Abstract
Several recent studies have revealed previously unknown complexity of the amphibian interferon (IFN) system. Being unique in vertebrate animals, amphibians not only conserve and multiply the fish-like intron-containing IFN genes, but also rapidly evolve amniote-like intronless IFN genes in each tested species. We postulate that the amphibian IFN system confers an essential model to study vertebrate immune evolution in molecular and functional diversity to cope with unprecedented pathophysiological requirement during terrestrial adaption. Studies so far have ascribed a potential role of these IFNs in immune regulation against intracellular pathogens, particularly viruses; however, many knowledge gaps remain elusive. Based on recent reports about IFN’s multifunctional properties in regulation of animal physiological and defense responses, we interpret that amphibian IFNs may evolve novel function pertinent to their superior molecular diversity. Such new function revealed by the emerging studies about antifungal and developmental regulation of amphibian IFNs will certainly promote our understanding of immune evolution in vertebrates to address current pathogenic threats causing amphibian decline.
1. The Unique Complexity and Role of Amphibian Interferons (IFNs) in Immune Evolution
The animal immune system relies on a series of specified immune molecules, collectively known as immunome, to detect and repel pathogenic invaders at the molecular, cellular, and organismal levels. Species-specific or cross-species characterization of the immunome lays a cornerstone in studying animal health and immune evolution [1,2,3]. Interferon (IFN)-mediated immune response involves IFNs and several hundreds of IFN-stimulated genes (ISGs) that play a pivotal role in antiviral defense [4,5,6,7]. Coincided with the evolution of adaptive immunity, IFN genes also originate in jawed fish from a few multi-exon progenitors [8]. Primitive IFN-like molecules have been previously determined in bony fish, with discernible IFN-γ- and IFN-ϕ-like genes, which are presumed to be progenitors of type II (IFN-II, IFN-γ) and type I IFNs (IFN-I), respectively, in tetrapods [9,10,11]. A new study discovered ancestral IFN orthologs of all three IFN types in a cartilaginous fish, and hence resolved that three types of IFNs were ramified before the bony fish [12]. We annotated IFN gene families across 110 vertebrate genomes, and showed that IFN genes, after originating in jawed fishes, had several significant evolutionary surges regarding the numbers of IFN-coding genes and subtypes in some species of amphibians, bats, and ungulates (particularly pigs and cattle of livestock) [13,14,15]. Here, we review the newly identified IFN molecular evolution and expansion in amphibians [12,13,16].
The evolution of intronless (or single-exon) IFN genes and further molecular diversification in amniotes are signature events in IFN evolution [11,12,13,14,15,16]. However, our understanding of this process is elusive, partly due to the unrecognized role of amphibians in IFN evolution [9,10,11]. Previous studies have: (1) Only discovered several intron-containing IFN progenitors in amphibians [9], and (2) underestimated IFN-mediated antimicrobial responses in amphibians and the unique role of amphibians in IFN evolution. Previously, the emergence of intronless IFN genes has been assumedly ascribed to reptiles, which further suggested a linear-increasing model for IFN diversification from fish to mammals [9,10]. Our recent research detected the unexpected emergence and expansion of intronless IFN genes in amphibians, exemplified by 40–50 predicated IFN-coding genes, of which 24–37 are intronless in each Xenopus species [13]. This remarkable coexistence of intron-containing and intronless IFNs in amphibians confirms that the emergence of intronless IFN genes actually occurred in amphibians (or even earlier in fish) in contrast to previously hypothesized in reptiles [13,16]. These findings determined previously unknown and species-independent expansion of intronless IFN genes in amphibians, which revises the established model of IFN evolution and highlights the critical role of amphibian IFN complex in studying IFN biology (Figure 1) [11,12,13,14,15,16].
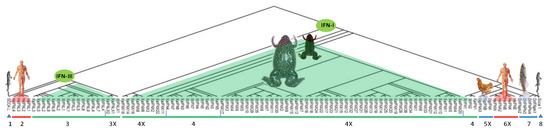
Figure 1.
Schematic of vertebrate interferon (IFN) evolution and recent discovery about amphibian IFN complex. Evolutionary analyses were conducted in MEGA X [17]. The minimum evolution (ME) method was used to infer the molecular phylogeny, and the evolutionary distances were computed using the p-distance method, shown in the units of the number of amino acid differences per site. The ME tree was searched (search level of 1) using the close-neighbor-interchange (CNI) algorithm. The analysis used 108 amino acid sequences. All ambiguous positions were removed for each sequence pair. There was a total of 437 positions in the final dataset. In contrast to the previously known several fish-like intron-containing amphibian IFNs, recent studies revealed that amphibians are unique to have both fish-like intron-containing and amniotic intronless IFNs, which molecularly and immunologically stamp their amphibiotic position in vertebrate evolution [12,13,16]. Legends and Abbreviations: IFN-I, type I IFNs (including amniotic intronless IFNA, IFNB, IFNE, IFNK, IFNW, respectively, for the genes of IFN-α, -β, -ε, -κ, -ω; and diverse amphibian intron-containing, XaIFN or XtIFN, and intronless XaIFNX or XtIFNX, listed here); IFN-III, type III IFNs (including amniotic intron-containing IFNL gene for IFN-λ; and amphibian intron-containing XaIFNL or XtIFNL, and intronless XaIFNLX or XtIFNLX, listed here); SCCA 1, Scyliorhinus canicula (catshark) ancestral IFN-I; SCCA L, Scyliorhinus canicula (catshark) ancestral IFN-III; Dr, Danio rerio (zebrafish); Gg, Gallus gallus (chicken); Hs, Homo sapiens; Xa, Xenopus laevis; Xt, Xenopus tropicalis; Group designation: 1 and 7 & 8, intron-containing ancestral IFN-I (7 & 8) and IFN-III (1); 2, intron-containing IFN-III in amniotes; 3 and 4, amphibian intron-containing IFN-III (3) and IFN-I(4); 3X and 4X, intronless amphibian IFN-III (3X) and IFN-I (4X); 5X and 6X, intronless IFN-I in amniotes.
