Recovery of Soil Hydraulic Properties for Assisted Passive and Active Restoration: Assessing Historical Land Use and Forest Structure

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
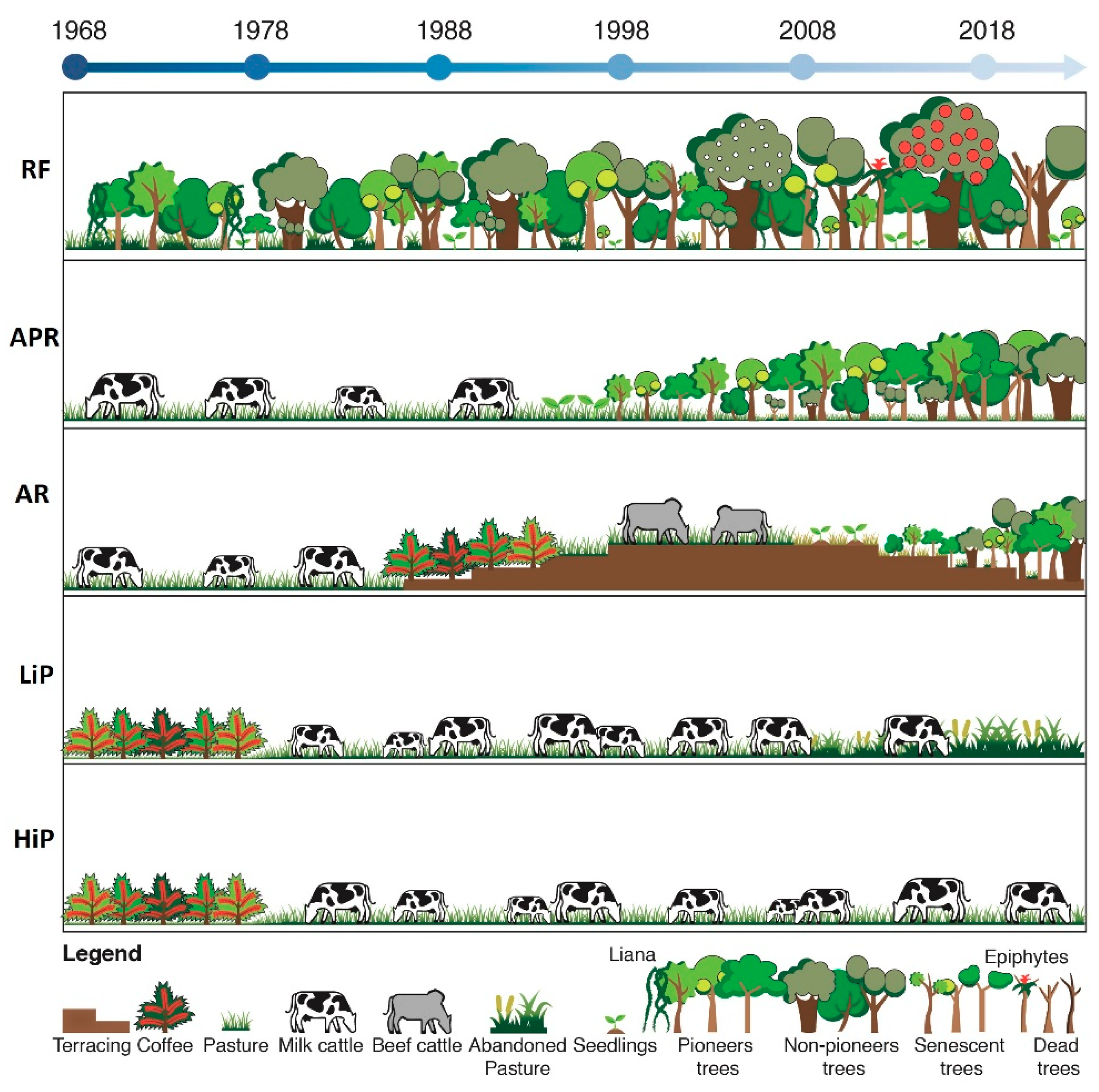
2.2. Experimental Design
2.3. Vegetation Sampling
2.4. Soil Sampling
2.5. Data Analysis
3. Results
3.1. Vegetation Attributes
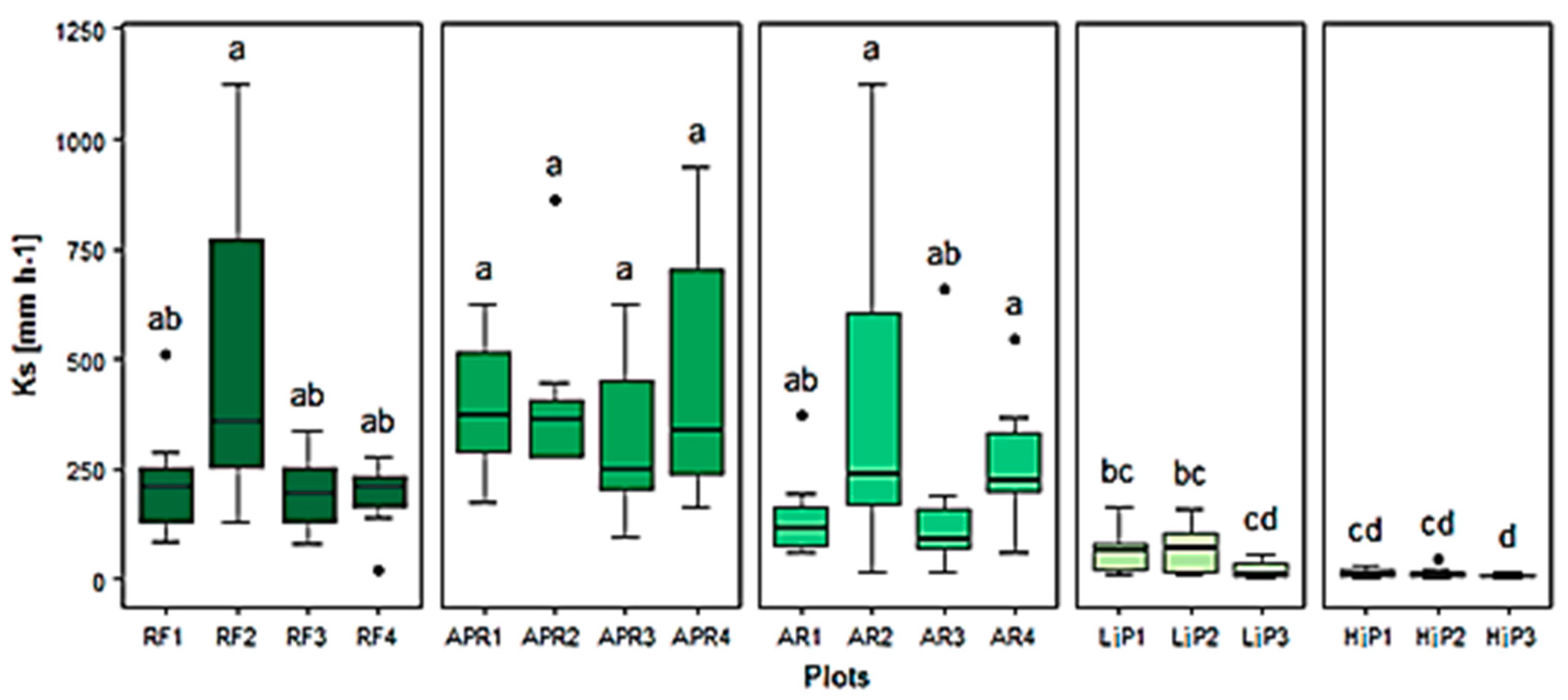
3.2. Soil Physical and Hydraulic Properties
4. Discussion
4.1. Effects of Land-Cover Type and Land Use History on Soil Physical and Hydraulic Properties
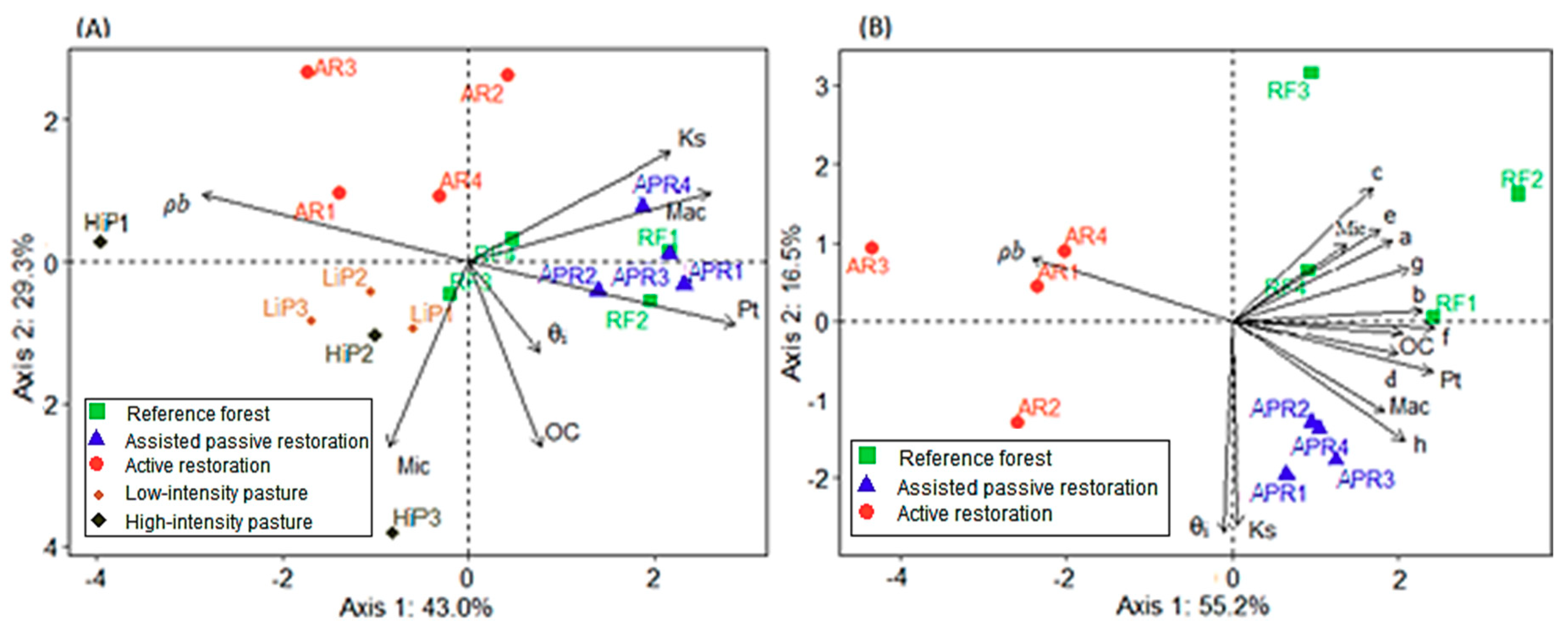
4.2. Relationships between Soil, Vegetation and Land Use History
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chazdon, R.L. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 2008, 320, 1458–1460. [Google Scholar] [CrossRef]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Gandolfi, S.; Nave, A.G.; Aronson, J.; Barreto, T.E.; Vidal, C.Y.; Brancalion, P.H.S. Large-scale ecological restoration of high-diversity tropical forests in SE Brazil. For. Ecol. Manag. 2011, 261, 1605–1613. [Google Scholar] [CrossRef]
- Shono, K.; Cadaweng, E.A.; Durst, P.B. Application of assisted natural regeneration to restore degraded tropical forestlands. Restor. Ecol. 2007, 15, 620–626. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Reid, J.L.; Holl, K.D. Hidden costs of passive restoration: Passive restoration. Restor. Ecol. 2014, 22, 284–287. [Google Scholar] [CrossRef]
- Holl, K.D.; Aide, T.M. When and where to actively restore ecosystems? For. Ecol. Manag. 2011, 261, 1558–1563. [Google Scholar] [CrossRef]
- Badalamenti, E.; da Silveira Bueno, R.; Campo, O.; Gallo, M.; La Mela Veca, D.; Pasta, S.; Sala, G.; La Mantia, T. Pine stand density influences the regeneration of Acacia saligna Labill. H.L. Wendl. and native woody species in a mediterranean coastal pine plantation. Forests 2018, 9, 359. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Gageler, R.; Bonner, M.; Kirchhof, G.; Amos, M.; Robinson, N.; Schmidt, S.; Shoo, L.P. Early response of soil properties and function to riparian rainforest restoration. PLoS ONE 2014, 9, e104198. [Google Scholar] [CrossRef]
- Mendes, M.S.; Latawiec, A.E.; Sansevero, J.B.B.; Crouzeilles, R.; de Moraes, L.F.D.; Castro, A.; Pinto, H.N.A.; Brancalion, P.H.S.; Rodrigues, R.R.; Chazdon, R.L.; et al. Look down—There is a gap—The need to include soil data in Atlantic Forest restoration: Scarcity of soil data in restoration. Restor. Ecol. 2018. [Google Scholar] [CrossRef]
- Zimmermann, B.; Elsenbeer, H.; De Moraes, J.M. The influence of land-use changes on soil hydraulic properties: Implications for runoff generation. For. Ecol. Manag. 2006, 222, 29–38. [Google Scholar] [CrossRef]
