

Being a Dog: A Review of the Domestication Process

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Canine Domestication Process
2.1. Sources of Selective Pressure: The Environment and Humans
2.2. One Experimental Domestication Model: Belyaev’s Foxes
2.3. A Possible Role for Oxytocin in Domestication
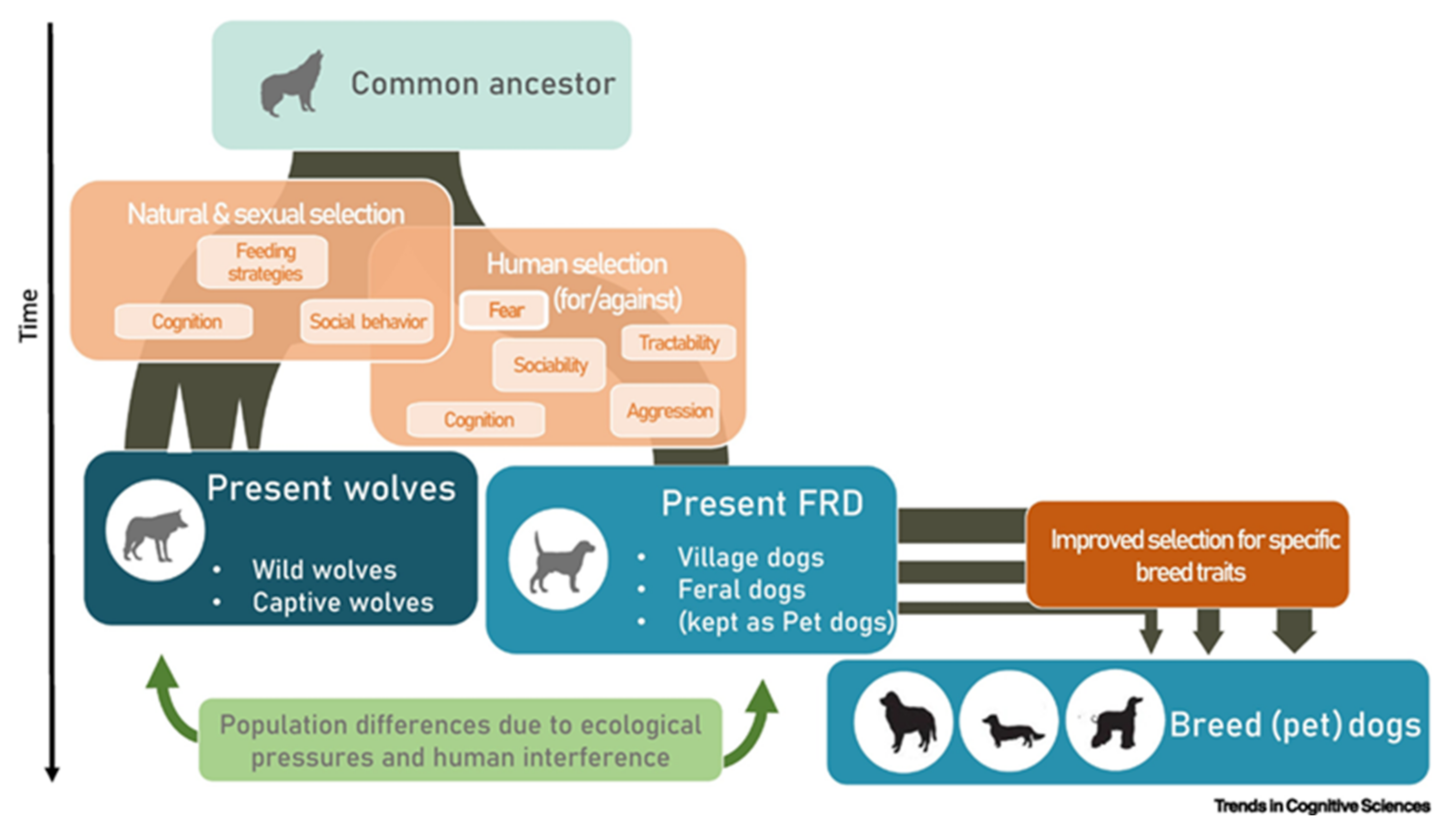
3. Wolves vs. Dogs: A Socio-Ecological Clash
4. Genetics of Ancient European Dogs
4.1. Canine Y Chromosome Marker
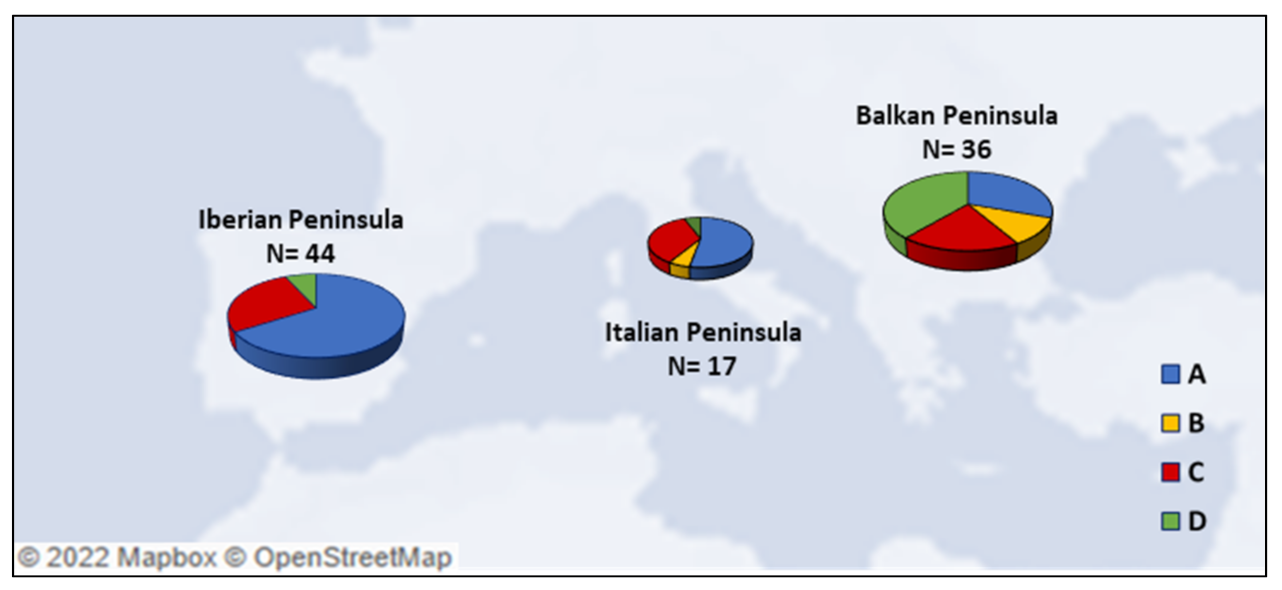
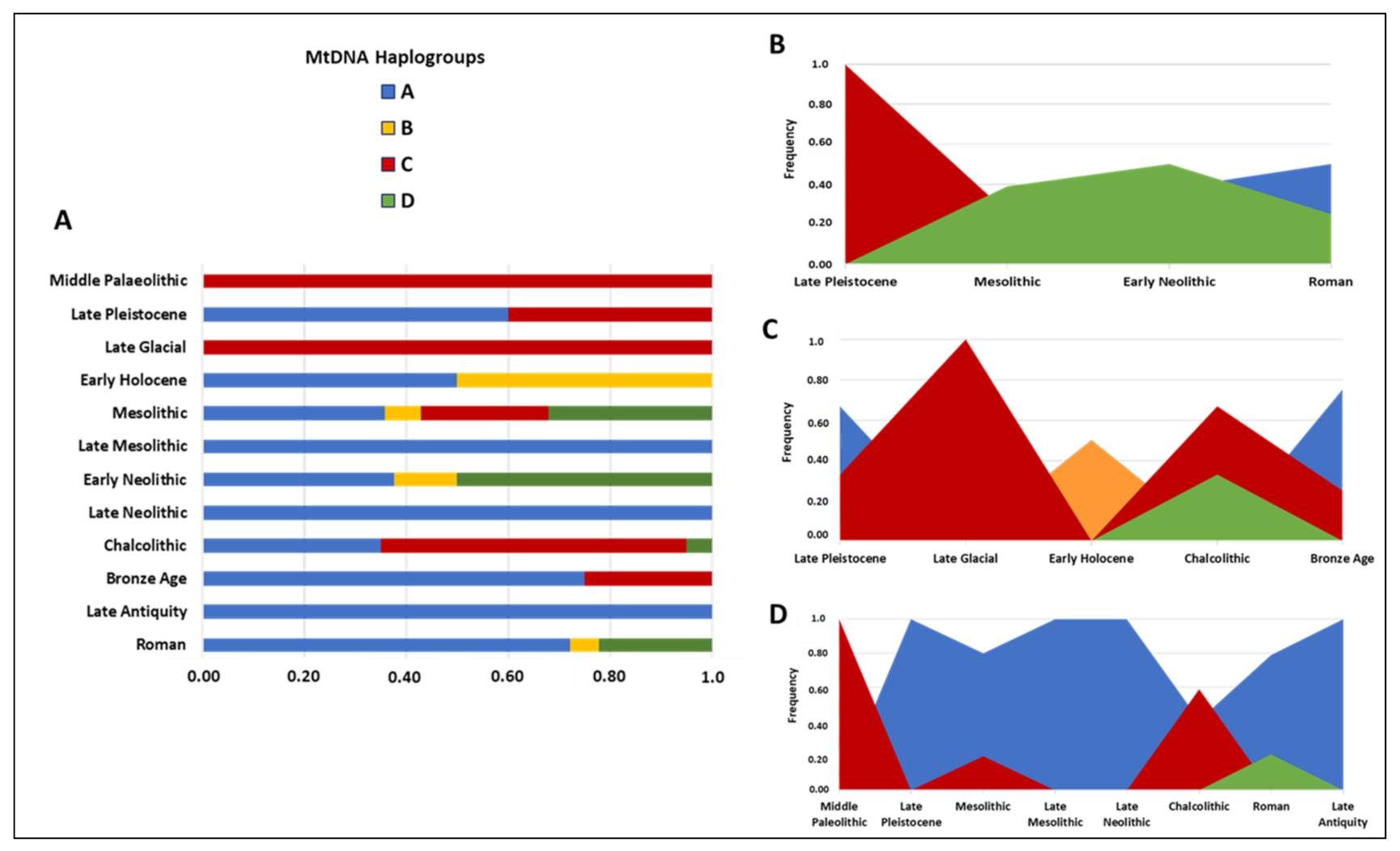
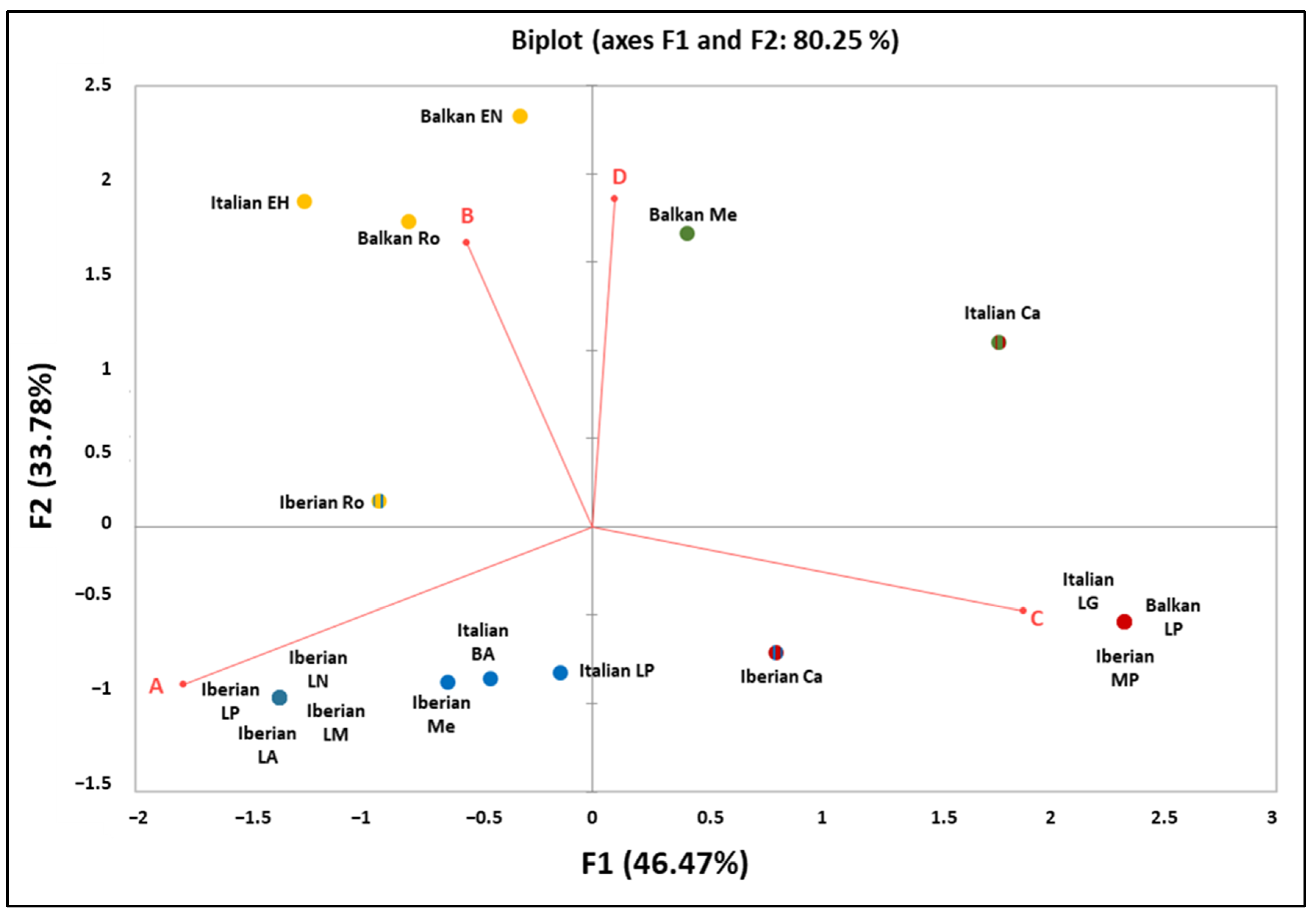
4.2. A Focus on the Ancient mtDNA Phylogeny in Three Mediterranean Peninsulas
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Purugganan, M.D. What Is Domestication? Trends Ecol. Evol. 2022, 37, 663–671. [Google Scholar] [CrossRef]
- Belyaev, D.K. Destabilizing Selection as a Factor in Domestication. J. Hered. 1979, 70, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current Perspectives and the Future of Domestication Studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Fuller, D.Q. The Evolution of Animal Domestication. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 115–136. [Google Scholar] [CrossRef]
- Vigne, J.-D. Early Domestication and Farming: What Should We Know or Do for a Better Understanding? Anthropozoologica 2015, 50, 123–150. [Google Scholar] [CrossRef]
- Zeder, M.A. Core Questions in Domestication Research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef]
- Rehfeld, K.; Münch, T.; Ho, S.L.; Laepple, T. Global Patterns of Declining Temperature Variability from the Last Glacial Maximum to the Holocene. Nature 2018, 554, 356–359. [Google Scholar] [CrossRef]
- Prato Previde, E.; Valsecchi, P. The Immaterial Cord. In The Social Dog; Elsevier: Amsterdam, The Netherlands, 2014; pp. 165–189. ISBN 978-0-12-407818-5. [Google Scholar]
- Oskarsson, M. Analysis of Origin and Spread of the Domestic Dog Using Y-Chromosome DNA and MtDNA Sequence Data; KTH Royal Institute of Technology, School of Biotechnology, Gene Technology: Stockholm, Sweden, 2012; ISBN 978-91-7501-364-0. [Google Scholar]
- McHugo, G.P.; Dover, M.J.; MacHugh, D.E. Unlocking the Origins and Biology of Domestic Animals Using Ancient DNA and Paleogenomics. BMC Biol. 2019, 17, 98. [Google Scholar] [CrossRef]
- Freedman, A.H.; Gronau, I.; Schweizer, R.M.; Ortega-Del Vecchyo, D.; Han, E.; Silva, P.M.; Galaverni, M.; Fan, Z.; Marx, P.; Lorente-Galdos, B.; et al. Genome Sequencing Highlights the Dynamic Early History of Dogs. PLoS Genet. 2014, 10, e1004016. [Google Scholar] [CrossRef]
- Wang, G.-D.; Zhai, W.; Yang, H.-C.; Wang, L.; Zhong, L.; Liu, Y.-H.; Fan, R.-X.; Yin, T.-T.; Zhu, C.-L.; Poyarkov, A.D.; et al. Out of Southern East Asia: The Natural History of Domestic Dogs across the World. Cell Res. 2016, 26, 21–33. [Google Scholar] [CrossRef]
- Herbeck, Y.E.; Eliava, M.; Grinevich, V.; MacLean, E.L. Fear, Love, and the Origins of Canid Domestication: An Oxytocin Hypothesis. Compr. Psychoneuroendocrinol. 2022, 9, 100100. [Google Scholar] [CrossRef] [PubMed]
- Serpell, J.A. Commensalism or Cross-Species Adoption? A Critical Review of Theories of Wolf Domestication. Front. Vet. Sci. 2021, 8, 662370. [Google Scholar] [CrossRef]
- Mech, L.D.; Janssens, L.A.A. An Assessment of Current Wolf Canis Lupus Domestication Hypotheses Based on Wolf Ecology and Behaviour. Mammal Rev. 2022, 52, 304–314. [Google Scholar] [CrossRef]
- Shipman, P. How Do You Kill 86 Mammoths? Taphonomic Investigations of Mammoth Megasites. Quat. Int. 2015, 359–360, 38–46. [Google Scholar] [CrossRef]
- Wheat, C.H.; Larsson, L.; Berner, P.; Temrin, H. Hand-Reared Wolves Show Attachment Comparable to Dogs and Use Human Caregiver as a Social Buffer in the Strange Situation Test. bioRxiv 2020. [Google Scholar]
