
Effects of Exogenous (K+) Potassium Application on Plant Hormones in the Roots of Tamarix ramosissima under NaCl Stress
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Treatment of Experimental Seedlings
2.3. Transcriptome Sequencing and Differentially Expressed Gene Screening
2.4. Quantitative Real-Time PCR (qRT-PCR) Validation
2.5. Experiment Data Processing
3. Results
3.1. Screening of Differentially Expressed Genes (DEGs)
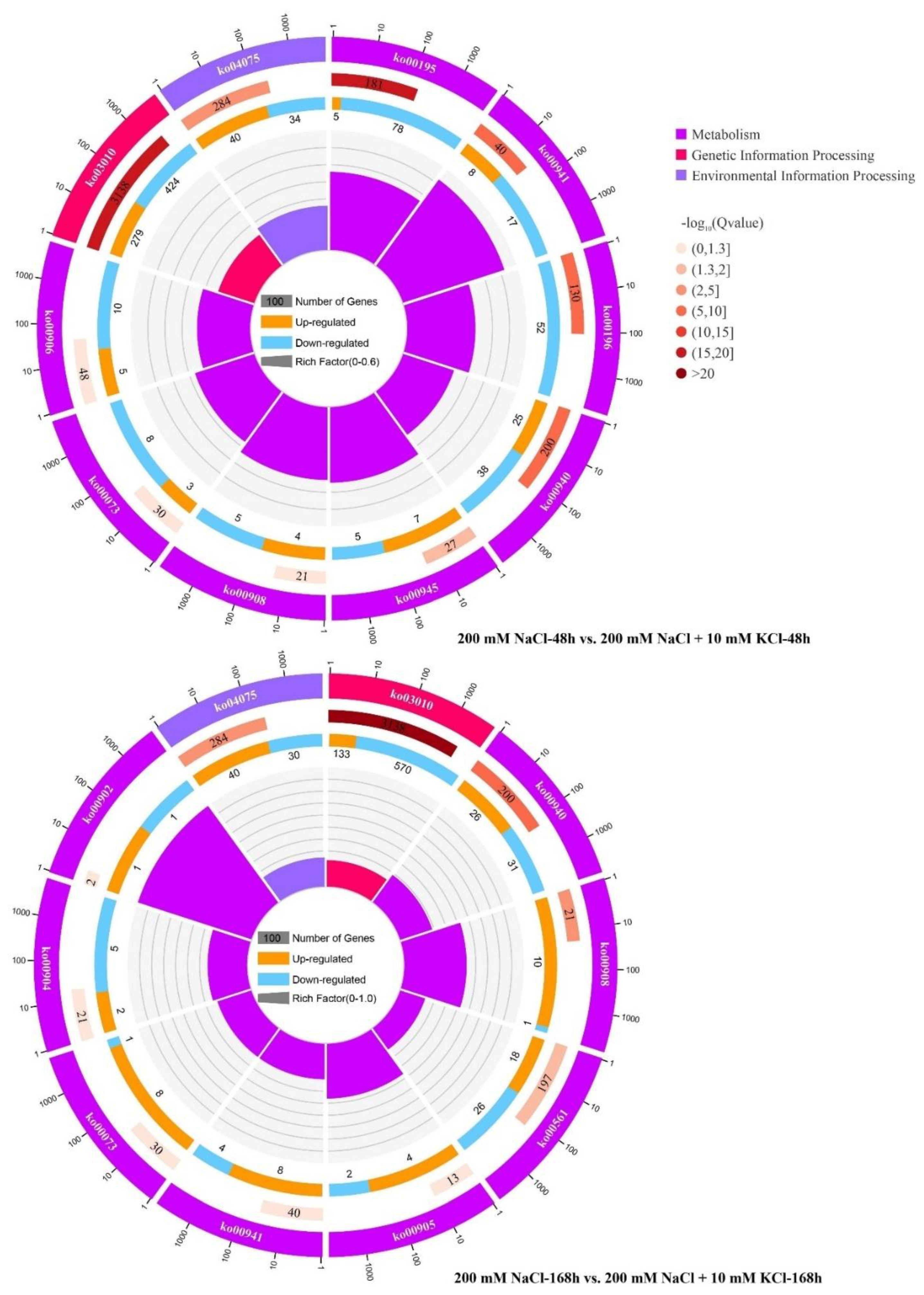
3.2. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
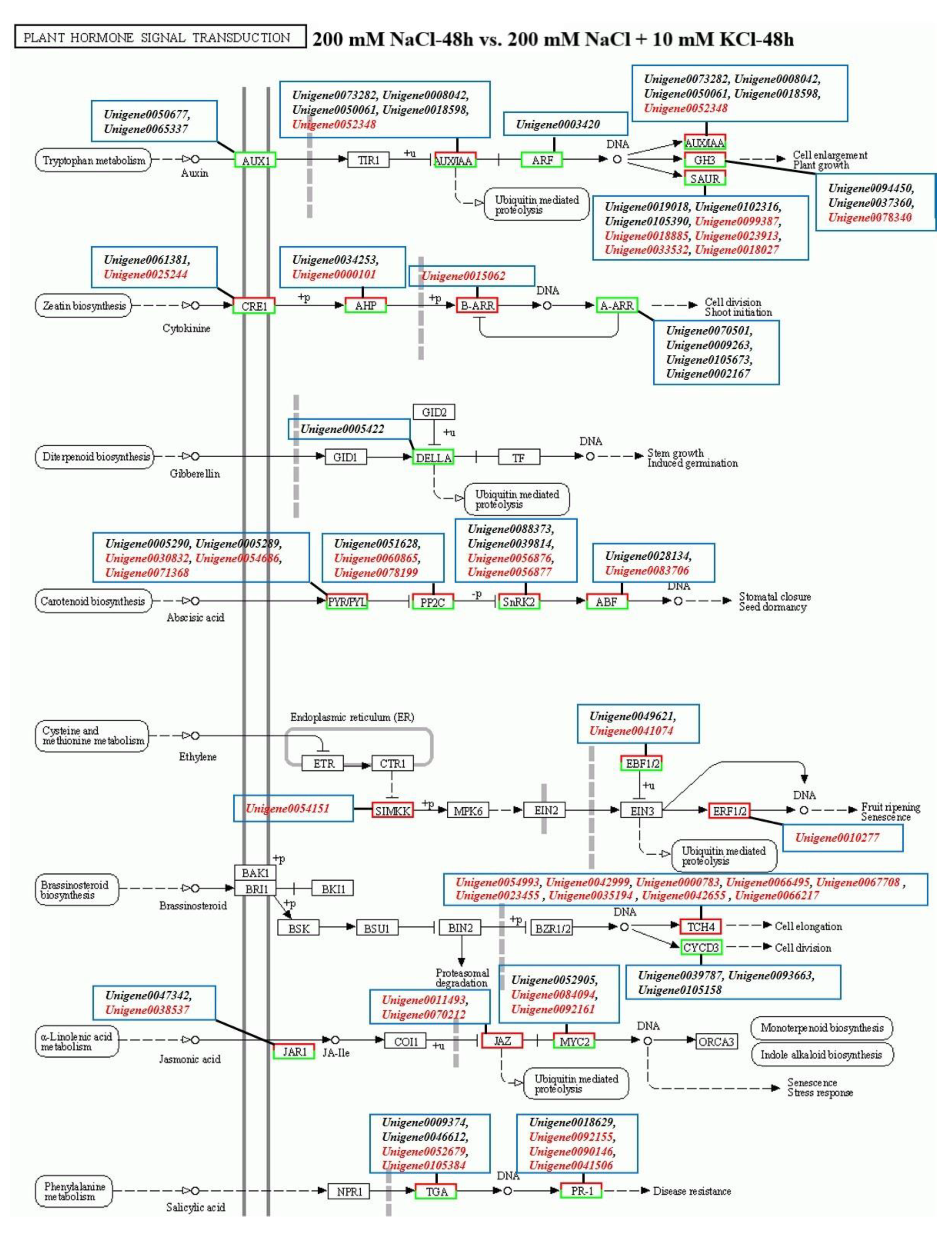
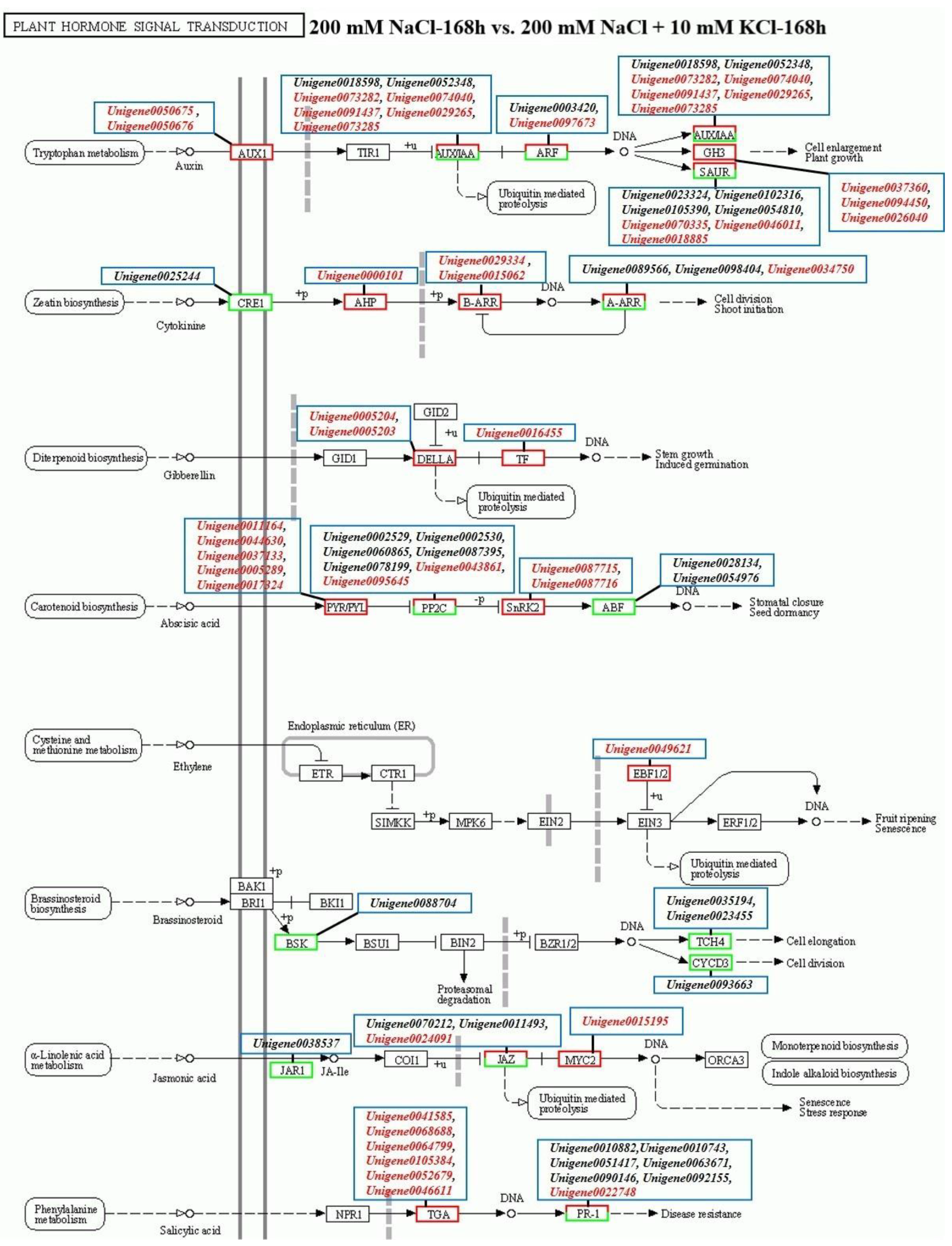
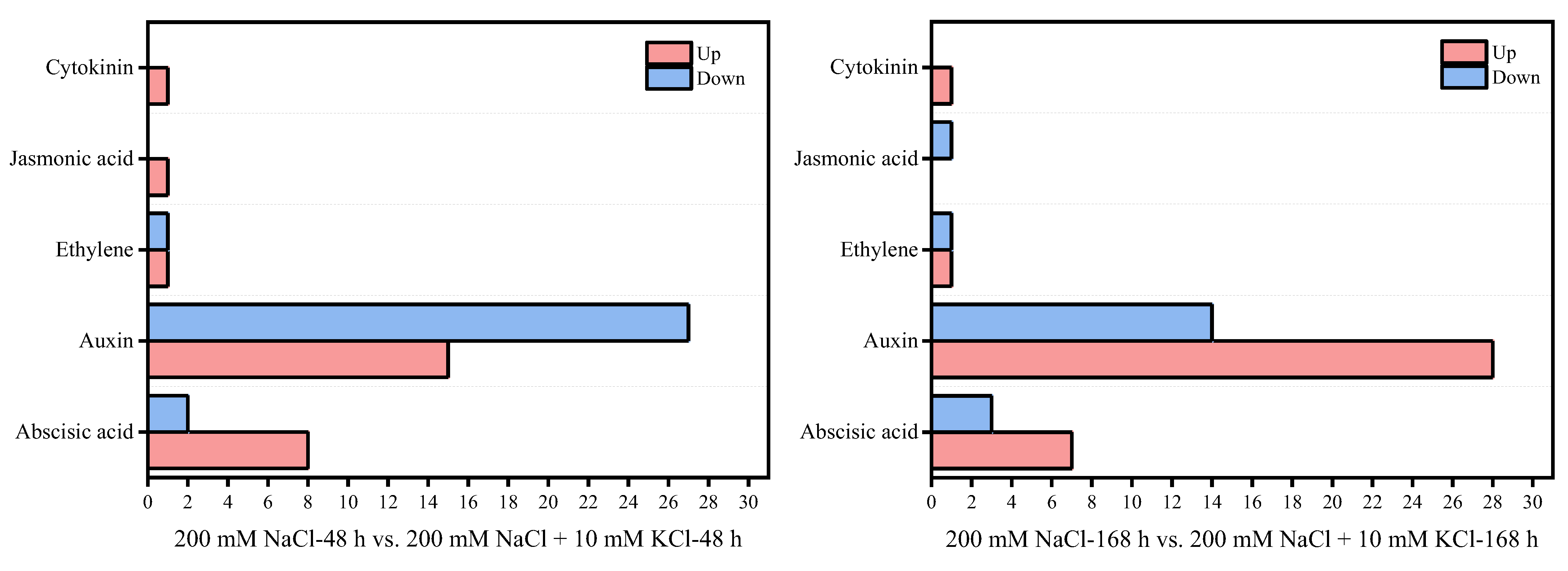
3.3. Plant Hormone Signal Transduction Pathway Analysis
3.4. Analysis of Candidate Genes Related to Plant Hormones in the Roots of T. ramosissima by Exogenous Potassium Application under NaCl Stress
3.5. Phylogenetic Tree Analysis of Auxin Key Candidate Genes in Plant Hormones
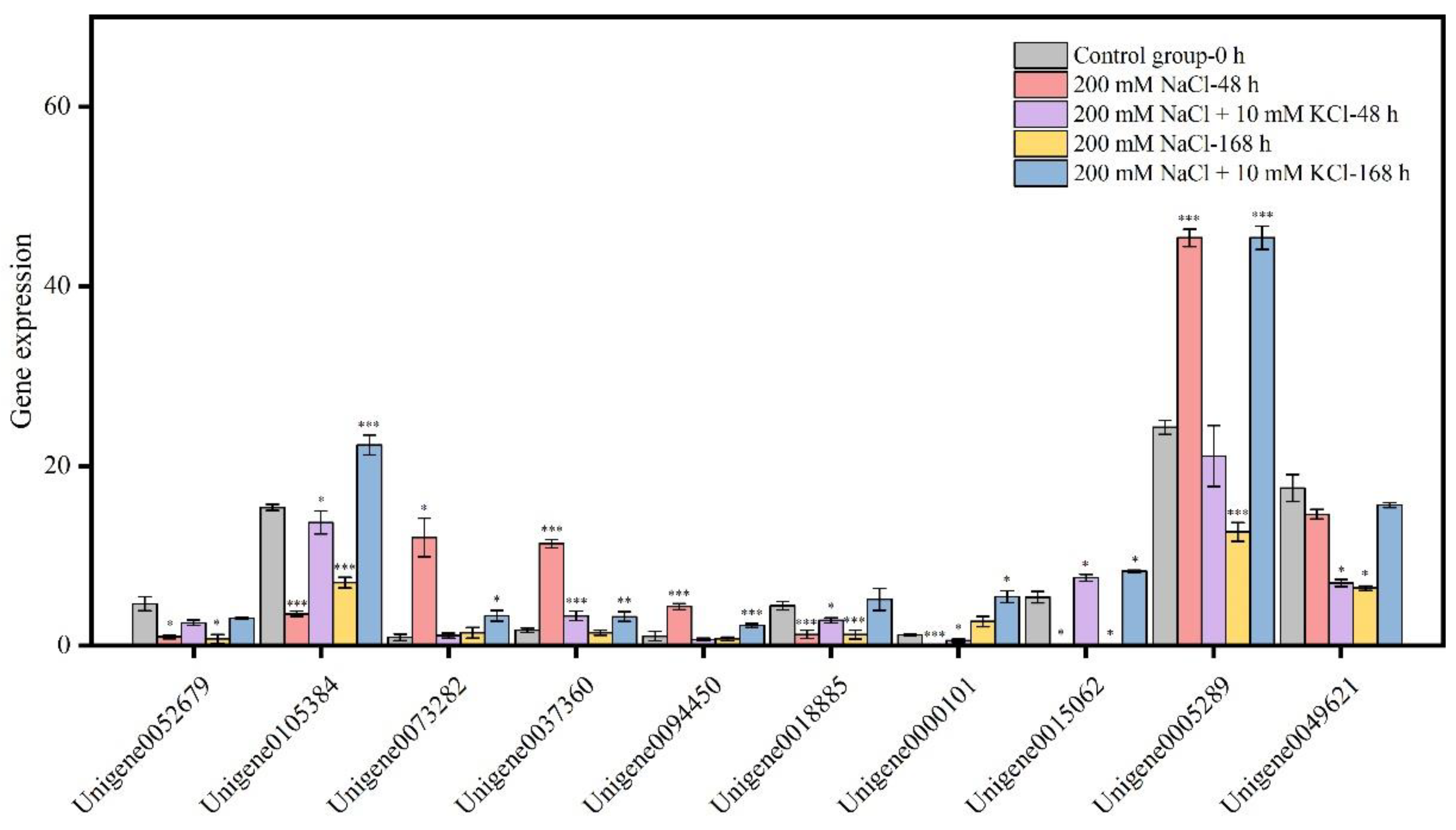
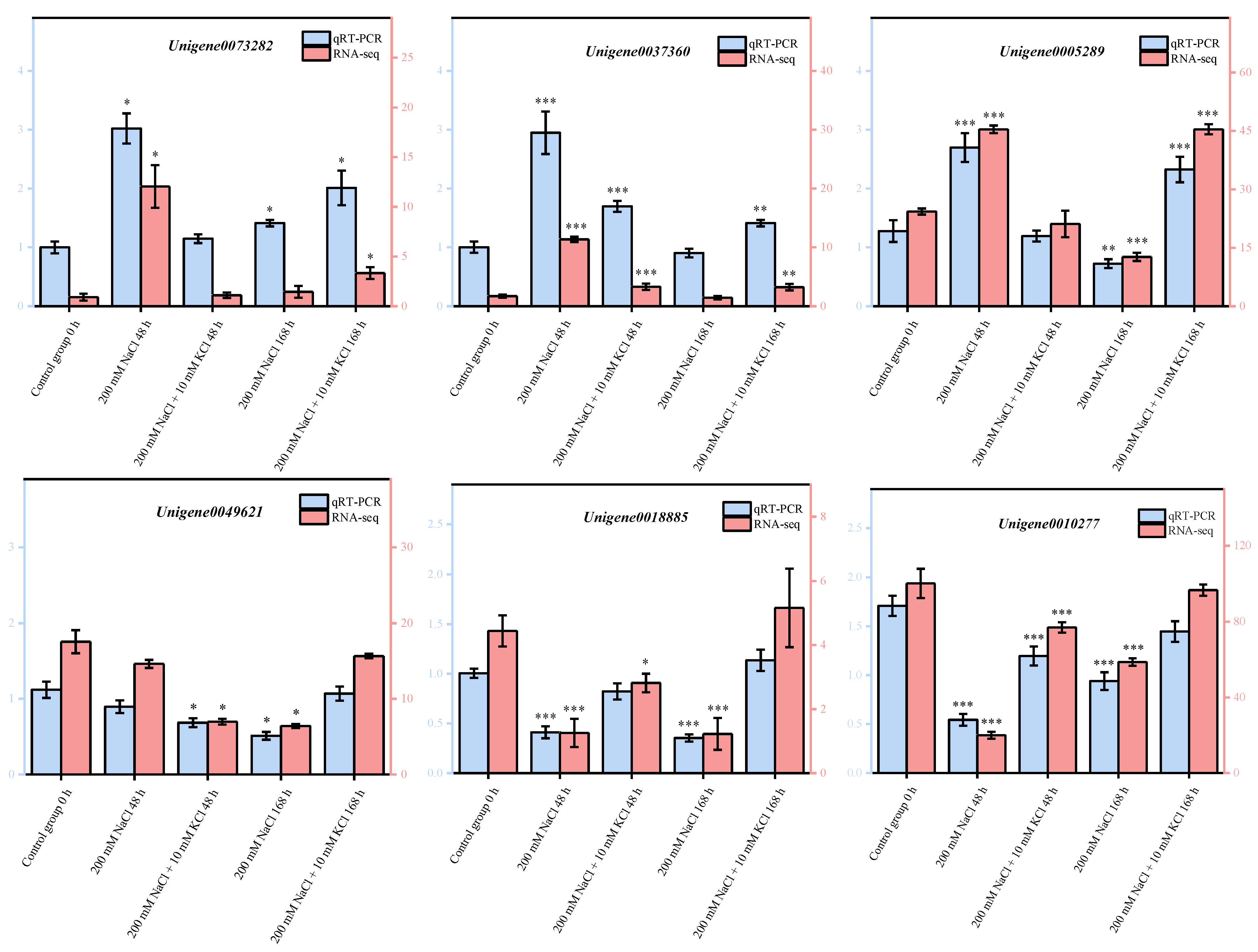
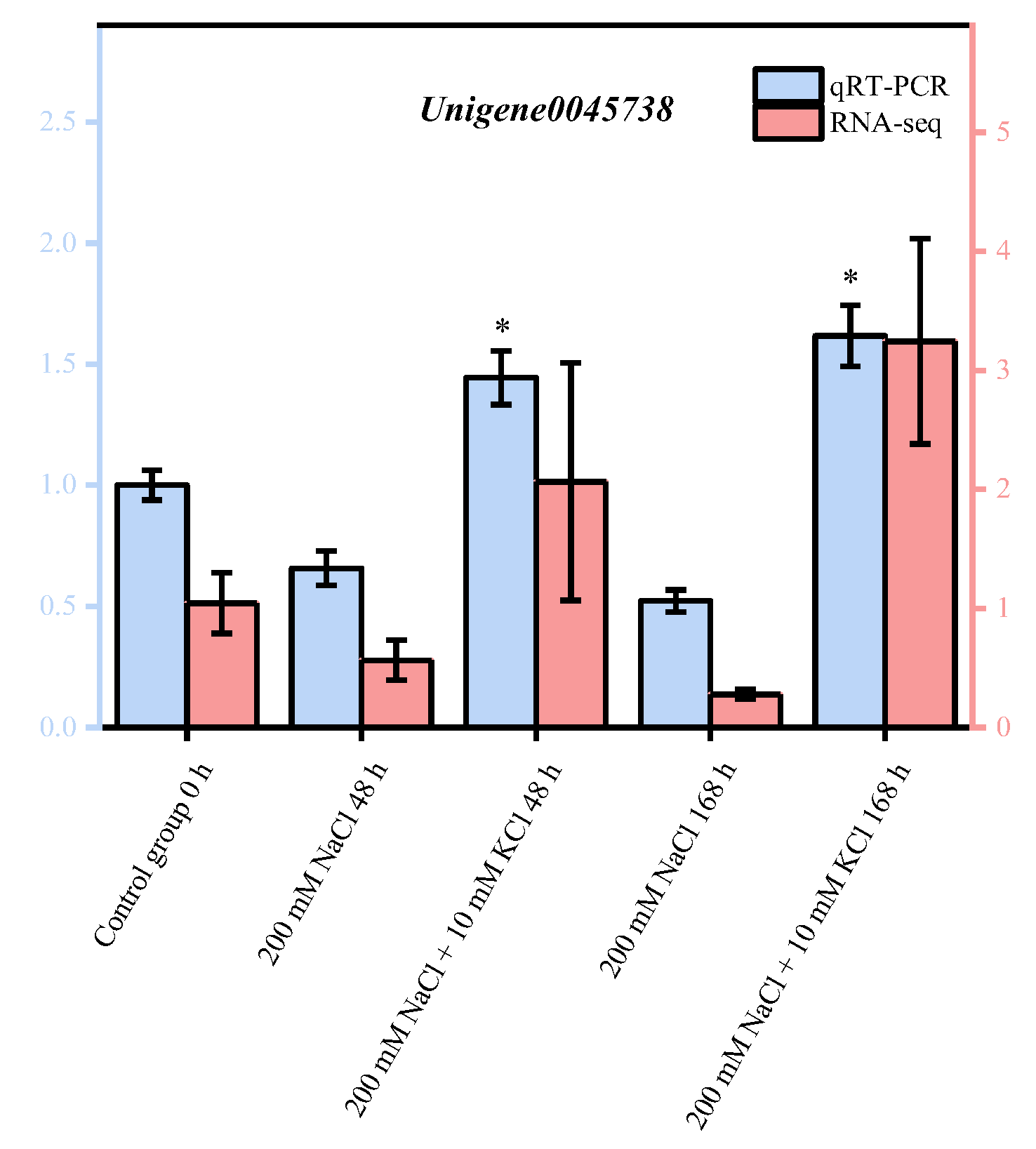
3.6. Quantitative Real-Time PCR (qRT-PCR) Validation of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Lubna; Khan, M.A.; Asaf, S.; Jan, R.; Waqas, M.; Kim, K.; Lee, I. Endophytic fungus Bipolaris sp. CSL-1 induces salt tolerance in Glycine max. L via modulating its endogenous hormones, antioxidative system and gene expression. J. Plant Interact. 2022, 17, 319–332. [Google Scholar] [CrossRef]
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity Stress in Arid and Semi-Arid Climates: Effects and Management in Field Crops. Clim. Change Agric. 2019, 13, 1–26. [Google Scholar] [CrossRef]
- Dhiman, P.; Rajora, N.; Bhardwaj, S.; Sudhakaran, S.S.; Kumar, A.; Raturi, G.; Chakraborty, K.; Gupta, O.P.; Devanna, B.N.; Tripathi, D.K.; et al. Fascinating role of silicon to combat salinity stress in plants: An updated overview. Plant Physiol. Biochem. 2021, 162, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Cimini, S.; Locato, V.; Giacinti, V.; Molinari, M.; De Gara, L. A Multifactorial Regulation of Glutathione Metabolism behind Salt Tolerance in Rice. Antioxidants 2022, 11, 1114. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, T.U.D.L.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. Vitr. Cell. Dev. Biol. Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Islam, M.S.; Hossain, A.; Iqbal, M.A.; Mubeen, M.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as Growth Regulators During Abiotic Stress Tolerance in Plants. Front. Agron. 2022, 4, 765068. [Google Scholar] [CrossRef]
- Asami, T.; Nakagawa, Y. Preface to the Special Issue: Brief review of plant hormones and their utilization in agriculture. J. Pestic. Sci. 2018, 43, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.H.; Islam, S.; Parrey, Z.A.; Mohammad, F. Role of Exogenously Applied Plant Growth Regulators in Growth and Development of Edible Oilseed Crops Under Variable Environmental Conditions: A Review. J. Soil Sci. Plant Nut. 2021, 21, 3284–3308. [Google Scholar] [CrossRef]
