


Dendroclimatic Reconstruction of Seasonal Precipitation from Two Endangered Spruce Species in Northeastern Mexico


Abstract
1. Introduction
2. Materials and Methods
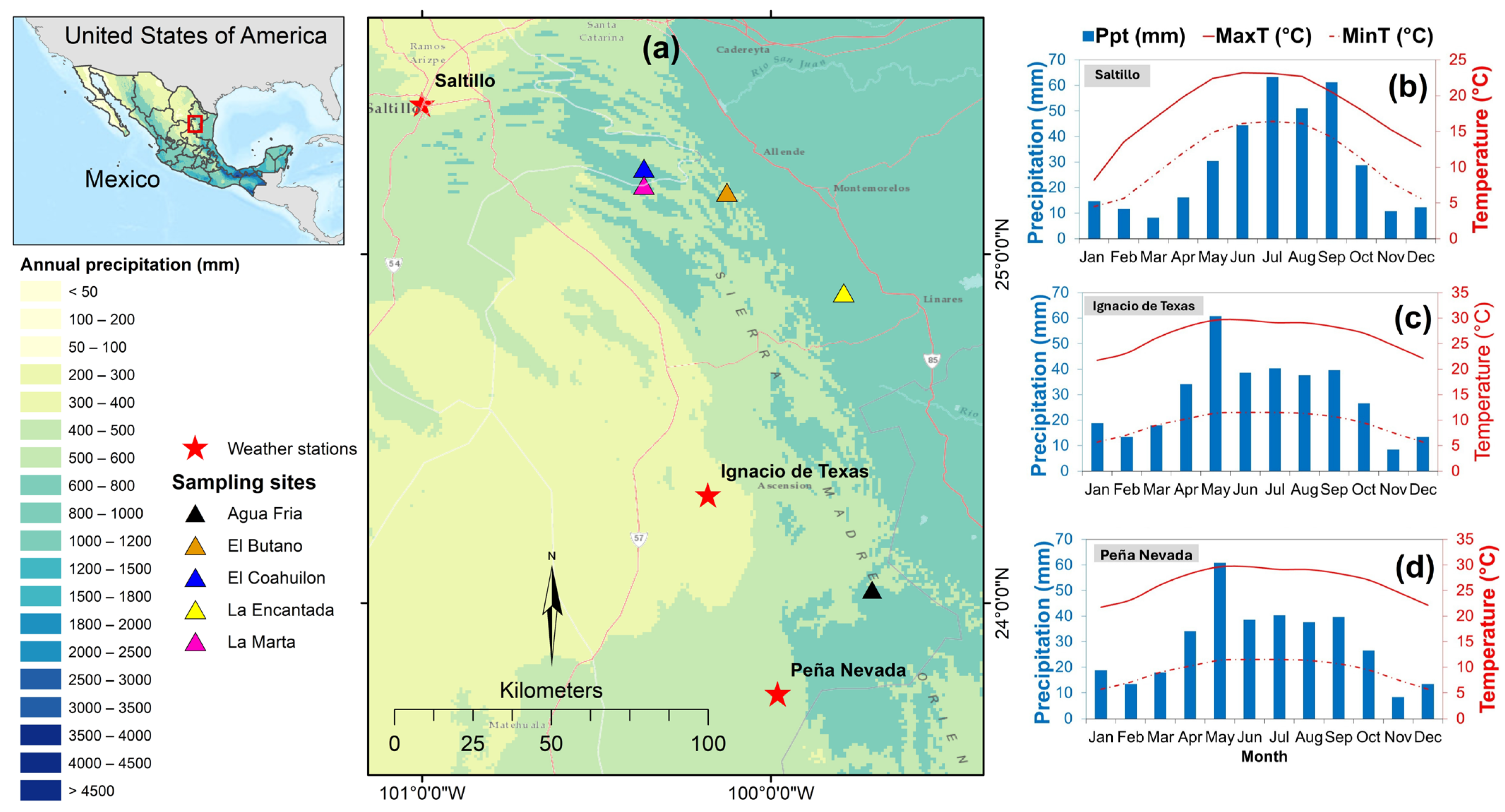
2.1. Study Area
2.2. Collection and Processing of Dendrochronological Samples
2.3. Climate Data Source
2.4. Correlation Analysis
2.5. Reconstruction of Climatic Variables
2.6. Comparison with Other Reconstructions
2.7. Analysis of the Influence of ENSO on Precipitation
3. Results