- Freedman, A.H.; Wayne, R.K. Deciphering the Origin of Dogs: From Fossils to Genomes. Annu. Rev. Anim. Biosci. 2017, 5, 281–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-D.; Fan, R.-X.; Zhai, W.; Liu, F.; Wang, L.; Zhong, L.; Wu, H.; Yang, H.-C.; Wu, S.-F.; Zhu, C.-L.; et al. Genetic Convergence in the Adaptation of Dogs and Humans to the High-Altitude Environment of the Tibetan Plateau. Genome Biol. Evol. 2014, 6, 2122–2128. [Google Scholar] [CrossRef] [PubMed]
- Range, F.; Marshall-Pescini, S. Comparing Wolves and Dogs: Current Status and Implications for Human ‘Self-Domestication. ’ Trends Cogn. Sci. 2022, 26, 337–349. [Google Scholar] [CrossRef]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The Genomic Signature of Dog Domestication Reveals Adaptation to a Starch-Rich Diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Cafazzo, S.; Bonanni, R.; Valsecchi, P.; Natoli, E. Social Variables Affecting Mate Preferences, Copulation and Reproductive Outcome in a Pack of Free-Ranging Dogs. PLoS ONE 2014, 9, e98594. [Google Scholar] [CrossRef]
- Marshall-Pescini, S.; Cafazzo, S.; Virányi, Z.; Range, F. Integrating Social Ecology in Explanations of Wolf–Dog Behavioral Differences. Curr. Opin. Behav. Sci. 2017, 16, 80–86. [Google Scholar] [CrossRef]
- Statham, M.J.; Trut, L.N.; Sacks, B.N.; Kharlamova, A.V.; Oskina, I.N.; Gulevich, R.G.; Johnson, J.L.; Temnykh, S.V.; Acland, G.M.; Kukekova, A.V. On the Origin of a Domesticated Species: Identifying the Parent Population of Russian Silver Foxes (Vulpes Vulpes): The Origin of Russian Silver Foxes. Biol. J. Linn. Soc. 2011, 103, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Lord, K.A.; Larson, G.; Coppinger, R.P.; Karlsson, E.K. The History of Farm Foxes Undermines the Animal Domestication Syndrome. Trends Ecol. Evol. 2020, 35, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Trut, L.N. Early Canid Domestication: The Farm-Fox Experiment: Foxes Bred for Tamability in a 40-Year Experiment Exhibit Remarkable Transformations That Suggest an Interplay between Behavioral Genetics and Development. Am. Sci. 1999, 87, 160–169. [Google Scholar] [CrossRef]
- Dugatkin, L.A. The Silver Fox Domestication Experiment. Evol. Edu. Outreach 2018, 11, 16. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal Evolution during Domestication: The Domesticated Fox as a Model. BioEssays 2009, 31, 349–360. [Google Scholar] [CrossRef]
- Herbeck, Y.E.; Gulevich, R.G.; Eliava, M.; Shepeleva, D.V.; Trut, L.N.; Grinevich, V. Domestication: Neuroendocrine Mechanisms of Canidae -Human Bonds. In Model Animals in Neuroendocrinology; Ludwig, M., Levkowitz, G., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2018; pp. 313–334. ISBN 978-1-119-39112-8. [Google Scholar]
- Sánchez-Villagra, M.R.; Geiger, M.; Schneider, R.A. The Taming of the Neural Crest: A Developmental Perspective on the Origins of Morphological Covariation in Domesticated Mammals. R. Soc. Open Sci. 2016, 3, 160107. [Google Scholar] [CrossRef]
- Wilkins, A.S.; Wrangham, R.; Fitch, W.T. The Neural Crest/Domestication Syndrome Hypothesis, Explained: Reply to Johnsson, Henriksen, and Wright. Genetics 2021, 219, iyab098. [Google Scholar] [CrossRef]
- Hecht, E.E.; Smaers, J.B.; Dunn, W.D.; Kent, M.; Preuss, T.M.; Gutman, D.A. Significant Neuroanatomical Variation Among Domestic Dog Breeds. J. Neurosci. 2019, 39, 7748–7758. [Google Scholar] [CrossRef]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef]
- Pendleton, A.L.; Shen, F.; Taravella, A.M.; Emery, S.; Veeramah, K.R.; Boyko, A.R.; Kidd, J.M. Comparison of Village Dog and Wolf Genomes Highlights the Role of the Neural Crest in Dog Domestication. BMC Biol. 2018, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Hansen Wheat, C.; van der Bijl, W.; Wheat, C.W. Morphology Does Not Covary with Predicted Behavioral Correlations of the Domestication Syndrome in Dogs. Evol. Lett. 2020, 4, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Lord, K.A.; Larson, G.; Karlsson, E.K. Brain Size Does Not Rescue Domestication Syndrome. Trends Ecol. Evol. 2020, 35, 1061–1062. [Google Scholar] [CrossRef]
