 0.878, TLI = 0.839, RMSEA = 0.212, SRMR = 0.046) and RM (CFI = 0.888, TLI = 0.852, RMSEA = 0.208, SRMR = 0.051) systems.
In both systems, soil nutrients exhibited strong positive loadings on their indicator variables (TN, TP, Na, Pa, SOM), with standardized coefficients ranging from 0.874 to 0.940 in the RLS system and 0.875 to 0.980 in the RM system. The relationship between soil nutrients and enzyme activity was strongly positive in both systems (RLS: β = 0.972, p < 0.001; RM: β = 0.978, p < 0.001).
Soil pH showed a significant negative effect on enzyme activity in both systems (RLS: β = −0.134, p = 0.013; RM: β = −0.150, p = 0.028). The impact of soil nutrients on microbial population was positive and significant in both systems, but the effect was stronger in the RM system (β = 0.835, p < 0.001) compared to the RLS system (β = 0.740, p = 0.001).
Notably, the covariance between soil nutrients and soil pH was significant in the RM system (standardized covariance = 0.408, p = 0.014) but not in the RLS system (standardized covariance = 0.114, p = 0.456), suggesting that the integrated RLS system may decouple the relationship between soil nutrients and pH.
Enzyme activities showed a strong positive effect on microbial populations in both systems (RLS: β = 0.307, p < 0.001; RM: β = 0.140, p = 0.015), with a notably stronger effect in the RLS system.
4. Discussion
4.1. Soil Health and Nutrient Dynamics in Integrated Farming Systems
Our multivariate analyses reveal that the Rice-Loach-Shrimp (RLS) integrated farming system exhibits generally higher values for key soil health indicators compared to conventional Rice Monoculture (RM), although the overall correlation patterns are similar. The stronger correlations among soil health indicators in the RLS system, particularly between soil organic matter (SOM), enzyme activities, and microbial populations, suggest a more tightly coupled and potentially more resilient soil ecosystem [
31]. This aligns with the ecological intensification principle, where increased biological complexity can enhance ecosystem functioning and stability [
5].
The pronounced positive associations between SOM and microbial parameters in the RLS system provide valuable insights. These relationships indicate that the integrated system may be more effective at building and maintaining soil organic matter, which serves as a crucial energy source and habitat for soil microorganisms [
32]. The presence of aquatic animals in the RLS system likely contributes to this effect through the input of organic residues and the stimulation of microbial activity via bioturbation [
33,
34].
The consistency of soil-microbial interaction patterns over the five-year study period, as revealed by the PCA, suggests that the benefits of the RLS system in terms of enhanced soil health and microbial diversity are not transient but persist over time. This temporal stability is crucial for the long-term sustainability of agricultural systems and aligns with the findings of other long-term studies on integrated farming approaches [
35,
36].
Further studies should investigate the persistence of soil health benefits when transitioning from RLS to conventional rice monoculture systems, as this information would be valuable for farmers considering adopting or discontinuing integrated farming practices.
4.2. Nutrient Dynamics and Microbial Community Structure
The PCA and random forest analyses consistently identified available phosphorus (Pa) and SOM as key drivers of microbial community structure and abundance in the RLS system. This finding underscores the critical role of phosphorus in shaping soil microbial communities, particularly in aquatic-terrestrial interface ecosystems like rice paddies [
37]. The higher levels of available phosphorus in the RLS system may be attributed to the feeding and excretion activities of loach and shrimp, which can enhance nutrient cycling and availability [
10].
The strong influence of soil enzyme activities, particularly acid phosphatase (ACP), on microbial populations in the RLS system suggests a potential positive feedback loop between microbial activity and nutrient availability. Enhanced enzyme activities can increase nutrient mineralization rates, supporting larger and more diverse microbial communities, which in turn produce more enzymes [
38]. This synergistic relationship may contribute to the overall improved soil health observed in the RLS system.
Our results show weak correlation between soil pH and most indicators in both systems. However, the RLS system had a slightly higher mean pH (6.264 vs. 5.976) and exhibited weaker correlations between pH and other indicators compared to RM. This suggests that the integrated farming approach may create a more stable pH environment, potentially buffering against pH-induced changes in nutrient availability and microbial activity. The decoupling of the relationship between soil nutrients and pH in the RLS system, as revealed by the SEM analysis, further supports this hypothesis and warrants further investigation.
