
On the Origin of Tetraploid Vernal Grasses (Anthoxanthum) in Europe


Abstract
1. Introduction
2. Materials and Methods
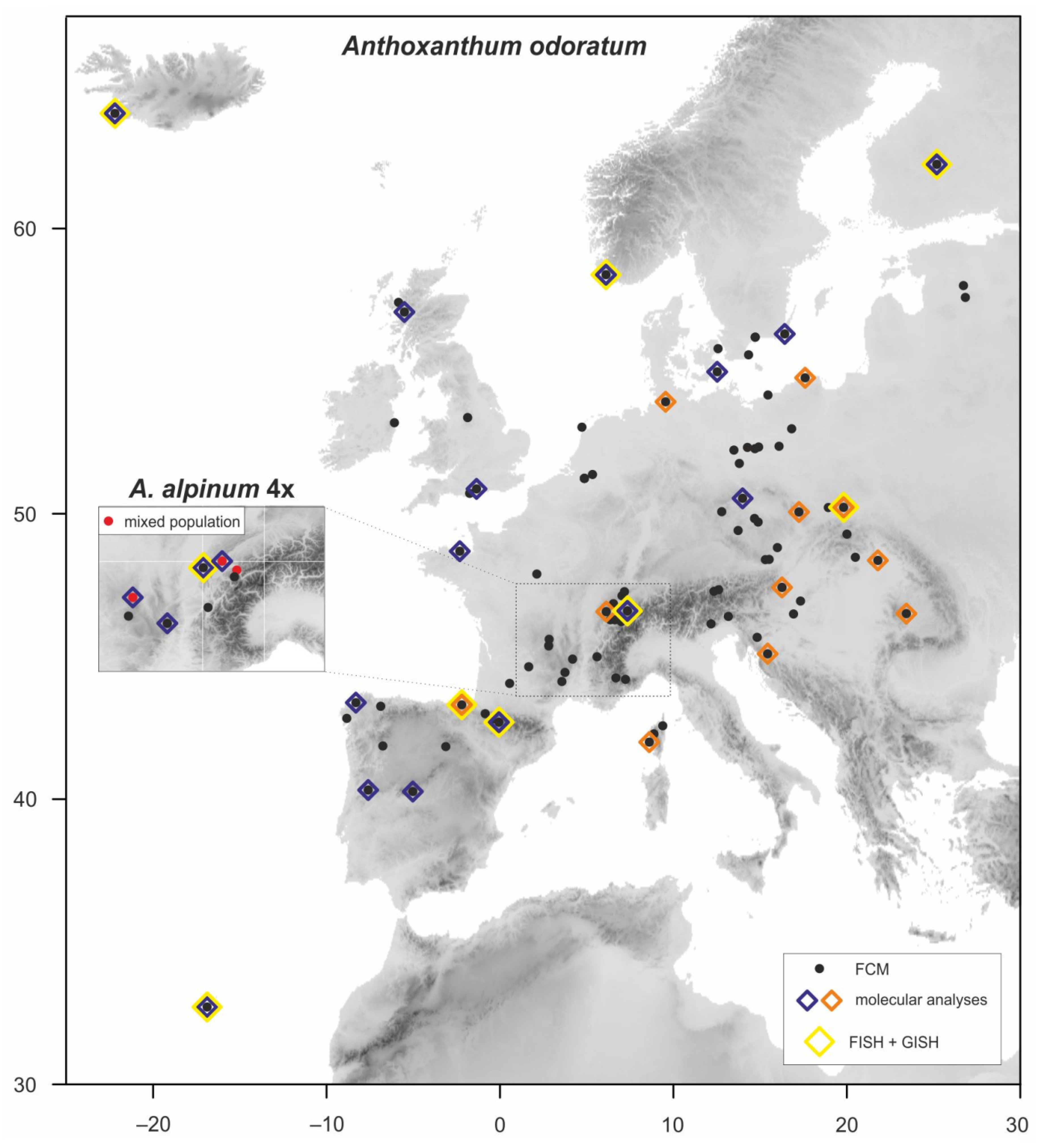
2.1. Plant Material
2.2. Flow Cytometry
2.3. Molecular Data Collection
2.4. Molecular Data Analyses (cpDNA and nrDNA)
2.5. Chromosome Preparations
2.6. DNA Probes
2.7. In Situ Hybridization
3. Results
3.1. Intraspecific Variation in Nuclear Genome Size
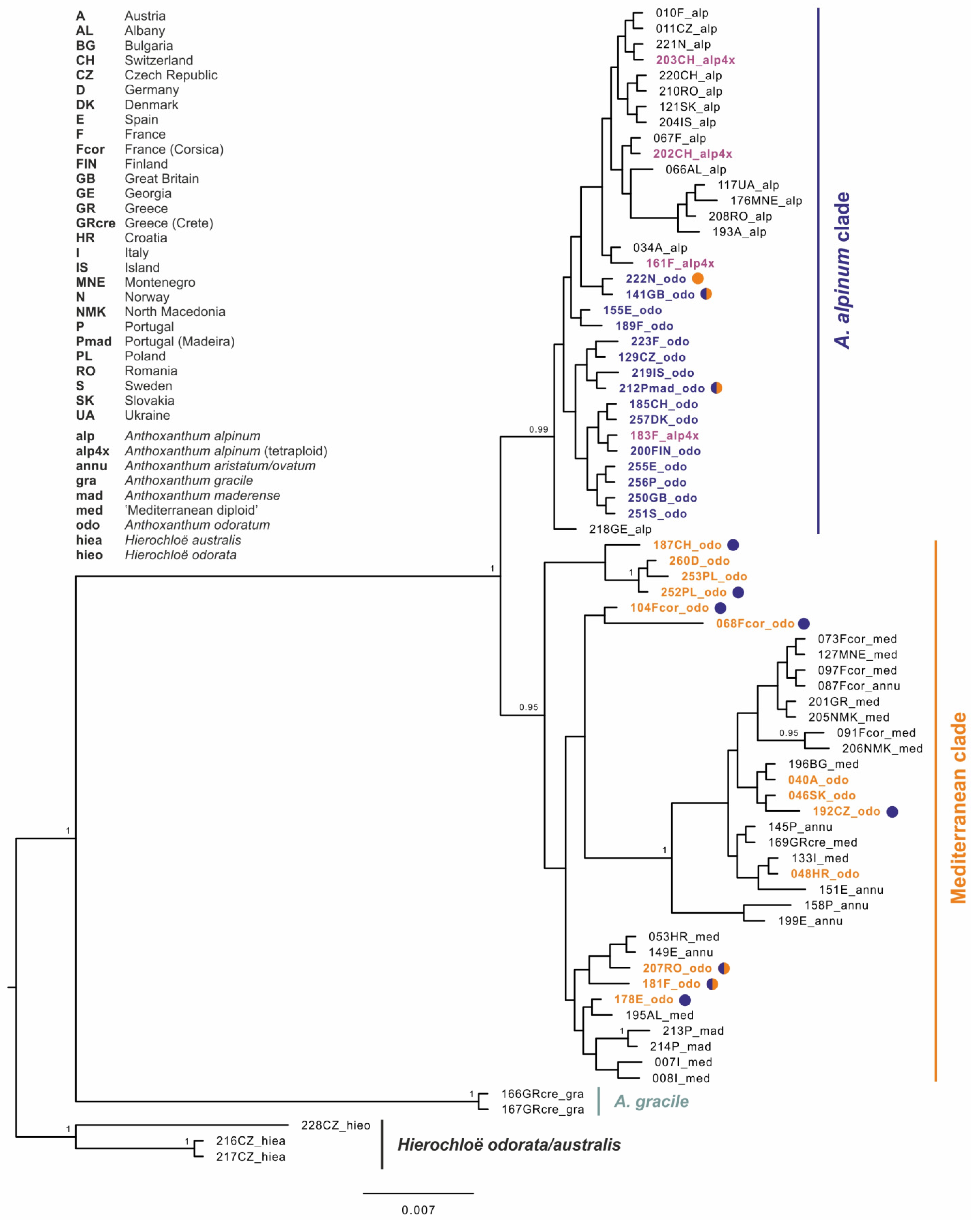
3.2. Molecular Analyses–Plastid DNA
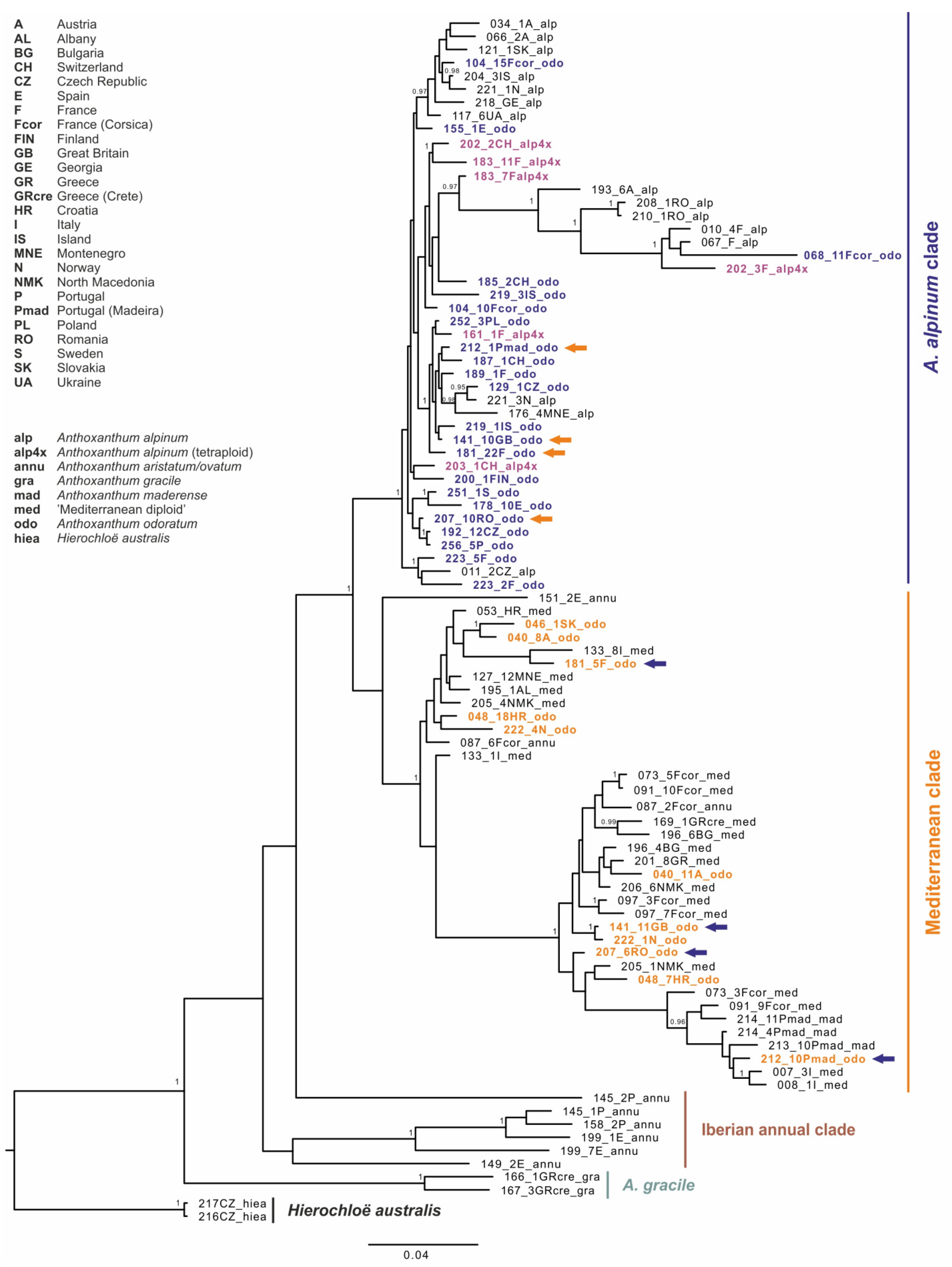
3.3. Molecular Analyses–nrDNA–GBSSI
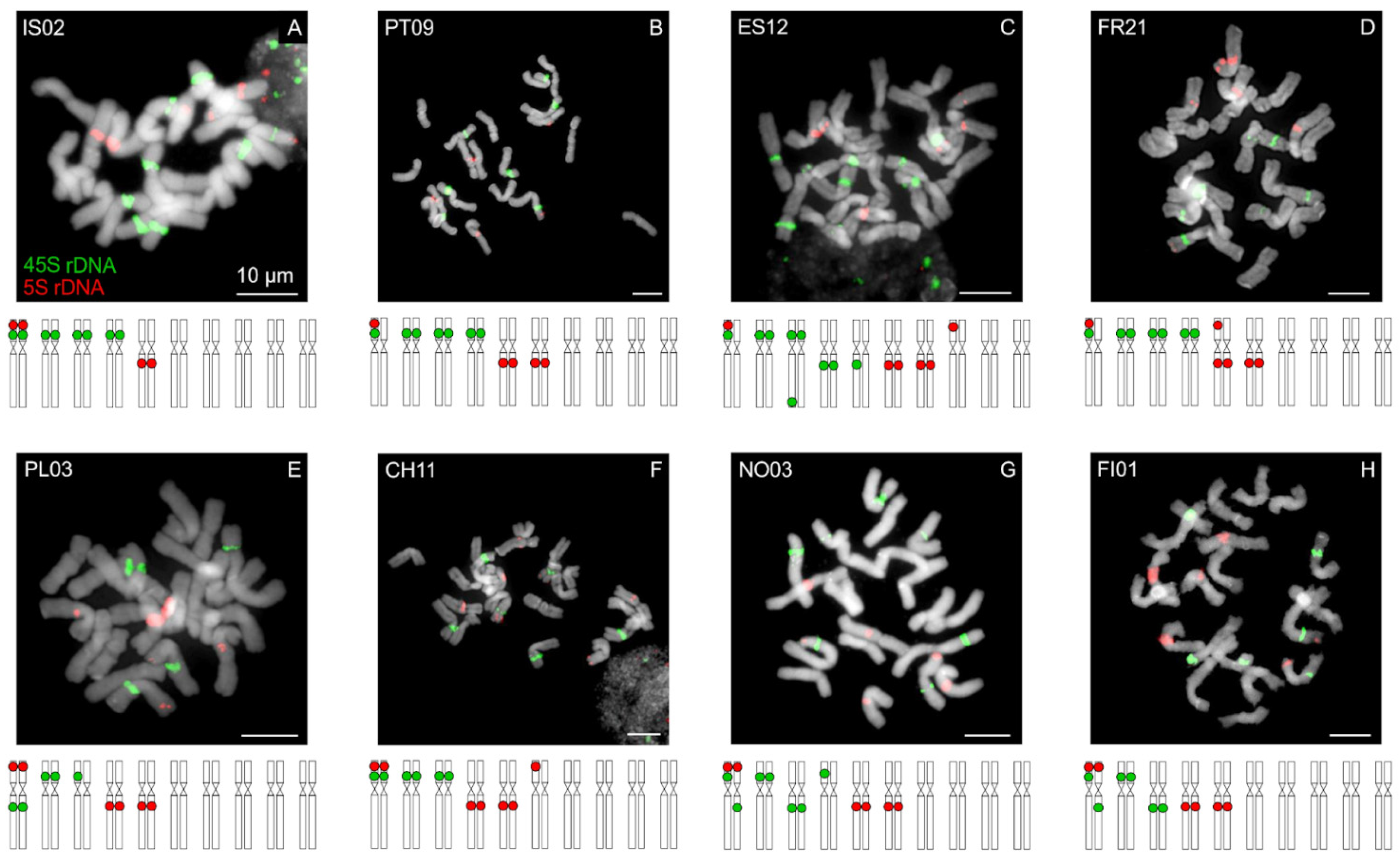
3.4. Chromosome Localization of rDNA Loci
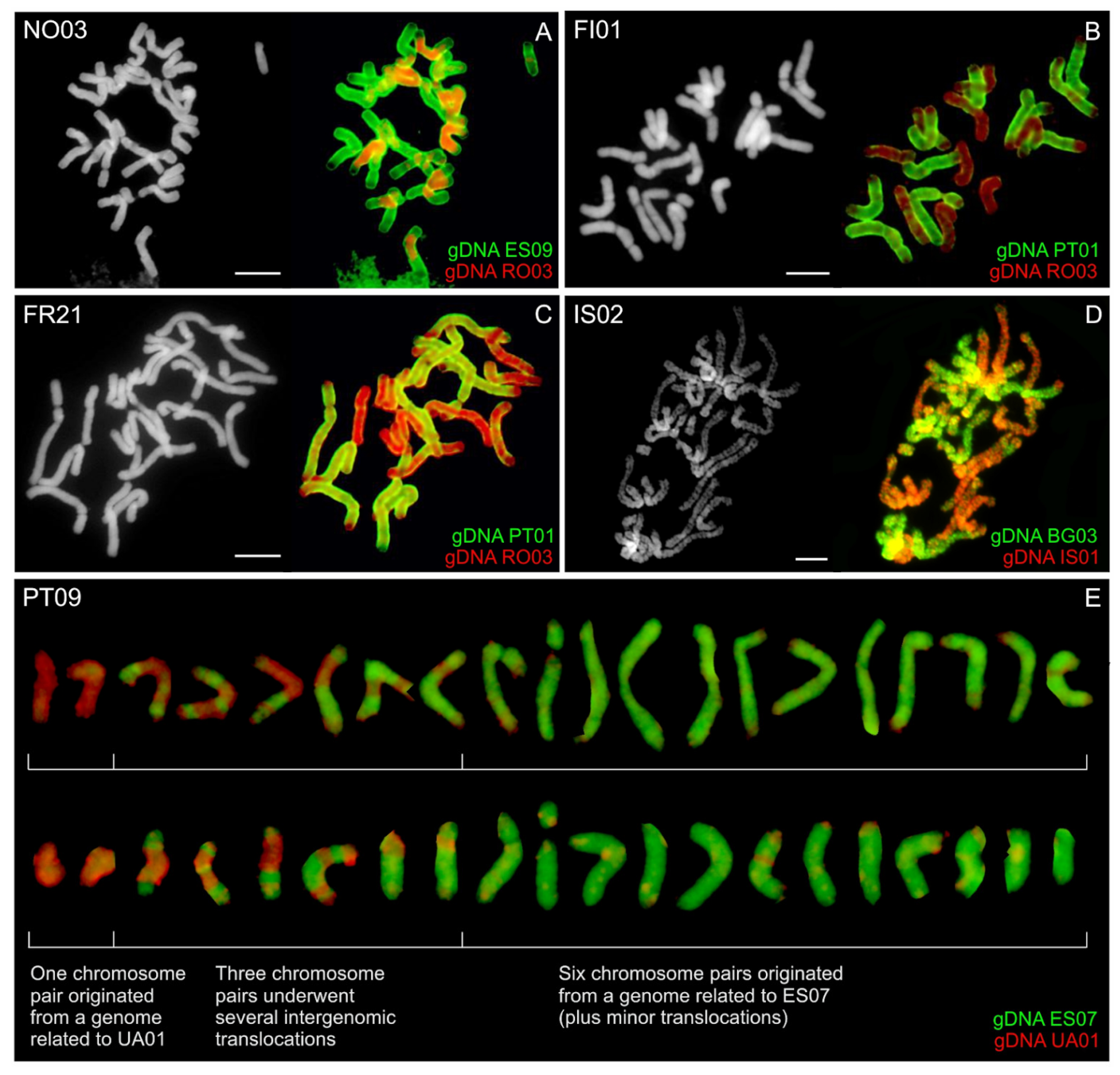
3.5. GISH in Polyploid Anthoxanthum Species’
4. Discussion
4.1. Intraspecific Variation in Nuclear Genome Size
4.2. Intraspecific Variation in Molecular and Cytogenomic Analyses
4.3. On the Origin of Anthoxanthum odoratum
4.4. Autopolyploid Origin of Anthoxanthum alpinum
4.5. Consequences of Allopolyploidy in A. odoratum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borrill, M. Experimental Studies of Evolution in Anthoxanthum (Gramineae). Genetica 1963, 34, 183–210. [Google Scholar] [CrossRef]
- Pimentel, M.; Sahuquillo, E. Relationships among Some Populations of Anthoxanthum alpinum and A. odoratum (Poaceae, Pooideae): A Morphological/Anatomical Approach. Aliso 2007, 23, 472–484. [Google Scholar] [CrossRef][Green Version]
- Chumová, Z.; Krejčíková, J.; Mandáková, T.; Suda, J.; Trávníček, P. Evolutionary and Taxonomic Implications of Variation in Nuclear Genome Size: Lesson from the Grass Genus Anthoxanthum (Poaceae). PLoS ONE 2015, 10, e0133748. [Google Scholar] [CrossRef] [PubMed]
- Paunero, E. Las Especies Españoles Del Género Anthoxanthum L. An. Inst. Botánico AJ Cavanilles 1953, 12, 401–442. [Google Scholar]