- Wilkins, A.S. A Striking Example of Developmental Bias in an Evolutionary Process: The “Domestication Syndrome”. Evol. Dev. 2020, 22, 143–153. [Google Scholar] [CrossRef]
- Zeder, M.A. Straw Foxes: Domestication Syndrome Evaluation Comes Up Short. Trends Ecol. Evol. 2020, 35, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, M.; Henriksen, R.; Wright, D. The Neural Crest Cell Hypothesis: No Unified Explanation for Domestication. Genetics 2021, 219, iyab097. [Google Scholar] [CrossRef]
- Wilson, L.A.B.; Balcarcel, A.; Geiger, M.; Heck, L.; Sánchez-Villagra, M.R. Modularity Patterns in Mammalian Domestication: Assessing Developmental Hypotheses for Diversification. Evol. Lett. 2021, 5, 385–396. [Google Scholar] [CrossRef]
- von Holdt, B.M.; Shuldiner, E.; Koch, I.J.; Kartzinel, R.Y.; Hogan, A.; Brubaker, L.; Wanser, S.; Stahler, D.; Wynne, C.D.L.; Ostrander, E.A.; et al. Structural Variants in Genes Associated with Human Williams-Beuren Syndrome Underlie Stereotypical Hypersociability in Domestic Dogs. Sci. Adv. 2017, 3, e1700398. [Google Scholar] [CrossRef]
- Dai, L.; Carter, C.S.; Ying, J.; Bellugi, U.; Pournajafi-Nazarloo, H.; Korenberg, J.R. Oxytocin and Vasopressin Are Dysregulated in Williams Syndrome, a Genetic Disorder Affecting Social Behavior. PLoS ONE 2012, 7, e38513. [Google Scholar] [CrossRef]
- Knobloch, H.S.; Grinevich, V. Evolution of Oxytocin Pathways in the Brain of Vertebrates. Front. Behav. Neurosci. 2014, 8, 31. [Google Scholar] [CrossRef]
- Banerjee, P.; Joy, K.P.; Chaube, R. Structural and Functional Diversity of Nonapeptide Hormones from an Evolutionary Perspective: A Review. Gen. Comp. Endocrinol. 2017, 241, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Kenkel, W.M.; Paredes, J.; Yee, J.R.; Pournajafi-Nazarloo, H.; Bales, K.L.; Carter, C.S. Neuroendocrine and Behavioural Responses to Exposure to an Infant in Male Prairie Voles: Exposure to an Infant in Male Prairie Voles. J. Neuroendocrinol. 2012, 24, 874–886. [Google Scholar] [CrossRef]
- Feldman, R. Oxytocin and Social Affiliation in Humans. Horm. Behav. 2012, 61, 380–391. [Google Scholar] [CrossRef]
- Nave, G.; Camerer, C.; McCullough, M. Does Oxytocin Increase Trust in Humans? A Critical Review of Research. Perspect. Psychol. Sci. 2015, 10, 772–789. [Google Scholar] [CrossRef] [PubMed]
- Kosfeld, M.; Heinrichs, M.; Zak, P.J.; Fischbacher, U.; Fehr, E. Oxytocin Increases Trust in Humans. Nature 2005, 435, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Guastella, A.J.; Mitchell, P.B.; Dadds, M.R. Oxytocin Increases Gaze to the Eye Region of Human Faces. Biol. Psychiatry 2008, 63, 3–5. [Google Scholar] [CrossRef]
- Domes, G.; Steiner, A.; Porges, S.W.; Heinrichs, M. Oxytocin Differentially Modulates Eye Gaze to Naturalistic Social Signals of Happiness and Anger. Psychoneuroendocrinology 2013, 38, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Guo, C.; Han, S. Oxytocin Enhances Inter-Brain Synchrony during Social Coordination in Male Adults. Soc. Cogn. Affect. Neurosci. 2016, 11, 1882–1893. [Google Scholar] [CrossRef]
- Feldman, R. Sensitive Periods in Human Social Development: New Insights from Research on Oxytocin, Synchrony, and High-Risk Parenting. Dev. Psychopathol. 2015, 27, 369–395. [Google Scholar] [CrossRef]
- Lefevre, A.; Richard, N.; Jazayeri, M.; Beuriat, P.-A.; Fieux, S.; Zimmer, L.; Duhamel, J.-R.; Sirigu, A. Oxytocin and Serotonin Brain Mechanisms in the Nonhuman Primate. J. Neurosci. 2017, 37, 6741–6750. [Google Scholar] [CrossRef]
- Yoshida, M.; Takayanagi, Y.; Inoue, K.; Kimura, T.; Young, L.J.; Onaka, T.; Nishimori, K. Evidence That Oxytocin Exerts Anxiolytic Effects via Oxytocin Receptor Expressed in Serotonergic Neurons in Mice. J. Neurosci. 2009, 29, 2259–2271. [Google Scholar] [CrossRef]