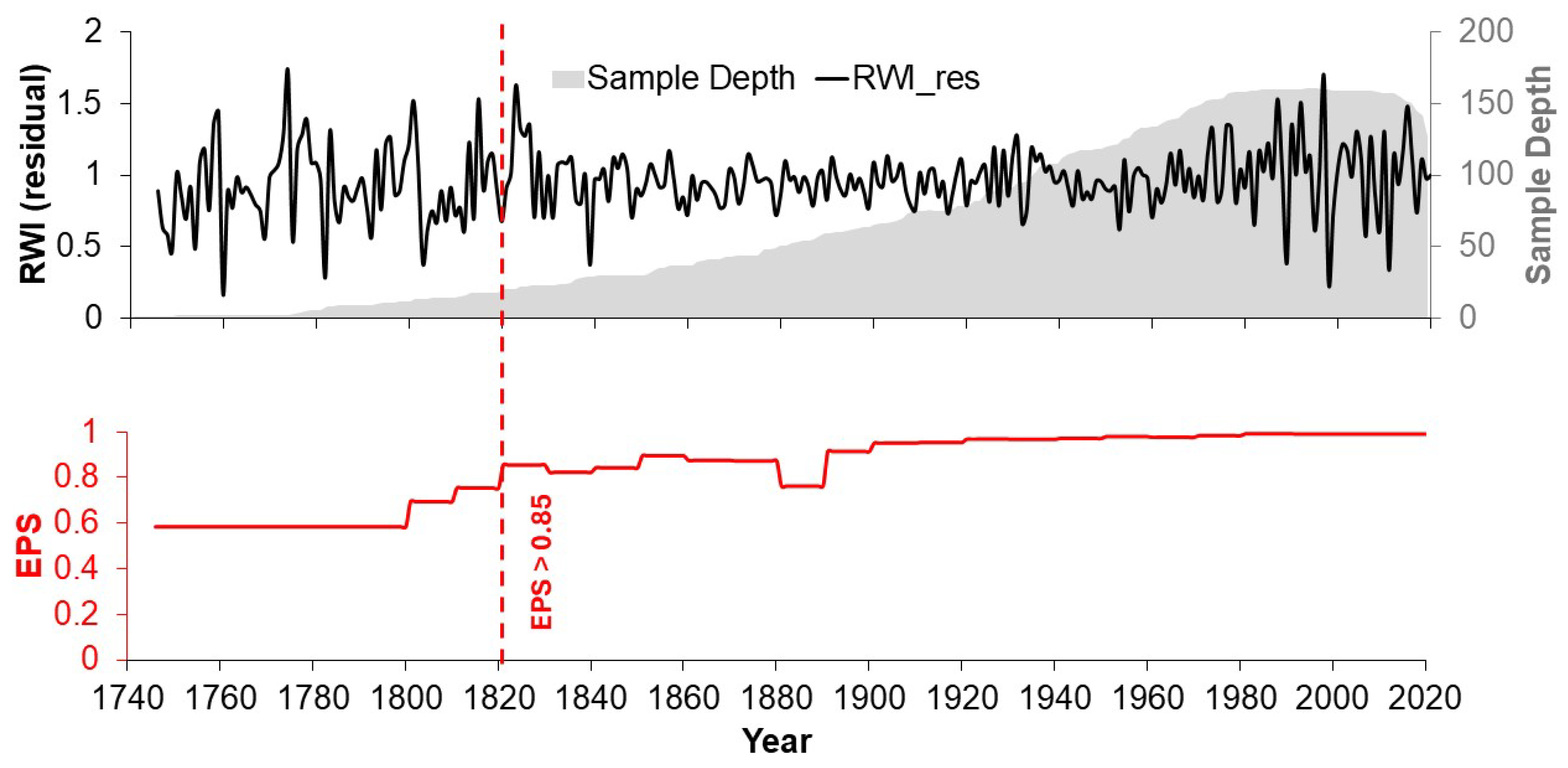
3.1. Age of Sampled Trees and Dendrochronological Parameters of the Ring Width Series
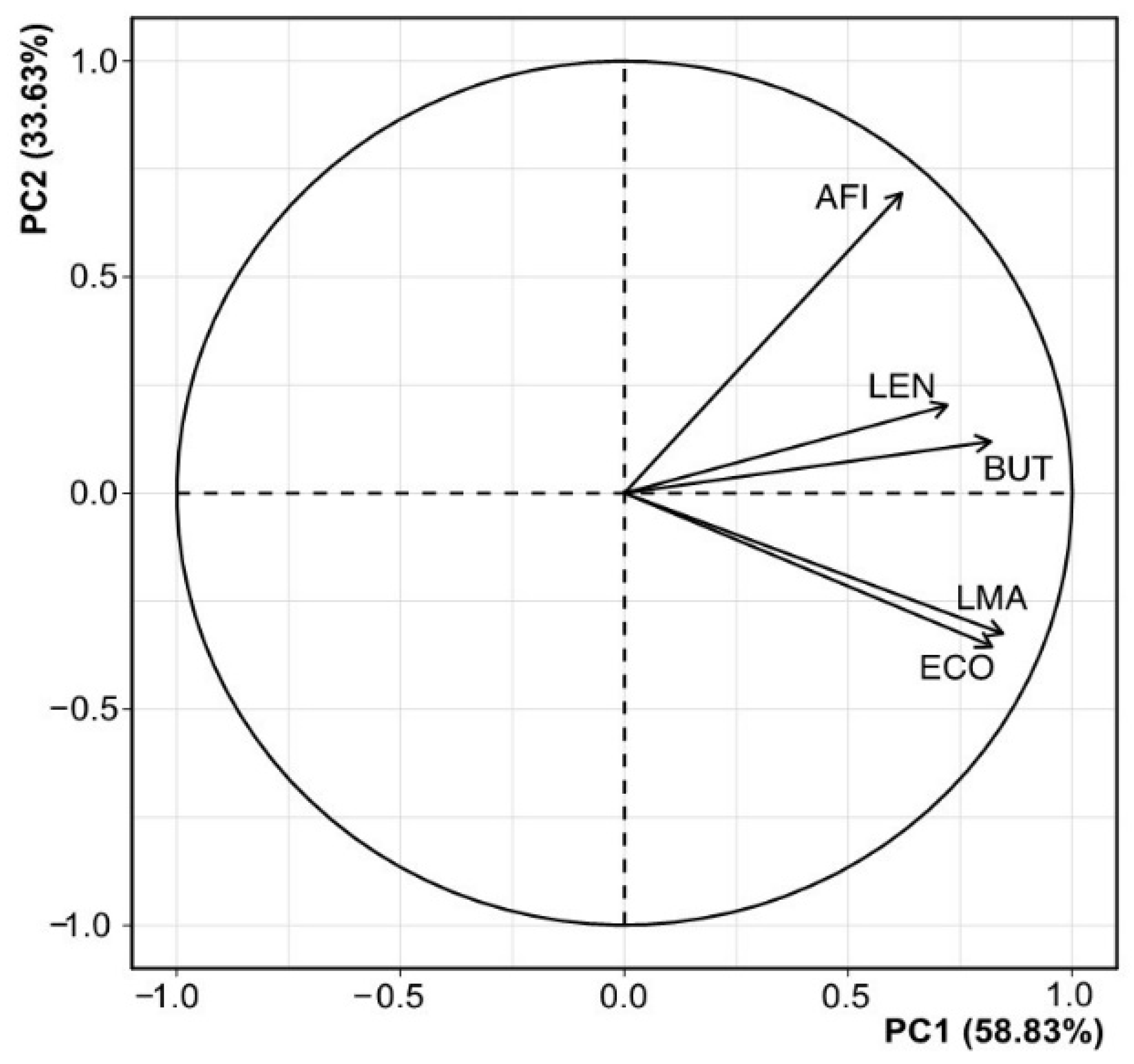
3.2. Correlation Analysis Between Spruce Series
3.3. Climatic Response of the Chronologies
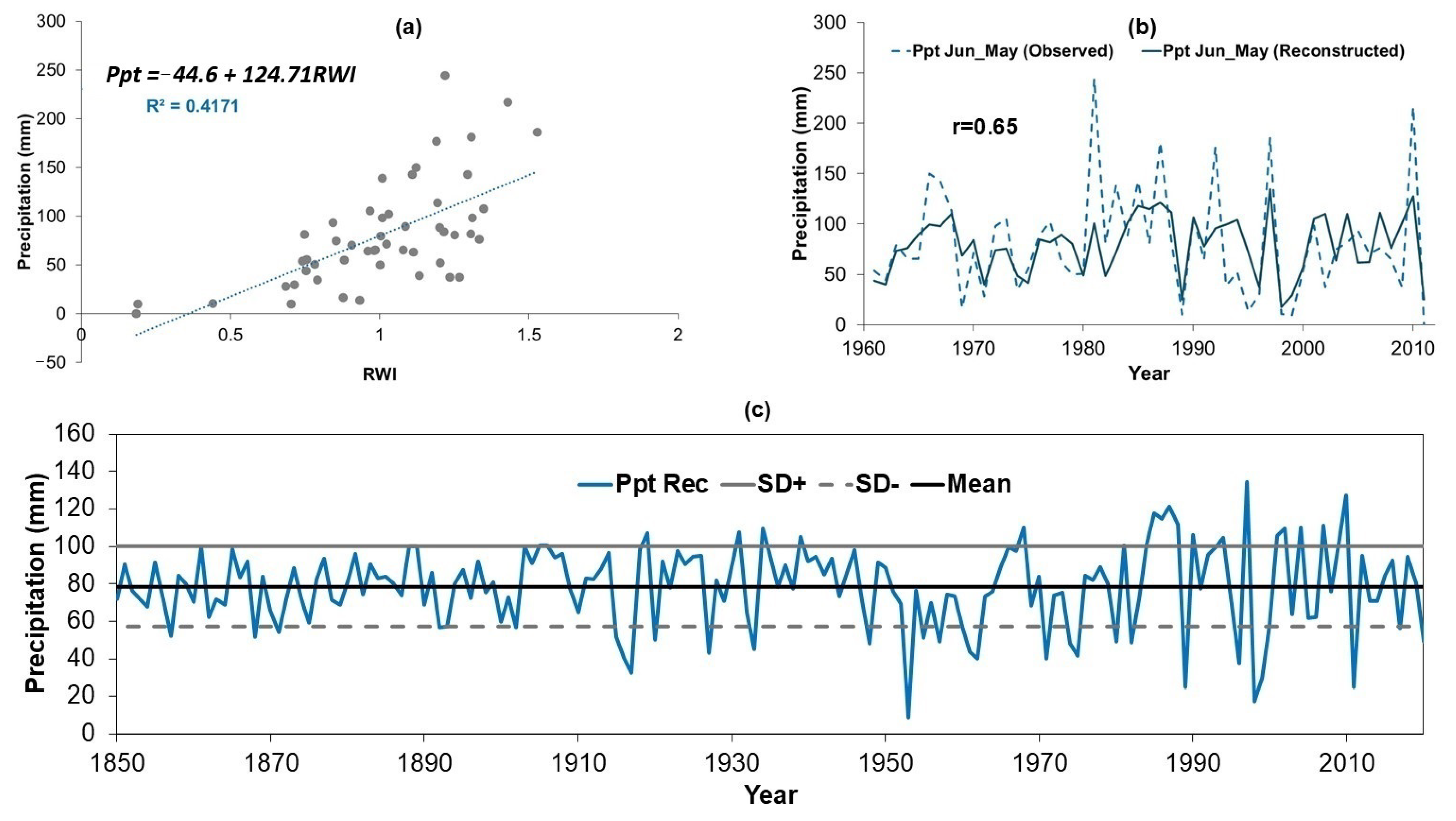
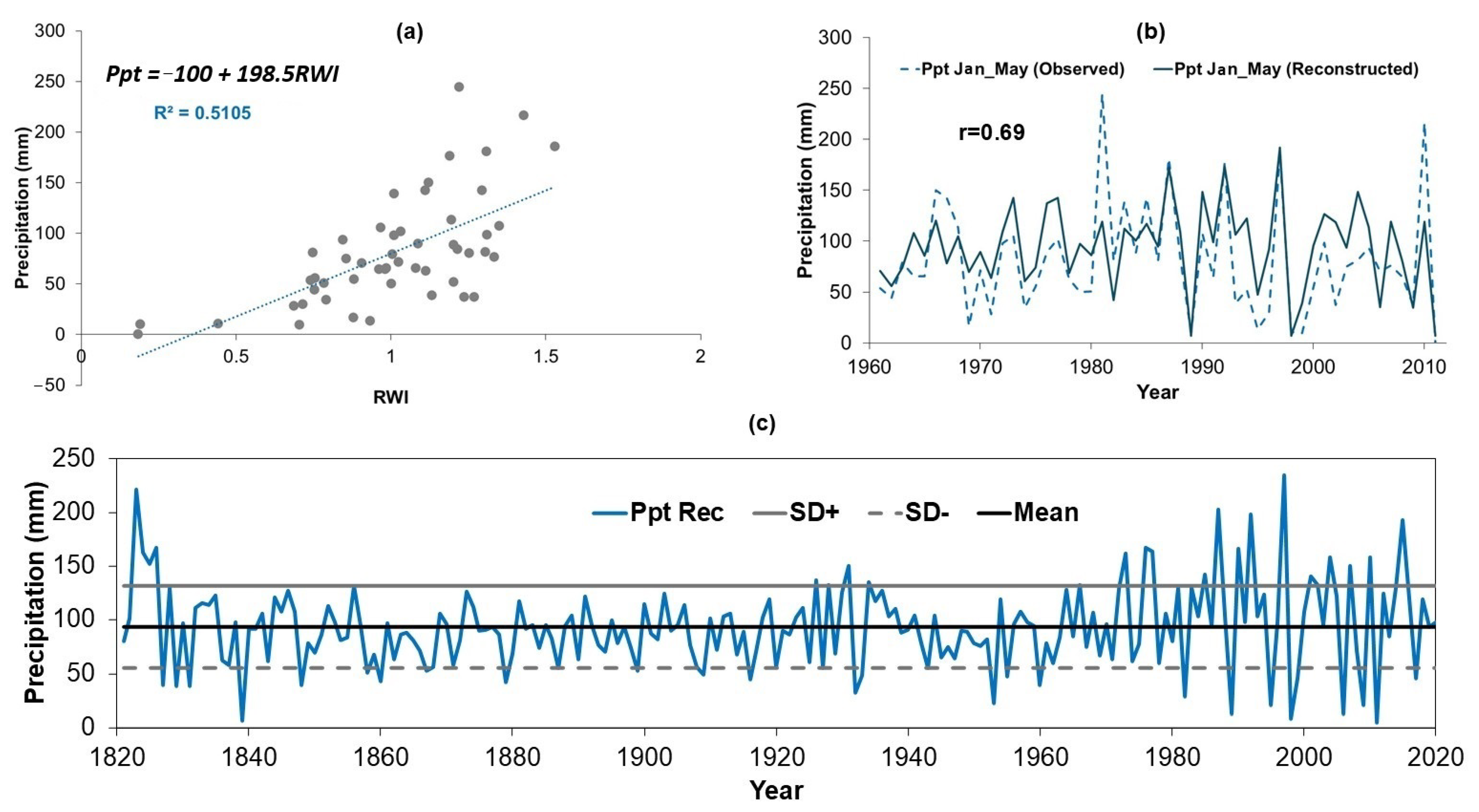
3.4. Development of the Model Used to Reconstruct Precipitation
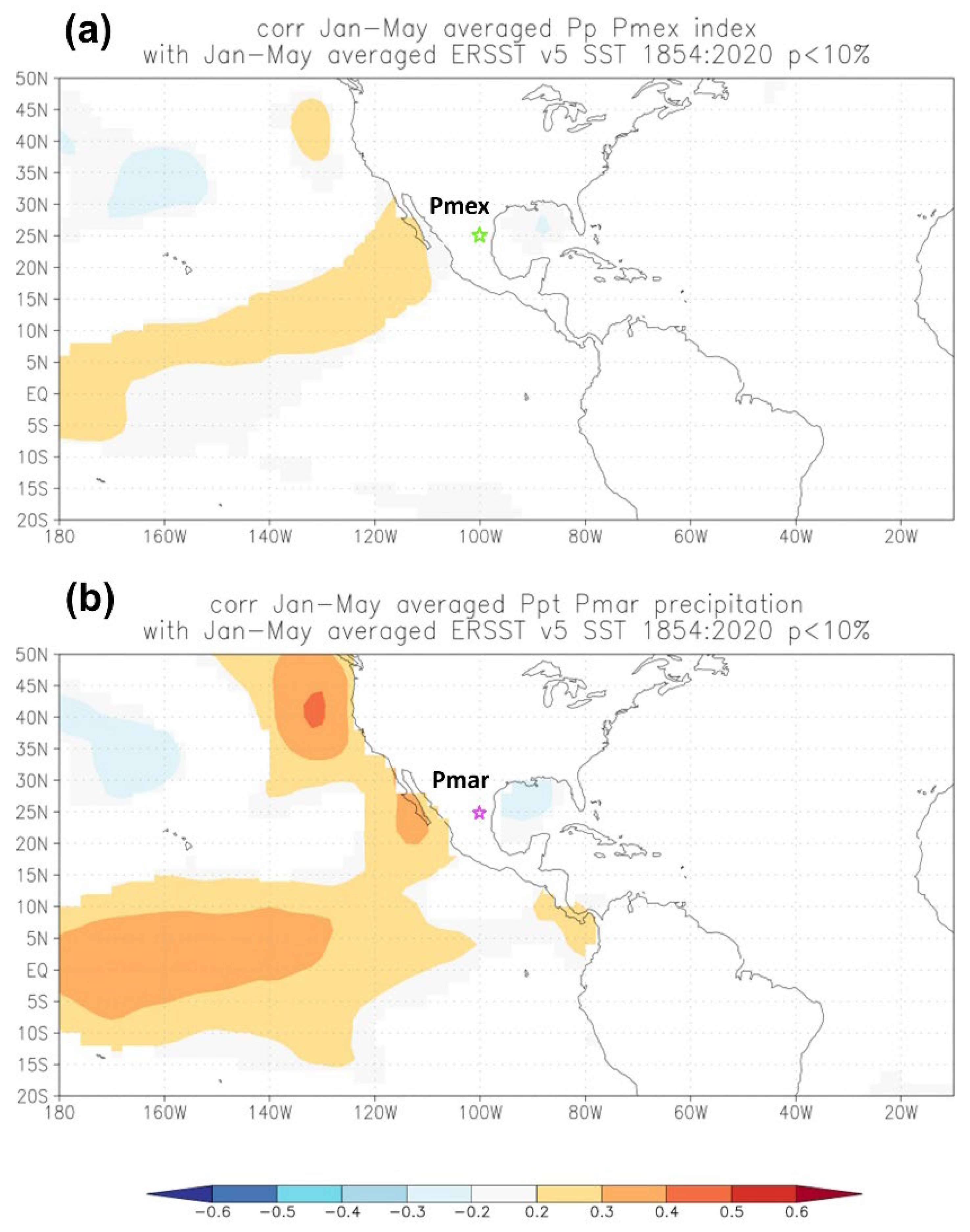
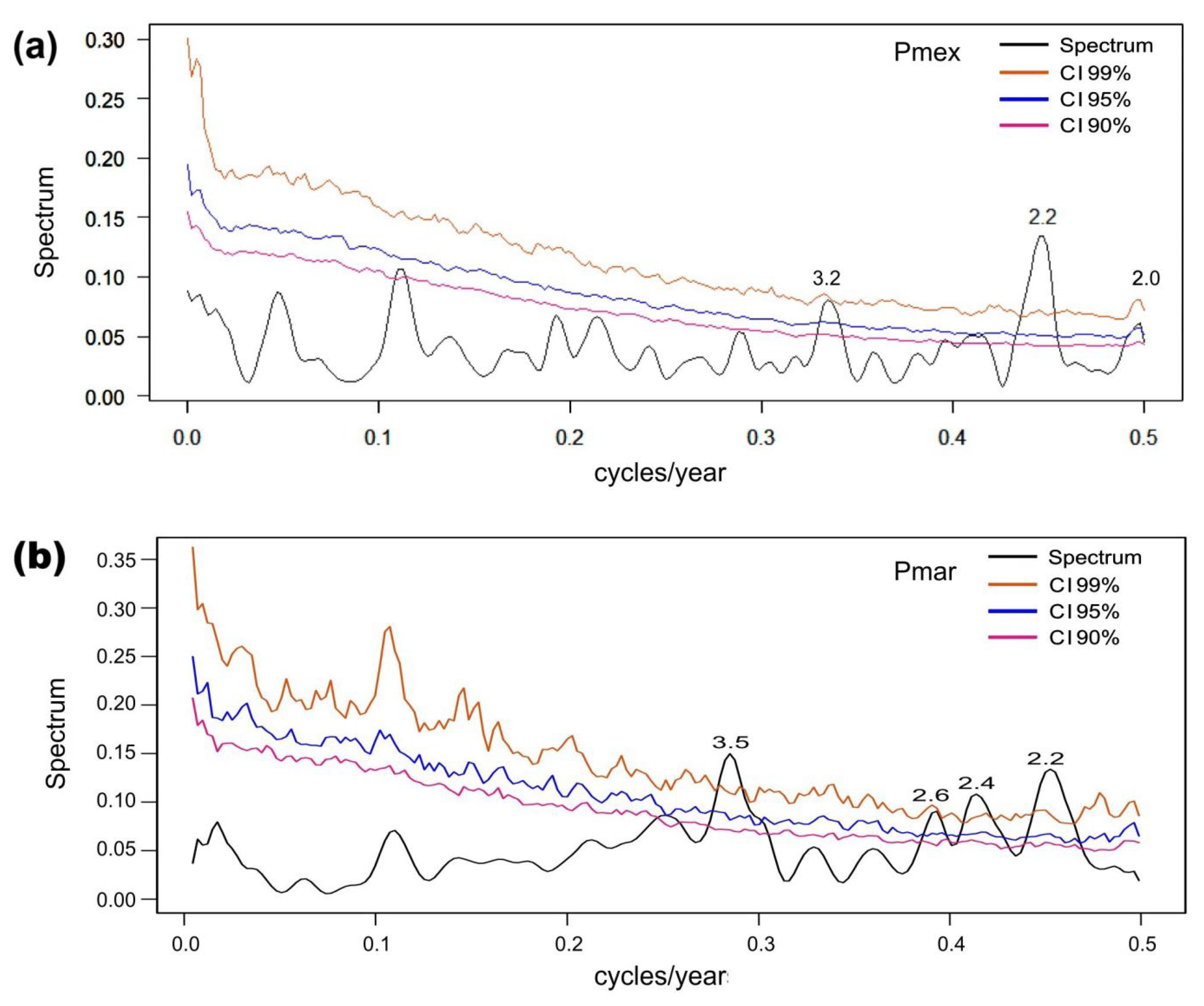
3.5. ENSO Modulations on Reconstructed Precipitation