- Tutin, T. Anthoxanthum L. In Flora Europaea; Tutin, T., Heywood, V., Burges, N., Moore, D., Valentine, D., Walters, S., Webb, D., Eds.; Cambridge University Press: Cambridge, UK, 1980; Volume 5. [Google Scholar]
- Teppner, H. Anthoxanthum maderense Spec. Nova and A. odoratum (Poaceae-Aveneae) from Madeira and Their Chromosome Morphology. Phyton Ann. Rei Bot. 1998, 38, 307–321. [Google Scholar]
- Pimentel, M.; Catalan, P.; Sahuquillo, E. Morphological and Molecular Taxonomy of the Annual Diploids Anthoxanthum aristatum and A. ovatum (Poaceae) in the Iberian Peninsula. Evidence of Introgression in Natural Populations. Bot. J. Linn. Soc. 2010, 164, 53–71. [Google Scholar] [CrossRef][Green Version]
- Valdés Valdés, B. Revisión de Las Especies Anuales Del Género Anthoxanthum (Graminae). Lagascalia 1973, 3, 99–141. [Google Scholar]
- Pimentel, M.; Sahuquillo, E.; Torrecilla, Z.; Popp, M.; Catalan, P.; Brochmann, C. Hybridization and Long-Distance Colonization at Different Time Scales: Towards Resolution of Long-Term Controversies in the Sweet Vernal Grasses (Anthoxanthum). Ann. Bot. 2013, 112, 1015–1030. [Google Scholar] [CrossRef]
- Clayton, W.D. Gramineae (Part I). In Flora of Tropical East Africa; Redhead, E.M., Polhill, E.M., Eds.; Ministry for Overseas Development: London, UK, 1970. [Google Scholar]
- Tusiime, F.M.; Gizaw, A.; Wondimu, T.; Masao, C.A.; Abdi, A.A.; Muwanika, V.; Trávníček, P.; Nemomissa, S.; Popp, M.; Eilu, G.; et al. Sweet Vernal Grasses (Anthoxanthum) Colonized African Mountains along Two Fronts in the Late Pliocene, Followed by Secondary Contact, Polyploidization and Local Extinction in the Pleistocene. Mol. Ecol. 2017, 26, 3513–3532. [Google Scholar] [CrossRef]
- Hedberg, I. Cytotaxonomic Reconnaissance of Tropical African Anthoxanthum L. (Gramineae). Bot. Not. 1976, 129, 85–90. [Google Scholar]
