Community Structure and Influencing Factors of Airborne Microbial Aerosols over Three Chinese Cities with Contrasting Social-Economic Levels

 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Microbial Aerosol Sampling
2.2. Bacterial Genomic DNA Extraction
2.3. Amplification and Sequencing
2.4. Data Processing and Statistical Analysis
2.5. Meteorological Data
3. Results
3.1. Sample Sequencing Data
3.2. Microbial Biodiversity and Community Structures
3.3. Variation of Bacterial Taxa based on the Phylum and Genus Levels
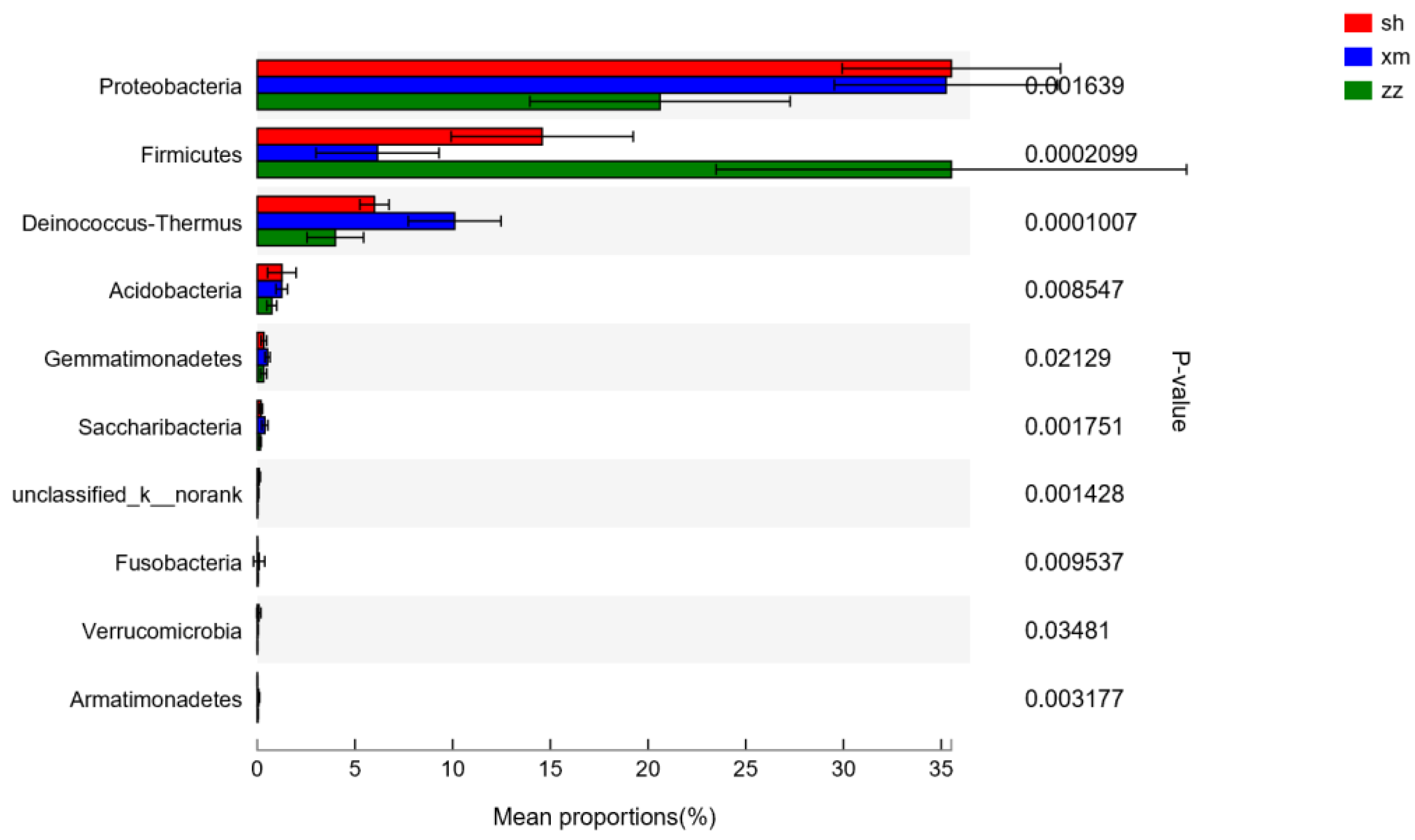
3.3.1. Characteristics of the Bacterial Community Structure at the Phylum Level
3.3.2. Characteristic of the Bacterial Community Structure at the Genus Level
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delort, A.; Vaïtilingom, M.; Amato, P.; Sancelme, M.; Parazols, M.; Mailhot, G.; Laj, P.; Deguillaume, L. A short overview of the microbial population in clouds: Potential roles in atmospheric chemistry and nucleation processes. Atmos. Res. 2010, 98, 249–260. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the Earth system: Climate, health, and ecosystem interactions. Atmos. Res. 1997, 15, 346–376. [Google Scholar] [CrossRef]
- Peccia, J.; Hospodsky, D.; Bibby, K. New Directions: A revolution in DNA sequencing now allows for the meaningful integration of biology with aerosol science. Atmos. Environ. 2010, 45, 1896–1897. [Google Scholar] [CrossRef]
- Den Boer, J.W.; Yzerman, E.P.F.; Schellekens, J.; Lettinga, K.D.; Boshuizen, H.C.; Van Steenbergen, J.E.; Bosman, A.; Van den Hof, S.; Van Vliet, H.A.; Peeters, M.F.; et al. A large outbreak of Legionnaires’ disease at a flower show, the Netherlands, 1999. Emerg. Infect. Dis. 2001, 8, 37–43. [Google Scholar] [CrossRef]
- Douwes, J.; Wouters, I.; Dubbeld, H.; van Zwieten, L.; Steerenberg, P.; Doekes, G.; Heederik, D. Upper airway inflammation assessed by nasal lavage in compost workers: A relation with bio-aerosol exposure. Am. J. Ind. Med. 2000, 37, 459–468. [Google Scholar] [CrossRef]
- Brągoszewska, E. Exposure to Bacterial and Fungal Aerosols: Microorganism Indices in A Waste-Sorting Plant in Poland. Int. J. Environ. Res. Public Health 2019, 16, 3308. [Google Scholar] [CrossRef]
- Wright, T.J.; Greene, V.W.; Paulus, H.J. Viable microorganisms in an urban atmosphere. J. Air Pollut. Control Assoc. 1969, 19, 337–341. [Google Scholar] [CrossRef]
- Thorne, P.S.; Kulhánková, K.; Yin, M.; Cohn, R.; Arbse, S.J.; Zeldin, D.C. Endotoxin exposure is a risk factor for asthma: The national survey of endotoxin in United States housing. Am. J. Respir Crit. Care Med. 2005, 172, 1371–1377. [Google Scholar] [CrossRef]
- Wan, G.H.; Chung, F.F.; Tang, C.S. Long-term surveillance of air quality in medical center operating rooms. Am. J. Infect. Control. 2010, 39, 302–308. [Google Scholar] [CrossRef]
- Urbano, R.; Palenik, B.; Gaston, C.J.; Prather, K.A. Detection and phylogenetic analysis of coastal bioaerosols using culture dependent and independent techniques. Biogeosci. Discuss. 2010, 7, 301–309. [Google Scholar] [CrossRef]