- Wang, L.; Zou, Y.; Kaw, H.Y.; Wang, G.; Sun, H.; Cai, L.; Li, C.; Meng, L.Y.; Li, D. Recent developments and emerging trends of mass spectrometric methods in plant hormone analysis: A review. Plant Methods 2020, 16, 54. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, Y. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Yan, Z.; Li, X.; Chen, J.; Tam, N.F. Combined toxicity of cadmium and copper in Avicennia marina seedlings and the regulation of exogenous jasmonic acid. Ecotoxicol. Environ. Saf. 2015, 113, 124–132. [Google Scholar] [CrossRef]
- Ondrasek, G.; Rathod, S.; Manohara, K.K.; Gireesh, C.; Anantha, M.S.; Sakhare, A.S.; Parmar, B.; Yadav, B.K.; Bandumula, N.; Raihan, F.; et al. Salt Stress in Plants and Mitigation Approaches. Plants 2022, 11, 717. [Google Scholar] [CrossRef]
- Muller, M.; Munne-Bosch, S. Hormonal impact on photosynthesis and photoprotection in plants. Plant Physiol. 2021, 185, 1500–1522. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Parveen, A.; Ahmar, S.; Kamran, M.; Malik, Z.; Ali, A.; Riaz, M.; Abbasi, G.H.; Khan, M.; Sohail, A.B.; Rizwan, M.; et al. Abscisic acid signaling reduced transpiration flow, regulated Na+ ion homeostasis and antioxidant enzyme activities to induce salinity tolerance in wheat (Triticum aestivum L.) seedlings. Environ. Technol. Innov. 2021, 24, 101808. [Google Scholar] [CrossRef]
- Sripinyowanich, S.; Klomsakul, P.; Boonburapong, B.; Bangyeekhun, T.; Asami, T.; Gu, H.; Buaboocha, T.; Chadchawan, S. Exogenous ABA induces salt tolerance in indica rice (Oryza sativa L.): The role of OsP5CS1 and OsP5CR gene expression during salt stress: Phytohormones and Stress. Environ. Exp. Bot. 2013, 86, 94–105. [Google Scholar] [CrossRef]
- Khalid, A.; Aftab, F. Effect of exogenous application of IAA and GA3 on growth, protein content, and antioxidant enzymes of Solanum tuberosum L. grown in vitro under salt stress. Vitr. Cell. Dev. Biol. Plant 2020, 56, 377–389. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Tahjib-Ul-Arif, M.; Rhaman, M.S. Exogenous Auxin-Mediated Salt Stress Alleviation in Faba Bean (Vicia faba L.). Agronomy 2021, 11, 547. [Google Scholar] [CrossRef]
- Guo, Q.; Major, I.T.; Howe, G.A. Resolution of growth-defense conflict: Mechanistic insights from jasmonate signaling. Curr. Opin. Plant Biol. 2018, 44, 72–81. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Hamayun, M.; Lee, S.; Lee, I. Methyl jasmonate alleviated salinity stress in soybean. J. Crop. Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, W.; Chan, Z.; Wu, Y. Endogenous Cytokinin Overproduction Modulates ROS Homeostasis and Decreases Salt Stress Resistance in Arabidopsis Thaliana. Front. Plant Sci. 2015, 6, 1004. [Google Scholar] [CrossRef]
- Abdullah, Z.; Ahmad, R. Effect of Pre- and Post-Kinetin Treatments on Salt Tolerance of Different Potato Cultivars Growing on Saline Soils. J. Agron. Crop Sci. 1990, 165, 94–102. [Google Scholar] [CrossRef]