- Hassler, S.K.; Zimmermann, B.; van Breugel, M.; Hall, J.S.; Elsenbeer, H. Recovery of saturated hydraulic conductivity under secondary succession on former pasture in the humid tropics. For. Ecol. Manag. 2011, 261, 1634–1642. [Google Scholar] [CrossRef]
- Zimmermann, A.; Schinn, D.S.; Francke, T.; Elsenbeer, H.; Zimmermann, B. Uncovering patterns of near-surface saturated hydraulic conductivity in an overland flow-controlled landscape. Geoderma 2013, 195–196, 1–11. [Google Scholar] [CrossRef]
- Alagna, V.; Di Prima, S.; Rodrigo-Comino, J.; Iovino, M.; Pirastru, M.; Keesstra, S.; Novara, A.; Cerdà, A. The impact of the age of vines on soil hydraulic conductivity in vineyards in eastern Spain. Water 2017, 10, 14. [Google Scholar] [CrossRef]
- Di Prima, S.; Marrosu, R.; Lassabatere, L.; Angulo-Jaramillo, R.; Pirastru, M. In situ characterization of preferential flow by combining plot- and point-scale infiltration experiments on a hillslope. J. Hydrol. 2018, 563, 633–642. [Google Scholar] [CrossRef]
- Cullotta, S.; Bagarello, V.; Baiamonte, G.; Gugliuzza, G.; Iovino, M.; La Mela Veca, D.S.; Maetzke, F.; Palmeri, V.; Sferlazza, S. Comparing different methods to determine soil physical quality in a mediterranean forest and pasture land. Soil Sci. Soc. Am. J. 2016, 80, 1038–1056. [Google Scholar] [CrossRef]
- Di Prima, S.; Bagarello, V.; Angulo-Jaramillo, R.; Bautista, I.; Cerdà, A.; del Campo, A.; González-Sanchis, M.; Iovino, M.; Lassabatere, L.; Maetzke, F. Impacts of thinning of a Mediterranean oak forest on soil properties influencing water infiltration. J. Hydrol. Hydromech. 2017, 65, 276–286. [Google Scholar] [CrossRef]
- Elrick, D.E.; Reynolds, W.D. Methods for analyzing constant-head well permeameter data. Soil Sci. Soc. Am. J. 1992, 56, 320–323. [Google Scholar] [CrossRef]
- Deb, S.K.; Shukla, M.K. Variability of hydraulic conductivity due to multiple factors. Am. J. Environ. Sci. 2012, 8, 489–502. [Google Scholar] [CrossRef]
- Godsey, S.; Elsenbeer, H. The soil hydrologic response to forest regrowth: A case study from southwestern Amazonia. Hydrol. Process. 2002, 16, 1519–1522. [Google Scholar] [CrossRef]
- Ziegler, A.D.; Giambelluca, T.W.; Tran, L.T.; Vana, T.T.; Nullet, M.A.; Fox, J.; Vien, T.D.; Pinthong, J.; Maxwell, J.; Evett, S. Hydrological consequences of landscape fragmentation in mountainous northern Vietnam: Evidence of accelerated overland flow generation. J. Hydrol. 2004, 287, 124–146. [Google Scholar] [CrossRef]
- Paul, M.; Catterall, C.P.; Pollard, P.C.; Kanowski, J. Recovery of soil properties and functions in different rainforest restoration pathways. For. Ecol. Manag. 2010, 259, 2083–2092. [Google Scholar] [CrossRef]
