Heme, A Metabolic Sensor, Directly Regulates the Activity of the KDM4 Histone Demethylase Family and Their Interactions with Partner Proteins



Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Plasmids
2.2. Cell Growth and β-Galactosidase Assays
2.3. Purification of KDM4 and Gis1 proteins, Spectroscopic Analyses, and Protein Binding to Heme–Agarose Beads
2.4. Measurement of KDM4A/B/C and Gis1 Demethylase Activity
2.5. Identification of Gis1 Interactors, Mass Spectrometry (MS), and Proteomic Data Analysis
2.6. Preparation of Yeast Extracts, TAP-Pulldown, and Western Blotting
3. Results
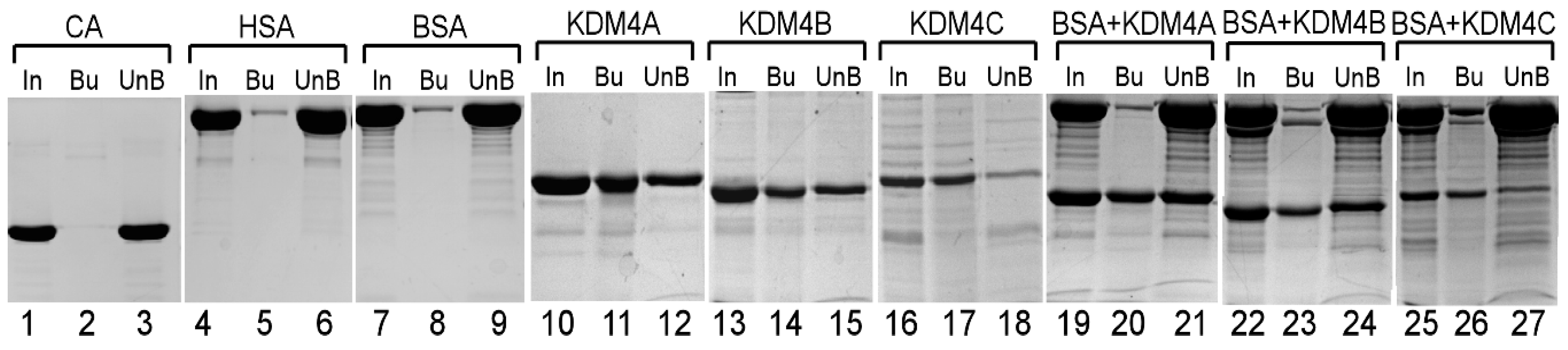
3.1. Heme Binds to the JmjN/C Domain of KDM4A/B/C and Regulates the Histone Demethylase Activity of KDM4A and C, but not KDM4B
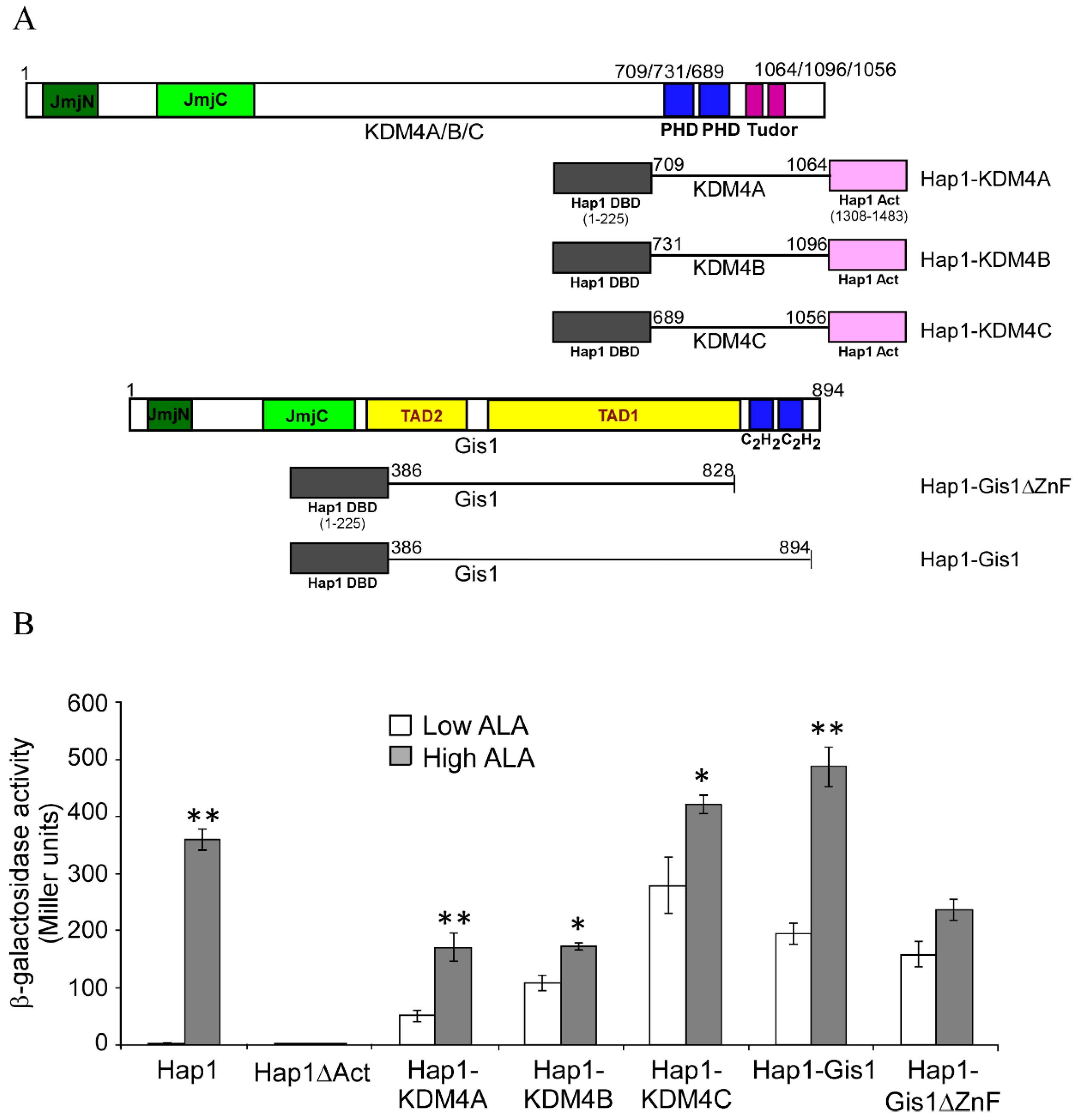
3.2. The C-terminal Regions of KDM4A/B/C, like that of Gis1, Have the Potential to Modulate the Activity of the JmjN/C Domain and Heme Regulation
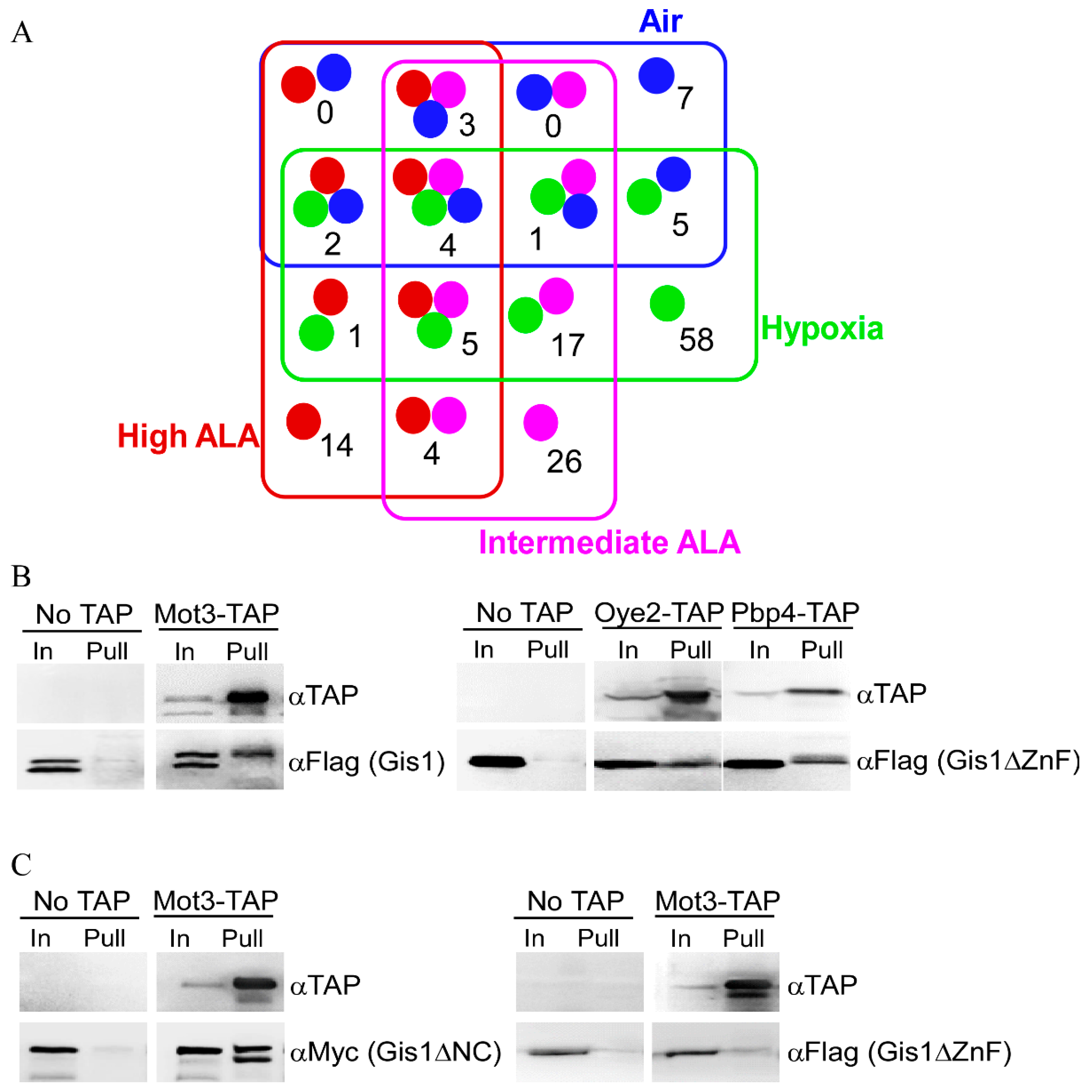
3.3. Mass Spectrometry (MS) Identified 147 Proteins that can Interact with Gis1 In Vitro
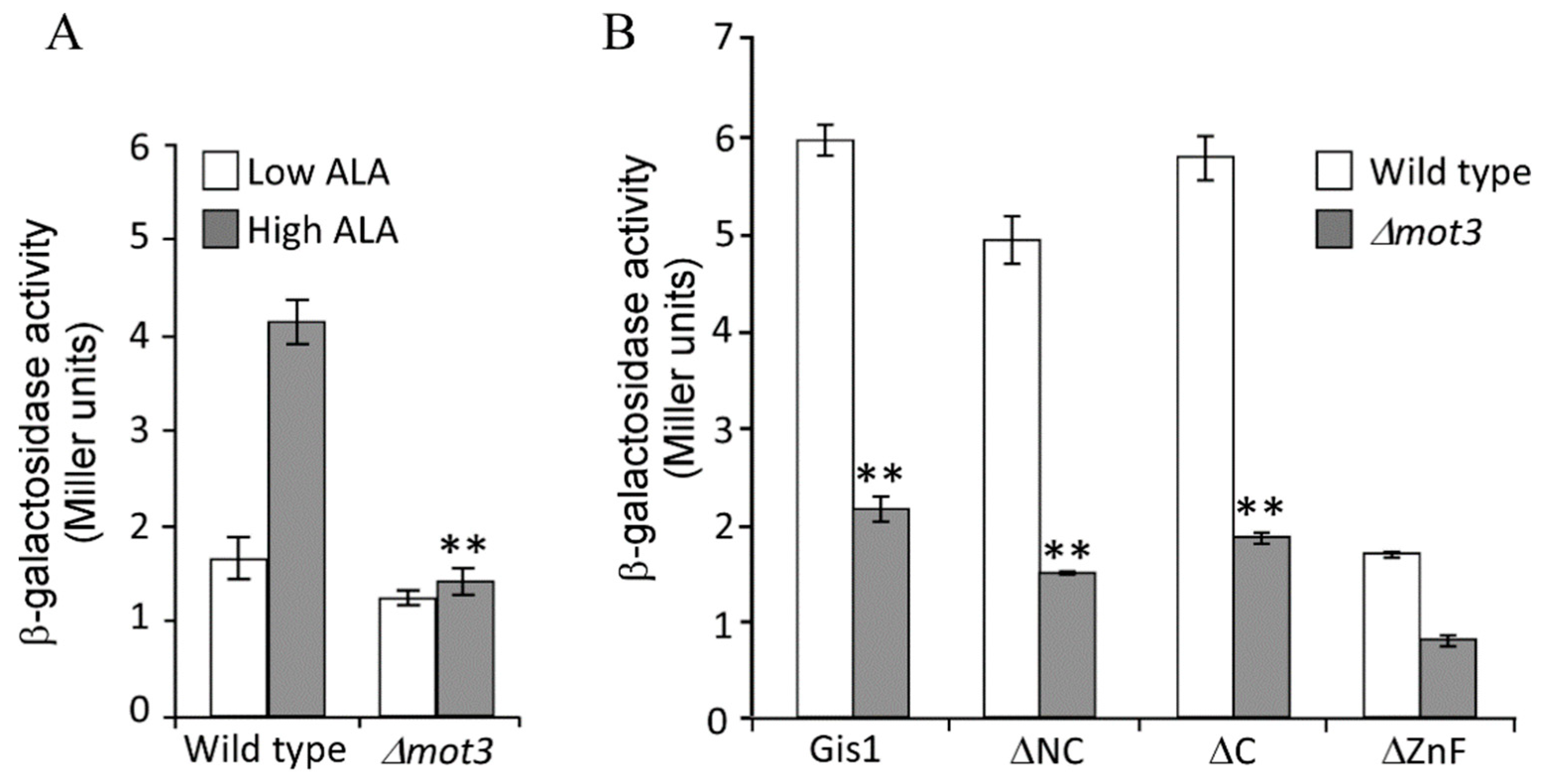
3.4. Mot3 is Essential for Heme Activation of Gis1 Transcriptional Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shi, Y. Histone lysine demethylases: Emerging roles in development, physiology and disease. Nat. Rev. Genet. 2007, 8, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Klose, R.J.; Zhang, Y. Regulation of histone methylation by demethylimination and demethylation. Nat. Rev. Mol. Cell Biol. 2007, 8, 307–318. [Google Scholar] [CrossRef]
- Chen, Z.; Zang, J.; Whetstine, J.; Hong, X.; Davrazou, F.; Kutateladze, T.G.; Simpson, M.; Mao, Q.; Pan, C.H.; Dai, S.; et al. Structural insights into histone demethylation by JMJD2 family members. Cell 2006, 125, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.; Tumber, A.; Che, K.; Cain, P.; Nowak, R.; Gileadi, C.; Oppermann, U. The roles of Jumonji-type oxygenases in human disease. Epigenomics 2014, 6, 89–120. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Walport, L.J.; Hopkinson, R.J.; Madden, S.K.; Chowdhury, R.; Schofield, C.J.; Kawamura, A. Studies on the catalytic domains of multiple JmjC oxygenases using peptide substrates. Epigenetics 2014, 9, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Zhang, J.; Yonezawa, M.; Schmidt, A.; Zheng, H.; Jenuwein, T.; Reinberg, D. Dynamic Histone H1 Isotype 4 Methylation and Demethylation by Histone Lysine Methyltransferase G9a/KMT1C and the Jumonji Domain-containing JMJD2/KDM4 Proteins. J. Biol. Chem. 2009, 284, 8395–8405. [Google Scholar] [CrossRef]
- Hillringhaus, L.; Yue, W.W.; Rose, N.R.; Ng, S.S.; Gileadi, C.; Loenarz, C.; Bello, S.H.; Bray, J.E.; Schofield, C.J.; Oppermann, U. Structural and evolutionary basis for the dual substrate selectivity of human KDM4 histone demethylase family. J. Biol. Chem. 2011, 286, 41616–41625. [Google Scholar] [CrossRef]
- Labbe, R.M.; Holowatyj, A.; Yang, Z.Q. Histone lysine demethylase (KDM) subfamily 4: Structures, functions and therapeutic potential. Am. J. Transl. Res. 2014, 6, 1–15. [Google Scholar]
- Cloos, P.A.; Christensen, J.; Agger, K.; Maiolica, A.; Rappsilber, J.; Antal, T.; Hansen, K.H.; Helin, K. The putative oncogene GASC1 demethylates tri- and dimethylated lysine 9 on histone H3. Nature 2006, 442, 307–311. [Google Scholar] [CrossRef]
- Wissmann, M.; Yin, N.; Muller, J.M.; Greschik, H.; Fodor, B.D.; Jenuwein, T.; Vogler, C.; Schneider, R.; Gunther, T.; Buettner, R.; et al. Cooperative demethylation by JMJD2C and LSD1 promotes androgen receptor-dependent gene expression. Nat. Cell Biol. 2007, 9, 347–353. [Google Scholar] [CrossRef]
- Klose, R.J.; Kallin, E.M.; Zhang, Y. JmjC-domain-containing proteins and histone demethylation. Nat. Rev. Genet. 2006, 7, 715–727. [Google Scholar] [CrossRef]
- Whetstine, J.R.; Nottke, A.; Lan, F.; Huarte, M.; Smolikov, S.; Chen, Z.; Spooner, E.; Li, E.; Zhang, G.; Colaiacovo, M.; et al. Reversal of histone lysine trimethylation by the JMJD2 family of histone demethylases. Cell 2006, 125, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Janknecht, R. Diversity within the JMJD2 histone demethylase family. Biochem. Biophys. Res. Commun. 2007, 353, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Agger, K.; Nishimura, K.; Miyagi, S.; Messling, J.E.; Rasmussen, K.D.; Helin, K. The KDM4/JMJD2 histone demethylases are required for hematopoietic stem cell maintenance. Blood 2019, 134, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Filiu-Braga, L.D.C.; Serejo, T.R.T.; Lucena-Araujo, A.R.; Neves, F.A.R.; de Carvalho, J.L.; Rego, E.M.; Saldanha-Araujo, F. Unraveling KDM4 histone demethylase expression and its association with adverse cytogenetic findings in chronic lymphocytic leukemia. Med. Oncol 2018, 36, 3. [Google Scholar] [CrossRef]
- Kang, M.K.; Mehrazarin, S.; Park, N.H.; Wang, C.Y. Epigenetic gene regulation by histone demethylases: Emerging role in oncogenesis and inflammation. Oral Dis 2017, 23, 709–720. [Google Scholar] [CrossRef]