For instance, extensive analysis of current genome assemblies of Xenopus tropicalis and Xenopus laevis at NCBI (https://www.ncbi.nlm.nih.gov/genome/?term=Xenopus) and Xenobase (http://www.xenbase.org/) led to the identification of 37 intronless IFN-like genes (XtIFNX) (including 4 near intronless IFN-like genes retaining only a short (<50 bp) intron) and 3 pseudogenes in X. tropicalis [13]. Similarly, 26 intronless IFN-I (XaIFNX) genes (including 8 with a short remaining intron and 4 pseudogenes) were identified in the X. laevis genome. Notably, those amphibian short-intron-containing IFN gene-like sequences were indicated to be rudimentary intermediates of the retro(trans)position event that further became intronless IFN genes. Our annotation of the current genome assemblies at NCBI and Xenbase not only determined intronless IFNs in frogs for the first time, but also discovered several more intron-containing IFNs, which were not revealed in previous studies. We therefore updated the current repertoire of IFN gene loci in X. tropicalis: The co-existence of 14 intron-containing IFNs (7 IFN-I, 1 IFN-II, and 6 IFN-III) and the expansion of 37 intronless IFN-like genes (36 IFN-I and 1 IFN-III); and in X. laevis: 17 intron-containing (7 IFN-I, 1 IFN-II, and 9 IFN-III) and 24 intronless IFN-coding genes (22 IFN-I and 2 IFN-III) (Table 1 and Figure 1) [13]. A new study in a Tibet frog species (Nanorana parkeri) reinforced the co-existence of both intron-containing and intronless IFN genes in amphibians [16]. However, the expansion of antiviral IFN genes in different amphibian species, especially the intronless ones, seems species-independent. These intronless amphibian IFN genes seem to have a paralogous rather than an orthologous relationship, as well as to the amniote intronless IFNs [12,13,16]. All of these studies collectively determined the unique coexistence of intron-containing and intronless IFNs in both IFN-I and IFN-III in amphibians [12,13,14,15,16]. These findings support that the retroposition process, which initiated the evolution and expansion of intronless IFNs in higher amniotes, evidently originated in amphibians or earlier, rather than in reptiles as previously believed [9,10,11,12,13,16]. The species-independent expansion of intronless type I IFNs in amphibians suggests a unique role of amphibians in IFN evolution. Indeed, collected evidence also indicates an evolutionary benefit of these intronless IFN genes in gene expression and efficient contest to infectious and developmental pressures during adaption to terrestrial lives [11,12,13,14,16,18,19,20].

Table 1.
Molecular composition of amphibian IFN complex compared with IFNs in amniotes (human) and fish (catshark).
Amphibians are an irreplaceable link in animal evolution toward terrestrial tetrapods. They reside in aquatic, terrestrial, and arboreal habitats, and are sensitive indicators for monitoring the environmental and habitat changes in our bio-ecosystem. In basic science research, amphibians have long been utilized as prominent models for studying physiological and developmental processes [21,22,23,24]. Due to the crucial role that amphibians play in both the ecosystem and experimental sciences, it is imperative to gain a better understanding of their immunobiology. Recent investigations have indicated that pathogenic factors (such as by chytrid fungus and Ranavirus) are causing amphibian population decline and threaten global biodiversity [25,26,27,28,29,30,31]. This further emphasizes the need to study the amphibian immunome and its interaction with epidemic pathogens [1,2,3,32,33,34,35,36,37,38,39,40]. Amphibians have several unusual characteristics that are stimulating for immunobiological studies [32,33,35,39]. For example, amphibians have moist and permeable skin, which also serves as an organ for respiration [33,34]. Previous studies have discovered that amphibian skin is one of the most generous sources of antimicrobial peptides, of which some are broadly effective and potentially therapeutic against bacteria, fungi, viruses, and even cancers [37,38,41,42,43,44,45]. Recent studies in the amphibian IFN system uncovered another unique advantage—that amphibians give rise to the emergence and expansion of intronless IFN genes [12,13,16], which archive the primitive diversity of the IFN system prior to further evolve in higher vertebrates, including humans [1,12]. Therefore, not only are amphibians unique for the study of the signature event of IFN evolution that leads to antiviral superiority of intronless IFNs in amniotes, but amphibian models are also irreplaceable in order to determine the evolutionary coordination and functional diversity among the IFN subgroups [12,13,14,15,16]. This is because amphibians represent the only animal clade to have both IFN gene subgroups (intronless vs. intron-containing) in either IFN-I or IFN-III types (Table 1 and Figure 1) [12,13,14,15,16].
2. The Fish-Like Intron-Containing IFN Genes in Amphibians
Two types of fish IFN genes, generally one to several IFN-γ and several IFN-ϕ (current nomenclature as fish IFN-a/-b/-c/-d, etc.) genes in each species have been previously only found in teleosts [10,11,12]. As fish IFN-γ manifests orthologous phylogeny with the IFN-II identified in tetrapods, it is uncertain about IFN-ϕ genes, which were proposed to be ancestral molecules for either IFN-I or IFN-III, or both, found in tetrapods. Per gene structure, all fish IFN-ϕ genes contain about four introns, which resemble what have been observed in most IFN-III genes but not IFN-I genes in amniotes, where most IFN-I genes are intronless [10,11,12,46,47,48,49]. Examination of the proteins coded by fish IFN-ϕ genes indicated that they contain either two or four cysteines to form into intra-molecular disulfide bonds critical for IFN tertiary conformations and potential antiviral potency [10,11,12]. A recent study identified an ancestral IFN-III gene in searching transcriptome of catshark, a cartilaginous fish [12]. This resolves that the jawed vertebrate ancestor has orthologs of all three interferon types [12]. Therefore, the IFN-ϕ genes previously discovered in bony fish, including both two- and four-cysteine groups, are determined to be in the IFN-I lineage, even though they may not be orthologous to tetrapod intronless IFN-I genes [46,47,48,49].