- Nyberg, G.; Bargués Tobella, A.; Kinyangi, J.; Ilstedt, U. Soil property changes over a 120-yr chronosequence from forest to agriculture in western Kenya. Hydrol. Earth Syst. Sci. 2012, 16, 2085–2094. [Google Scholar] [CrossRef]
- Leite, P.A.M.; de Souza, E.S.; dos Santos, E.S.; Gomes, R.J.; Cantalice, J.R.; Wilcox, B.P. The influence of forest regrowth on soil hydraulic properties and erosion in a semiarid region of Brazil. Ecohydrology 2017, 11, 1–12. [Google Scholar] [CrossRef]
- Mapa, R.B. Effect of reforestation using Tectona grandis on infiltration and soil water retention. For. Ecol. Manag. 1995, 77, 119–125. [Google Scholar] [CrossRef]
- Bonell, M.; Purandara, B.K.; Venkatesh, B.; Krishnaswamy, J.; Acharya, H.A.K.; Singh, U.V.; Jayakumar, R.; Chappell, N. The impact of forest use and reforestation on soil hydraulic conductivity in the Western Ghats of India: Implications for surface and sub-surface hydrology. J. Hydrol. 2010, 391, 47–62. [Google Scholar] [CrossRef]
- Ghimire, C.P.; Bruijnzeel, L.A.; Bonell, M.; Coles, N.; Lubczynski, M.W.; Gilmour, D.A. The effects of sustained forest use on hillslope soil hydraulic conductivity in the Middle Mountains of Central Nepal: Sustained forest use and soil hydraulic conductivity. Ecohydrology 2014, 7, 478–495. [Google Scholar] [CrossRef]
- Lozano-Baez, S.; Cooper, M.; Ferraz, S.; Ribeiro Rodrigues, R.; Pirastru, M.; Di Prima, S. Previous land use affects the recovery of soil hydraulic properties after forest restoration. Water 2018, 10, 453. [Google Scholar] [CrossRef]
- Zwartendijk, B.W.; van Meerveld, H.J.; Ghimire, C.P.; Bruijnzeel, L.A.; Ravelona, M.; Jones, J.P.G. Rebuilding soil hydrological functioning after swidden agriculture in eastern Madagascar. Agric. Ecosyst. Environ. 2017, 239, 101–111. [Google Scholar] [CrossRef]
- Cooper, M.; Rosa, J.D.; Medeiros, J.C.; de Oliveira, T.C.; Toma, R.S.; Juhász, C.E.P. Hydro-physical characterization of soils under tropical semi-deciduous forest. Sci. Agricol. 2012, 69, 152–159. [Google Scholar] [CrossRef]
- Ziter, C.; Graves, R.A.; Turner, M.G. How do land-use legacies affect ecosystem services in United States cultural landscapes? Landsc. Ecol. 2017, 32, 2205–2218. [Google Scholar] [CrossRef]
- Foster, D.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. BioScience 2003, 53, 77–88. [Google Scholar] [CrossRef]
- Zimmermann, B.; Elsenbeer, H. Spatial and temporal variability of soil saturated hydraulic conductivity in gradients of disturbance. J. Hydrol. 2008, 361, 78–95. [Google Scholar] [CrossRef]
- Mello, M.H.; Pedro Junior, M.J.; Ortolani, A.A.; Alfonsi, R.R. Chuva e Temperatura: Cem Anos de Observações em Campinas; Boletim Tecnico; IAC: Campinas, Brazil, 1994. [Google Scholar]