- Teperino, R.; Schoonjans, K.; Auwerx, J. Histone methyl transferases and demethylases; can they link metabolism and transcription? Cell Metab. 2010, 12, 321–327. [Google Scholar] [CrossRef]
- Kaelin, W.G., Jr. Cancer and altered metabolism: Potential importance of hypoxia-inducible factor and 2-oxoglutarate-dependent dioxygenases. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 335–345. [Google Scholar] [CrossRef]
- Gut, P.; Verdin, E. The nexus of chromatin regulation and intermediary metabolism. Nature 2013, 502, 489–498. [Google Scholar] [CrossRef]
- Carbonneau, M.; Gagné, L.M.; Lalonde, M.E.; Germain, M.A.; Motorina, A.; Guiot, M.C.; Secco, B.; Vincent, E.E.; Tumber, A.; Hulea, L.; et al. The oncometabolite 2-hydroxyglutarate activates the mTOR signalling pathway. Nat. Commun. 2016, 7, 12700. [Google Scholar] [CrossRef]
- Filipp, F.V. Crosstalk between epigenetics and metabolism-Yin and Yang of histone demethylases and methyltransferases in cancer. Brief. Funct. Genomics 2017, 16, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Hiltunen, M.; Kaarniranta, K. Krebs cycle intermediates regulate DNA and histone methylation: Epigenetic impact on the aging process. Ageing Res. Rev. 2014, 16, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Lal, S.; Comer, J.M.; Konduri, P.C.; Shah, A.; Wang, T.; Lewis, A.; Shoffner, G.; Guo, F.; Zhang, L. Heme promotes transcriptional and demethylase activities of Gis1, a member of the histone demethylase JMJD2/KDM4 family. Nucleic Acids Res. 2018, 46, 215–228. [Google Scholar] [CrossRef]
- Kwon, D.W.; Ahn, S.H. Role of yeast JmjC-domain containing histone demethylases in actively transcribed regions. Biochem. Biophys. Res. Commun. 2011, 410, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Sein, H.; Varv, S.; Kristjuhan, A. Distribution and maintenance of histone H3 lysine 36 trimethylation in transcribed locus. PLoS ONE 2015, 10, e0120200. [Google Scholar] [CrossRef]
- Tu, S.; Bulloch, E.M.; Yang, L.; Ren, C.; Huang, W.C.; Hsu, P.H.; Chen, C.H.; Liao, C.L.; Yu, H.M.; Lo, W.S.; et al. Identification of histone demethylases in Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 14262–14271. [Google Scholar] [CrossRef]
- Cameroni, E.; Hulo, N.; Roosen, J.; Winderickx, J.; De Virgilio, C. The novel yeast PAS kinase Rim 15 orchestrates G0-associated antioxidant defense mechanisms. Cell Cycle 2004, 3, 462–468. [Google Scholar] [CrossRef]
- Roosen, J.; Engelen, K.; Marchal, K.; Mathys, J.; Griffioen, G.; Cameroni, E.; Thevelein, J.M.; De Virgilio, C.; De Moor, B.; Winderickx, J. PKA and Sch9 control a molecular switch important for the proper adaptation to nutrient availability. Mol. Microbiol. 2005, 55, 862–880. [Google Scholar] [CrossRef]
- Cheng, C.; Fabrizio, P.; Ge, H.; Longo, V.D.; Li, L.M. Inference of transcription modification in long-live yeast strains from their expression profiles. BMC Genomics 2007, 8, 219. [Google Scholar] [CrossRef]
- Zhang, N.; Wu, J.; Oliver, S.G. Gis1 is required for transcriptional reprogramming of carbon metabolism and the stress response during transition into stationary phase in yeast. Microbiology 2009, 155, 1690–1698. [Google Scholar] [CrossRef]
- Wei, M.; Fabrizio, P.; Madia, F.; Hu, J.; Ge, H.; Li, L.M.; Longo, V.D. Tor1/Sch9-regulated carbon source substitution is as effective as calorie restriction in life span extension. PLoS Genet. 2009, 5, e1000467. [Google Scholar] [CrossRef] [PubMed]
- Orzechowski Westholm, J.; Tronnersjo, S.; Nordberg, N.; Olsson, I.; Komorowski, J.; Ronne, H. Gis1 and Rph1 regulate glycerol and acetate metabolism in glucose depleted yeast cells. PLoS ONE 2012, 7, e31577. [Google Scholar] [CrossRef] [PubMed]
- Boorstein, W.R.; Craig, E.A. Regulation of a yeast HSP70 gene by a cAMP responsive transcriptional control element. EMBO J. 1990, 9, 2543–2553. [Google Scholar] [CrossRef] [PubMed]
- Pedruzzi, I.; Burckert, N.; Egger, P.; De Virgilio, C. Saccharomyces cerevisiae Ras/cAMP pathway controls post-diauxic shift element-dependent transcription through the zinc finger protein Gis1. EMBO J. 2000, 19, 2569–2579. [Google Scholar] [CrossRef]
- Jang, Y.K.; Wang, L.; Sancar, G.B. RPH1 and GIS1 are damage-responsive repressors of PHR1. Mol. Cell Biol. 1999, 19, 7630–7638. [Google Scholar] [CrossRef]
- Yu, Y.; Neiman, A.M.; Sternglanz, R. The JmjC domain of Gis1 is dispensable for transcriptional activation. FEMS Yeast Res. 2010, 10, 793–801. [Google Scholar] [CrossRef]
- Quan, Z.; Oliver, S.G.; Zhang, N. JmjN interacts with JmjC to ensure selective proteolysis of Gis1 by the proteasome. Microbiology 2011, 157, 2694–2701. [Google Scholar] [CrossRef]
- Pascual, J.; Martinez-Yamout, M.; Dyson, H.J.; Wright, P.E. Structure of the PHD zinc finger from human Williams-Beuren syndrome transcription factor. J. Mol. Biol. 2000, 304, 723–729. [Google Scholar] [CrossRef]