- Kirkham, M.B.; Gardner, W.R.; Gerloff, G.C. Internal Water Status of Kinetin-treated, Salt-stressed Plants. Plant Physiol. 1974, 53, 241–243. [Google Scholar] [CrossRef]
- Barciszewski, J.; Siboska, G.; Rattan, S.I.S.; Clark, B.F.C. Occurrence, biosynthesis and properties of kinetin (N6-furfuryladenine). Plant Growth Regul. 2000, 32, 257–265. [Google Scholar] [CrossRef]
- Yu, Y.; Li, Y.; Yan, Z.; Duan, X. The Role of Cytokinins in Plant Under Salt Stress. J. Plant Growth Regul. 2021, 41, 2279–2291. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Meng, Z.; Wang, B.; Chen, M. Research Progress on the Roles of Cytokinin in Plant Response to Stress. Int. J. Mol. Sci. 2020, 21, 6574. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Belfield, E.J.; Cao, Y.; Smith, J.A.; Harberd, N.P. An Arabidopsis soil-salinity-tolerance mutation confers ethylene-mediated enhancement of sodium/potassium homeostasis. Plant Cell 2013, 25, 3535–3552. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Schroeder, J.I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants. Nature 1994, 370, 655–658. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef]
- 38. Chen, G.; Hu, Q.; Luo, L.E.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef]
- Song, Z.Z.; Yang, S.; Zhu, H.; Jin, M.; Su, Y.H. Heterologous expression of an alligatorweed high-affinity potassium transporter gene enhances salinity tolerance in Arabidopsis thaliana. Am. J. Bot. 2014, 101, 840–850. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Du, S.; Jiang, J.; Wang, G. Transcriptome and Metabonomic Analysis of Tamarix ramosissima Potassium (K+) Channels and Transporters in Response to NaCl Stress. Genes 2022, 13, 1313. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Bahramsoltani, R.; Kalkhorani, M.; Abbas, Z.S.; Farzaei, M.H.; Rahimi, R. The genus Tamarix: Traditional uses, phytochemistry, and pharmacology. J. Ethnopharmacol. 2020, 246, 112245. [Google Scholar] [CrossRef] [PubMed]
- Thomson, W.W.; Liu, L.L. Ultrastructural features of the salt gland of Tamarix aphylla L. Planta 1967, 73, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yan, X.; Yang, Z.; Han, G.; Wang, L.; Yuan, F.; Wang, B. Salt glands of recretohalophyte Tamarix under salinity: Their evolution and adaptation. Ecol. Evol. 2020, 10, 9384–9395. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Wang, Y.; Jiang, B.; Liu, G.; Yu, L.; Wei, Z.; Yang, C. A novel vacuolar membrane H+-ATPase c subunit gene (ThVHAc1) from Tamarix hispida confers tolerance to several abiotic stresses in Saccharomyces cerevisiae. Mol. Biol. Rep. 2011, 38, 957–963. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Che, B.; Cheng, C.; Fang, J.; Liu, Y.; Jiang, L.; Yu, B. The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1. Int. J. Mol. Sci. 2019, 20, 2930. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Zhu, Z.; Wang, B.; Chen, M. Recent Progress on the Salt Tolerance Mechanisms and Application of Tamarisk. Int. J. Mol. Sci. 2022, 23, 3325. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, G.; Zhang, H.; Zhang, N.; Jiang, J.; Song, Z. Transcriptome analysis of Tamarix ramosissima leaves in response to NaCl stress. PLoS ONE 2022, 17, e265653. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef]