- Chumová, Z.; Záveská, E.; Mandáková, T.; Krak, K.; Trávníček, P. The Mediterranean: The Cradle of Anthoxanthum (Poaceae) Diploid Diversity. Ann. Bot. 2017, 120, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Jones, K. Chromosomes and the Origin of Anthoxanthum odoratum L. Chromosoma 1964, 15, 248–274. [Google Scholar] [CrossRef]
- Hedberg, O. Afro-Alpine Vascular Plants. Symb. Bot. Ups. 1957, 15, 54. [Google Scholar]
- Felber, F. Distribution des cytodèmes d’Anthoxanthum odoratum L. s lat. en Suisse. Les relations Alpes-Jura. Bot. Helvetica 1986, 96, 145–158. [Google Scholar]
- Phillips, S. Poaceae (Graminae). In Flora of Ethiopia and Eritreia; Hedberg, I., Edwards, S., Eds.; Swedish Science Press: Upsalla, Sweden, 1995; pp. 40–42. [Google Scholar]
- Teppner, H. Poaceae in the Greenhouses of the Botanic Garden Institute of Botany in Graz (Austria, Europe). Fritschiana 2002, 31, 1–42. [Google Scholar]
- Mashau, A.C. A Synopsis of Anthoxanthum (Poaceae: Pooideae: Poeae) in Southern Africa and Description of a New Subspecies. Kew Bull. 2016, 71, 18. [Google Scholar] [CrossRef][Green Version]
- Katterman, G. Über Die Bildung Polyvalenter Chromosomenverbände Bei Einigen Gramineen. Planta 1931, 12, 734–774. [Google Scholar] [CrossRef]
- Parthasarathy, N. Cytogenetical Studies in Oryzeae and Phalaridineae. Ann. Bot. Lond. NS 1939, 3, 43–76. [Google Scholar] [CrossRef]
- Östergren, G. Chromosome Numbers in Anthoxanthum. Hereditas 1942, 33, 242–243. [Google Scholar]
- Hedberg, I. Cytotaxonomic Studies on Anthoxanthum odoratum L. s. Lat. 2. Investigations of Some Swedish and of a Few Swiss Population Samples. Symb. Bot. Ups. 1967, 18, 5–97. [Google Scholar]
- Hedberg, I. Cytotaxonomic Studies on Anthoxanthum odoratum L. s. Lat. IV. Karyotypes, Meiosis and Origin of Tetraploid A. odoratum. Hereditas 1970, 48, 471–502. [Google Scholar]
- Hedberg, I. The Genesis of Tetraploid Anthoxanthum odoratum. Symb. Bot. Ups. 1986, 27, 147–154. [Google Scholar]