- Lu, R.; Wang, G.G. Tudor: A versatile family of histone methylation ‘readers’. Trends Biochem. Sci. 2013, 38, 546–555. [Google Scholar] [CrossRef]
- Musselman, C.A.; Kutateladze, T.G. PHD fingers: Epigenetic effectors and potential drug targets. Mol. Interv. 2009, 9, 314–323. [Google Scholar] [CrossRef]
- Musselman, C.A.; Kutateladze, T.G. Handpicking epigenetic marks with PHD fingers. Nucleic Acids Res. 2011, 39, 9061–9071. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.; Zhou, M.M. The PHD finger: A versatile epigenome reader. Trends Biochem. Sci. 2011, 36, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Adams-Cioaba, M.A.; Min, J. Structure and function of histone methylation binding proteins. Biochem. Cell Biol. 2009, 87, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W.; De Matteis, F.; Aldridge, W.N. Heme and Hemoproteins; Springer: Berlin/Heidelberg, Germany, 1978; pp. 49–80. [Google Scholar]
- Ponka, P. Cell biology of heme. Am. J. Med. Sci. 1999, 318, 241–256. [Google Scholar] [CrossRef]
- Ortiz de Montellano, P.R. Hemes in Biology. In Wiley Encyclopedia of Chemical Biology; John Wiley & Sons, Ltd.: Chichester, West Sussex, UK; Hoboken, NJ, USA, 2009; pp. 240–249. [Google Scholar]
- Padmanaban, G.; Venkateswar, V.; Rangarajan, P.N. Haem as a multifunctional regulator. Trends Biochem. Sci. 1989, 14, 492–496. [Google Scholar] [CrossRef]
- Sassa, S. Novel effects of heme and heme-related compounds in biological systems. Current Med. Chem. 1996, 3, 273–290. [Google Scholar]
- Yin, L.; Wu, N.; Curtin, J.C.; Qatanani, M.; Szwergold, N.R.; Reid, R.A.; Waitt, G.M.; Parks, D.J.; Pearce, K.H.; Wisely, G.B.; et al. Rev-erbalpha, a heme sensor that coordinates metabolic and circadian pathways. Science 2007, 318, 1786–1789. [Google Scholar] [CrossRef]
- Weitz, S.H.; Gong, M.; Barr, I.; Weiss, S.; Guo, F. Processing of microRNA primary transcripts requires heme in mammalian cells. Proc. Natl. Acad. Sci. USA 2014, 111, 1861–1866. [Google Scholar] [CrossRef]
- Zhang, L.; Guarente, L. Heme binds to a short sequence that serves a regulatory function in diverse proteins. EMBO J. 1995, 14, 313–320. [Google Scholar] [CrossRef]
- Zhang, L.; Hach, A. Molecular mechanism of heme signaling in yeast: The transcriptional activator Hap1 serves as the key mediator. Cell Mol. Life Sci. 1999, 56, 415–426. [Google Scholar] [CrossRef]
- Raghuram, S.; Stayrook, K.R.; Huang, P.; Rogers, P.M.; Nosie, A.K.; McClure, D.B.; Burris, L.L.; Khorasanizadeh, S.; Burris, T.P.; Rastinejad, F. Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta. Nat. Struct. Mol. Biol. 2007, 14, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, D.; Fiorito, V.; Petrillo, S.; Tolosano, E. Unraveling the Role of Heme in Neurodegeneration. Front. Neurosci. 2018, 12, 712. [Google Scholar] [CrossRef] [PubMed]
- Sohoni, S.; Ghosh, P.; Wang, T.; Kalainayakan, S.P.; Vidal, C.; Dey, S.; Konduri, P.C.; Zhang, L. Elevated Heme Synthesis and Uptake Underpin Intensified Oxidative Metabolism and Tumorigenic Functions in Non-Small Cell Lung Cancer Cells. Cancer Res. 2019, 79, 2511–2525. [Google Scholar] [CrossRef] [PubMed]
- Galmozzi, A.; Kok, B.P.; Kim, A.S.; Montenegro-Burke, J.R.; Lee, J.Y.; Spreafico, R.; Mosure, S.; Albert, V.; Cintron-Colon, R.; Godio, C.; et al. PGRMC2 is an intracellular haem chaperone critical for adipocyte function. Nature 2019, 576, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Rothenberg, M.; Downs, M.A.; Blumer, K.J. Mot3, a Zn finger transcription factor that modulates gene expression and attenuates mating pheromone signaling in Saccharomyces cerevisiae. Genetics 1998, 149, 879–892. [Google Scholar] [PubMed]
- Martinez-Montanes, F.; Rienzo, A.; Poveda-Huertes, D.; Pascual-Ahuir, A.; Proft, M. Activator and repressor functions of the Mot3 transcription factor in the osmostress response of Saccharomyces cerevisiae. Eukaryot. Cell 2013, 12, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, J.; Han, G.S.; Iwanyshyn, W.M.; Conover, K.; Carman, G.M. Regulation of the yeast DPP1-encoded diacylglycerol pyrophosphate phosphatase by transcription factor Gis1p. J. Biol. Chem. 2003, 278, 31495–31503. [Google Scholar] [CrossRef]
- Zhang, N.; Oliver, S.G. The transcription activity of Gis1 is negatively modulated by proteasome-mediated limited proteolysis. J. Biol. Chem. 2010, 285, 6465–6476. [Google Scholar] [CrossRef]
- Turcotte, B.; Guarente, L. HAP1 positive control mutants specific for one of two binding sites. Genes Dev. 1992, 6, 2001–2009. [Google Scholar] [CrossRef]
- Zhang, L.; Guarente, L. The C6 zinc cluster dictates asymmetric binding by HAP1. EMBO J. 1996, 15, 4676–4681. [Google Scholar] [CrossRef]
- Hon, T.; Lee, H.C.; Zhang, L. The actions of Hsp90 and Hsp70 molecular chaperones in heme signaling in eukaryotes. In Recent Research Developments in Molecular and Cellular Biology; Fagan, J., Shimizu, N., Davidson, J.N., Eds.; Research Signpost: Trivandrum, India, 2003; pp. 11–21. [Google Scholar]