Amphibians bridge vertebrate evolution from the water to terrestrial lives. From the perspective of IFN evolution, previous studies only found several intron-containing IFN genes belonging to IFN types, which thus assumed more ancestral conservation than rapid IFN evolution in amphibians [9,11]. In spite of this, extensive inspection of current Xenopus genome assemblies identified 6–9 intron-containing IFN genes in both IFN-I and IFN-III types, which indicated a moderate gene expansion to compare to that identified in most fish species (Figure 1 and Table 1) [9,12,13]. Most amphibian intron-containing IFN genes encode a protein with four cysteines, with some exceptions of two or three cysteines in both IFN-I and IFN-III types [9,12,13,16]. Phylogenically, Xenopus intron-containing IFN-I genes belong to two groups (Group 4s in Figure 1): One (consisting of only three IFN genes) falls sister to fish ancestral and tetrapod IFN-I genes, and another group sisters amphibian intronless IFN-I genes (Figure 1). This indicates that amphibian intron-containing IFN genes might experience less birth–death rates than their amniotic intronless counterparts to preserve potential orthologs that derived into IFN lineages discovered in tetrapod [12]. In addition, these primitive amphibian IFN genes also adapted to evolve into different subgroups of intronless IFN genes, which believably have more rapid birth–death dynamics to evolutionarily cope with pathogenic and physiological pressure during the amphibiotic stage of vertebrate evolution [11,12].
3. The Expansion of Amniote-Like Intronless IFN Genes in Amphibians
In addition to the conservation of intron-containing IFN genes, the co-existence of intronless IFN genes inscribes the amphibiotic role of amphibians in vertebrate evolution [12,13,16,50,51]. Prior to identification of intronless IFN genes in amphibians, a prevalent model for IFN evolution described that a retroposition process happened in reptiles and led to the origin and expansion of intronless IFN-I genes in amniotes [9,10]. The discovery of intronless IFN genes in amphibians revised this model and confirmed that the emergence of intronless IFN might happen in amphibians or even earlier. Indeed, the earliest existence of intronless IFN genes so far was only detected independently in different species of amphibians [12,13,16,50,51]. Genes without introns are a characteristic feature of prokaryotes; however, a small portion of intronless genes also exist (even emerge) in animal genomes, including all mammalian species. In eukaryotic cells, genes use introns to enhance transcription fidelity, i.e., intron–exon splicing inhibits topoisomerase-I cutting activity to reduce mutagenesis generated during gene transcription [52]. On the other hand, intronless genes express more rapidly upon stimuli, and duplicate or mutate more efficiently for new gene evolution [50,52]. In this line, intronless IFN gene evolution may represent a prokaryote-wise arm race adapted by animal immunity at a molecular level [50].
Amniotic IFN-I genes are almost all intronless, which have been classified into multiple subtypes, including IFN-α, IFN-β, IFN-δ, IFN-ε, IFN-κ, IFN-τ, IFN-ω, and IFN-ζ, commonly (IFN-α, IFN-β, IFN-ε, and IFN-κ) or species-specifically (IFN-δ, IFN-τ, IFN-ω, and IFN-ζ) expressed in mammalians [11,12,13,14,15,16]. Birds and reptiles also possess several to a dozen intronless IFN-I genes in each species, which have also been named such as IFN-α and IFN-β, however, are not necessarily orthologs (but paralogs) to their mammal counterparts [11,12]. Amphibians contain intronless IFN genes of both IFN-I and IFN-III types. This is unique among all studied vertebrate species because amniotic IFN-I genes are generally intronless, and intron-containing IFN genes are common for all fish IFN genes and amniotic IFN-III genes [11,12,13,14,15,16]. In the three studied amphibian species (i.e., Xenopus laevis, Xenopus tropicalis, and Nanorana parkeri), each species may harbor several to near 40 intronless IFN genes, and the majority of them belonging to IFN-I types [12,13,14,16,50,51]. Intronless Xenopus IFN-I genes were grouped into four groups based on their intermolecular sequence similarity, and further defined into three clades based on their phylogenic relationship to other tetrapod species [12,13]. For example, about 10 intronless Xenopus IFN-I genes were located within the mammal and reptile IFN-I clade, next to a clade containing only reptile sequences [12,13]. This suggests that amphibians contain orthologs of amniote intronless IFN-I genes, and thus provide the earliest intronless ancestors (identified so far) of tetrapod IFN-I genes [12,13]. The other intronless amphibian IFN-I genes were nested or fell sister to fish intron-containing IFN genes, indicating their close phylogeny to these primitive ancestors as well [12]. Intriguingly, several Xenopus IFN-I genes were found to contain a single short intron and were assumed to be intermediate molecules recoding the intron loss process [13]. One of these small-intron containing Xenopus IFN-I genes located sister to the group of cartilaginous fish IFN-I genes that were next to the intronless amniote IFN-Is in a recent phylogenic analysis [12]. Thus, the intronless amphibian IFN-I genes consist of a complex of tetrapod IFN intermediates between fish intron-containing ancestors and amniotic IFN-I genes; however, a one-to-one orthologous relationship is yet to be determined between amphibian and amniotic intronless IFN genes [12,13].
In addition, amphibians are among the few tetrapod species having intronless IFN-III genes [12,13,51]. Studies indicated that intronless IFN-III genes have evolved at least twice during amphibian evolution [12]. We also detected the co-existence of intronless IFN-III genes in many amniotic species, and hence, at least three times intron loss has happened throughout vertebrate evolution [14]. However, intronless IFN-III genes experienced much less expansion potency than intronless IFN-I genes in tetrapods. It is unclear what kind of molecular or phylogenic mechanisms underlie this difference between IFN-I and IFN-III genes (Table 1 and Figure 2) [11,12,13,14,15,16].
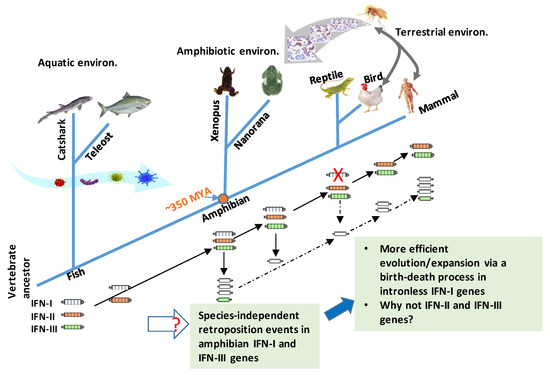
Figure 2.