- Teppner, H. Caryotypes of European, Perennial Species of Gramineae Genus Anthoxanthum. Osterreichische Bot. Z. 1970, 118, 280–292. [Google Scholar] [CrossRef]
- Hedberg, I. Morphological, Cytotaxonomic and Evolutionary Studies in Anthoxanthum odoratum L. s. Lat.—A Critical Review. Sommerfeltia 1990, 11, 97–107. [Google Scholar]
- Hodkinson, T.R.; Chase, M.W.; Takahashi, C.; Leitch, I.J.; Bennett, M.D.; Renvoize, S.A. The Use of DNA Sequencing (ITS and TrnL-F), AFLP, and Fluorescent in Situ Hybridization to Study Allopolyploid Miscanthus (Poaceae). Am. J. Bot. 2002, 89, 279–286. [Google Scholar] [CrossRef]
- Ellneskog-Staam, P.; Salomon, B.; von Bothmer, R.; Anamthawat-Jonsson, K. The Genome Composition of Hexaploid Psammopyrum athericum and Octoploid Psammopyrum pungens (Poaceae: Triticeae). Genome 2003, 46, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, J.; Kopecky, D.; Castro, S.; Santos, C.; Silveira, P. Flow Cytometric and Cytogenetic Analyses of Iberian Peninsula Festuca Spp. Plant Syst. Evol. 2007, 269, 89–105. [Google Scholar] [CrossRef]
- Mahelka, V.; Kopecký, D. Gene Capture from across the Grass Family in the Allohexaploid Elymus repens (L.) Gould (Poaceae, Triticeae) as Evidenced by ITS, GBSSI, and Molecular Cytogenetics. Mol. Biol. Evol. 2010, 27, 1370–1390. [Google Scholar] [CrossRef]
- Felber, F. Phenology of Flowering in Diploid and Tetraploid Populations of Anthoxanthum alpinum and Anthoxanthum odoratum. Can. J. Bot. Rev. Can. Bot. 1988, 66, 2258–2264. [Google Scholar] [CrossRef]
- Felber, F. Sensitivity of the Four Cytodemes of Anthoxanthum odoratum L. s. Lat. (Poaceae) to Puccinia sardonensis Gaumann (Uredinales). Taxon 1987, 36, 573–577. [Google Scholar] [CrossRef]
- Zeroual-Humbert-Droz, C.; Felber, F. Evidence from Isozyme Analysis of Autopolyploidy in Anthoxanthum alpinum A. & D. Löve. Bot. Helvetica 1999, 109, 217–227. [Google Scholar]