- Kundaje, A.; Xin, X.; Lan, C.; Lianoglou, S.; Zhou, M.; Zhang, L.; Leslie, C. A predictive model of the oxygen and heme regulatory network in yeast. PLoS Comput. Biol. 2008, 4, e1000224. [Google Scholar] [CrossRef] [PubMed]
- Hon, T.; Hach, A.; Lee, H.C.; Chen, T.; Zhang, L. Functional analysis of heme regulatory elements of the transcriptional activator Hap1. Biochem. Biophys Res. Commun. 2000, 273, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Hon, T.; Hach, A.; Tamalis, D.; Zhu, Y.; Zhang, L. The yeast heme-responsive transcriptional activator Hap1 is a preexisting dimer in the absence of heme. J. Biol. Chem. 1999, 274, 22770–22774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Guarente, L. HAP1 is nuclear but is bound to a cellular factor in the absence of heme. J. Biol. Chem. 1994, 269, 14643–14647. [Google Scholar] [PubMed]
- Zhang, L.; Hach, A.; Wang, C. Molecular mechanism governing heme signaling in yeast: A higher-order complex mediates heme regulation of the transcriptional activator HAP1. Mol. Cell Biol. 1998, 18, 3819–3828. [Google Scholar] [CrossRef] [PubMed]
- Ghosh Dastidar, R.; Hooda, J.; Shah, A.; Cao, T.M.; Henke, R.M.; Zhang, L. The nuclear localization of SWI/SNF proteins is subjected to oxygen regulation. Cell Biosci. 2012, 2, 30. [Google Scholar] [CrossRef]
- Mellacheruvu, D.; Wright, Z.; Couzens, A.L.; Lambert, J.P.; St-Denis, N.A.; Li, T.; Miteva, Y.V.; Hauri, S.; Sardiu, M.E.; Low, T.Y.; et al. The CRAPome: A contaminant repository for affinity purification-mass spectrometry data. Nat. Methods 2013, 10, 730–736. [Google Scholar] [CrossRef]
- Hon, T.; Lee, H.C.; Hach, A.; Johnson, J.L.; Craig, E.A.; Erdjument-Bromage, H.; Tempst, P.; Zhang, L. The Hsp70-Ydj1 Molecular Chaperone Represses the Activity of the Transcriptional Activator Hap1 in the Absence of Heme. Mol. Cell Biol. 2001, 21, 7923–7932. [Google Scholar] [CrossRef]
- Gerace, E.; Moazed, D. Affinity Purification of Protein Complexes Using TAP Tags. Methods Enzymol. 2015, 559, 37–52. [Google Scholar]
- Ishimori, K.; Watanabe, Y. Unique Heme Environmental Structures in Heme-regulated Proteins Using Heme as the Signaling Molecule. Chem. Lett. 2014, 43, 1680–1689. [Google Scholar] [CrossRef]
- Yao, X.; Balamurugan, P.; Arvey, A.; Leslie, C.; Zhang, L. Heme controls the regulation of protein tyrosine kinases Jak2 and Src. Biochem. Biophys Res. Commun. 2010, 403, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Liew, C.K.; Simpson, R.J.; Kwan, A.H.; Crofts, L.A.; Loughlin, F.E.; Matthews, J.M.; Crossley, M.; Mackay, J.P. Zinc fingers as protein recognition motifs: Structural basis for the GATA-1/friend of GATA interaction. Proc. Natl. Acad. Sci. USA 2005, 102, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Tronnersjo, S.; Hanefalk, C.; Balciunas, D.; Hu, G.Z.; Nordberg, N.; Muren, E.; Ronne, H. The jmjN and jmjC domains of the yeast zinc finger protein Gis1 interact with 19 proteins involved in transcription, sumoylation and DNA repair. Mol. Genet. Genomics 2007, 277, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hon, T.; Dodd, A.; Dirmeier, R.; Gorman, N.; Sinclair, P.R.; Zhang, L.; Poyton, R.O. A mechanism of oxygen sensing in yeast. Multiple oxygen-responsive steps in the heme biosynthetic pathway affect Hap1 activity. J. Biol. Chem. 2003, 278, 50771–50780. [Google Scholar] [CrossRef] [PubMed]
- Balciunas, D.; Ronne, H. Yeast genes GIS1-4: Multicopy suppressors of the Gal- phenotype of snf1 mig1 srb8/10/11 cells. Mol. Gen. Genet. 1999, 262, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Mehta, M.; Kuo, D.; Sung, M.K.; Chuang, R.; Jaehnig, E.J.; Bodenmiller, B.; Licon, K.; Copeland, W.; Shales, M.; et al. Rewiring of genetic networks in response to DNA damage. Science 2010, 330, 1385–1389. [Google Scholar] [CrossRef]
- Costanzo, M.; Baryshnikova, A.; Bellay, J.; Kim, Y.; Spear, E.D.; Sevier, C.S.; Ding, H.; Koh, J.L.; Toufighi, K.; Mostafavi, S.; et al. The genetic landscape of a cell. Science 2010, 327, 425–431. [Google Scholar] [CrossRef]
- Costanzo, M.; VanderSluis, B.; Koch, E.N.; Baryshnikova, A.; Pons, C.; Tan, G.; Wang, W.; Usaj, M.; Hanchard, J.; Lee, S.D.; et al. A global genetic interaction network maps a wiring diagram of cellular function. Science 2016, 353, aaf1420. [Google Scholar] [CrossRef]
- Kastaniotis, A.J.; Zitomer, R.S. Rox1 mediated repression. Oxygen dependent repression in yeast. Adv. Exp. Med. Biol. 2000, 475, 185–195. [Google Scholar]
- Lai, L.C.; Kosorukoff, A.L.; Burke, P.V.; Kwast, K.E. Metabolic-state-dependent remodeling of the transcriptome in response to anoxia and subsequent reoxygenation in Saccharomyces cerevisiae. Eukaryot Cell 2006, 5, 1468–1489. [Google Scholar] [CrossRef]
- Krupp, M.; Marquardt, J.U.; Sahin, U.; Galle, P.R.; Castle, J.; Teufel, A. RNA-Seq Atlas--a reference database for gene expression profiling in normal tissue by next-generation sequencing. Bioinformatics 2012, 28, 1184–1185. [Google Scholar] [CrossRef] [PubMed]