- de Oliveira, L.H.d.S.; Valladares, G.S.; Coelho, R.M.; Criscuolo, C. Soil vulnerability to degradation at Campinas municipality, SP. Geografia (Londrina) 2014, 22, 65–79. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Rodrigues, R.R.; Lima, R.A.F.; Gandolfi, S.; Nave, A.G. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol. Conserv. 2009, 142, 1242–1251. [Google Scholar] [CrossRef]
- Molin, P.G.; Gergel, S.E.; Soares-Filho, B.S.; Ferraz, S.F.B. Spatial determinants of Atlantic Forest loss and recovery in Brazil. Landsc. Ecol. 2017, 32, 857–870. [Google Scholar] [CrossRef]
- Preiskorn, G.M.; Pimenta, D.; Amazonas, N.T.; Nave, A.G.; Gandolfi, S.; Rodrigues, R.R.; Belloto, A.; Cunha, M.C.S. Metodologia de restauração para fins de aproveitamento econômico (reservas legais e áreas agrícolas). In Pacto Pela Restauração da Mata Atlântica—Referencial dos Conceitos e ações de Restauração Florestal; Rodrigues, R.R., Brancalion, P.H.S., Eds.; LERF/ESALQ: Instituto BioAtlântica: São Paulo, Brazil, 2009; pp. 158–175. ISBN 978-85-60840-02-1. [Google Scholar]
- Suganuma, M.S.; Durigan, G. Indicators of restoration success in riparian tropical forests using multiple reference ecosystems: Indicators of riparian forests restoration success. Restor. Ecol. 2015, 23, 238–251. [Google Scholar] [CrossRef]
- Toledo, R.M.; Santos, R.F.; Baeten, L.; Perring, M.P.; Verheyen, K. Soil properties and neighbouring forest cover affect above-ground biomass and functional composition during tropical forest restoration. Appl. Veg. Sci. 2018, 21, 179–189. [Google Scholar] [CrossRef]
- Viani, R.A.G.; Barreto, T.E.; Farah, F.T.; Rodrigues, R.R.; Brancalion, P.H.S. Monitoring young tropical forest restoration sites: How much to measure? Trop. Conserv. Sci. 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Chaves, R.B.; Durigan, G.; Brancalion, P.H.S.; Aronson, J. On the need of legal frameworks for assessing restoration projects success: New perspectives from São Paulo state (Brazil): Legal instruments for assessing restoration. Restor. Ecol. 2015, 23, 754–759. [Google Scholar] [CrossRef]
- Canfield, R. Application of line interception method in sampling range vegetation. J. For. 1941, 39, 388–394. [Google Scholar] [CrossRef]
- Gee, G.; Or, D. Particle-size analysis. In Methods of Soil Analysis: Physical Methods; Dane, J.H., Topp, C., Eds.; Soil Science Society of America: Madison, WI, USA, 2002; pp. 255–293. ISBN 978-0-89118-841-4. [Google Scholar]
- Flint, A.L.; Flint, L.E. Particle density. In Methods of Soil Analysis: Physical Methods; Dane, J., Topp, G.C., Eds.; Soil Science Society of America: Madison, WI, USA, 2002; pp. 229–240. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Lassabatère, L.; Angulo-Jaramillo, R.; Soria Ugalde, J.M.; Cuenca, R.; Braud, I.; Haverkamp, R. Beerkan estimation of soil transfer parameters through infiltration experiments—BEST. Soil Sci. Soc. Am. J. 2006, 70, 521. [Google Scholar] [CrossRef]