4. Discussion
4.1. Climatic Response Function for the Dendrochronological Series
4.2. Reconstructed Precipitations
4.3. Influence of the SST’s and ENSO Phenomenon on the Precipitation Reconstructions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cook, B.I.; Mankin, J.S.; Anchukaitis, K.J. Climate change and drought: From past to future. Curr. Clim. Chang. Rep. 2018, 4, 164–170. [Google Scholar] [CrossRef]
- Wilhite, D.A.; Svoboda, M.D.; Hayes, M.J. Understanding the complex impacts of drought: A key to enhancing drought mitigation and preparedness. Water Resour. Manag. 2007, 21, 763–774. [Google Scholar] [CrossRef]
- Robb, K. The Rio Grande/Rio Bravo Water Delivery Under the 1944 Treaty: A Compendium of Ideas. Executive Summary Prepared at the Request of the United States International Boundary and Water Commission. 2022. Available online: https://www.ibwc.gov/wp-content/uploads/2023/04/Exec_Rio_Grande_White_Paper_-Summary-FINAL.pdf (accessed on 15 May 2025).
- Instituto Mexicano para la Competitividad (IMCO). México Enfrenta Sequías Sin Recursos Suficientes: Presupuesto 2024 para la Comisión Nacional del Agua. 2023. Available online: https://imco.org.mx/mexico-enfrenta-las-sequias-sin-recursos/ (accessed on 9 May 2025).
- Comision Nacional del Agua [CONAGUA]. Información Estadística Climatológica. 2023. Available online: https://smn.conagua.gob.mx/es/climatologia/informacion-climatologica/informacion-estadistica-climatologica (accessed on 8 May 2025).