- Després, V.R.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.S.; Buryak, G.; Fröhlich Nowoisky, J.; Elbert, W.; Andreae, M.O.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B Chem. Phys. Meteorol. 2011, 64, 145–153. [Google Scholar] [CrossRef]
- Zhai, J.H.; Zhai, H. Analysis and identification of airborne fungi in Beijing and Nanjing. China Public Health 2000, 16, 1026. [Google Scholar]
- Yamaguchi, N.; Park, J.; Kodama, M.; Ichijo, T.; Baba, T.; Nasu, M. Changes in the airborne bacterial community in outdoor environments following Asian dust events. Microbes Environ. 2014, 29, 82–88. [Google Scholar] [CrossRef]
- Mayol, E.; Arrieta, J.M.; Jiménez, M.A.; Martínez-Asensio, A.; Garcias-Bonet, N.; Dachs, J.; González-Gaya, B.; Royer, S.-J.; Benítez-Barrios, V.M.; Fraile-Nuez, E.; et al. Long-range transport of airborne microbes over the global tropical and subtropical ocean. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Jones, S.E.; Newton, R.J.; McMahon, K.D. Potential for atmospheric deposition of bacteria to influence bacterioplankton communities. FEMS Microbiol. Ecol. 2008, 64, 388–394. [Google Scholar] [CrossRef]
- Monteil, C.L.; Bardin, M.; Morris, C.E. Features of air masses associated with the deposition of Pseudomonas syringae and Botrytis cinerea by rain and snowfall. ISME J. 2014, 8, 2290–2304. [Google Scholar] [CrossRef]
- Peter, H.; Hortnagl, P.; Reche, I.; Sommaruga, R. Bacterial diversity and composition during rain events with and without Saharan dust influence reaching a high mountain lake in the Alps. Environ. Microbiol. Rep. 2014, 6, 618–624. [Google Scholar] [CrossRef]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition rates of viruses and bacteria above the atmospheric boundary layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef]
- Yahya, R.Z.; Arrieta, J.M.; Cusack, M.; Duarte, C.M. Airborne Prokaryote and Virus Abundance Over the Red Sea. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Rahav, E.; Ovadia, G.; Paytan, A.; Herut, B. Contribution of airborne microbes to bacterial production and N2 fixation in seawater upon aerosol deposition. Geophys. Res. Lett. 2016, 43, 719–727. [Google Scholar] [CrossRef]
- Marín-Beltrán, I.; Logue, J.B.; Andersson, A.F.; Peters, F. Atmospheric deposition impact on bacterial community composition in the NW Mediterranean. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Brągoszewska, E.; Pastuszka, J.S. Influence of meteorological factors on the level and characteristics of culturable bacteria in the air in Gliwice, Upper Silesia (Poland). Aerobiologia 2018, 34, 241–255. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef]
- García Blázquez, G.; Göker, M.; Voglmayr, H.; Martín, M.P.; Tellería, M.T.; Oberwinkler, F. Phylogeny of Peronospora, parasitic on Fabaceae, based on ITS sequences. Mycol. Res. 2008, 112, 502–512. [Google Scholar] [CrossRef]