- Berry, W.L.; Janknecht, R. KDM4/JMJD2 histone demethylases: Epigenetic regulators in cancer cells. Cancer Res. 2013, 73, 2936–2942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Chen, H.Z.; Wang, L.; Liu, D.P.; Hill, J.A.; Liu, Z.P. The histone trimethyllysine demethylase JMJD2A promotes cardiac hypertrophy in response to hypertrophic stimuli in mice. J. Clin. Invest. 2011, 121, 2447–2456. [Google Scholar] [CrossRef] [PubMed]
- Young, L.C.; Hendzel, M.J. The oncogenic potential of Jumonji D2 (JMJD2/KDM4) histone demethylase overexpression. Biochem. Cell Biol. 2013, 91, 369–377. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, G.W.; Jeon, Y.H.; Yoo, J.; Lee, S.W.; Kwon, S.H. Advances in histone demethylase KDM4 as cancer therapeutic targets. FASEB J. 2020, 34, 3461–3484. [Google Scholar] [CrossRef]
- Bishop, D.F.; Anderson, K.E.; Desnick, R.J.; Sassa, S. Disorders of heme biosynthesis: X-linked sideroblastic anemia and the porphyrias. In The Online Metabolic and Molecular Bases of Inherited Disease; The McGraw-Hill Companies, Inc.: New York, NY, USA, 2016; pp. 2991–3062. [Google Scholar]
- Hooda, J.; Shah, A.; Zhang, L. Heme, an essential nutrient from dietary proteins, critically impacts diverse physiological and pathological processes. Nutrients 2014, 6, 1080–1102. [Google Scholar] [CrossRef]
- Mattoon, J.; Lancashire, W.; Sanders, H.; Carvajal, E.; Malamud, D.; Braz, G.; Panek, A. Oxygen and catabolite regulation of hemoprotein biosynthesis in the yeast Saccharomyces cerevisiae. In Biosynthesis of Heme and Cholorophylls; Caughey, W.J., Ed.; Academic Press: Cambridge, MA, USA, 1979; pp. 421–435. [Google Scholar]
- Zheng, B.; Albrecht, U.; Kaasik, K.; Sage, M.; Lu, W.; Vaishnav, S.; Li, Q.; Sun, Z.S.; Eichele, G.; Bradley, A.; et al. Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock. Cell 2001, 105, 683–694. [Google Scholar] [CrossRef]
- Burch, J.S.; Marcero, J.R.; Maschek, J.A.; Cox, J.E.; Jackson, L.K.; Medlock, A.E.; Phillips, J.D.; Dailey, H.A., Jr. Glutamine via alpha-ketoglutarate dehydrogenase provides succinyl-CoA for heme synthesis during erythropoiesis. Blood 2018, 132, 987–998. [Google Scholar] [CrossRef]
- Leurs, U.; Clausen, R.P.; Kristensen, J.L.; Lohse, B. Inhibitor scaffold for the histone lysine demethylase KDM4C (JMJD2C). Bioorg Med. Chem. Lett. 2012, 22, 5811–5813. [Google Scholar] [CrossRef]
- Metzger, E.; Stepputtis, S.S.; Strietz, J.; Preca, B.T.; Urban, S.; Willmann, D.; Allen, A.; Zenk, F.; Iovino, N.; Bronsert, P.; et al. KDM4 Inhibition Targets Breast Cancer Stem-like Cells. Cancer Res. 2017, 77, 5900–5912. [Google Scholar] [CrossRef]
- Tan, M.K.; Lim, H.J.; Harper, J.W. SCF(FBXO22) regulates histone H3 lysine 9 and 36 methylation levels by targeting histone demethylase KDM4A for ubiquitin-mediated proteasomal degradation. Mol. Cell Biol. 2011, 31, 3687–3699. [Google Scholar] [CrossRef] [PubMed]
- Ipenberg, I.; Guttmann-Raviv, N.; Khoury, H.P.; Kupershmit, I.; Ayoub, N. Heat shock protein 90 (Hsp90) selectively regulates the stability of KDM4B/JMJD2B histone demethylase. J. Biol. Chem. 2013, 288, 14681–14687. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Vasilatos, S.N.; Bhargava, R.; Fine, J.L.; Oesterreich, S.; Davidson, N.E.; Huang, Y. Functional interaction of histone deacetylase 5 (HDAC5) and lysine-specific demethylase 1 (LSD1) promotes breast cancer progression. Oncogene 2017, 36, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.; Seo, J.; Lee, H.S.; Stanton, L.W.; Kim, D.; Choi, J.K. Global mapping of the regulatory interactions of histone residues. FEBS Lett. 2015, 589, 4061–4070. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | ORF Name | Protein ID (gi #) | Description | Human Orthologues | HCB 1 | Mtb 2 | Ubl 3 | Acet 4 |
|---|---|---|---|---|---|---|---|---|
| ACT1 | YFL039C | 14318479 | Structural protein involved in cell polarization | ACTA1, ACTA2, ACTB, ACTBL2, ACTC1, ACTG1, ACTG2, ACTR1A, ACTR1B, ACTRT1, ACTRT2, ACTRT3, POTEE, POTEKP | + | + | + | |
| AHA1 | YDR214W | 398366155 | Activator of heat-shock protein 90 ATPase | AHSA1 | ||||
| AIM6 | YDL237W | 330443495 | Altered inheritance of mitochondria protein | − | ||||
| ARC40 | YBR234C | 6319711 | Subunit of the ARP2/3 complex | ARPC1A, ARPC1B | ||||
| BCK1 | YJL095W | 6322366 | Serine/threonine protein kinase | MAP3K1, MAP3K2, MAP3K3, NRBP1, NRBP2, WNK1, WNK2, WNK3, WNK4 | + | |||
| CAF20 | YOR276W | 398366043 | Cap-associated phosphoprotein of the mRNA cap-binding complex | - | + | |||
| CDC33 | YOL139C | 6324433 | mRNA cap binding protein and translation initiation factor | EIF4E, EIF4E2, EIF4E3 | + | + | ||
| CKS1 | YBR135W | 398364999 | Cyclin-dependent protein kinase regulatory subunit and adaptor | CKS1B, CKS2 | ||||
| CPR1 | YDR155C | 6320359 | Cytoplasmic peptidyl-prolyl cis–trans isomerase | PPIA, PPIAL4E, PPIE, PPIF, RANBP2, RGPD1, RGPD3, RGPD4, RGPD5, RGPD6 | + | + | + | |