- Pacheco-Treviño, S.; Manzano-Camarillo, M. The Socioeconomic dimensions of water scarcity in urban and rural Mexico: A comprehensive assessment of sustainable development. Sustainability 2024, 16, 1011. [Google Scholar] [CrossRef]
- Wright, K.T.; Johnson, K.R.; Bhattacharya, T.; Marks, G.S.; McGee, D.; Elsbury, D.; Peings, Y.; Lacaille-Muzquiz, J.-L.; Lum, G.; Beramendi-Orozco, L.; et al. Precipitation in Northeast Mexico primarily controlled by the relative warming of Atlantic SSTs. Geophys. Res. Lett. 2022, 49, e2022GL098186. [Google Scholar] [CrossRef]
- Meko, D.M.; Touchan, R.; Villanueva-Diaz, J.; Griffin, D.; Woodhouse, C.A.; Castro, C.L.; Carrillo, C.; Leavitt, S.O. Sierra San Pedro Martir, Baja California cool-season precipitation reconstructed from earlywood width of Abies concolor tree rings. JGR Biogeosci. 2013, 118, 1660–1673. [Google Scholar] [CrossRef]
- Mendoza-Maya, E.; Espino-Espino, J.; Quiñonez-Pérez, C.Z.; Flores-López, C.; Wehenkel, C.; Vargas-Hernández, J.; Sáenz-Romero, C. Proposal for conservation of three endangered species of Mexican spruce. Rev. Fitotec. Mex. 2015, 38, 235–247. [Google Scholar]
- Mendoza-Maya, E.; Gómez-Pineda, E.; Sáenz-Romero, C.; Hernández-Díaz, J.C.; López-Sánchez, C.A.; Vargas-Hernández, J.J.; Prieto-Ruiz, J.A.; Wehenkel, C. Assisted migration and the rare endemic plant species: The case of two endangered Mexican spruces. PeerJ 2022, 10, e13812. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Romero, C.; Larter, M.; González-Muñoz, N.; Wehenkel, C.; García-Blanco, A.; Castellanos-Acuña, D.; Burlett, R.; Delzon, S. Mexican conifers differ in their capacity to face climate. J. Plant Hydraul. 2024, 4, e203. [Google Scholar] [CrossRef]
- Ledig, T.; Hodgskiss, P.D.; Jacob-Cervantes, V. Genetic diversity, mating system, and conservation of a Mexican subalpine relicty, Picea mexicana Martínez. Conserv. Genet. 2002, 3, 113–122. [Google Scholar] [CrossRef]
- Cabral-Alemán, C.; Pompa-García, M.; Acosta-Hernández, A.C.; Zúñiga-Vásquez, J.M.; Camarero, J.J. Earlywood and latewood widths of Picea chihuahuana show contrasting sensitivity to seasonal climate. Forests 2017, 8, 173. [Google Scholar] [CrossRef]
- Díaz-Carrillo, Ó.A.; Wehenkel, C.A.; Villanueva-Díaz, J.; Hernández-Díaz, J.C. Serie dendrocronológica de Picea mexicana Martínez en La Sierra Madre Oriental. Rev. Mex. Agroecosistemas 2022, 9, 1–7. [Google Scholar]
- Segura-Sanchez, C.A.; Hernández-Velasco, J.; Villanueva-Díaz, J.; Chano, V.; Hernández-Díaz, J.C.; Mendoza-Maya, E.; Carrillo-Parra, A.; Wehenkel, C. Dendrogenomic resilience mechanisms of two endangered Mexican spruces. Plant Diverity 2025, in press. [Google Scholar] [CrossRef]
- Arreola-Ortíz, M.R.; González-Elizondo, M.; Návar-Cháidez, J. Dendrocronología de Pseudotsuga menziesii (Mirb.) Franco de la Sierra Madre Oriental en Nuevo León, México. Madera Y Bosques 2009, 16, 71–85. [Google Scholar] [CrossRef]
- Ledig, F.T.; Mapúla-Larreta, M.; Bermejo-Velázquez, B.V.; Reyes-Hernández, V.; Flores-López, C.; Capó-Ortega, M.A. Locations of endangered spruce populations in Mexico and the demography of Picea chihuahuana. Madroño 2000, 47, 71–88. [Google Scholar]
- Fritts, H. Tree and Climate; Academic Press: New York, NY, USA, 1976; p. 567. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; The University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Douglas, A.E. Crossdating in dendrochronology. J. For. 1941, 39, 825–832. [Google Scholar]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 69. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Aplications in Dendroclimatology and Hidrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Merian, P.; Qeadan, F.; Zang, C.; Pucha, C.; Wernick, J. dplR: Dendrochronology Program Library in R. R Package Version 1.6.7. 2018. Available online: https://r-forge.r-project.org/projects/dplr/ (accessed on 10 April 2025).
- Cook, E.R. The decomposition of tree-ring series for environmental studies. Tree-Ring Bull. 1987, 47, 37–59. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. (Eds.) Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Fritts, H. Reconstructing Large-Scale Climatic Patterns from Tree-Ring Data: A Diagnostic Analysis; The University of Arizona Press: Tucson, AZ, USA, 1991; p. 286. [Google Scholar]
- Stahle, D.W.; Cook, E.R.; Burnette, D.J.; Torbenson, M.C.A.; Howard, I.M.; Griffin, D.; Villanueva-Diaz, J.; Cook, B.I.; Williams, A.P.; Watson, E.; et al. Dynamics, variability, and change in seasonal precipitation reconstructions for North America. J. Clim. 2020, 33, 3173–3194. [Google Scholar] [CrossRef]
- Burnette, D.J. The Tree-Ring Drought Atlas Portal: Gridded drought reconstructions for the past 500–2000 years. BAMS 2021, 102, 953–956. [Google Scholar] [CrossRef]
- Trouet, V.; Van Oldenborgh, G.J. KNMI Climate Explorer: A web-based research tool for high-resolution paleoclimatology. Tree-Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef]
- Mann, M.E.; Lees, J.M. Robust estimation of background noise and signal detection in climatic time series. Clim. Chang. 1996, 33, 409–445. [Google Scholar] [CrossRef]