An updated model for antiviral IFN (IFN-I and IFN-III) gene evolution in vertebrates. The ancestral genes of three type of IFNs (IFN-I, -II, and -III) have been identified in cartilaginous fish. All fish IFN genes are intron-containing ones generally with four introns (or five exons) [12]. In contrast, amniotic IFN-I genes are generally intronless. Co-existence of multiple intronless and intron-containing IFN genes were only detected independently in different species of amphibians [12,13,14,15,16,50,51]. The current evidences support that at least two retroposition events might have happened in amphibians, at about 180.70 MYA (million years ago) in Xenopus and 87.57 MYA in N. parkeri, for example [12,13,14,15,16,50,51]. These two independent retroposition events might have occurred much later than the divergence between amphibians and amniotes (i.e., ~350 MYA, orange dot). Another retroposition event leading to intronless IFN orthologs in amniotes could also happen in accompaniment to loss of intron-containing IFN genes in reptiles; however, it could be unlikely if an orthologous relationship exists between different intronless amphibian IFNs with amniotic ones [12]. The emergence and expansion of intronless amphibian IFNs likely reflect the increasing evolution pressure (especially the air-borne pathogens and physiological requirement for terrestrial adaptation, shown with broad curve arrows) during the transition period, when vertebrates migrated from aquatic to terrestrial environments. Retroposition defines a reverse-transcription process of cellular mRNA and reintegration into the genome to enhance gene copying and evolution in molecular evolution. Compared to intronless IFN-I genes, one paradox in IFN evolution is why IFN-II and –III genes conserve the ancestral gene structures, and do not show gene expansion even after few intronless IFN-III genes originated in amphibians and other amniotes. Solid or dashed black arrows: Indicating certain or uncertain orthologous relationships discovered, respectively.
4. Phylogenic View and Evolutionary Postulation about Amphibian IFN Evolution
In Figure 2, we summarize the current understanding of IFN evolution to highlight several key points and the newly discovered IFN complex in amphibians [12,13,14,15,16,50,51]. First, the ancestral molecules of three types of IFNs were identified in cartilaginous fish, indicating the earlier ramification of IFN primitive genes in fish [12]. This elegant discovery concludes a dispute about whether the fish IFN-ϕ genes are ancestral to tetrapod IFN-Is or IFN-IIIs to indicate that they all belong to IFN-Is, except a recently found SCCA-L from catshark, which is a fish IFN-III gene [12]. A question remains: Is that why no other IFN-III genes have been detected in bony fish so far? Extinct after the emergence or originated in a species- or taxon-independent manner? Second, intronless IFN genes in both IFN-I and IFN-III genes co-exist with intron-containing IFN genes, and this extensive co-existence has only been detected in different species of amphibians [12,13,16,50,51]. This indicates that the retroposition events that led to the emergence of intronless IFN genes happened several times in a species-independent way in amphibians [12,13,16,51]. Therefore, amphibians also comprise a unique animal taxon, containing both intronless and intron-containing IFN genes in both IFN-I and IFN-III types. Notably, intronless IFN-I genes but not IFN-III genes have experienced a rapid gene expansion, probably through a recently proposed birth–death process in Xenopus [12,13,16,51]. We propose that the emergence and expansion of intronless amphibian IFNs likely reflect the increase evolution pressure (especially the air-borne pathogens and physiological requirement for terrestrial adaptation) during the transition period when vertebrates migrated from aquatic to terrestrial environments. So why did intronless gene expansion happened in Xenopus but not obviously in Nanorana [13,16,51]? Maybe habitats close to human or domestic areas provides an epidemiological explanation [14,15]. Compared to intronless IFN-I genes, one paradox in IFN evolution is why IFN-II and –III genes conserve the ancestral gene structures, and do not show gene expansion even after few intronless IFN-III genes originated in amphibians and other ammonites. Recent studies discovered the epithelia-specified expression pattern and less-inflammatory activity of IFN-IIIs in mammalian studies [53,54,55,56,57]. It deserves investigation of whether this anatomic or physiological ramification limits the expansion potency of intronless IFN-III genes in tetrapods.
5. Amphibian IFN Complex: Expression and Function in Antiviral and Other Immune Responses
Described first for “interfering” with influenza virus in cultured chicken cells [58], IFNs are key antiviral cytokines and have multiple functions in immune regulation [4,5,6,7]. Nevertheless, most IFN functional studies have been conducted using mammal models, and amphibian IFNs remain largely uncharacterized. Previously, only several intron-containing IFN-I and IFN-III genes were identified in amphibians; and Xenopus IFN-III genes were ubiquitously detected in multiple tissues from healthy adult frogs [9,59,60]. Xenopus tissues or cells treated with poly (I:C) or infected with Ranavirus (FV3) robustly stimulated the expression of both types of IFN genes and their corresponding receptors. In Xenopus kidney cells and tadpole models, administration of recombinant IFN peptides of either Xenopus IFN1 or an IFNλ induced the expression of ISGs, including IFIT5, PKR, and MX1 [9,59,60]. The recombinant Xenopus IFN1 peptides protected the kidney-derived A6 cell line and tadpoles against FV3 infection, showing the decrease of the infectious viral burdens in both cases [59,60]. The study also observed that adult frogs were naturally more resistant to FV3 and were able to clear the infection within a few weeks, whereas tadpoles typically could not get over the viral infections. Correspondingly, adult X. laevis frogs demonstrated significantly higher FV3-elicited IFN gene expression than tadpoles, indicating that IFN response at least partly determined the antiviral consequence [59,60]. Even though IFN treatment markedly impaired viral replication and viral burdens in tadpoles, it only transiently increased tadpole survival time and did not change the final mortality rate of infected tadpoles due to considerable organ damage, regardless of IFN treatment in FV3-infected tadpoles [59,60]. Therefore, these studies revealed that amphibian intron-containing IFNs may consist of a basic immune barrier, even in healthy frogs; however, the undeveloped or irresponsive IFN status may contribute to the vulnerable consequence in tadpoles, which ascribe to a major portion of animal death in amphibian decline [59,60,61,62].