- Lu, Y.; Lei, J.Q.; Zeng, F.J.; Xu, L.S.; Peng, S.L.; Gao, H.H.; Liu, G.J. Effects of NaCl treatment on growth and Ec-ophysiology Characteristics of Tamarix ramossisma. J. Des. Res. 2014, 34, 1509–1515. [Google Scholar] [CrossRef]
- Garthwaite, A.J.; von Bothmer, R.; Colmer, T.D. Salt tolerance in wild Hordeum species is associated with restricted entry of Na+ and Cl− into the shoots. J. Exp. Bot. 2005, 56, 2365–2378. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Du, S.; Zhang, X.; Wang, G.; Huang, J.; Jiang, J. Effects of Exogenous Potassium (K+) Application on the Antioxidant Enzymes Activities in Leaves of Tamarix ramosissima under NaCl Stress. Genes 2022, 13, 1507. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Du, S.; Zhang, X.; Jiang, J.; Wang, G. Analysis of Amino Acids in the Roots of Tamarix ramosissima by Application of Exogenous Potassium (K+) under NaCl Stress. Int. J. Mol. Sci. 2022, 23, 9331. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani Javid, M.; Sorooshzadeh, A.; Modarres Sanavy, S.A.M.; Allahdadi, I.; Moradi, F. Effects of the exogenous application of auxin and cytokinin on carbohydrate accumulation in grains of rice under salt stress. Plant Growth Regul. 2011, 65, 305–313. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, D.; Feng, N.; Zhou, H.; Mu, D.; Zhao, L.; Shen, X.; Rao, G.; Meng, F.; Huang, A. Physiological mechanisms of ABA-induced salinity tolerance in leaves and roots of rice. Sci. Rep. 2022, 12, 8228. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Jin, Y.; Wang, M.; Yang, H.; Ma, H.; Jiang, C.; Liang, Z. Abscisic acid primes rice seedlings for enhanced tolerance to alkaline stress by upregulating antioxidant defense and stress tolerance-related genes. Plant Soil 2019, 438, 39–55. [Google Scholar] [CrossRef]
- Ribba, T.; Garrido-Vargas, F.; O’Brien, J.A. Auxin-mediated responses under salt stress: From developmental regulation to biotechnological applications. J. Exp. Bot. 2020, 71, 3843–3853. [Google Scholar] [CrossRef]
- Guo, Y.; Jiang, Q.; Hu, Z.; Sun, X.; Fan, S.; Zhang, H. Function of the auxin-responsive gene TaSAUR75 under salt and drought stress. Crop J. 2018, 6, 181–190. [Google Scholar] [CrossRef]
- Jain, M.; Khurana, J.P. Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 2009, 276, 3148–3162. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A.L. Alleviation of salt stress-induced adverse effects on maize plants by exogenous application of indoleacetic acid (IAA) and inorganic nutrients-A field trial. Aust. J. Crop Sci. 2013, 7, 249–254. [Google Scholar]
- Campos, M.L.; Kang, J.H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, D.; Wang, Y.; Xie, D. Jasmonate action in plant defense against insects. J. Exp. Bot. 2019, 70, 3391–3400. [Google Scholar] [CrossRef]