- Pagani, J.H.; Williams Avram, S.K.; Cui, Z.; Song, J.; Mezey, É.; Senerth, J.M.; Baumann, M.H.; Young, W.S. Raphe Serotonin Neuron-Specific Oxytocin Receptor Knockout Reduces Aggression without Affecting Anxiety-like Behavior in Male Mice Only: 5-HT Oxtr KO Decouples Aggression from Anxiety. Genes Brain Behav. 2015, 14, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Voitenko, N.N.; Kulikov, A.V.; Avgustinovich, D.F. Evidence for the Involvement of Central Serotonin in Mechanism of Domestication of Silver Foxes. Pharmacol. Biochem. Behav. 1991, 40, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Audero, E.; Mlinar, B.; Baccini, G.; Skachokova, Z.K.; Corradetti, R.; Gross, C. Suppression of Serotonin Neuron Firing Increases Aggression in Mice. J. Neurosci. 2013, 33, 8678–8688. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Miczek, K.A. Neurogenetics of Aggressive Behavior: Studies in Rodents. In Neuroscience of Aggression; Miczek, K.A., Meyer-Lindenberg, A., Eds.; Current Topics in Behavioral Neurosciences; Springer: Heidelberg, Berlin, 2013; Volume 17, pp. 3–44. ISBN 978-3-662-44280-7. [Google Scholar]
- MacLean, E.L.; Gesquiere, L.R.; Gee, N.R.; Levy, K.; Martin, W.L.; Carter, C.S. Effects of Affiliative Human–Animal Interaction on Dog Salivary and Plasma Oxytocin and Vasopressin. Front. Psychol. 2017, 8, 1606. [Google Scholar] [CrossRef]
- Handlin, L.; Hydbring-Sandberg, E.; Nilsson, A.; Ejdebäck, M.; Jansson, A.; Uvnäs-Moberg, K. Short-Term Interaction between Dogs and Their Owners: Effects on Oxytocin, Cortisol, Insulin and Heart Rate—An Exploratory Study. Anthrozoös 2011, 24, 301–315. [Google Scholar] [CrossRef]
- Nagasawa, M.; Kikusui, T.; Onaka, T.; Ohta, M. Dog’s Gaze at Its Owner Increases Owner’s Urinary Oxytocin during Social Interaction. Horm. Behav. 2009, 55, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Odendaal, J.S.J.; Meintjes, R.A. Neurophysiological Correlates of Affiliative Behaviour between Humans and Dogs. Vet. J. 2003, 165, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, T.; Mogi, K.; Kikusui, T. Oxytocin-Gaze Positive Loop and the Coevolution of Human-Dog Bonds. Science 2015, 348, 333–336. [Google Scholar] [CrossRef]
- Romero, T.; Nagasawa, M.; Mogi, K.; Hasegawa, T.; Kikusui, T. Oxytocin Promotes Social Bonding in Dogs. Proc. Natl. Acad. Sci. USA 2014, 111, 9085–9090. [Google Scholar] [CrossRef]
- Hernádi, A.; Kis, A.; Kanizsár, O.; Tóth, K.; Miklósi, B.; Topál, J. Intranasally Administered Oxytocin Affects How Dogs (Canis familiaris) React to the Threatening Approach of Their Owner and an Unfamiliar Experimenter. Behav. Process 2015, 119, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Crespi, B.J. Oxytocin, Testosterone, and Human Social Cognition: Oxytocin and Social Behavior. Biol. Rev. 2016, 91, 390–408. [Google Scholar] [CrossRef] [PubMed]
- Egito, J.H.; Nevat, M.; Shamay-Tsoory, S.G.; Osório, A.A.C. Oxytocin Increases the Social Salience of the Outgroup in Potential Threat Contexts. Horm. Behav. 2020, 122, 104733. [Google Scholar] [CrossRef]
- Anton, C.B.; Smith, D.W.; Suraci, J.P.; Stahler, D.R.; Duane, T.P.; Wilmers, C.C. Gray Wolf Habitat Use in Response to Visitor Activity along Roadways in Yellowstone National Park. Ecosphere 2020, 11, e03164. [Google Scholar] [CrossRef]
- Zepeda, E.; Payne, E.; Wurth, A.; Sih, A.; Gehrt, S. Early Life Experience Influences Dispersal in Coyotes (Canis latrans). Behav. Ecol. 2021, 32, 728–737. [Google Scholar] [CrossRef]
- Berghänel, A.; Lazzaroni, M.; Cimarelli, G.; Marshall-Pescini, S.; Range, F. Cooperation and Cognition in Wild Canids. Curr. Opin. Behav. Sci. 2022, 46, 101173. [Google Scholar] [CrossRef]
- Bentosela, M.; Wynne, C.D.L.; D’Orazio, M.; Elgier, A.; Udell, M.A.R. Sociability and Gazing toward Humans in Dogs and Wolves: Simple Behaviors with Broad Implications: Sociability and Gazing in Dogs and Wolves. J. Exp. Anal. Behav. 2016, 105, 68–75. [Google Scholar] [CrossRef]