| CYR1 | YJL005W | 398364701 | Adenylate cyclase | LRCH1, LRR1, LRRC59, LRRK1, PHLPP1, PHLPP2 | + | + | ||
| DIG1 | YPL049C | 6325208 | Downregulator of invasive growth | − | + | |||
| DOT6 | YER088C | 398364577 | Disruptor of telomeric silencing | − | + | |||
| ERG6 | YML008C | 6323635 | Delta(24)-sterol C-methyltransferase | COQ3, WBSCR27 | + | + | ||
| ESS1 | YJR017C | 37362669 | Peptidyl-prolyl cis–trans isomerase | PIN1 | ||||
| GOR1 | YNL274C | 6325144 | Glyoxylate reductase | GRHPR | + | + | ||
| GPD2 | YOL059W | 6324513 | Mitochondrial glycerol-3-phosphate dehydrogenase (NAD(+)) 2 | GPD1 | + | + | ||
| GRX2 | YDR513W | 6320720 | Mitochondrial glutaredoxin-2 | GLRX, GLRX2 | ||||
| GYP1 | YOR070C | 6324644 | Cis-golgi GTPase-activating protein (GAP) for yeast Rabs | − | ||||
| HIT1 | YJR055W | 6322515 | Protein involved in C/D snoRNP assembly | ZNHIT3 | ||||
| HPA3 | YEL066W | 6320768 | D-amino-acid N-acetyltransferase | − | + | |||
| HTB2 | YBL002W | 6319471 | Core histone protein required for chromatin assembly and chromosome function | H2BFWT, HIST1H2BA, HIST1H2BB, HIST1H2BC, HIST1H2BD, HIST1H2BE, HIST1H2BF, HIST1H2BG, HIST1H2BH, HIST1H2BI, HIST1H2BJ, HIST1H2BK, HIST1H2BL, HIST1H2BM, HIST1H2BN, HIST1H2BO, HIST2H2BE, HIST3H2BB | + | + | + | |
| IMP2 | YMR035-W | 398364225 | Catalytic subunit of mitochondrial inner membrane peptidase complex | IMMP2L | ||||
| KCS1 | YDR017C | 398365025 | Inositol hexakisphosphate and inositol heptakisphosphate kinase | IP6K1, IP6K2, IP6K3, IPMK | + | + | ||
| MHP1 | YJL042W | 6322419 | MAP-homologous protein | CARD8, CIITA, NLRC3, NLRC5, NLRP1, NLRP10, NLRP11, NLRP12, NLRP13, NLRP14, NLRP2, NLRP3, NLRP4, NLRP5, NLRP6, NLRP7, NLRP9, NLRX1, NOD1, NOD2 | + | + | ||
| MIG1 | YGL035C | 6321403 | Transcription factor involved in glucose repression | EGR1, EGR2, EGR3, EGR4, WT1 | + | |||
| MOT2 | YER068W | 398364501 | General negative regulator of transcription subunit | CNOT4 | + | + | ||
| MRM1 | YOR201C | 398365713 | Mitochondrial rRNA methyltransferase | − | + | |||
| MRPL32 | YCR003W | 6319848 | Mitochondrial ribosomal protein of the large subunit | MRPL32 | ||||
| MRPS12 | YNR036C | 6324364 | Mitochondrial ribosomal protein of the small subunit | MRPS12 | ||||
| MSC3 | YLR219W | 6323248 | Meiotic sister chromatid recombination protein | − | ||||
| NBA1 | YOL070C | 6324502 | Protein of unknown function | − | ||||
| NUP60 | YAR002W | 6319318 | Component of central core of the nuclear pore complex | − | ||||
| OSH3 | YHR073W | 6321864 | Member of an oxysterol-binding protein family | OSBPL3, OSBPL6, OSBPL7 | ||||
| OYE2 | YHR179W | 6321973 | NADPH dehydrogenase | − | + | |||
| PBP4 | YDL053C | 6320150 | Pbp1p-binding protein | − | ||||
| PDI1 | YCL043C | 6319806 | Protein disulfide isomerase | ERP27, ERP44, P4HB, PDIA2, PDIA3, TMX3 | ||||
| PGA2 | YNL149C | 6324180 | Essential protein required for maturation of Gas1p and Pho8p | − | + | |||
| PGI1 | YBR196C | 6319673 | Glycolytic enzyme phosphoglucose isomerase | GPI | + | + | ||
| PGK1 | YCR012W | 10383781 | 3-phosphoglycerate kinase | PGK1, PGK2 | + | + | + | + |
| PHO8 | YDR481C | 398366635 | Repressible vacuolar alkaline phosphatase | ALPI, ALPL, ALPP, ALPPL2 | + | |||
| POL4 | YCR014C | 7839197 | DNA polymerase IV | DNTT, POLL, POLM | + | |||
| PSP2 | YML017W | 41629688 | Asn-rich cytoplasmic protein containing RGG motifs | − | + | |||
| PTC4 | YBR125C | 6319601 | Cytoplasmic type 2C protein phosphatase (PP2C) | PPM1D, PPM1G | ||||
| RPS18A | YDR450W | 6320658 | Protein component of the small (40S) ribosomal subunit | RPS18 | + | |||
| SIP1 | YDR422C | 398366583 | Alternate beta-subunit of the Snf1p kinase complex | − | ||||
| SNX41 | YDR425W | 6320633 | Sorting nexin | SNX4 | ||||
| SPC42 | YKL042W | 398364591 | Central plaque component of spindle pole body | − | ||||
| SSD1 | YDR293C | 6320499 | Translational repressor with a role in polar growth and wall integrity | DIS3L2 | + | + | ||
| TMC1 | YOR052C | 6324626 | AN1-type zinc finger protein, effector of proteotoxic stress response | − | + | |||
| TOD6 | YBL054W | 6319417 | Twin of dot6p | − | + | |||
| TOS1 | YBR162C | 6319638 | Covalently bound cell wall protein of unknown function | − | ||||
| UTR1 | YJR049C | 6322509 | ATP–NADH kinase | NADK | + | + | ||
| YBL081W | YBL081W | 6319390 | Uncharacterized protein | − | ||||
| YCL020W | YCL020W | 10383771 | Retrotransposon TYA Gag gene co-transcribed with TYB Pol | − | + | |||
| YDR365-WB | YDR365WB | 7839164 | Retrotransposon TYA Gag and TYB Pol genes | − | ||||
| YPK2 | YMR104C | 6323751 | Serine/threonine protein kinase | AKT1, AKT2, AKT3, PRKCA, PRKCB, PRKCD, PRKCE, PRKCG, PRKCH, PRKCI, PRKCQ, PRKCZ, RPS6KB1, RPS6KB2, SGK1, SGK2, SGK3 | + | + | ||
| YPL247C | YPL247C | 6325009 | WD repeat-containing protein | |||||
| ZDS1 | YMR273C | 6323929 | Protein with a role in regulating Swe1p-dependent polarized growth | − |