Genome-wide identification of intron-containing and intronless IFN genes in Xenopus and Nanorana species allows characterizing amphibian IFNs more extensively, in particular for the intronless subgroups [13,16]. The constitutive expression of multiple intronless IFN genes in X. laevis was demonstrated in most tested tissues, with a particular high expression in lung and heart [13]. The two intronless IFN-III genes were expressed in skin, stomach, and kidney. Similarly, the intron-containing IFN4 had the highest expression in kidney and heart. In contrast, IFN genes (such as the intronless IFNX3 and IFNLX group) that showed high inductive response (see context next) generally had a low expression level in normal tissues [13,50]. In addition to our gene-targeting expression analysis, most intronless IFN genes in X. tropicalis genome were also heavily mapped by RNA-Seq reads deposited at the public domains of NCBI, indicating functional expression of these less-studied amphibian IFNX genes (Table 2) [13].
Table 2.
Functional knowns and unknowns about amphibian IFN complex referring to well-studied amniotic IFNs.
Similarly, in the N. parkeri, an intron-containing IFN-I gene was highly expressed in kidney and intestine, as well as two intronless IFN-I genes in the lung, intestine, and skin, constitutively [16]. More than a 100-fold stimulation of the two intronless IFN genes was observed post-treatment of tissues (spleen, kidney, and liver) using poly(I:C) for 3–24 h [16]. More extensive inductive expression of amphibian IFN genes was compared in a kidney cell line from X. laevis (A6) after treatment of the cells with representative microbial mimics [13]. For example, intron-containing IFNs (represented by IFN4) were constitutively expressed at 5 h, but clearly showed inductive expression after cells were treated for 24 h. The viral dsRNA mimic, poly (I:C), most effectively stimulated several subgroups of IFNs, including both intron-containing and intronless IFNs in both IFN types at either 5 h or 24 h post-treatment. On the other hand, TLR2 and TLR4 ligands (Pam2/3CSK4/LTA and LPS, respectively) induced these same IFN expressions especially at 24 h post-treatment. Treatments with the ligands for TLR2/6 (Pam2CSK4) and TLR4 (LPS), but not ligands for TLR2/1 (Pam3CSK4) or TLR2 only (LTA), stimulated significant IFN expression. This indicates the differential regulation among amphibian IFN subgroups regarding defense responses against different pathogens [13].
The results from Xenopus tadpoles and adult frogs were more conclusive toward biological significance [61,62]. Using primary skin and kidney cells, Wendel et al. showed that most IFN genes were differentially expressed in the two cell types treated with various pathogenic mimics [62]. The findings include: (1) With few exceptions, most intron-containing IFN-I genes were more inductive in tadpole skin cells; (2) most intronless IFN-I and all IFN-III genes were more inductive in adult skin cells; and (3) the majority of IFN genes showed stronger induction in tadpole kidney cells; however, CpG DNA stimulated more robust IFN response in the adult kidney cells (Table 2). Post-FV3 infection, the expression of nine representative IFN genes (including six IFN-I and three IFN-III of both intron-containing and intronless groups) were increased 3–25-fold in the FV3-infected skin cells from both tadpoles and adult frogs; however, a robust stimulation (10–400-fold increase) of these IFN genes was detected in FV3-infected kidney cells from adult frogs, whereas negligible increases were detected in FV3-infected tadpole kidney cells [61,62]. Not only susceptible to amphibian FV3 infections, our data showed that frog kidney cells were also permissive to an influenza virus (H1N1 TX98) [13]. The flu virus infection induced a moderate increase of IFN4 and IFN-III gene expression at a low virus titer (multiplicity of infection or MOI at 1). A higher than 100-fold increase of these virus-responsive IFNs was detected in cells infected with the virus at a high MOI for 48 h. In addition, a strong expression of most intronless type I IFNs was shown in cells infected with the virus at 10 MOI, indicating a robust IFN reaction upon a massive viral infection [13]. In summary, the expression of these amphibian IFNs, particularly the intronless subgroup, was experimentally validated and shows differential expression in the ex vivo stimulated cells and animal tissues. Both the constitutive and inductive expressed patterns under various stimuli indicate the functional diversity of amphibian IFN complex as components of the innate immune system [13,16,59,60,61,62].
The antiviral activity of amphibian IFN peptides has been tested in both cells and animals [13,16,59,60,61,62]. Grayfer et al. produced a recombinant Xenopus IFN1 peptide using an insect cell-expression system [59,60]. The IFN1 peptide actively protected frog kidney cells against FV3 infection, and strongly induced MX1 (a robust antiviral ISG) expression in multiple tissues after administration into tadpoles for 24 h. In animal tests, the recombinant IFN1 peptide reduced viral burdens; however, did not improve the final mortality rate caused by FV3 infection in tadpoles [59,60]. Gan et al. tested the antiviral activity of N. parkeri intronless IFNi1 and IFNi2 by direct transfection of the IFN genes in a mammalian-expression vector into Xenopus kidney cells (A6 cells) [16]. They demonstrated that the transfection of IFNi1 and IFNi2 constructs induced the expression of serval typical ISGs and protected the cells against FV3 infection in the Xenopus kidney cells only if the IFNAR receptor genes of N. parkeri were co-transfected [16]. This indicates that there is a cross-species barrier of IFN antiviral activity due to species-specific IFN–IFNAR engagement in amphibians, further supporting the species-specific co-evolution of amphibian intronless IFN genes and corresponding IFN receptors [11,12,13,14,15,16]. We also used a mammalian-expression system to produce 15 Xenopus IFN peptides and determined their different antiviral activity against an influenza virus [13,14,15]. Antiviral assays by direct transfection of A6 cells using IFN-expressing constructs indicated that typical candidates of each amphibian IFN subgroup induced antiviral protection and reduced virus titers in A6 cells. In conclusion, the tested intron-containing IFN group (including IFN4, IFN6, and IFN7), which had higher activity against the viral cytopathic effect, are generally less active in suppression of viral replication. In contrast, the intronless IFNs demonstrated higher virus suppression, but were less effective to protect cell death from the viral infection [13,14]. Comparatively speaking, most Xenopus type III IFNs were less active in antiviral activity, except an IFNL4 that showed stronger virus-suppressive activity than most type I IFNs tested. Another significant IFN example was IFNX4, which demonstrated remarkable virus-suppressive and cell protective activity similar to most intron-containing IFNs tested. Clearly, all of these antiviral analyses validate that the Xenopus IFN complex comprises a profound antiviral diversity considering their antiviral activity against the influenza and FV3 infections [13,16,59,60,61,62].