- Greilhuber, J.; Dolezel, J.; Lysak, M.A.; Bennett, M.D. The Origin, Evolution and Proposed Stabilization of the Terms “genome Size” and “C-Value” to Describe Nuclear DNA Contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef]
- Otto, S.P. The Evolutionary Consequences of Polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of Nuclear DNA Content in Plants Using Flow Cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Trávníček, P.; Ponert, J.; Urfus, T.; Jersáková, J.; Vrána, J.; Hřibová, E.; Doležel, J.; Suda, J. Challenges of Flow-Cytometric Estimation of Nuclear Genome Size in Orchids, a Plant Group with Both Whole-Genome and Progressively Partial Endoreplication. Cytom. A 2015, 87A, 958–966. [Google Scholar] [CrossRef]
- Suda, J.; Krahulcová, A.; Trávníček, P.; Krahulec, F. Ploidy Level versus DNA Ploidy Level: An Appeal for Consistent Terminology. Taxon 2006, 55, 447–450. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal Primers for Amplification of 3 Noncoding Regions of Chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of Whole Chloroplast Genome Sequences to Choose Noncoding Regions for Phylogenetic Studies in Angiosperms: The Tortoise and the Hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Mason-Gamer, R.J.; Weil, C.F.; Kellogg, E.A. Granule-Bound Starch Synthase: Structure, Function, and Phylogenetic Utility. Mol. Biol. Evol. 1998, 15, 1658–1673. [Google Scholar] [CrossRef] [PubMed]
- Záveská, E.; Fér, T.; Šída, O.; Krak, K.; Marhold, K.; Leong-Škorničková, J. Phylogeny of Curcuma (Zingiberaceae) Based on Plastid and Nuclear Sequences: Proposal of the New Subgenus Ecomata. Taxon 2012, 61, 747–763. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit, Biological Sequence Alignment Editor for Win95/98/NT/2K/XP, Version 7.0.4.1. 2004. Available online: http://www.Mbio.Ncsu.Edu/BioEdit/Bioedit.Html (accessed on 12 December 2020).