- Braud, I.; De Condappa, D.; Soria, J.M.; Haverkamp, R.; Angulo-Jaramillo, R.; Galle, S.; Vauclin, M. Use of scaled forms of the infiltration equation for the estimation of unsaturated soil hydraulic properties (the Beerkan method). Eur. J. Soil Sci. 2005, 56, 361–374. [Google Scholar] [CrossRef]
- Bagarello, V.; Di Prima, S.; Iovino, M. Estimating saturated soil hydraulic conductivity by the near steady-state phase of a Beerkan infiltration test. Geoderma 2017, 303, 70–77. [Google Scholar] [CrossRef]
- Alagna, V.; Iovino, M.; Bagarello, V.; Mataix-Solera, J.; Lichner, L. Alternative analysis of transient infiltration experiment to estimate soil water repellency. Hydrol. Process. 2018. [Google Scholar] [CrossRef]
- Di Prima, S.; Concialdi, P.; Lassabatere, L.; Angulo-Jaramillo, R.; Pirastru, M.; Cerdà, A.; Keesstra, S. Laboratory testing of Beerkan infiltration experiments for assessing the role of soil sealing on water infiltration. Catena 2018, 167, 373–384. [Google Scholar] [CrossRef]
- Danielson, R.E.; Sutherland, P.L. Porosity. In Methods of Soil Analysis. Part I. Physical and Mineralogical Methods. Agronomy Monograph No. 9; Klute, A., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1986; pp. 443–461. [Google Scholar]
- EMBRAPA. Manual of Methods of Soil Analysis, 2nd ed.; Embrapa Soils: Rio de Janeiro, Brazil, 2011. [Google Scholar]
- Reynolds, W.D.; Drury, C.F.; Yang, X.M.; Tan, C.S. Optimal soil physical quality inferred through structural regression and parameter interactions. Geoderma 2008, 146, 466–474. [Google Scholar] [CrossRef]
- Lee, D.M.; Elrick, D.E.; Reynolds, W.D.; Clothier, B.E. A comparison of three field methods for measuring saturated hydraulic conductivity. Can. J. Soil Sci. 1985, 65, 563–573. [Google Scholar] [CrossRef]
- Scheffler, R.; Neill, C.; Krusche, A.V.; Elsenbeer, H. Soil hydraulic response to land-use change associated with the recent soybean expansion at the Amazon agricultural frontier. Agric. Ecosyst. Environ. 2011, 144, 281–289. [Google Scholar] [CrossRef]
- Salemi, L.F.; Groppo, J.D.; Trevisan, R.; de Moraes, J.M.; de Barros Ferraz, S.F.; Villani, J.P.; Duarte-Neto, P.J.; Martinelli, L.A. Land-use change in the Atlantic rainforest region: Consequences for the hydrology of small catchments. J. Hydrol. 2013, 499, 100–109. [Google Scholar] [CrossRef]
- Cooper, M.; Medeiros, J.C.; Rosa, J.D.; Soria, J.E.; Toma, R.S. Soil functioning in a toposequence under rainforest in São Paulo, Brazil. Rev. Bras. Ciência Solo 2013, 37, 392–399. [Google Scholar] [CrossRef]