The immune role of amphibian IFNs against other intracellular pathogens has also been recently tested [13]. Listeria monocytogenes represents a prevalent intracellular bacterial pathogen such as in food safety and public health. We detected that in the bacteria-infected A6 cells, most amphibian IFNs were highly stimulated when the cells were intracellularly infected with the bacteria. In all tests where cells were with different bacterial dosages, the intronless IFNX3 group was highly stimulated. This indicated that this intracellular IFN subtype (lack of signal peptides for secretion) might have evolved to regulate antibacterial activity inside cells. In cell cultures with a gel-overlay to restrict the bacterial intracellular growth, most IFN-expressing constructs showed antibacterial to some extent. However, most IFNs lost their antibacterial activity without a gel-overlay to limit the bacterial extracellular spreading. We also noted that IFNX2, the intracellular IFN isoform, developed significant antibacterial activity in both test conditions (with or without gel-overlay). Indeed, both intronless IFNX2 and IFNX3 are among the IFNX3 group, which showed the most antibacterial activity and were among the most inductive IFN genes in cells infected with L. monocytogenes, or treated with the ligands for either TLR2 (Pam2CSK4) or TLR4 (LPS) [13]. Considering that IFNX2 and IFNX3 are intracellular IFN peptides that lack a signal peptide for secretion, they potentially induce protective activity against intracellular bacteria via intracellular signaling pathways that are different from canonic IFN signaling through binding the membrane-bound IFNAR receptors [13,14,63]. Interferons and some ISGs (such as numerous IFN induced very large GTPase gene analogs) were found to be upregulated throughout Chytridiomycosis, a fungal skin infection that caused amphibian declines in several amphibian species [34,64]. Currently, there is lack of studies about the antifungal effects of amphibian IFNs. Several recent studies revealed a coordination between mouse type I IFNs and type III IFNs to regulate antifungal immune responses. Showing that during a fungal infection caused by Aspergillus fumigatus, type I IFNs produced by monocytes signaled the epithelial production of type III IFNs, which in turn played a critical role in regulating the antifungal neutrophil responses [65,66,67]. Given the diverse IFN-III genes evolved in amphibians, we expect to determine some amphibian IFN-IIIs that are important in antifungal immune response. In summary, recent studies conclude that the amphibian IFN family has a differential expression pattern and exerts a regulatory role in antiviral, antibacterial, and potential antifungal immune responses [13,16,59,60,61,62]. In addition, studies in amniotic IFNs also determined the multifunctional properties of antiviral IFNs in the regulation of immune cell development and metabolism, as well as animal pregnancy [68,69,70,71]. Whether amphibian IFN complex evolves to have similar functional diversification warrants further investigations in vertebrate physiology and development.
6. Conclusive Remarks: Amphibian IFN Complex—A Molecular Signature of Vertebrate Immune Evolution
Several recent studies have revealed a previously unknown complexity of the amphibian IFN system [11,12,13,14,15,16]. Being unique to animals from any other classes of vertebrates, amphibians not only conserve and multiply the fish-like intron-containing IFN genes, but also rapidly evolve amniote-like intronless IFN genes in three tested species [13,16,51]. Amphibians are thus critical intermediate species in IFN (and potentially other immune gene families too) evolution, in addition to their key position in studies of vertebrate development [24,35,40]. We postulate that the amphibian IFN system provides an irreplaceable model to study vertebrate immune evolution in molecular and functional diversity to cope with unprecedented immuno-physiological requirement during terrestrial adaption [11,12,13,14,15,16]. Studies so far have ascribed a defense role of these IFNs in immune regulation against intracellular pathogens particularly viruses and bacteria [13,16,59,60,61,62]; however, many knowledge gaps remain to be completed. Based on recent reports about IFN’s multifunctional properties in regulation of animal physiological and defense responses, we interpret that amphibian IFNs may evolve novel function pertinent to their superior molecular diversity [4,5,6,7,11,12,13,14,15,16]. Such new function revealed by the emerging studies of antifungal and developmental regulation of amphibian IFNs will certainly promote our understanding of immune evolution in vertebrates to address current pathogenic threats causing amphibian decline [25,26,27,28,29]. Xenopus is proposed as an alternative model for cancer studies due to its evolutionary tumor immunity [35]. Antitumor activity constitutes another key function of the antiviral IFNs [6]. We therefore expect that the amphibian IFN complex may have evolved IFN molecules bearing superior antitumor activity, and the Xenopus tumor immunity model can contribute to the development of IFN-based cancer immunotherapies [6,35].
Funding
This work was primarily supported by NSF-IOS-1831988, and in part by USDA NIFA 2018-67016-28313 and USDA NIFA Evans-Allen-1013186 to Y.S. through a resource sharing plan.