- Lang-Yona, N.; Lehahn, Y.; Herut, B.; Burshtein, N.; Rudich, Y. Marine aerosol as a possible source for endotoxins in coastal areas. Sci. Total Environ. 2014, 499, 311–318. [Google Scholar] [CrossRef]
- Robertson, C.E.; Baumgartner, L.K.; Harris, J.K.; Peterson, K.L.; Stevens, M.J.; Frank, D.N.; Pace, N.R. Culture-independent analysis of aerosol microbiology in a metropolitan subway system. Appl. Environ. Microbiol. 2013, 79, 3485–3493. [Google Scholar] [CrossRef]
- Bertolini, V.; Gandolfi, I.; Ambrosini, R.; Bestetti, G.; Innocente, E.; Rampazzo, G.; Franzetti, A. Temporal variability and effect of environmental variables on airborne bacterial communities in an urban area of Northern Italy. Appl. Microbiol. Biotechnol. 2013, 97, 6561–6570. [Google Scholar] [CrossRef]
- Gao, M.; Qiu, T.; Jia, R.; Han, M.; Song, Y.; Wang, X. Concentration and size distribution of viable bioaerosols during non-haze and haze days in Beijing. Environ. Sci. Pollut. Res. 2015, 22, 4359–4368. [Google Scholar] [CrossRef]
- Shao, C.C.; Zhen, Y.; Mi, T.Z.; Ma, M.M.; Feng, W.R. Characterization of Airborne Bacterial Communities in Qingdao during Haze and Non-haze Episodes in Winter. Urban Environ. Urban Ecol. 2016, 29, 7–13. [Google Scholar]
- Zhong, X.; Qi, J.H.; Li, H.T.; Dong, L.J.; Gao, D.M. Seasonal distribution of microbial activity in bioaerosols in the outdoor environment of the Qingdao coastal region. Atmos. Environ. 2016, 140, 506–513. [Google Scholar] [CrossRef]
- Liao, X.; Hu, A.Y.; Yang, X.Y.; Chen, J.S.; Yu, C.P.; Lin, J.Q. Community Composition and environmental sources of bacterial and eukaryotic microbiomes in fine air particles(PM2.5) of xiamen in winter. Ecol. Environ. Sci. 2013, 22, 1395–1400. [Google Scholar]
- Liao, X. Study on the Community Composition and Distribution Characteristics of Microorganisms in the Air of Xiamen; Jimei University: Xiamen, China, 2010. [Google Scholar]
- Li, Y.P.; Fu, H.L.; Wang, W.; Liu, J.; Meng, Q.L.; Wang, W.K. Characteristics of bacterial and fungal aerosols during the autumn haze days in Xi’an, China. Atmos. Environ. 2015, 122, 439–447. [Google Scholar] [CrossRef]
- Gou, H.G.; Lu, J.J.; Li, S.M.; Tong, Y.B.; Xie, C.B.; Zheng, X.W. Assessment of microbial communities in PM1 and PM10 of Urumqi during winter. Environ. Pollut. 2016, 214, 202–210. [Google Scholar] [CrossRef]
- Xu, C.; Wei, M.; Chen, J.; Wang, X.; Zhu, C.; Li, J.; Zheng, L.; Sui, G.; Li, W.; Wang, W.; et al. Bacterial characterization in ambient submicron particles during severe haze episodes at Ji’nan, China. Sci. Total Environ. 2016, 580, 188–196. [Google Scholar] [CrossRef]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef]
- Friedrich, V.W.; Göbel Ulf, B.; Erko, S. Determination of microbial diversity in environmental samples: Pitfalls of pcr-based rrna analysis. FEMS Microbiol. Rev. 1997, 21, 213–229. [Google Scholar] [CrossRef]
- Trimmomatic: A Flexible Read Trimming Tool for Illumina NGS Data. Available online: http://www.usadellab.org/cms/?page=trimmomatic (accessed on 2 February 2020).