- Ferraz, S.F.B.; Ferraz, K.M.P.M.B.; Cassiano, C.C.; Brancalion, P.H.S.; da Luz, D.T.A.; Azevedo, T.N.; Tambosi, L.R.; Metzger, J.P. How good are tropical forest patches for ecosystem services provisioning? Landsc. Ecol. 2014, 29, 187–200. [Google Scholar] [CrossRef]
- Ilstedt, U.; Malmer, A.; Verbeeten, E.; Murdiyarso, D. The effect of afforestation on water infiltration in the tropics: A systematic review and meta-analysis. For. Ecol. Manag. 2007, 251, 45–51. [Google Scholar] [CrossRef]
- Filoso, S.; Bezerra, M.O.; Weiss, K.C.; Palmer, M.A. Impacts of forest restoration on water yield: A systematic review. PLoS ONE 2017, 12, 1–26. [Google Scholar] [CrossRef]
- Wei, W.; Chen, D.; Wang, L.; Daryanto, S.; Chen, L.; Yu, Y.; Lu, Y.; Sun, G.; Feng, T. Global synthesis of the classifications, distributions, benefits and issues of terracing. Earth-Sci. Rev. 2016, 159, 388–403. [Google Scholar] [CrossRef]
- Atta, H.A.E.; Aref, I. Effect of terracing on rainwater harvesting and growth of Juniperus procera Hochst. ex Endlicher. Int. J. Environ. Sci. Technol. 2010, 7, 59–66. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- Martínez, L.; Zinck, J. Temporal variation of soil compaction and deterioration of soil quality in pasture areas of Colombian Amazonia. Soil Tillage Res. 2004, 75, 3–18. [Google Scholar] [CrossRef]
- Nogueira, L.R.; da Silva, C.F.; Pereira, M.G.; Gaia-Gomes, J.H.; da Silva, E.M.R. Biological Properties and Organic Matter Dynamics of Soil in Pasture and Natural Regeneration Areas in the Atlantic Forest Biome. Rev. Bras. Ciência Solo 2016, 40. [Google Scholar] [CrossRef]
- César, R.G.; Moreno, V.S.; Coletta, G.D.; Chazdon, R.L.; Ferraz, S.F.B.; de Almeida, D.R.A.; Brancalion, P.H.S. Early ecological outcomes of natural regeneration and tree plantations for restoring agricultural landscapes. Ecol. Appl. 2018, 28, 373–384. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Finegan, B.; Capers, R.S.; Salgado-Negret, B.; Casanoves, F.; Boukili, V.; Norden, N. Composition and dynamics of functional groups of trees during tropical forest succession in Northeastern Costa Rica: Functional groups of trees. Biotropica 2010, 42, 31–40. [Google Scholar] [CrossRef]
- Garcia, L.C.; Hobbs, R.J.; Ribeiro, D.B.; Tamashiro, J.Y.; Santos, F.A.M.; Rodrigues, R.R. Restoration over time: Is it possible to restore trees and non-trees in high-diversity forests? Appl. Veg. Sci. 2016, 19, 655–666. [Google Scholar] [CrossRef]
- de Souza, F.M.; Batista, J.L.F. Restoration of seasonal semideciduous forests in Brazil: Influence of age and restoration design on forest structure. For. Ecol. Manag. 2004, 191, 185–200. [Google Scholar] [CrossRef]