| Common Name | ORF Name | Protein ID (gi #) | Description | Human Orthologues | HCB 1 | Mtb 2 | Ubl 3 | Acet 4 |
|---|---|---|---|---|---|---|---|---|
| ADE1 | YAR015W | 6319326 | Phosphoribosylaminoimidazole-succinocarboxamide synthase | PAICS | + | + | + | |
| ADH3 | YMR083-W | 6323729 | Mitochondrial alcohol dehydrogenase isozyme III | SORD, TP53I3 | + | |||
| AGE1 | YDR524C | 398366675 | ADP-ribosylation factor (ARF) GTPase activating protein (GAP) effector | ACAP1, ACAP2, AGAP1, AGAP2, ARAP1, ARAP2, ARAP3, ASAP1, ASAP2, ASAP3 | ||||
| ALT1 | YLR089C | 6323118 | Alanine transaminase (glutamic pyruvic transaminase) | CCBL1, CCBL2, GPT, GPT2 | + | + | ||
| ATG18 | YFR021W | 16740527 | Autophagy-related protein | WIPI1, WIPI2 | ||||
| AYR1 | YIL124W | 398364253 | Bifunctional triacylglycerol lipase and 1-acyl DHAP reductase | DECR1, DHRS7, DHRS7B, DHRS7C, HSD11B1, HSD17B2 | ||||
| BOI1 | YBL085W | 6319386 | Protein implicated in polar growth | NEB, NEBL, PSD, PSD4, SH3RF2 | ||||
| BRE2 | YLR015W | 6323043 | Subunit of COMPASS (Set1C) complex | ASH2L | + | |||
| BUD5 | YCR038C | 10383799 | Bud site selection protein | RAPGEF1, SOS1, SOS2 | ||||
| COA6 | YMR244C-A | 6323902 | Protein involved in cytochrome c oxidase (Complex IV) assembly | COA6 | ||||
| CPA1 | YOR303W | 398366173 | Carbamoyl-phosphate synthase arginine-specific small chain | CAD, CPS1, OTC | + | + | ||
| ERG26 | YGL001C | 6321437 | Sterol-4-alpha-carboxylate 3-dehydrogenase | GMDS, HSD3B1, HSD3B2, HSD3B7, NSDHL | + | |||
| ERV41 | YML067C | 6323573 | Protein localized to COPII-coated vesicles | ERGIC2 | ||||
| GIS2 | YNL255C | 6324074 | Translational activator for mRNAs with internal ribosome entry sites | CNBP | + | |||
| GVP36 | YIL041W | 398364435 | BAR domain protein | TAF7, TAF7L, FECH | + | + | ||
| HEM15 | YOR176W | 398365579 | Ferrochelatase | FECH | + | |||
| HFD1 | YMR110C | 6323757 | Fatty aldehyde dehydrogenase | ALDH3A1, ALDH3A2, ALDH3B1, ALDH3B2 | + | |||
| HSP42 | YDR171W | 6320376 | Small heat-shock protein (sHSP) with chaperone activity | − | ||||
| INA22 | YIR024C | 6322215 | Inner membrane assembly complex subunit 22 | − | ||||
| IPP1 | YBR011C | 6319483 | Cytoplasmic inorganic pyrophosphatase | PPA1, PPA2 | + | |||
| IRC22 | YEL001C | 6320836 | Increased recombination centers protein | − | ||||
| IRC5 | YFR038W | 42742173 | Uncharacterized ATP-dependent helicase | HELLS, SMARCAL1 | + | |||
| LYS12 | YIL094C | 6322097 | Mitochondrial homoisocitrate dehydrogenase | IDH3A | + | + | ||
| LYS20 | YDL182W | 6320019 | Homocitrate synthase isozyme | HMGCL | + | |||
| LYS21 | YDL131W | 6320071 | Mitochondrial homocitrate synthase | HMGCL | + | |||
| MIG3 | YER028C | 6320866 | Transcription corepressor | EGR1, EGR2, EGR3, EGR4, WT1 | + | |||
| MLF3 | YNL074C | 398365051 | Serine-rich protein of unknown function | − | ||||
| MPE1 | YKL059C | 6322791 | Essential conserved subunit of CPF cleavage and polyadenylation factor | RBBP6 | + | |||
| MSC1 | YML128C | 6323507 | Protein of unknown function | − | ||||
| NAM9 | YNL137C | 398364685 | 37S ribosomal protein | − | + | |||
| NPA3 | YJR072C | 398365155 | GPN-loop GTPase | GPN1 | + | + | ||
| OSM1 | YJR051W | 6322511 | Fumarate reductase | SDHA | ||||
| PHB2 | YGR231C | 50593217 | Prohibitin | PHB2 | ||||
| PNO1 | YOR145C | 6324720 | Pre-rRNA-processing protein | − | + | |||
| PUB1 | YNL016W | 6324312 | Nuclear and cytoplasmic polyadenylated RNA-binding protein | RBM42, TIA1 | + | + | ||
| RNH1 | YMR234W | 6323890 | Ribonuclease | RNASEH1 | ||||
| RPC40 | YPR110C | 6325367 | RNA polymerase subunit AC40 | POLR1C | + | + | ||
| RRP3 | YHR065C | 37362659 | ATP-dependent rRNA helicase | DDX28, DDX47 | + | |||
| RSA1 | YPL193W | 6325063 | Protein involved in the assembly of 60S ribosomal subunits | NUFIP1 | + | |||
| RTN1 | YDR233C | 398366209 | Reticulon-like protein 1 | RTN1, RTN2, RTN3, RTN4 | ||||
| RVS167 | YDR388W | 6320596 | Calmodulin-binding actin-associated protein | − | + | + | ||
| SCS2 | YER120W | 398364741 | Integral ER membrane protein | MOSPD3, VAPA, VAPB | + | |||
| SDS22 | YKL193C | 6322655 | Regulatory subunit of the type 1 protein phosphatase | CD180, CNTRL, DNAAF1, LRRC32, NRROS, PPP1R7, TLR2, TLR3, TLR4, TLR5, TLR7 | ||||
| SHE1 | YBL031W | 6319440 | Mitotic spindle protein | − | ||||
| SIP2 | YGL208W | 6321230 | One of three beta subunits of the Snf1 kinase complex | PRKAB1, PRKAB2 | + | |||
| SNF4 | YGL115W | 6321323 | Activating gamma subunit of the AMP-activated Snf1p kinase complex | PRKAG1, PRKAG2, PRKAG3 | + | + | ||
| SOK1 | YDR006C | 398364967 | Protein of unknown function | TCP11 | ||||
| SRB4 | YER022W | 6320860 | Subunit of the RNA polymerase II mediator complex | MED17 | + | |||
| SSO1 | YPL232W | 6325024 | Plasma membrane t-SNARE | STX11, STX12, STX17, STX1A, STX1B, STX2, STX3, STX4, STX7 | ||||
| SUA7 | YPR086W | 6325343 | Transcription factor TFIIB | GTF2B | ||||
| TDH1 | YJL052W | 398364523 | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH, GAPDHS | + | + | ||