- FLASh: Fast Length Adjustment of Short Reads. Available online: https://ccb.jhu.edu/software/FLASH/ (accessed on 2 February 2020).
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef]
- QIIME: Quantitative Insights into Microbial Ecology. Available online: http://qiime.org/scripts/assign_taxonomy.html (accessed on 2 February 2020).
- USEARCH v11. Available online: http://www.drive5.com/usearch/manual/chimera_formation.html (accessed on 2 February 2020).
- UCHIME Algorithm. Available online: http://www.drive5.com/usearch/manual/uchime_algo.html (accessed on 2 February 2020).
- WU: Weather Underground. Available online: https://www.wunderground.com (accessed on 2 February 2020).
- Draxler, R.; Rolph, G. HYSPLIT-Hybrid Single Particle Lagrangian Integrated Trajectory Model. 2012. Available online: https://ready.arl.noaa.gov/HYSPLIT.php (accessed on 2 February 2020).
- Mu, F.; Li, Y.; Lu, R.; Qi, Y.; Xie, W.; Bai, W. Source identification of airborne bacteria in the mountainous area and the urban areas. Atmos. Res. 2020, 231, 104676. [Google Scholar] [CrossRef]
- Genitsaris, S.; Stefanidou, N.; Katsiapi, M.; Kormas, K.; Sommer, U.; Moustaka-Gouni, M. Variability of airborne bacteria in an urban Mediterranean area (Thessaloniki, Greece). Atmos. Environ. 2017, 157, 101–110. [Google Scholar] [CrossRef]
- Amato, P.; Parazols, M.; Sancelme, M.; Laj, P.; Mailhot, G.; Delort, A.M. Microorganisms isolated from the water phase of tropospheric clouds at the Puy de Dôme: Major groups and growth abilities at low temperatures. FEMS Microbiol. Ecol. 2007, 59, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Reeve, J.N. Bacterial recovery from ancient glacial ice. Environ. Microbiol. 2003, 5, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.L.; Wang, S.G.; Guo, W. Effect of urea application amount on microbes and enzymes in soil. Chin. J. Soil Sci. 2004, 35, 303–306. [Google Scholar] [CrossRef]
- Amatoab, P.; Parazolsbc, M.; Sancelmea, M.; Mailhotc, G.; Lajb, P.; Delort, A.M. An important oceanic source of micro-organisms for cloud water at the Puy de Dôme (France). Atmos. Environ. 2007, 41, 8253–8263. [Google Scholar] [CrossRef]
- Côté, V.; Kos, G.; Mortazavi, R.; Ariya, P.A. Microbial and “de novo” transformation of dicarboxylic acids by three airborne fungi. Sci. Total Environ. 2007, 390, 530–537. [Google Scholar] [CrossRef]
- Perfumo, A.; Marchant, R. Global transport of thermophilic bacteria in atmospheric dust. Environ. Microbiol. Rep. 2010, 2, 333–339. [Google Scholar] [CrossRef]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Zagorodnov, V.; Reeve, J.N. Recovery and Identification of Viable Bacteria Immured in Glacial Ice. Iearus 2000, 144, 479–485. [Google Scholar] [CrossRef]