- Chapla, T.E.; Campos, J.B. Allelopathic evidence in exotic guava (Psidium guajava L.). Braz. Arch. Biol. Technol. 2010, 53, 1359–1362. [Google Scholar] [CrossRef]
- Niemeyer, R.J.; Fremier, A.K.; Heinse, R.; Chávez, W.; DeClerck, F.A.J. Woody vegetation increases saturated hydraulic conductivity in dry tropical Nicaragua. Vadose Zone J. 2014, 13, 1–11. [Google Scholar] [CrossRef]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Mommer, L.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Holl, K.D.; Zahawi, R.A. Factors explaining variability in woody above-ground biomass accumulation in restored tropical forest. For. Ecol. Manag. 2014, 319, 36–43. [Google Scholar] [CrossRef]
- Rocha, G.P.E.; Vieira, D.L.M.; Simon, M.F. Fast natural regeneration in abandoned pastures in southern Amazonia. For. Ecol. Manag. 2016, 370, 93–101. [Google Scholar] [CrossRef]
- Bürgi, M.; Östlund, L.; Mladenoff, D.J. Legacy Effects of Human Land Use: Ecosystems as Time-Lagged Systems. Ecosystems 2017, 20, 94–103. [Google Scholar] [CrossRef]
- Owuor, S.O.; Butterbach-Bahl, K.; Guzha, A.C.; Jacobs, S.; Merbold, L.; Rufino, M.C.; Pelster, D.E.; Díaz-Pinés, E.; Breuer, L. Conversion of natural forest results in a significant degradation of soil hydraulic properties in the highlands of Kenya. Soil Tillage Res. 2018, 176, 36–44. [Google Scholar] [CrossRef]
- Bargués Tobella, A.; Reese, H.; Almaw, A.; Bayala, J.; Malmer, A.; Laudon, H.; Ilstedt, U. The effect of trees on preferential flow and soil infiltrability in an agroforestry parkland in semiarid Burkina Faso. Water Resour. Res. 2014, 50, 3342–3354. [Google Scholar] [CrossRef] [PubMed]
- Ilstedt, U.; Bargués Tobella, A.; Bazié, H.R.; Bayala, J.; Verbeeten, E.; Nyberg, G.; Sanou, J.; Benegas, L.; Murdiyarso, D.; Laudon, H.; et al. Intermediate tree cover can maximize groundwater recharge in the seasonally dry tropics. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RF | APR | AR | |
|---|---|---|---|
| Basal area (m2/ha−1) | 26.4 ± 4.49 a | 20.8 ± 2.53 a | 12.5 ± 3.32 a |
| Canopy cover (%) | 95.8 ± 2.17 a | 91.3 ± 1.49 a | 77.5 ± 3.11 b |
| Vegetation height of trees (m) | 10.1 ± 1.16 a | 7.79 ± 0.57 a | 7.00 ± 0.11 a |
| Density of trees (ind·ha−1) | 1,325 ± 137 a | 1,300 ± 72 a | 610 ± 72 b |
| Density of saplings (ind·ha−1) | 3,950 ± 172 a | 1,963 ± 959 ab | 850 ± 119 b |
| Total richness (tree and non-tree) | 82 ± 4 a | 62 ± 1 a | 38 ± 2 b |
| Overstory richness | 50 ± 2 a | 41 ± 1 a | 30 ± 1 b |
| Richness of saplings | 62 ± 2 a | 39 ± 3 b | 15 ± 1 b |
| Land Cover | Clay (%) | Silt (%) | Sand (%) | Sand (%) | Textural Class | ||||
|---|---|---|---|---|---|---|---|---|---|
| Very Fine | Fine | Medium | Coarse | Very Coarse | |||||
| RF | 24.8 b | 25.9 b | 49.3 a | 6.23 a | 14.0 abc | 12.8 a | 9.21 a | 6.99 a | Sandy clay loam |