| VMA13 | YPR036W | 6325293 | Subunit H of the V1 peripheral membrane domain of V-ATPase | ATP6V1H | ||||
| YBL086C | YBL086C | 6319385 | Uncharacterized protein | FAM102A | ||||
| YCR087C-A | YCR087C-A | 6319930 | Putative protein of unknown function | − | + |
| Common Name | ORF Name | Protein ID (gi#) | Description | Human Orthologues | HCB 1 | Mtb 2 | Ubl 3 | Acet 4 |
|---|---|---|---|---|---|---|---|---|
| ATP1 | YBL099W | 330443397 | Alpha subunit of the F1 sector of mitochondrial F1F0 ATP synthase | ATP5A1, ATP5B | + | + | ||
| BEM2 | YER155C | 398364959 | GTPase-activating protein | ARHGAP10, ARHGAP12, ARHGAP15, ARHGAP21, ARHGAP23, ARHGAP26, ARHGAP27, ARHGAP35, ARHGAP42, ARHGAP9, CHN1, CHN2, GMIP, HMHA1, OPHN1, RACGAP1 | + | + | ||
| DBP5 | YOR046C | 6324620 | Cytoplasmic ATP-dependent RNA helicase of the DEAD-box family | DDX19B, DDX25 | + | |||
| DLD3 | YEL071W | 6320764 | D-2-hydroxyglutarate--pyruvate transhydrogenase | AGPS, D2HGDH, LDHD | + | + | + | |
| DUF1 | YOL087C | 6324485 | Uncharacterized WD repeat-containing protein | WDR48 | ||||
| ERG11 | YHR007C | 6321795 | Lanosterol 14-alpha-demethylase | CYP51A1 | + | + | + | |
| GAL83 | YER027C | 6320865 | One of three possible beta-subunits of the Snf1 kinase complex | PRKAB1, PRKAB2 | + | |||
| GUA1 | YMR217W | 6323873 | GMP synthase | GMPS | + | + | + | |
| HBT1 | YDL223C | 6319978 | Shmoo tip protein | − | ||||
| HRK1 | YOR267C | 6324841 | Serine/threonine protein kinase | HUNK, PRKAA2 | + | |||
| ILV1 | YER086W | 6320930 | Mitochondrial threonine dehydratase | − | ||||
| IST2 | YBR086C | 27808701 | Increased sodium tolerance protein | ANO1, ANO2, ANO3, ANO4, ANO5, ANO6, ANO7 | ||||
| MOT3 | YMR070W | 6323715 | Transcriptional activator/repressor | SMARCA2, SMARCA4 | + | + | ||
| NMA1 | YLR328W | 6323360 | Nicotinic acid mononucleotide adenylyltransferase | NMNAT1, NMNAT2, NMNAT3 | + | + | ||
| NMA2 | YGR010W | 6321447 | Nicotinic acid mononucleotide adenylyltransferase | NMNAT1, NMNAT2, NMNAT3 | + | + | ||
| NOG2 | YNR053C | 6324381 | Nucleolar GTP-binding protein | GNL2 | + | |||
| OLE1 | YGL055W | 398364717 | Acyl-CoA desaturase | SCD, SCD5 | + | + | ||
| OPY1 | YBR129C | 6319605 | Protein of unknown function | PLEK, PLEK2 | ||||
| PAL2 | YHR097C | 6321889 | pH-response regulator protein | − | ||||
| PBP1 | YGR178C | 398366039 | PAB1-binding protein | ATXN2, ATXN2L | + | + | ||
| RPT1 | YKL145W | 6322704 | ATPase of the 19S regulatory particle of the 26S proteasome | PSMC2 | + | |||
| RTK1 | YDL025C | 6320179 | Probable serine/threonine protein kinase | HUNK, PRKAA2 | + | + | ||
| RVB2 | YPL235W | 6325021 | RuvB-like protein | RUVBL2 | + | |||
| SNF1 | YDR477W | 398366631 | AMP-activated S/T protein kinase | HUNK, MELK, PRKAA1, PRKAA2, STK40, TRIB1, TRIB2, TRIB3 | + | + | + | |
| SOF1 | YLL011W | 6323018 | Protein required for biogenesis of 40S (small) ribosomal subunit | DCAF13 | + | |||
| SRO9 | YCL037C | 37362625 | Cytoplasmic RNA-binding protein | LARP1, LARP4B | + | + | ||
| SRV2 | YNL138W | 6324191 | CAP (cyclase-associated protein) | − | ||||
| TUB1 | YML085C | 6323554 | Tubulin alpha-1 chain | TUBA8 | + | |||
| TUB2 | YFL037W | 14318481 | Beta-tubulin, associates with alpha-tubulin | TUBB, TUBB1, TUBB2A, TUBB2B, TUBB3, TUBB4A, TUBB4B, TUBB6, TUBE1 | + | |||
| UGP1 | YKL035W | 398364619 | UTP--glucose-1-phosphate uridylyltransferase | UAP1, UGP2 | + | + | ||
| VTS1 | YOR359W | 398366369 | Flap-structured DNA-binding and RNA-binding protein | SAMD4A | + | |||
| WHI2 | YOR043W | 6324617 | Growth regulation protein | − | ||||
| YBR238C | YBR238C | 6319715 | Mitochondrial membrane protein | − | + | |||
| YCK2 | YNL154C | 6324175 | Palmitoylated plasma membrane-bound casein kinase I (CK1) isoform | CSNK1G2 | + | + | + | |
| YGR237C | YGR237C | 6321676 | Uncharacterized protein | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konduri, P.C.; Wang, T.; Salamat, N.; Zhang, L. Heme, A Metabolic Sensor, Directly Regulates the Activity of the KDM4 Histone Demethylase Family and Their Interactions with Partner Proteins. Cells 2020, 9, 773. https://doi.org/10.3390/cells9030773
Konduri PC, Wang T, Salamat N, Zhang L. Heme, A Metabolic Sensor, Directly Regulates the Activity of the KDM4 Histone Demethylase Family and Their Interactions with Partner Proteins. Cells. 2020; 9(3):773. https://doi.org/10.3390/cells9030773
Chicago/Turabian StyleKonduri, Purna Chaitanya, Tianyuan Wang, Narges Salamat, and Li Zhang. 2020. "Heme, A Metabolic Sensor, Directly Regulates the Activity of the KDM4 Histone Demethylase Family and Their Interactions with Partner Proteins" Cells 9, no. 3: 773. https://doi.org/10.3390/cells9030773
APA StyleKonduri, P. C., Wang, T., Salamat, N., & Zhang, L. (2020). Heme, A Metabolic Sensor, Directly Regulates the Activity of the KDM4 Histone Demethylase Family and Their Interactions with Partner Proteins. Cells, 9(3), 773. https://doi.org/10.3390/cells9030773