4.3. Mechanisms Underlying Enhanced Soil Health in RLS
The structural equation modeling (SEM) results provide insights into the potential mechanisms driving improved soil health in the RLS system. The strong positive relationship between soil nutrients and enzyme activities, coupled with the significant influence of these factors on microbial populations, suggests a cascading effect where improved nutrient status enhances microbial activity, leading to increased enzyme production and further nutrient cycling [
39].
The differing strengths of relationships between soil nutrients and microbial populations in the RLS and RM systems, as revealed by SEM, indicate that the integrated system may foster more complex ecological interactions. This complexity could contribute to the higher microbial diversity and biomass observed in the RLS system, potentially enhancing ecosystem resilience and functioning [
40].
These findings align with the concept of ecological stoichiometry, which posits that the balance of elements in an ecosystem influences its structure and function [
41]. The RLS system appears to create a more balanced and efficient nutrient cycle, supporting a more diverse and active microbial community. This enhanced microbial activity may, in turn, contribute to improved nutrient use efficiency and crop productivity, as observed in other integrated farming systems [
11].
4.4. Implications for Sustainable Agriculture
Our findings have significant implications for the development of sustainable agricultural practices. The RLS system demonstrates the potential of ecological intensification approaches to enhance soil health, microbial diversity, and potentially, long-term agricultural sustainability. By fostering a more diverse and active soil microbiome, such integrated systems may reduce the need for external inputs, enhance nutrient use efficiency, and improve crop resilience to environmental stresses [
42].
The identified key drivers of soil-microbial interactions in the RLS system, such as available phosphorus and soil organic matter, provide valuable insights for the management of integrated farming systems. Practices that enhance these factors, such as optimizing aquatic animal stocking densities or incorporating crop residues, could be pivotal in maximizing the benefits of integrated approaches.
Furthermore, the RLS system’s ability to maintain higher levels of soil organic matter and microbial biomass could contribute to carbon sequestration and climate change mitigation in agricultural landscapes [
43]. This aspect of integrated farming systems warrants further investigation, particularly in the context of developing climate-smart agricultural practices.
4.5. Limitations and Future Directions
While our study provides comprehensive insights into soil-microbial interactions in the RLS system, some limitations should be addressed in future research. First, our analysis focused on culturable microbial populations, which represent only a fraction of the total soil microbiome. Future studies employing metagenomic approaches could provide a more complete picture of microbial community structure and function [
44].
Subsequent research should also consider a more comprehensive approach to understanding the microbial ecology of RLS systems. This could include evaluating the gut microbiomes of fish and shrimp, and assessing their interactions with soil microorganisms. Additionally, analyzing different nitrogen compounds in the water could help trace correlations with soil organic matter content, further elucidating nutrient cycling processes in RLS systems. These approaches would offer a more complete picture of the ecological dynamics in integrated aquaculture-agriculture systems.
Additional studies should investigate the effects of RLS systems on blue-green algae populations and activity, given their crucial role in rice ecosystems. Investigating the impacts of the RLS system on broader ecosystem services, such as greenhouse gas emissions and biodiversity conservation, would provide a more holistic understanding of its environmental implications.
Economic analyses comparing the RLS system with conventional rice monoculture would also be crucial for assessing its viability and potential for widespread adoption. These diverse research directions will contribute to a more comprehensive understanding of RLS systems and their potential role in sustainable agriculture.
5. Conclusions
This five-year study, employing advanced multivariate analyses, suggests that the Rice-Loach-Shrimp (RLS) integrated farming system generally exhibits higher values for key soil health indicators compared to conventional Rice Monoculture (RM). The RLS system demonstrates enhanced soil organic matter content, nutrient availability, enzyme activities, and microbial populations, with available phosphorus and soil organic matter emerging as primary drivers of microbial community structure. Structural equation modeling indicated complex interactions among soil properties, enzyme activities, and microbial populations. These results support the potential of RLS as an effective ecological intensification strategy for rice production, promoting a more sustainable soil ecosystem. As agriculture faces the challenges of food security and environmental impact, integrated farming systems like RLS offer a promising pathway towards sustainable food production. Future research should explore the broader implications of RLS across diverse agro-ecological zones to fully realize its potential.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}