Acknowledgments
The authors thank Jacques Roberts and his laboratory at the University of Rochester Medical Center for collaborating in animal/material resources and idea discussion during the completion of relevant research projects that generated the results reviewed here.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ortutay, C.; Vihinen, M. Immunome: A reference set of genes and proteins for systems biology of the human immune system. Cell. Immunol. 2006, 244, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Biancotto, A.; McCoy, J.P. Studying the human immunome: The complexity of comprehensive leukocyte immunophenotyping. Curr. Top. Microbiol. Immunol. 2014, 377, 23–60. [Google Scholar] [PubMed]
- Savage, A.E.; Kiemnec-Tyburczy, K.M.; Ellison, A.R.; Fleischer, R.C.; Zamudio, K.R. Conservation and divergence in the frog immunome: Pyrosequencing and de novo assembly of immune tissue transcriptomes. Gene 2014, 542, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Snell, L.M.; McGaha, T.L.; Brooks, D.G. Type I interferon in chronic virus infection and cancer. Trends Immunol. 2017, 38, 542–557. [Google Scholar] [CrossRef]
- Levy, D.E.; Marié, I.J.; Durbin, J.E. Induction and function of type I and III interferon in response to viral infection. Curr. Opin. Virol. 2011, 1, 476–486. [Google Scholar] [CrossRef]
- Boehm, T.; Swann, J.B. Origin and evolution of adaptive immunity. Annu. Rev. Anim. Biosci. 2014, 2, 259–283. [Google Scholar] [CrossRef]
- Qi, Z.; Nie, P.; Secombes, C.J.; Zou, J. Intron-containing type I and type III IFN coexist in amphibians: Refuting the concept that a retroposition event gave rise to type I IFNs. J. Immunol. 2010, 184, 5038–5046. [Google Scholar] [CrossRef]
- Zou, J.; Tafalla, C.; Truckle, J.; Secombes, C.J. Identification of a second group of type I IFNs in fish sheds light on IFN evolution in vertebrates. J. Immunol. 2007, 179, 3859–3871. [Google Scholar] [CrossRef]
- Secombes, C.J.; Zou, J. Evolution of interferons and interferon receptors. Front. Immunol. 2017, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Redmond, A.K.; Zou, J.; Secombes, C.J.; Macqueen, D.J.; Dooley, H. Discovery of all three types in cartilaginous fishes enables phylogenetic resolution of the origins and evolution of interferons. Front. Immunol. 2019, 10, 1558. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Liu, Q.; Lee, J.; Ma, W.; McVey, D.S.; Blecha, F. Expansion of amphibian intronless interferons revises the paradigm for interferon evolution and functional diversity. Sci. Rep. 2016, 6, 29072. [Google Scholar] [CrossRef] [PubMed]
- Shields, L.E.; Jennings, J.; Liu, Q.; Lee, J.; Ma, W.; Blecha, F.; Miller, L.C.; Sang, Y. Cross-Species genome-wide analysis reveals molecular and functional diversity of the unconventional interferon-ω subtype. Front. Immunol. 2019, 10, 1431. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.; Sang, Y. Porcine Interferon Complex and Co-Evolution with Increasing Viral Pressure after Domestication. Viruses 2019, 11, 555. [Google Scholar] [CrossRef]
- Gan, Z.; Yang, Y.C.; Chen, S.N.; Hou, J.; Laghari, Z.A.; Huang, B.; Li, N.; Nie, P. Unique composition of intronless and intron-containing type I IFNs in the Tibetan frog Nanorana parkeri provides new evidence to support independent retroposition hypothesis for type I IFN genes in amphibians. J. Immunol. 2018, 201, 3329–3342. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hoffmann, H.H.; Schneider, W.M.; Rice, C.M. Interferons and viruses: An evolutionary arms race of molecular interactions. Trends Immunol. 2015, 36, 124–138. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Alves, J.M.; Carneiro, M.; Cheng, J.Y.; Lemos de Matos, A.; Rahman, M.M.; Loog, L.; Campos, P.F.; Wales, N.; Eriksson, A.; Manica, A.; et al. Parallel adaptation of rabbit populations to myxoma virus. Science 2019, 363, 1319–1326. [Google Scholar] [CrossRef]
- Burggren, W.W.; Warburton, S. Amphibians as animal models for laboratory research in physiology. ILAR J. 2007, 48, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Gurdon, J.B.; Hopwood, N. The introduction of Xenopus laevis into developmental biology: Of empire, pregnancy testing and ribosomal genes. Int. J. Dev. Biol. 2000, 44, 43–50. [Google Scholar] [PubMed]
- Callery, E.M. There’s more than one frog in the pond: A survey of the Amphibia and their contributions to developmental biology. Semin. Cell Dev. Biol. 2006, 17, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Horb, M.; Wlizla, M.; Abu-Daya, A.; McNamara, S.; Gajdasik, D.; Igawa, T.; Suzuki, A.; Ogino, H.; Noble, A.; Centre de Ressource Biologique Xenope team in France; et al. Xenopus Resources: Transgenic, Inbred and Mutant Animals, Training Opportunities, and Web-Based Support. Front. Physiol. 2019, 10, 387. [Google Scholar] [CrossRef]
- Pounds, J.A.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- Green, D.E.; Converse, K.A.; Schrader, A.K. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996–2001. Ann. N. Y. Acad. Sci. 2002, 969, 323–339. [Google Scholar] [CrossRef]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Organ. 2010, 92, 93–99. [Google Scholar] [CrossRef]
- Fites, J.S.; Ramsey, J.P.; Holden, W.M.; Collier, S.P.; Sutherland, D.M.; Reinert, L.K.; Gayek, A.S.; Dermody, T.S.; Aune, T.M.; Oswald-Richter, K.; et al. The invasive chytrid fungus of amphibians paralyzes lymphocyte responses. Science 2013, 342, 366–369. [Google Scholar] [CrossRef]
- Rosenblum, E.B.; James, T.Y.; Zamudio, K.R.; Poorten, T.J.; Ilut, D.; Rodriguez, D.; Eastman, J.M.; Richards-Hrdlicka, K.; Joneson, S.; Jenkinson, T.S.; et al. Complex history of the amphibian-killing chytrid fungus revealed with genome resequencing data. Proc. Natl. Acad. Sci. USA 2013, 110, 9385–9390. [Google Scholar] [CrossRef]
- Miller, D.; Gray, M.; Storfer, A. Ecopathology of ranaviruses infecting amphibians. Viruses 2011, 3, 2351–2373. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of amphibian communities due to an introduced Ranavirus. Curr. Biol. 2014, 24, 2586–2591. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.F.A.; Bui-Marinos, M.P.; Katzenback, B.A. Frog skin innate immune defences: Sensing and surviving pathogens. Front. Immunol. 2019, 9, 3128. [Google Scholar] [CrossRef] [PubMed]