- Noor, J.; Ullah, A.; Saleem, M.H.; Tariq, A.; Ullah, S.; Waheed, A.; Okla, M.K.; Al-Hashimi, A.; Chen, Y.; Ahmed, Z.; et al. Effect of Jasmonic Acid Foliar Spray on the Morpho-Physiological Mechanism of Salt Stress Tolerance in Two Soybean Varieties (Glycine max L.). Plants 2022, 11, 651. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.J.; Seo, Y.J.; Lee, J.D.; Ishii, R.; Kim, U.; Shin, D.H.; Park, S.K.; Jang, S.W.; Lee, I.J. Jasmonic acid differentially affects growth, ion uptake and abscisic acid concentration in salt-tolerant and salt-sensitive rice cultivars. J. Agron. Crop Sci. 2005, 191, 273–282. [Google Scholar] [CrossRef]
- Ali, M.S.; Baek, K. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, H.; Ma, B.; Zhang, W.; Chen, S.; Zhang, J. The Role of Ethylene in Plants Under Salinity Stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef]
- Cao, W.H.; Liu, J.; He, X.J.; Mu, R.L.; Zhou, H.L.; Chen, S.Y.; Zhang, J.S. Modulation of ethylene responses affects plant salt-stress responses. Plant Physiol. 2007, 143, 707–719. [Google Scholar] [CrossRef]
- Peng, J.; Li, Z.; Wen, X.; Li, W.; Shi, H.; Yang, L.; Zhu, H.; Guo, H. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. PLoS Genet. 2014, 10, e1004664. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.; Gong, W.; Sun, J.; Pan, Z.; Xu, F.; Lu, Y.; Du, X. Comprehensive analysis of differentially expressed genes and transcriptional regulation induced by salt stress in two contrasting cotton genotypes. BMC Genom. 2014, 15, 760. [Google Scholar] [CrossRef]
- Nishiyama, R.; Watanabe, Y.; Fujita, Y.; Le, D.T.; Kojima, M.; Werner, T.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Kakimoto, T.; et al. Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 2011, 23, 2169–2183. [Google Scholar] [CrossRef]
- Tran, L.S.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef]
- Ha, S.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Cytokinins: Metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci. 2012, 17, 172–179. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Gaudinova, A.; Hosek, P.; Soudek, P.; Knirsch, V.; Vankova, R. Hormonal dynamics during salt stress responses of salt-sensitive Arabidopsis thaliana and salt-tolerant Thellungiella salsuginea. Plant Sci. 2017, 264, 188–198. [Google Scholar] [CrossRef]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflorescence meristem-specific cytokinin oxidase—OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Qin, L.; Wang, L.; Guo, Y.; Li, Y.; Umut, H.; Wang, Y. An ERF transcription factor from Tamarix hispida, ThCRF1, can adjust osmotic potential and reactive oxygen species scavenging capability to improve salt tolerance. Plant Sci. 2017, 265, 154–166. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Pathway | Pathway Annotated Genes | p-Value | Pathway ID |
|---|---|---|---|---|
| 200 mM NaCl-48 h vs. 200 mM NaCl + 10 mM KCl-48 h | ||||
| 1 | Photosynthesis | 83 | 0.000000 | ko00195 |
| 2 | Ribosome | 703 | 0.000000 | ko03010 |