- McPhaden, M.J.; Zebiak, S.E.; Glantz, M.H. ENSO as an integrating concept in earth science. Science 2006, 314, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Mata, L.; Astudillo-Sánchez, C.C.; Alanís-Rodríguez, E.; Treviño-Carreón, J.; Mora-Olivo, A.; Soria-Díaz, L. Dendrochronological potential of weeping pinyon (Pinus pinceana Gordon) in Tamaulipas, Mexico. Rev. Chapingo Ser. Cienc. For. Y Del Ambiente 2024, 30, 107. [Google Scholar] [CrossRef]
- Villanueva-Díaz, J.; Vázquez-Selem, L.; Estrada-Ávalos, J.; Martínez-Sifuentes, A.R.; Cerano-Paredes, J.; Canizales-Velázquez, P.A.; Franco-Ramos, O.; Reyes-Camarillo, F.R. Hydroclimatic Behavior of Conifers at Cerro El Potosí, Nuevo León, México. Rev. Mex. Cien. For. 2018, 9, 166–187. [Google Scholar]
- Arroyo-Morales, S.; Astudillo-Sanchez, C.C.; Aguirrre-Calderon, O.A.; Villanueva-Diaz, J.; Soria-Diaz, L.; Martinez-Sifuentes, A.R. A precipitation reconstruction based on pinyon pine tree rings from the northeastern Mexican subtropic. Theor. Appl. Climatol. 2023, 151, 635–649. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Lebrija-Trejo, E.; Zuidema, P.A.; Martinez-Ramos, M. Climate-growth analysis for a Mexican dry forest tree shows strong impact of sea surface temperatures and predicts future growth declines. Glob. Change Biol. 2010, 16, 2001–2012. [Google Scholar] [CrossRef]
- Stahle, D.W.; Villanueva-Diaz, J.; Burnette, D.J.; Cerano-Paredes, J.; Heim, R.R., Jr.; Fye, F.K.; Acuna-Soto, R.; Therrell, M.D.; Cleaveland, M.K.; Stahle, D.K. Major Mesoamerican droughts of the past millennium. Geophys. Res. Lett. 2011, 38, 2–5. [Google Scholar] [CrossRef]
- Gutiérrez-García, G.; Ricker, M. Climate influence on radial growth in four conifer species in the Sierra de San Antonio Peña Nevada (Nuevo León, Mexico). Rev. Mex. Biodivers. 2019, 90, e902676. [Google Scholar]
- Panthi, S.; Bräuning, A.; Zhou, Z.K.; Fan, Z.X. Tree rings reveal recent intensified spring drought in the central Himalaya, Nepal. Glob. Planet. Chang. 2017, 157, 26–34. [Google Scholar] [CrossRef]
- Zhu, L.; Cooper, D.J.; Yang, J.; Zhang, X.; Wang, X. Rapid warming induces the contrasting growth of Yezo spruce (Picea jezoensis var. microsperma) at two elevation gradient sites of northeast China. Dendrochronologia 2018, 50, 52–63. [Google Scholar] [CrossRef]
- Lefèvre, F.; Boivin, T.; Bontemps, A.; Courbet, F.; Davi, H.; Durand-Gillmann, M.; Fady, B.; Gauzere, J.; Gidoin, C.; Karam, M.-J.; et al. Considering evolutionary processes in adaptive forestry. Ann. For. Sci. 2013, 71, 723–739. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Fonti, P.; Shishov, V.; Rozenberg, P. Wood formation and tree adaptation to climate. Ann. For. Sci. 2019, 76, 109. [Google Scholar] [CrossRef]
- Meko, D.M.; Stockton, C.W.; Boggess, W.R. The tree-ring record of severe sustained drought. J. Am. Water Resour. Assoc. 1995, 31, 789–801. [Google Scholar] [CrossRef]
- Lauer, W.; Klaus, D. Geoecological investigations on the timberline of Pico de Orizaba, México. Artic Alp. Res. 1975, 7, 315–330. [Google Scholar] [CrossRef]
- IPCC. Climate Change: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carphians (Romania). Trees 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Sánchez-Santillán, N.; Garduño-López, R.; Vidal-Zepeda, R.; Sánchez-Trejo, R. Climate change in NE Mexico: Influence of the North Atlantic Oscillation. Investig. Geográficas 2012, 78, 7–18. [Google Scholar] [CrossRef]
- Wehenkel, C.; González-Elizondo, M.S.; Mendoza-Maya, E.; Hernández-Diaz, J.C.; Quiñones-Pérez, C.Z. Distribución, ecología y estado de conservación de Las Piceas Mexicanas. Las Piceas (Picea, Pinaceae) de México, 1st ed.; González Elizondo, M.S., Wehenkel, C., Eds.; Comisión Nacional Forestal: Zapopan, México, 2022; pp. 28–55. [Google Scholar]
- Rosengaus, M.M.; Jimenez-Espinoza, M.; Vázquez-Conde, M.T. Atlas Climatológico de Ciclones Tropicales en Mexico. Centro Nacional de Prevención de Desatres. Cd. de México, México. 2022. Available online: https://www.cenapred.unam.mx/es/Publicaciones/archivos/37.pdf (accessed on 22 April 2025).
- Schweingruber, F.H. Tree Rings and Environment: Dendroecology; Birmennsdorf, Swish Federal Institute for Forest, Snow, and Landscape Research; Berne, Stuttgart, Vienna, Haupt; WSL/FNP: Birmendsdorf, Switzerland, 1996; p. 609. [Google Scholar]
- Stahle, D.W.; Cook, E.R.; Villanueva-Diaz, J.; Fye, F.K.; Burnnette, D.J.; Griffin, D.; Acuna-Soto, R.; Seager, R.; Heim, R.R., Jr. Early 21st-century drought in Mexico. Eos Trans. Am. Geophys. Union 2009, 90, 69–90. [Google Scholar] [CrossRef]
- Florescano, M.E.; Swan, S. Breve Historia de la Sequía en México; Biblioteca Universidad Veracruzana: Veracruz, México, 1995; p. 246. [Google Scholar]
- Seager, R.; Ting, M.; Davis, M.; Cane, M.; Naik, N.; Nakamura, J.; Li, C.; Cook, E.; Stahle, D.W. Mexican drought: An observational modeling and tree ring study of variability and climate change. Atmósfera 2009, 22, 1–31. [Google Scholar]
- Cleaveland, M.K.; Stahle, D.W.; Therrell, M.D.; Villanueva-Diaz, J.; Burns, B.T. Tree-ring reconstructed winter precipitation and tropical teleconnections in Durango, Mexico. Clim. Chang. 2003, 59, 369–388. [Google Scholar] [CrossRef]
- Mijarez-Fajardo, R.; Lobato-Sánchez, R.; Patiño-Gómez, C.; Guevara-Polo, D.E. Atlantic and Pacific Sea surface temperature correlations with precipitation over northern Mexico. Atmósfera 2023, 38, 217–234. [Google Scholar] [CrossRef]