- Popp, M.; Erixon, P.; Eggens, F.; Oxelman, B. Origin and Evolution of a Circumpolar Polyploid Species Complex in Silene (Caryophyllaceae) Inferred from Low Copy Nuclear RNA Polymerase Introns, RDNA, and Chloroplast DNA. Syst. Bot. 2005, 30, 302–313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.J. ModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model Selection and Model Averaging in Phylogenetics: Advantages of Akaike Information Criterion and Bayesian Approaches over Likelihood Ratio Tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A Plant DNA Minipreparation: Version II. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Mandáková, T.; Lysak, M.A. Painting of Arabidopsis chromosomes with chromosome-specific BAC clones. In Current Protocols in Plant Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; ISBN 978-1-119-07736-7. [Google Scholar]
- Greilhuber, J. Intraspecific Variation in Genome Size in Angiosperms: Identifying Its Existence. Ann. Bot. 2005, 95, 91–98. [Google Scholar] [CrossRef]
- Šmarda, P.; Bureš, P. Understanding Intraspecific Variation in Genome Size in Plants. Preslia 2010, 82, 41–61. [Google Scholar]
- Chumová, Z.; Mandáková, T.; Trávníček, P. Are B-Chromosomes Responsible for the Extraordinary Genome Size Variation in Selected Anthoxanthum Annuals? Plant Syst. Evol. 2016, 302, 731–738. [Google Scholar] [CrossRef]
- Bennett Michael David Nuclear DNA Content and Minimum Generation Time in Herbaceous Plants. Proc. R. Soc. Lond. B Biol. Sci. 1972, 181, 109–135. [CrossRef]
- Creber, H.M.C.; Davies, M.S.; Francis, D.; Walker, H.D. Variation in DNA C Value in Natural Populations of Dactylis glomerata L. New Phytol. 1994, 128, 555–561. [Google Scholar] [CrossRef]
- Reeves, G.; Francis, D.; Davies, M.S.; Rogers, H.J.; Hodkinson, T.R. Genome Size Is Negatively Correlated with Altitude in Natural Populations of Dactylis glomerata. Ann. Bot. 1998, 82, 99–105. [Google Scholar] [CrossRef]