| 3 | Flavonoid biosynthesis | 25 | 0.000000 | ko00941 |
| 4 | Photosynthesis—antenna proteins | 52 | 0.000000 | ko00196 |
| 5 | Phenylpropanoid biosynthesis | 63 | 0.000000 | ko00940 |
| 6 | Plant hormone signal transduction | 74 | 0.000079 | ko04075 |
| 7 | Stilbenoid, diarylheptanoid and gingerol biosynthesis | 12 | 0.000824 | ko00945 |
| 8 | Zeatin biosynthesis | 9 | 0.004980 | ko00908 |
| 9 | Cutin, suberine and wax biosynthesis | 11 | 0.008045 | ko00073 |
| 10 | Carotenoid biosynthesis | 15 | 0.011468 | ko00906 |
| 200 mM NaCl-168 h vs. 200 mM NaCl + 10 mM KCl-168 h | ||||
| 1 | Ribosome | 703 | 0.000000 | ko03010 |
| 2 | Phenylpropanoid biosynthesis | 57 | 0.000000 | ko00940 |
| 3 | Plant hormone signal transduction | 70 | 0.000002 | ko04075 |
| 4 | Zeatin biosynthesis | 11 | 0.000042 | ko00908 |
| 5 | Glycerolipid metabolism | 44 | 0.001344 | ko00561 |
| 6 | Brassinosteroid biosynthesis | 6 | 0.005758 | ko00905 |
| 7 | Flavonoid biosynthesis | 12 | 0.007853 | ko00941 |
| 8 | Cutin, suberine and wax biosynthesis | 9 | 0.020172 | ko00073 |
| 9 | Monoterpenoid biosynthesis | 2 | 0.020236 | ko00902 |
| 10 | Diterpenoid biosynthesis | 7 | 0.021957 | ko00904 |
| Pathway | Gene ID | Description | Log2 Fold-Change | |
|---|---|---|---|---|
| 200 mM NaCl-48 h vs. 200 mM NaCl + 10 mM KCl-48 h | 200 mM NaCl-168 h vs. 200 mM NaCl + 10 mM KCl-168 h | |||
| Abscisic Acid | ||||
| ko00906 | Unigene0079211 | Abscisic acid 8′-hydroxylase 2-like | 0.44 | 4.17 |
| Unigene0063711 | Abscisic acid 8′-hydroxylase | 4.62 | 1.50 | |
| Unigene0008844 | Abscisic acid 8′-hydroxylase 2-like | 0.58 | 2.74 | |
| Unigene0027741 | Abscisic acid 8′-hydroxylase CYP707A2 isoform X1 | 1.70 | −0.01 | |
| ko04016; ko04075 | Unigene0080236 | Abscisic acid receptor PYL2-like | 0.05 | 7.85 |
| Unigene0039063 | Abscisic acid receptor PYR1-like | −0.49 | 0.28 | |
| Unigene0044630 | Abscisic acid receptor PYL4-like | 0.44 | 1.14 | |
| Unigene0054686 | Abscisic acid receptor PYR1-like | 2.01 | −0.97 | |
| Unigene0071368 | Abscisic acid receptor PYL4 | 2.09 | 0.03 | |
| ko04075 | Unigene0028134 | Abscisic acid-insensitive 5-like protein 2 | −1.79 | −1.33 |
| Auxin | ||||
| ko04075 | Unigene0000292 | Auxin-responsive protein IAA14-like | −0.25 | 0.06 |
| Unigene0003420 | Auxin response factor 3-like | −1.73 | −1.08 | |
| Unigene0008042 | Auxin-responsive protein IAA14 | −1.51 | 0.50 | |
| Unigene0009423 | Auxin | −0.76 | 0.17 | |
| Unigene0011116 | Auxin-responsive protein SAUR32-like | −0.13 | −0.07 | |
| Unigene0018027 | Auxin-responsive protein SAUR21 | 10.01 | 8.49 | |
| Unigene0018598 | Auxin-induced protein AUX22-like | −1.04 | −2.11 | |
| Unigene0018885 | Auxin-responsive protein SAUR36 | 1.16 | 2.07 | |
| Unigene0019018 | Auxin-responsive protein SAUR50-like | −3.55 | −0.04 | |
| Unigene0029265 | Auxin-induced protein 22D-like | −0.35 | 2.06 | |
| Unigene0030611 | Auxin-responsive protein SAUR50 | 7.32 | −9.42 | |
| Unigene0031005 | Auxin response factor 1-like | −0.98 | −0.12 | |