| APR | 30.2 a | 31.9 a | 37.9 c | 6.28 a | 12.1 c | 9.11 c | 5.54 c | 4.84 b | Clay loam |
| AR | 30.0 ab | 23.9 b | 46.1 ab | 5.73 a | 14.2 ab | 11.7 ab | 7.67 b | 6.68 a | Sandy clay loam |
| LiP | 31.7 a | 22.6 b | 45.7 ab | 6.60 a | 15.1 a | 12.6 ab | 6.87 bc | 4.44 b | Clay loam |
| HiP | 33.6 a | 23.2 b | 43.1 bc | 5.82 a | 12.8 bc | 10.7 bc | 7.43 b | 6.34 ab | Clay loam |
| Land Cover | ρb | Pd | OC | Ks | Mic | Mac | Pt | θi | θs |
|---|---|---|---|---|---|---|---|---|---|
| RF | 1.04 b | 2.66 ab | 16.2 a | 215 ab | 0.29 ab | 0.32 a | 0.61 a | 0.18 c | 0.48 ab |
| (7.06) | (1.17) | (24.3) | (90.2) | (14.6) | (9.82) | (4.54) | (12.6) | (22.2) | |
| APR | 1.04 b | 2.68 a | 16.4 a | 351 a | 0.31 bc | 0.29 a | 0.60 a | 0.24 a | 0.45 b |
| (6.50) | (1.11) | (21.4) | (58.4) | (9.12) | (9.18) | (2.53) | (14.1) | (19.4) | |
| AR | 1.19 a | 2.68 a | 10.3 b | 163 b | 0.29 c | 0.25 b | 0.56 b | 0.20 bc | 0.38 c |
| (7.20) | (0.49) | (35.5) | (135.5) | (12.8) | (10.1) | (4.49) | (13.7) | (14.7) | |
| LiP | 1.14 a | 2.65 ab | 15.1 ab | 32.6 c | 0.33 ab | 0.22 c | 0.57 b | 0.22 ab | 0.54 a |
| (7.12) | (0.82) | (12.4) | (155.0) | (10.8) | (11.7) | (5.31) | (10.8) | (15.0) | |
| HiP | 1.18 a | 2.64 b | 18.6 a | 10.4 d | 0.34 a | 0.20 c | 0.55 b | 0.22 ab | 0.50 ab |
| (12.0) | (0.67) | (28.4) | (82.9) | (11.9) | (9.90) | (9.59) | (27.0) | (15.5) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano-Baez, S.E.; Cooper, M.; Frosini de Barros Ferraz, S.; Ribeiro Rodrigues, R.; Castellini, M.; Di Prima, S. Recovery of Soil Hydraulic Properties for Assisted Passive and Active Restoration: Assessing Historical Land Use and Forest Structure. Water 2019, 11, 86. https://doi.org/10.3390/w11010086
Lozano-Baez SE, Cooper M, Frosini de Barros Ferraz S, Ribeiro Rodrigues R, Castellini M, Di Prima S. Recovery of Soil Hydraulic Properties for Assisted Passive and Active Restoration: Assessing Historical Land Use and Forest Structure. Water. 2019; 11(1):86. https://doi.org/10.3390/w11010086
Chicago/Turabian StyleLozano-Baez, Sergio Esteban, Miguel Cooper, Silvio Frosini de Barros Ferraz, Ricardo Ribeiro Rodrigues, Mirko Castellini, and Simone Di Prima. 2019. "Recovery of Soil Hydraulic Properties for Assisted Passive and Active Restoration: Assessing Historical Land Use and Forest Structure" Water 11, no. 1: 86. https://doi.org/10.3390/w11010086
APA StyleLozano-Baez, S. E., Cooper, M., Frosini de Barros Ferraz, S., Ribeiro Rodrigues, R., Castellini, M., & Di Prima, S. (2019). Recovery of Soil Hydraulic Properties for Assisted Passive and Active Restoration: Assessing Historical Land Use and Forest Structure. Water, 11(1), 86. https://doi.org/10.3390/w11010086