- Wirobski, G.; Range, F.; Schaebs, F.S.; Palme, R.; Deschner, T.; Marshall-Pescini, S. Life Experience Rather than Domestication Accounts for Dogs’ Increased Oxytocin Release during Social Contact with Humans. Sci. Rep. 2021, 11, 14423. [Google Scholar] [CrossRef]
- Lord, K. A Comparison of the Sensory Development of Wolves (Canis lupus Lupus) and Dogs (Canis lupus familiaris). Ethology 2013, 119, 110–120. [Google Scholar] [CrossRef]
- Hansen Wheat, C.; van der Bijl, W.; Temrin, H. Dogs, but Not Wolves, Lose Their Sensitivity Toward Novelty With Age. Front. Psychol. 2019, 10, 2001. [Google Scholar] [CrossRef]
- Moretti, L.; Hentrup, M.; Kotrschal, K.; Range, F. The Influence of Relationships on Neophobia and Exploration in Wolves and Dogs. Anim. Behav. 2015, 107, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Mech, L.D.; Cluff, H.D. Prolonged Intensive Dominance Behavior Between Gray Wolves, Canis Lupus. Can. Field Nat. 2010, 124, 215. [Google Scholar] [CrossRef]
- Rao, A.; Bernasconi, L.; Lazzaroni, M.; Marshall-Pescini, S.; Range, F. Differences in Persistence between Dogs and Wolves in an Unsolvable Task in the Absence of Humans. PeerJ 2018, 6, e5944. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Pescini, S.; Besserdich, I.; Kratz, C.; Range, F. Exploring Differences in Dogs’ and Wolves’ Preference for Risk in a Foraging Task. Front. Psychol. 2016, 7, 1241. [Google Scholar] [CrossRef] [PubMed]
- Street, M.; Napierala, H.; Janssens, L. The Late Palaeolithic Dog from Bonn-Oberkassel in Context. In The Late Glacial Burial from Oberkassel Revisited; Giemsch, L., Schmitz, R.W., Eds.; Rheinische Ausgrabungen: Bonn, Germany, 2015; Volume 72, pp. 253–274. ISBN 978-3-8053-4970-3. [Google Scholar]
- Pionnier-Capitan, M.; Bemilli, C.; Bodu, P.; Célérier, G.; Ferrié, J.-G.; Fosse, P.; Garcià, M.; Vigne, J.-D. New Evidence for Upper Palaeolithic Small Domestic Dogs in South-Western Europe. J. Archaeol. Sci. 2011, 38, 2123–2140. [Google Scholar] [CrossRef]
- Koupadi, K.; Fontani, F.; Ciucani, M.M.; Maini, E.; De Fanti, S.; Cattani, M.; Curci, A.; Nenzioni, G.; Reggiani, P.; Andrews, A.J.; et al. Population Dynamics in Italian Canids between the Late Pleistocene and Bronze Age. Genes 2020, 11, 1409. [Google Scholar] [CrossRef]
- Wilczyński, J.; Haynes, G.; Sobczyk, Ł.; Svoboda, J.; Roblíčková, M. Friend or foe? Large canid remains from Pavlovian sites and their archaeozoological context. J. Anthropol. Archaeol. 2020, 59, 101197. [Google Scholar] [CrossRef]
- da Silva Coelho, F.A.; Gill, S.; Tomlin, C.M.; Heaton, T.H.; Lindqvist, C. An Early Dog from Southeast Alaska Supports a Coastal Route for the First Dog Migration into the Americas. Proc. R. Soc. B 2021, 288, 20203103. [Google Scholar] [CrossRef]
- Perri, A.R.; Feuerborn, T.R.; Frantz, L.A.F.; Larson, G.; Malhi, R.S.; Meltzer, D.J.; Witt, K.E. Dog Domestication and the Dual Dispersal of People and Dogs into the Americas. Proc. Natl. Acad. Sci. USA 2021, 118, e2010083118. [Google Scholar] [CrossRef]
- Vilà, C.; Savolainen, P.; Maldonado, J.E.; Amorim, I.R.; Rice, J.E.; Honeycutt, R.L.; Crandall, K.A.; Lundeberg, J.; Wayne, R.K. Multiple and Ancient Origins of the Domestic Dog. Science 1997, 276, 1687–1689. [Google Scholar] [CrossRef]
- Savolainen, P.; Zhang, Y.; Luo, J.; Lundeberg, J.; Leitner, T. Genetic Evidence for an East Asian Origin of Domestic Dogs. Science 2002, 298, 1610–1613. [Google Scholar] [CrossRef] [PubMed]
- Pakendorf, B.; Stoneking, M. Mitochondrial Dna and Human Evolution. Annu. Rev. Genom. Hum. Genet. 2005, 6, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, P.; Ersmark, E.; Palkopoulou, E.; Dalén, L. Ancient Wolf Genome Reveals an Early Divergence of Domestic Dog Ancestors and Admixture into High-Latitude Breeds. Curr. Biol. 2015, 25, 1515–1519. [Google Scholar] [CrossRef] [PubMed]