| Unigene0038393 | Auxin-responsive protein SAUR72 | 0.34 | 0.69 | |
| Unigene0046011 | Auxin-responsive protein SAUR32 | −0.01 | 1.59 | |
| Unigene0047953 | Auxin-responsive protein SAUR32 | −0.78 | 0.27 | |
| Unigene0048320 | Auxin response factor 9-like | −0.07 | 0.37 | |
| Unigene0048945 | Auxin-induced protein 15A-like | −2.51 | 0.37 | |
| Unigene0050061 | Auxin-responsive protein IAA9 | −1.17 | −0.97 | |
| Unigene0050675 | Auxin transporter-like protein 3 | −0.20 | 3.19 | |
| Unigene0050676 | Auxin transporter-like protein 3 | 0.23 | 3.33 | |
| Unigene0050677 | Auxin transporter-like protein 5 isoform X1 | −2.08 | 0.11 | |
| Unigene0052348 | Auxin-responsive protein IAA29-like | 1.39 | −1.38 | |
| Unigene0054810 | Auxin-responsive protein SAUR50 | 0.83 | −1.01 | |
| Unigene0059004 | Auxin-induced protein 22B | −0.13 | 0.23 | |
| Unigene0059322 | Auxin-responsive protein SAUR50 | −0.08 | 0.70 | |
| Unigene0059399 | Auxin-induced protein 6B-like | 0.41 | 0.59 | |
| Unigene0065338 | Auxin transporter-like protein 4 | 0.14 | −0.09 | |
| Unigene0065339 | Auxin transporter-like protein 2 | −0.92 | 0.26 | |
| Unigene0070335 | Auxin-responsive protein SAUR71-like | 0.71 | 1.27 | |
| Unigene0071678 | Auxin-responsive protein IAA26 | 0.29 | 0.78 | |
| Unigene0072291 | Auxin-responsive protein IAA26-like | 0.40 | 0.40 | |
| Unigene0073282 | Auxin-responsive protein IAA27-like protein | −3.44 | 1.20 | |
| Unigene0073285 | Auxin-responsive protein | 0.51 | 9.06 | |
| Unigene0074040 | Auxin-responsive protein IAA31 | −0.43 | 1.44 | |
| Unigene0077555 | Auxin-induced protein 6B-like | 0.14 | 0.74 | |
| Unigene0085809 | Auxin-responsive protein SAUR71-like | 0.98 | −0.61 | |
| Unigene0088743 | Auxin response factor 3-like | −0.22 | −0.51 | |
| Unigene0089917 | Auxin-responsive protein IAA8-like isoform X2 | −0.23 | 0.55 | |
| Unigene0091437 | Auxin-responsive protein IAA7 | −0.30 | 1.68 | |
| Unigene0097673 | Auxin response factor 11 | −0.42 | 1.35 | |
| Unigene0102316 | Auxin-induced protein 15A-like | −2.88 | −1.47 | |
| Unigene0105390 | Auxin-responsive protein SAUR71-like | −2.70 | −1.07 | |
| Ethylene | ||||
| ko04016; ko04075; | Unigene0010277 | Ethylene response factor 11 | 1.94 | 0.72 |
| ko04626 | Unigene0080149 | Ethylene response factor 6 | −0.47 | −0.47 |
| Jasmonic acid | ||||
| ko04075 | Unigene0038537 | Jasmonic acid-amido synthetase JAR1 | 1.82 | −1.54 |
| Cytokinin | ||||
| ko00908 | Unigene0045738 | Cytokinin oxidase/dehydrogenase | 1.87 | 3.55 |
| Family | Species | Description | Protein ID | CDS (bp) | ORF Length (aa) |
|---|---|---|---|---|---|
| Euphorbiaceae | Ricinus communis | Auxin-responsive protein IAA14 | XP_002517023.3 | 711 | 236 |
| Actinidiaceae | Actinidia chinensis var. chinensis | Auxin-responsive protein | PSS33813.1 | 681 | 226 |
| Anacardiaceae | Pistacia vera | Auxin response factor 5 | XP_031270171.1 | 2841 | 946 |
| Amaranthaceae | Chenopodium quinoa | Auxin-induced protein 22D-like | XP_021742258.1 | 546 | 181 |