- Bowers, R.M.; Lauber, C.L.; Wiedinmyer, C.; Hamady, M.; Hallar, A.G.; Fall, R.; Knight, R.; Fierer, N. Characterization of airborne microbial communities at a high-elevation site and their potential to act as atmospheric ice nuclei. Appl. Environ. Microbiol. 2009, 75, 5121–5130. [Google Scholar] [CrossRef]
- Fang, Z.G.; Ouyang, Z.Y.; Zhao, J.Z.; Wang, X.K.; Zheng, H. Community composition and dynamics of airborne bacteria in Beijing. Acta Microbiol. Sin. 2006, 46, 618–623. [Google Scholar] [CrossRef]
- Moon, K.W.; Huh, E.H.; Jeong, H.C. Seasonal evaluation of bioaerosols from indoor air of residential apartments within the metropolitan area in South Korea. Environ. Monit. Assess. 2014, 186, 2111–2120. [Google Scholar] [CrossRef]
- Zhao, Y.B. Studies on Air Bacterial Community Structure at Urban Streets and Wetland in Qingdao; Qingdao University of Technology: Shaidong, China, 2015. [Google Scholar]
- Bowers, R.M.; Sullivan, A.P.; Costello, E.K.; Collett, J.L.; Knight, R.; Fierer, N. Sources of bacteria in outdoor air across cities in the midwestern United States. Appl. Environ. Microbiol. 2011, 77, 6350–6356. [Google Scholar] [CrossRef]
- Goudarzi, G.; Shirmardi, M.; Khodarahmi, F.; Hashemi-Shahraki, A.; Alavi, N.; Ankali, K.A.; Babaei, A.A.; Soleimani, Z.; Marzouni, M.B. Particulate matter and bacteria characteristics of the Middle East Dust (MED) storms over Ahvaz, Iran. Aerobiologia 2014, 30, 345–356. [Google Scholar] [CrossRef]
- Attard, E.; Yang, H.; Delort, A.-M.; Amato, P.; Poschl, U.; Glaux, C.; Koop, T.; Morris, C.E. Effects of atmospheric conditions on ice nucleation activity of Pseudomonas. Atmos. Chem. Phys. 2012, 12, 10667–10677. [Google Scholar] [CrossRef]
- Mouli, P.C.; Mohan, S.V.; Reddy, S.J. Assessment of microbial (bacteria) concentrations of ambient air at semi-arid urban region: Influence of meteorological factors. Appl. Ecol. Environ. Res. 2005, 3, 139–149. [Google Scholar] [CrossRef]
- Zhang, X. Structure of Airborne Microbial Community and Influencing Factors; Zhejiang University: Zhejiang, China, 2016. [Google Scholar]
- Maki, T.; Hara, K.; Kobayashi, F.; Kurosaki, Y.; Kakikawa, M.; Matsuki, A.; Chen, B.; Shi, G.; Hasegawa, H.; Iwasaka, Y. Vertical distribution of airborne bacterial communities in an Asiandust downwind area, Noto Peninsula. Atmos. Environ. 2015, 119, 282–293. [Google Scholar] [CrossRef]
- Gat, D.; Mazar, Y.; Cytryn, E.; Rudich, Y. Origin-Dependent Variations in the Atmospheric Microbiome Community in Eastern Mediterranean Dust Storms. Environ. Sci. Technol. 2017, 51, 6709–6718. [Google Scholar] [CrossRef]
- Katra, I.; Arotsker, L.; Krasnov, H.; Zaritsky, A.; Kushmaro, A.; Ben-Dov, E. Richness and Diversity in Dust Stormborne Biomes at the Southeast Mediterranean. Sci. Rep. 2014, 4, 5265. [Google Scholar] [CrossRef]
- Zhangzhou People’s Government. Available online: www.zhangzhou.gov.cn/cms/html/zzsrmzf/2017 (accessed on 2 February 2020).