- Bergström, A.; Frantz, L.; Schmidt, R.; Ersmark, E.; Lebrasseur, O.; Girdland-Flink, L.; Lin, A.T.; Storå, J.; Sjögren, K.-G.; Anthony, D.; et al. Origins and Genetic Legacy of Prehistoric Dogs. Science 2020, 370, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Ollivier, M.; Tresset, A.; Frantz, L.A.F.; Bréhard, S.; Bălăşescu, A.; Mashkour, M.; Boroneanţ, A.; Pionnier-Capitan, M.; Lebrasseur, O.; Arbogast, R.-M.; et al. Dogs Accompanied Humans during the Neolithic Expansion into Europe. Biol. Lett. 2018, 14, 20180286. [Google Scholar] [CrossRef] [PubMed]
- Botigué, L.R.; Song, S.; Scheu, A.; Gopalan, S.; Pendleton, A.L.; Oetjens, M.; Taravella, A.M.; Seregély, T.; Zeeb-Lanz, A.; Arbogast, R.-M.; et al. Ancient European Dog Genomes Reveal Continuity since the Early Neolithic. Nat. Commun. 2017, 8, 16082. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Mullin, V.E.; Pionnier-Capitan, M.; Lebrasseur, O.; Ollivier, M.; Perri, A.; Linderholm, A.; Mattiangeli, V.; Teasdale, M.D.; Dimopoulos, E.A.; et al. Genomic and Archaeological Evidence Suggest a Dual Origin of Domestic Dogs. Science 2016, 352, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Kirkness, E.F.; Bafna, V.; Halpern, A.L.; Levy, S.; Remington, K.; Rusch, D.B.; Delcher, A.L.; Pop, M.; Wang, W.; Fraser, C.M.; et al. The Dog Genome: Survey Sequencing and Comparative Analysis. Science 2003, 301, 1898–1903. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J.; Zody, M.C.; et al. Genome Sequence, Comparative Analysis and Haplotype Structure of the Domestic Dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Jobling, M.A.; Tyler-Smith, C. The Human Y Chromosome: An Evolutionary Marker Comes of Age. Nat. Rev. Genet. 2003, 4, 598–612. [Google Scholar] [CrossRef]
- Underhill, P.A.; Kivisild, T. Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations. Annu. Rev. Genet. 2007, 41, 539–564. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.F.; Rozen, S. Genomics and Genetics of Human and Primate Y Chromosomes. Annu. Rev. Genom. Hum. Genet. 2012, 13, 83–108. [Google Scholar] [CrossRef]
- Cardinali, I.; Giontella, A.; Tommasi, A.; Silvestrelli, M.; Lancioni, H. Unlocking Horse Y Chromosome Diversity. Genes 2022, 13, 2272. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, C.; Genualdo, V.; Incarnato, D.; Mottola, F.; Perucatti, A.; Pauciullo, A. State of the Art on the Physical Mapping of the Y-Chromosome in the Bovidae and Comparison with Other Species—A Review. Anim Biosci 2022, 35, 1289–1302. [Google Scholar] [CrossRef]
- Lancioni, H.; Di Lorenzo, P.; Cardinali, I.; Ceccobelli, S.; Capodiferro, M.R.; Fichera, A.; Grugni, V.; Semino, O.; Ferretti, L.; Gruppetta, A.; et al. Survey of Uniparental Genetic Markers in the Maltese Cattle Breed Reveals a Significant Founder Effect but Does Not Indicate Local Domestication. Anim. Genet. 2016, 47, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Giontella, A.; Cardinali, I.; Pieramati, C.; Cherchi, R.; Biggio, G.P.; Achilli, A.; Silvestrelli, M.; Lancioni, H. A Genetic Window on Sardinian Native Horse Breeds through Uniparental Molecular Systems. Animals 2020, 10, 1544. [Google Scholar] [CrossRef] [PubMed]
- Bannasch, D.L.; Bannasch, M.J.; Ryun, J.R.; Famula, T.R.; Pedersen, N.C. Y Chromosome Haplotype Analysis in Purebred Dogs. Mamm. Genome 2005, 16, 273–280. [Google Scholar] [CrossRef]
- Lindgren, G.; Backström, N.; Swinburne, J.; Hellborg, L.; Einarsson, A.; Sandberg, K.; Cothran, G.; Vilà, C.; Binns, M.; Ellegren, H. Limited Number of Patrilines in Horse Domestication. Nat. Genet. 2004, 36, 335–336. [Google Scholar] [CrossRef]
- Ding, Z.-L.; Oskarsson, M.; Ardalan, A.; Angleby, H.; Dahlgren, L.-G.; Tepeli, C.; Kirkness, E.; Savolainen, P.; Zhang, Y.-P. Origins of Domestic Dog in Southern East Asia Is Supported by Analysis of Y-Chromosome DNA. Heredity 2012, 108, 507–514. [Google Scholar] [CrossRef]