- Šmarda, P.; Bureš, P. Intraspecific DNA Content Variability in Festuca pallens on Different Geographical Scales and Ploidy Levels. Ann. Bot. 2006, 98, 665–678. [Google Scholar] [CrossRef]
- Pečinka, A.; Suchánková, P.; Lysak, M.A.; Trávníček, B.; Doležel, J. Nuclear DNA Content Variation among Central European Koeleria Taxa. Ann. Bot. 2006, 98, 117–122. [Google Scholar] [CrossRef]
- Laurie, D.A.; Bennett, M.D. Nuclear DNA Content in the Genera Zea and Sorghum. Intergeneric, Interspecific and Intraspecific Variation. Heredity 1985, 55, 307–313. [Google Scholar] [CrossRef]
- Stritt, C.; Wyler, M.; Gimmi, E.L.; Pippel, M.; Roulin, A.C. Diversity, Dynamics and Effects of Long Terminal Repeat Retrotransposons in the Model Grass Brachypodium distachyon. New Phytol. 2020, 227, 1736–1748. [Google Scholar] [CrossRef]
- Zwyrtková, J.; Němečková, A.; Čížková, J.; Holušová, K.; Kapustová, V.; Svačina, R.; Kopecký, D.; Till, B.J.; Doležel, J.; Hřibová, E. Comparative Analyses of DNA Repeats and Identification of a Novel Fesreba Centromeric Element in Fescues and Ryegrasses. BMC Plant Biol. 2020, 20, 280. [Google Scholar] [CrossRef]
- Giraud, D.; Lima, O.; Huteau, V.; Coriton, O.; Boutte, J.; Kovarik, A.; Leitch, A.R.; Leitch, I.J.; Aïnouche, M.; Salmon, A. Evolutionary Dynamics of Transposable Elements and Satellite DNAs in Polyploid Spartina Species. Plant Sci. 2021, 302, 110671. [Google Scholar] [CrossRef]
- Vaio, M.; Speranza, P.; Valls, J.F.; Guerra, M.; Mazzella, C. Localization of the 5S and 45S RDNA Sites and CpDNA Sequence Analysis in Species of the Quadrifaria Group of Paspalum (Poaceae, Paniceae). Ann. Bot. 2005, 96, 191–200. [Google Scholar] [CrossRef]
- González, M.L.; Urdampilleta, J.D.; Fasanella, M.; Premoli, A.C.; Chiapella, J.O. Distribution of RDNA and Polyploidy in Deschampsia antarctica E. Desv. in Antarctic and Patagonic Populations. Polar Biol. 2016, 39, 1663–1677. [Google Scholar] [CrossRef]