- Hou, L.Y.; Hu, A.Y.; Ma, Y.; Yu, C.P. Distribution of potential pathogenic bacteria in the Jiulong River Watershed. Environ. Sci. 2014, 35, 1742–1749. [Google Scholar] [CrossRef]
- Peeler, K.A.; Opsahl, S.P.; Chanton, J.P. Tracking anthropogenic inputs using caffeine, indicator bacteria, and nutrients in rural freshwater and urban marine systems. Environ. Sci. Technol. 2006, 40, 7616–7622. [Google Scholar] [CrossRef]
- Iguchi, A.; Nagaya, Y.; Pradel, E.; Ooka, T.; Ogura, Y.; Katsura, K.; Kurokawa, K.; Oshima, K.; Hattori, M.; Parkhill, J.; et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol. Evol. 2014, 6, 2096–2110. [Google Scholar] [CrossRef]
- Hurtado, L.; Rodríguez, G.; Lopez, J.; Castillo, J.; Molina, L.; Zavala, M.; Quintana, P. Characterization of atmospheric bioaerosols at 9 sites in Tijuana, Mexico. Atmos. Environ. 2014, 96, 430–436. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Sh | Xm | Zz |
|---|---|---|---|
| Proteobacteria | 35.61 ± 5.59 | 35.16 ± 5.72 | 20.92 ± 6.66 |
| Actinobacteria | 25.69 ± 3.69 | 28.09 ± 6.44 | 23.87 ± 3.14 |
| Firmicutes | 13.82 ± 4.66 | 6.12 ± 3.15 | 34.82 ± 12.04 |
| Bacteroidetes | 5.10 ± 0.64 | 4.71 ± 2.55 | 4.04 ± 1.40 |
| Cyanobacteria | 9.96 ± 4.38 | 11.59 ± 5.36 | 9.55 ± 2.69 |
| Deinococcus-Thermus | 5.95 ± 0.75 | 10.18 ± 2.37 | 4.09 ± 1.45 |
| Acidobacteria | 1.29 ± 0.73 | 1.25 ± 0.29 | 0.75 ± 0.25 |
| Chloroflexi | 1.53 ± 0.94 | 1.53 ± 0.38 | 1.21 ± 0.34 |
| Gemmatimonadetes | 0.35 ± 0.15 | 0.52 ± 0.13 | 0.33 ± 0.16 |
| Saccharibacteria | 0.17 ± 0.08 | 0.39 ± 0.16 | 0.16 ± 0.05 |
| Planctomycetes | 0.26 ± 0.29 | 0.08 ± 0.04 | 0.09 ± 0.04 |
| Bacteria-unclassified | 0.10 ± 0.06 | 0.05 ± 0.03 | 0.01 ± 0.01 |
| Nitrospirae | 0.03 ± 0.02 | 0.05 ± 0.03 | 0.03 ± 0.02 |
| Fusobacteria | 0.01 ± 0.01 | 0.10 ± 0.29 | 0.03 ± 0.01 |
| Latescibacteria | 0.01 ± 0.03 | n.a | n.a |
| Chlorobi | 0.02 * | 0.02 ± 0.06 | n.a |
| Armatimonadetes | 0.06 ± 0.05 | 0.03 ± 0.02 | |
| Spirochaetes | 0.02 ± 0.04 | 0.04 ± 0.04 | |
| SHA-109 | 0.02 ± 0.03 | 0.01 * | |
| Verrucomicrobia | 0.08 ± 0.10 | 0.03 ± 0.02 | |
| Deferribacteres | n.a | 0.02 ± 0.04 | |
| Parcubacteria | n.a | 0.01 * | |
| Chlamydiae | 0.02 ± 0.02 | 0.01 ± 0.02 | |
| TM6 | n.a | n.a | |
| Fibrobacteres | n.a | n.a | |
| Tenericutes | n.a | n.a | 0.01 ± 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, Y.; Li, H.; Chen, M.; Huang, K.; Chen, J.; Xu, J.; Zhuang, G. Community Structure and Influencing Factors of Airborne Microbial Aerosols over Three Chinese Cities with Contrasting Social-Economic Levels. Atmosphere 2020, 11, 317. https://doi.org/10.3390/atmos11040317
Rao Y, Li H, Chen M, Huang K, Chen J, Xu J, Zhuang G. Community Structure and Influencing Factors of Airborne Microbial Aerosols over Three Chinese Cities with Contrasting Social-Economic Levels. Atmosphere. 2020; 11(4):317. https://doi.org/10.3390/atmos11040317
Chicago/Turabian StyleRao, Ying, Heyang Li, Mingxia Chen, Kan Huang, Jia Chen, Jian Xu, and Guoshun Zhuang. 2020. "Community Structure and Influencing Factors of Airborne Microbial Aerosols over Three Chinese Cities with Contrasting Social-Economic Levels" Atmosphere 11, no. 4: 317. https://doi.org/10.3390/atmos11040317
APA StyleRao, Y., Li, H., Chen, M., Huang, K., Chen, J., Xu, J., & Zhuang, G. (2020). Community Structure and Influencing Factors of Airborne Microbial Aerosols over Three Chinese Cities with Contrasting Social-Economic Levels. Atmosphere, 11(4), 317. https://doi.org/10.3390/atmos11040317