- Natanaelsson, C.; Oskarsson, M.C.; Angleby, H.; Lundeberg, J.; Kirkness, E.; Savolainen, P. Dog Y Chromosomal DNA Sequence: Identification, Sequencing and SNP Discovery. BMC Genet. 2006, 7, 45. [Google Scholar] [CrossRef]
- Shannon, L.M.; Boyko, R.H.; Castelhano, M.; Corey, E.; Hayward, J.J.; McLean, C.; White, M.E.; Abi Said, M.; Anita, B.A.; Bondjengo, N.I.; et al. Genetic Structure in Village Dogs Reveals a Central Asian Domestication Origin. Proc. Natl. Acad. Sci. USA 2015, 112, 13639–13644. [Google Scholar] [CrossRef]
- Li, G.; Davis, B.W.; Raudsepp, T.; Pearks Wilkerson, A.J.; Mason, V.C.; Ferguson-Smith, M.; O’Brien, P.C.; Waters, P.D.; Murphy, W.J. Comparative Analysis of Mammalian Y Chromosomes Illuminates Ancestral Structure and Lineage-Specific Evolution. Genome Res. 2013, 23, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Oetjens, M.T.; Martin, A.; Veeramah, K.R.; Kidd, J.M. Analysis of the Canid Y-Chromosome Phylogeny Using Short-Read Sequencing Data Reveals the Presence of Distinct Haplogroups among Neolithic European Dogs. BMC Genom. 2018, 19, 350. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpré, M.B.; Sablin, M.V.; López-Giráldez, F.; Domingo-Roura, X.; et al. Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef]
- von Holdt, B.M.; Pollinger, J.P.; Earl, D.A.; Knowles, J.C.; Boyko, A.R.; Parker, H.; Geffen, E.; Pilot, M.; Jędrzejewski, W.; Jędrzejewska, B.; et al. A Genome-Wide Perspective on the Evolutionary History of Enigmatic Wolf-like Canids. Genome Res. 2011, 21, 1294–1305. [Google Scholar] [CrossRef]
- von Holdt, B.M.; Cahill, J.A.; Fan, Z.; Gronau, I.; Robinson, J.; Pollinger, J.P.; Shapiro, B.; Wall, J.; Wayne, R.K. Whole-Genome Sequence Analysis Shows That Two Endemic Species of North American Wolf Are Admixtures of the Coyote and Gray Wolf. Sci. Adv. 2016, 2, e1501714. [Google Scholar] [CrossRef]
- Marsden, C.D.; Ortega-Del Vecchyo, D.; O’Brien, D.P.; Taylor, J.F.; Ramirez, O.; Vilà, C.; Marques-Bonet, T.; Schnabel, R.D.; Wayne, R.K.; Lohmueller, K.E. Bottlenecks and Selective Sweeps during Domestication Have Increased Deleterious Genetic Variation in Dogs. Proc. Natl. Acad. Sci. USA 2016, 113, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Deguilloux, M.F.; Moquel, J.; Pemonge, M.H.; Colombeau, G. Ancient DNA Supports Lineage Replacement in European Dog Gene Pool: Insight into Neolithic Southeast France. J. Archaeol. Sci. 2009, 36, 513–519. [Google Scholar] [CrossRef]
- Verginelli, F.; Capelli, C.; Coia, V.; Musiani, M.; Falchetti, M.; Ottini, L.; Palmirotta, R.; Tagliacozzo, A.; De Grossi Mazzorin, I.; Mariani-Costantini, R. Mitochondrial DNA from Prehistoric Canids Highlights Relationships Between Dogs and South-East European Wolves. Mol. Biol. Evol. 2005, 22, 2541–2551. [Google Scholar] [CrossRef]
- Pires, A.E.; Detry, C.; Chikhi, L.; Rasteiro, R.; Amorim, I.R.; Simões, F.; Matos, J.; Petrucci-Fonseca, F.; Ollivier, M.; Hänni, C.; et al. The Curious Case of the Mesolithic Iberian Dogs: An Archaeogenetic Study. J. Archaeol. Sci. 2019, 105, 116–129. [Google Scholar] [CrossRef]
- Pang, J.-F.; Kluetsch, C.; Zou, X.-J.; Zhang, A.-b.; Luo, L.-Y.; Angleby, H.; Ardalan, A.; Ekstrom, C.; Skollermo, A.; Lundeberg, J.; et al. MtDNA Data Indicate a Single Origin for Dogs South of Yangtze River, Less Than 16,300 Years Ago, from Numerous Wolves. Mol. Biol. Evol. 2009, 26, 2849–2864. [Google Scholar] [CrossRef] [PubMed]
- Bergström, A.; Stanton, D.W.G.; Taron, U.H.; Frantz, L.; Sinding, M.-H.S.; Ersmark, E.; Pfrengle, S.; Cassatt-Johnstone, M.; Lebrasseur, O.; Girdland-Flink, L.; et al. Grey Wolf Genomic History Reveals a Dual Ancestry of Dogs. Nature 2022, 607, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, G.; Di Gaetano, C.; Guarrera, S.; Rosa, F.; Feldman, M.W.; Piazza, A.; Matullo, G. The Italian Genome Reflects the History of Europe and the Mediterranean Basin. Eur. J. Hum. Genet. 2016, 24, 1056–1062. [Google Scholar] [CrossRef]