- González, M.L.; Chiapella, J.O.; Urdampilleta, J.D. The Antarctic and South American Species of Deschampsia: Phylogenetic Relationships and Cytogenetic Differentiation. Syst. Biodivers. 2021, 1–40. [Google Scholar] [CrossRef]
- Amosova, A.V.; Bolsheva, N.L.; Zoshchuk, S.A.; Twardovska, M.O.; Yurkevich, O.Y.; Andreev, I.O.; Samatadze, T.E.; Badaeva, E.D.; Kunakh, V.A.; Muravenko, O.V. Comparative Molecular Cytogenetic Characterization of Seven Deschampsia (Poaceae) Species. PLoS ONE 2017, 12, e0175760. [Google Scholar] [CrossRef]
- Drapikowska, M.; Susek, K.; Hasterok, R.; Szkudlarz, P.; Celka, Z.; Jackowiak, B. Variability of Stomata and 45s and 5s Rdnas Loci Characteristics in Two Species of Anthoxanthum Genus: A. aristatum and A. odoratum (Poaceae). Acta Biol. Hung. 2013, 64, 352–363. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Characteristics | t-Value (df = 101) | p-Value | Correlation Coefficient |
|---|---|---|---|
| Latitude | −3.42 | <0.001 | −0.322 |
| Longitude | −0.69 | 0.505 | −0.066 |
| Altitude [m a.s.l.] | −2.80 | 0.006 | −0.269 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chumová, Z.; Mandáková, T.; Trávníček, P. On the Origin of Tetraploid Vernal Grasses (Anthoxanthum) in Europe. Genes 2021, 12, 966. https://doi.org/10.3390/genes12070966
Chumová Z, Mandáková T, Trávníček P. On the Origin of Tetraploid Vernal Grasses (Anthoxanthum) in Europe. Genes. 2021; 12(7):966. https://doi.org/10.3390/genes12070966
Chicago/Turabian StyleChumová, Zuzana, Terezie Mandáková, and Pavel Trávníček. 2021. "On the Origin of Tetraploid Vernal Grasses (Anthoxanthum) in Europe" Genes 12, no. 7: 966. https://doi.org/10.3390/genes12070966
APA StyleChumová, Z., Mandáková, T., & Trávníček, P. (2021). On the Origin of Tetraploid Vernal Grasses (Anthoxanthum) in Europe. Genes, 12(7), 966. https://doi.org/10.3390/genes12070966