- Comrie, A.C.; Glenn, E.C. Principal components-based regionalization of precipitation regimes across the southwest United States and northern Mexico, with an application to monsoon precipitation variability. Clim. Res. 1999, 10, 201–215. [Google Scholar] [CrossRef]
- Stahle, D.W.; D’Arrigo, R.D.; Krusic, P.J.; Ceaveland, M.K.; Cook, E.R.; Allan, R.J.; Cole, J.E.; Dunbar, R.B.; Therrell, M.D.; Gay, D.A.; et al. Experimental dendroclimatic reconstruction of the Southern Oscillation. Bull. Am. Meteorol. Soc. 1998, 79, 2137–2152. [Google Scholar] [CrossRef]
- Magaña, V.; Pérez, J.L.; Conde, C. El Niño y la Oscilación de Sur, sus impactos en Mexico. Ciencias 2009, 51, 14–18. [Google Scholar]
- D’Arrigo, R.D.; Jacoby, G.C. A 1000-year record of winter precipitation from northwestern New Mexico, USA: A reconstruction from tree-rings and its relation to El Niño and the Southern Oscillation. Holocene 1991, 1, 95–101. [Google Scholar] [CrossRef]
- Cook, E.R.; Woodhouse, C.A.; Eakin, C.M.; Meko, D.M.; Stahle, D.W. Long-term aridity changes in the western United States. Science 2004, 306, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- George, S.S. An overview of tree-ring width records across the Northern Hemisphere. Quat. Sci. Rev. 2014, 95, 132–150. [Google Scholar] [CrossRef]
- Perone, A.; Lombardi, F.; Marchetti, M.; Tognetti, R.; Lasserre, B. Evidence of solar activity and El Niño signals in tree rings of Araucaria araucana and A. angustifolia in South America. Glob. Planet. Chang. 2016, 145, 1–10. [Google Scholar] [CrossRef]
- Rodriguez-Caton, M.; Andreu-Hayles, L.; Daux, V.; Vuille, M.; Varuolo-Clarke, A.M.; Oelkers, R.; Christie, D.A.; D’Arrigo, R.; Morales, M.S.; Rao, M.P.; et al. Hydroclimate and ENSO variability recorded by oxygen isotopes from tree rings in the South American Altiplano. Geophys. Res. Lett. 2022, 49, e2021GL095883. [Google Scholar] [CrossRef]
- Su, H.; Axmacher, J.C.; Yang, B.; Sang, W. Differential radial growth response of three coexisting dominant tree species to local and large-scale climate variability in a subtropical evergreen broad-leaved forest of China. Ecol. Res. 2015, 30, 745–754. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, M.; Wernicke, J.; Bräuning, A. Changes in sensitivity of tree-ring widths to climate in a tropical moist forest tree in Bangladesh. Forests 2018, 9, 761. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weather Station | Latitude N (°) | Longitude W (°) | Elevation (m) | Data Recording Period | Ppt (mm) | MaxT (°C) | MinT (°C) |
|---|---|---|---|---|---|---|---|
| Saltillo | 25.43 | −101.00 | 1700 | 1948–2013 | 578 | 25.7 | 11.1 |
| Ignacio de Texas | 24.31 | −100.18 | 1680 | 1981–2018 | 346 | 26.2 | 8.8 |
| Peña Nevada | 23.74 | −99.98 | 1491 | 1981–2018 | 392 | 24.8 | 7.6 |
| Site | ID | Age of Trees | Cores/Trees | Period | 1 MS | 2 AC1 | 3 EPS | 4 Rbar | 5 SNR |
|---|---|---|---|---|---|---|---|---|---|
| El Coahuilon | ECO | 11–224 | 53/25 | 1782–2020 | 0.25 | 0.03 | 0.91 | 0.32 | 11.3 |
| La Marta | LMA | 24–170 | 49/24 | 1844–2020 | 0.31 | 0.04 | 0.96 | 0.46 | 25.6 |
| Agua Fria | AFI | 34–173 | 40/25 | 1848–2020 | 0.38 | 0.01 | 0.86 | 0.30 | 6.6 |
| El Butano | BUT | 23–204 | 109/46 | 1736–2020 | 0.40 | 0.02 | 0.94 | 0.32 | 19.7 |
| La Encantada | LEN | 22–186 | 55/35 | 1833–2020 | 0.33 | 0.03 | 0.95 | 0.43 | 16.3 |
| Chronologies | ECO | LMA | AFI | BUT | LEN |
|---|---|---|---|---|---|
| ECO | 1.00 | - | - | - | - |
| LMA | 0.72 ** | 1.00 | - | - | - |
| AFI | 0.43 * | 0.51 ** | 1.00 | - | - |
| BUT | 0.53 ** | 0.58 ** | 0.51 ** | 1.00 | - |
| LEN | 0.28 * | 0.35 * | 0.39 * | 0.48 * | 1.00 |
| Pmex model: | |||||||
| Period | r | r2 | Reduction of Error | T-value | Sign-Products | Negative First Difference | |
| Calibration | 1961–1986 | 0.670 * | 0.449 * | 0.433 * | 3.16 * | 7 * | 9 NS |
| Verification | 1987–2011 | 0.560 * | 0.314 * | 0.147 * | 4.05 * | 13 NS | 8 NS |
| Pmar model: | |||||||
| Period | r | r2 | Reduction of Error | T-value | Sign-Products | Negative First Difference | |
| Calibration | 1961–1986 | 0.780 * | 0.610 * | 0.588 * | 3.69 * | 8 NS | 4 * |
| Verification | 1987–2011 | 0.630 * | 0.400 * | 0.415 * | 3.15 * | 5 * | 4 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wehenkel, C.; Díaz-Carrillo, O.A.; Villanueva-Díaz, J. Dendroclimatic Reconstruction of Seasonal Precipitation from Two Endangered Spruce Species in Northeastern Mexico. Atmosphere 2025, 16, 863. https://doi.org/10.3390/atmos16070863
Wehenkel C, Díaz-Carrillo OA, Villanueva-Díaz J. Dendroclimatic Reconstruction of Seasonal Precipitation from Two Endangered Spruce Species in Northeastern Mexico. Atmosphere. 2025; 16(7):863. https://doi.org/10.3390/atmos16070863
Chicago/Turabian StyleWehenkel, Christian, Oscar A. Díaz-Carrillo, and Jose Villanueva-Díaz. 2025. "Dendroclimatic Reconstruction of Seasonal Precipitation from Two Endangered Spruce Species in Northeastern Mexico" Atmosphere 16, no. 7: 863. https://doi.org/10.3390/atmos16070863
APA StyleWehenkel, C., Díaz-Carrillo, O. A., & Villanueva-Díaz, J. (2025). Dendroclimatic Reconstruction of Seasonal Precipitation from Two Endangered Spruce Species in Northeastern Mexico. Atmosphere, 16(7), 863. https://doi.org/10.3390/atmos16070863