- Raveane, A.; Aneli, S.; Montinaro, F.; Athanasiadis, G.; Barlera, S.; Birolo, G.; Boncoraglio, G.; Di Blasio, A.M.; Di Gaetano, C.; Pagani, L.; et al. Population Structure of Modern-Day Italians Reveals Patterns of Ancient and Archaic Ancestries in Southern Europe. Sci. Adv. 2019, 5, Eaaw3492. [Google Scholar] [CrossRef] [PubMed]
- Modi, A.; Lancioni, H.; Cardinali, I.; Capodiferro, M.R.; Rambaldi Migliore, N.; Hussein, A.; Strobl, C.; Bodner, M.; Schnaller, L.; Xavier, C.; et al. The Mitogenome Portrait of Umbria in Central Italy as Depicted by Contemporary Inhabitants and Pre-Roman Remains. Sci. Rep. 2020, 10, 10700. [Google Scholar] [CrossRef] [PubMed]
- Aneli, S.; Caldon, M.; Saupe, T.; Montinaro, F.; Pagani, L. Through 40,000 Years of Human Presence in Southern Europe: The Italian Case Study. Hum. Genet. 2021, 140, 1417–1431. [Google Scholar] [CrossRef] [PubMed]
- Ciucani, M.M.; Palumbo, D.; Galaverni, M.; Serventi, P.; Fabbri, E.; Ravegnini, G.; Angelini, S.; Maini, E.; Persico, D.; Caniglia, R.; et al. Old Wild Wolves: Ancient DNA Survey Unveils Population Dynamics in Late Pleistocene and Holocene Italian Remains. PeerJ 2019, 7, e6424. [Google Scholar] [CrossRef]
- Yankova, I.; Marinov, M.; Neov, B.; Petrova, M.; Spassov, N.; Hristov, P.; Radoslavov, G. Evidence for Early European Neolithic Dog Dispersal: New Data on Southeastern European Subfossil Dogs from the Prehistoric and Antiquity Ages. Genes 2019, 10, 757. [Google Scholar] [CrossRef]
- Pilot, M.; Branicki, W.; Jędrzejewski, W.; Goszczyński, J.; Jędrzejewska, B.; Dykyy, I.; Shkvyrya, M.; Tsingarska, E. Phylogeographic History of Grey Wolves in Europe. BMC Evol. Biol. 2010, 10, 104. [Google Scholar] [CrossRef]
- Carbajo Arana, M.; Fernandez Rodríguez, C. Los Carnívoros Del Recinto de Fosos de El Casetón de La Era (Villalba de Los Alcores, Valladolid). In Entre Ciencia e Cultura: Da Interdisciplinaridade a Transversalidade Da Arqueologia, Actas Das VIII Jornadas de Jovens Em Investigaçao Arqueologica, Celebradas En 2015 En La Universidade Nova de Lisboa; Pinto Coelho, I., Bento Torres, J., Serrãno Gil, L., Ramos, T., Eds.; Centro de Estudos de Patrim Onio, Departamento de Historia, Arqueologia e Patrimonio, Universidade Do Algarve: Faro, Portugal, 2016; pp. 277–282. [Google Scholar]
- Pires, A.E.; Detry, C.; Fernandez-Rodriguez, C.; Valenzuela-Lamas, S.; Arruda, A.M.; De Grossi Mazzorin, J.; Ollivier, M.; Hänni, C.; Simões, F.; Ginja, C. Roman Dogs from the Iberian Peninsula and the Maghreb—A Glimpse into Their Morphology and Genetics. Quat. Int. 2018, 471, 132–146. [Google Scholar] [CrossRef]
- Boschin, F.; Bernardini, F.; Pilli, E.; Vai, S.; Zanolli, C.; Tagliacozzo, A.; Fico, R.; Fedi, M.; Corny, J.; Dreossi, D.; et al. The First Evidence for Late Pleistocene Dogs in Italy. Sci. Rep. 2020, 10, 13313. [Google Scholar] [CrossRef] [PubMed]
- Perini, F.; Cardinali, I.; Ceccobelli, S.; Gruppetta, A.; José, C.S.; Cosenza, M.; Musso, N.; Martìnez, A.; Abushady, A.M.; Monteagudo, L.V.; et al. Phylogeographic and Population Genetic Structure of Hound-like Native Dogs of the Mediterranean Basin. Res. Vet. Sci. 2023, 155, 103–114. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tancredi, D.; Cardinali, I. Being a Dog: A Review of the Domestication Process. Genes 2023, 14, 992. https://doi.org/10.3390/genes14050992
Tancredi D, Cardinali I. Being a Dog: A Review of the Domestication Process. Genes. 2023; 14(5):992. https://doi.org/10.3390/genes14050992
Chicago/Turabian StyleTancredi, Domenico, and Irene Cardinali. 2023. "Being a Dog: A Review of the Domestication Process" Genes 14, no. 5: 992. https://doi.org/10.3390/genes14050992
APA StyleTancredi, D., & Cardinali, I. (2023). Being a Dog: A Review of the Domestication Process. Genes, 14(5), 992. https://doi.org/10.3390/genes14050992