| Cucurbitaceae | Cucumis sativus | Auxin-responsive protein SAUR32 | XP_011656853.1 | 303 | 100 |
| Solanaceae | Capsicum chinense | Auxin-responsive protein SAUR32 | PHT98376.1 | 501 | 166 |
| Pedaliaceae | Sesamum indicum | Auxin response factor 9 | XP_011074533.1 | 1890 | 629 |
| Rosaceae | Pyrus bretschneideri | Auxin-responsive protein SAUR50 | XP_009335221.1 | 318 | 105 |
| Oleaceae | Olea europaea subsp. Europaea | Auxin transporter 3 | CAA3012958.1 | 1398 | 465 |
| Fagaceae | Quercus suber | Auxin transporter-like protein 5 | XP_023883666.1 | 1473 | 490 |
| Anacardiaceae | Mangifera indica | Auxin-induced protein 22B-like | XP_044499499.1 | 570 | 189 |
| Vitaceae | Vitis vinifera | Auxin-responsive protein SAUR50 | RVW80229.1 | 561 | 186 |
| Euphorbiaceae | Manihot esculenta | Auxin transporter-like protein 2 | XP_021623668.1 | 1452 | 483 |
| Proteaceae | Telopea speciosissima | Auxin-responsive protein IAA27 | XP_043724596.1 | 981 | 326 |
| Rosaceae | Rosa chinensis | Auxin-responsive protein IAA31 | XP_024192080.1 | 633 | 210 |
| Rosaceae | Malus domestica | Auxin responsive protein | AZI15332.1 | 1092 | 363 |
| Brassicaceae | Brassica napus | Auxin-responsive protein IAA7 | XP_013749219.1 | 729 | 242 |
| Amaranthaceae | Spinacia oleracea | Auxin response factor 11 | XP_021846673.1 | 1995 | 664 |
| Papaveraceae | Macleaya cordata | Auxin-induced protein | OVA12640.1 | 285 | 94 |
| Malvaceae | Durio zibethinus | Auxin-induced protein 6B-like | XP_022772137.1 | 438 | 145 |
| Theaceae | Camellia sinensis | Auxin-responsive protein SAUR71-like | XP_028108662.1 | 402 | 133 |
| Myricaceae | Morella rubra | Auxin-responsive protein IAA26 | KAB1218495.1 | 1098 | 365 |
| Rosaceae | Prunus avium | Auxin-responsive protein IAA26-like | XP_021818768.1 | 1080 | 359 |
| Malvaceae | Herrania umbratica | Auxin-responsive protein IAA27 | XP_021276374.1 | 919 | 312 |
| Cucurbitaceae | Momordica charantia | Auxin-induced protein 6B-like | XP_022138000.1 | 453 | 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Zhang, S.; Du, S.; Wang, G.; Zhang, J.; Jiang, J. Effects of Exogenous (K+) Potassium Application on Plant Hormones in the Roots of Tamarix ramosissima under NaCl Stress. Genes 2022, 13, 1803. https://doi.org/10.3390/genes13101803
Chen Y, Zhang S, Du S, Wang G, Zhang J, Jiang J. Effects of Exogenous (K+) Potassium Application on Plant Hormones in the Roots of Tamarix ramosissima under NaCl Stress. Genes. 2022; 13(10):1803. https://doi.org/10.3390/genes13101803
Chicago/Turabian StyleChen, Yahui, Shiyang Zhang, Shanfeng Du, Guangyu Wang, Jinchi Zhang, and Jiang Jiang. 2022. "Effects of Exogenous (K+) Potassium Application on Plant Hormones in the Roots of Tamarix ramosissima under NaCl Stress" Genes 13, no. 10: 1803. https://doi.org/10.3390/genes13101803
APA StyleChen, Y., Zhang, S., Du, S., Wang, G., Zhang, J., & Jiang, J. (2022). Effects of Exogenous (K+) Potassium Application on Plant Hormones in the Roots of Tamarix ramosissima under NaCl Stress. Genes, 13(10), 1803. https://doi.org/10.3390/genes13101803