- Hyoe, R.K.; Robert, J. A Xenopus tadpole alternative model to study innate-like T cell-mediated anti-mycobacterial immunity. Dev. Comp. Immunol. 2019, 92, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Grogan, L.F.; Robert, J.; Berger, L.; Skerratt, L.F.; Scheele, B.C.; Castley, J.G.; Newell, D.A.; McCallum, H.I. Review of the amphibian immune response to Chytridiomycosis, and future directions. Front. Immunol. 2018, 9, 2536. [Google Scholar] [CrossRef] [PubMed]
- Banach, M.; Robert, J. Tumor immunology viewed from alternative animal models-the Xenopus story. Curr. Pathobiol. Rep. 2017, 5, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Sater, A.K.; Moody, S.A. Using Xenopus to understand human disease and developmental disorders. Genesis 2017, 55, e22997. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A. Amphibian immunity-stress, disease, and climate change. Dev. Comp. Immunol. 2017, 66, 111–119. [Google Scholar] [CrossRef]
- Rebollar, E.A.; Antwis, R.E.; Becker, M.H.; Belden, L.K.; Bletz, M.C.; Brucker, R.M.; Harrison, X.A.; Hughey, M.C.; Kueneman, J.G.; Loudon, A.H.; et al. Using “Omics” and integrated multi-omics approaches to guide probiotic selection to mitigate chytridiomycosis and other emerging infectious diseases. Front. Microbiol. 2016, 7, 68. [Google Scholar] [CrossRef]
- Grayfer, L.; Robert, J. Amphibian macrophage development and antiviral defenses. Dev. Comp. Immunol. 2016, 58, 60–67. [Google Scholar] [CrossRef]
- Robert, J.; Jancovich, J.K. Recombinant Ranaviruses for studying evolution of host-pathogen interactions in ectothermic vertebrates. Viruses 2016, 8, 187. [Google Scholar] [CrossRef]
- Barra, D.; Simmaco, M. Amphibian skin: A promising resource for antimicrobial peptides. Trends Biotechnol. 1995, 13, 205–209. [Google Scholar] [CrossRef]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochim. Biophys. Acta 2009, 1788, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Helbing, C.C.; Hammond, S.A.; Jackman, S.H.; Houston, S.; Warren, R.L.; Cameron, C.E.; Birol, I. Antimicrobial peptides from Rana [Lithobates] catesbeiana: Gene structure and bioinformatic identification of novel forms from tadpoles. Sci. Rep. 2019, 9, 1529. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef]
- Langevin, C.; Aleksejeva, E.; Passoni, G.; Palha, N.; Levraud, J.P.; Boudinot, P. The antiviral innate immune response in fish: Evolution and conservation of the IFN system. J. Mol. Biol. 2013, 425, 4904–4920. [Google Scholar] [CrossRef]
- Boudinot, P.; Langevin, C.; Secombes, C.J.; Levraud, J.P. The peculiar characteristics of fish type I interferons. Viruses 2016, 8, 298. [Google Scholar] [CrossRef]
- Robertsen, B. The role of type I interferons in innate and adaptive immunity against viruses in Atlantic salmon. Dev. Comp. Immunol. 2018, 80, 41–52. [Google Scholar] [CrossRef]
- Pereiro, P.; Figueras, A.; Novoa, B. Insights into teleost interferon-gamma biology: An update. Fish Shellfish Immunol. 2019, 90, 150–164. [Google Scholar] [CrossRef]
- Krause, C.D. Intron loss in interferon genes follows a distinct set of stages, and may confer an evolutionary advantage. Cytokine 2016, 83, 193–205. [Google Scholar] [CrossRef]
- Gan, Z.; Chen, S.N.; Huang, B.; Hou, J.; Nie, P. Intronless and intron-containing type I IFN genes coexist in amphibian Xenopus tropicalis: Insights into the origin and evolution of type I IFNs in vertebrates. Dev. Comp. Immunol. 2017, 67, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Yang, Y. Why eukaryotic cells use introns to enhance gene expression: Splicing reduces transcription associated mutagenesis by inhibiting topoisomerase I cutting activity. Biol. Direct 2011, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Rivera, A.; Parker, D.; Durbin, J.E. Type III IFNs: Beyond antiviral protection. Semin. Immunol. 2019, 43, 101303. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Odendall, C. Editorial: Interferon-λs: New regulators of inflammatory processes. Front. Immunol. 2019, 10, 2117. [Google Scholar] [CrossRef]
- Ye, L.; Schnepf, D.; Staeheli, P. Interferon-λ orchestrates innate and adaptive mucosal immune responses. Nat. Rev. Immunol. 2019, 19, 614–625. [Google Scholar] [CrossRef]
- Rivera, A. Interferon lambda’s new role as regulator of neutrophil function. J. Interferon Cytokine Res. 2019, 39, 609–617. [Google Scholar] [CrossRef]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and distinct functions of type I and type III interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- Isaacs, A.; Lindenmann, J. Virus interference. I. The interferon. Proc. R. Soc. Lond. Ser. Biol. Sci. 1957, 147, 258–267. [Google Scholar]
- Grayfer, L.; De Jesús Andino, F.; Robert, J. The amphibian (Xenopus laevis) type I interferon response to frog virus 3: New insight into ranavirus pathogenicity. J. Virol. 2014, 88, 5766–5777. [Google Scholar] [CrossRef]
- Grayfer, L.; De Jesús Andino, F.; Robert, J. Prominent amphibian (Xenopus laevis) tadpole type III interferon response to the frog virus 3 ranavirus. J. Virol. 2015, 89, 5072–5082. [Google Scholar] [CrossRef]
- Wendel, E.S.; Yaparla, A.; Koubourli, D.V.; Grayfer, L. Amphibian (Xenopus laevis) tadpoles and adult frogs mount distinct interferon responses to the Frog Virus 3 ranavirus. Virology 2017, 503, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wendel, E.S.; Yaparla, A.; Melnyk, M.L.S.; Koubourli, D.V.; Grayfer, L. Amphibian (Xenopus laevis) tadpoles and adult frogs differ in their use of expanded repertoires of type I and type III interferon cytokines. Viruses 2018, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.X.; Zou, J.; Nie, P.; Huang, B.; Yu, Z.; Collet, B.; Secombes, C.J. Intracellular interferons in fish: A unique means to combat viral infection. PLoS Pathog. 2013, 9, e1003736. [Google Scholar] [CrossRef] [PubMed]
- Grogan, L.F.; Cashins, S.D.; Skerratt, L.F.; Berger, L.; McFadden, M.S.; Harlow, P.; Hunter, D.A.; Scheele, B.C.; Mulvenna, J. Evolution of resistance to chytridiomycosis is associated with a robust early immune response. Mol. Ecol. 2018, 27, 919–934. [Google Scholar] [CrossRef]
- Espinosa, V.; Dutta, O.; McElrath, C.; Du, P.; Chang, Y.J.; Cicciarelli, B.; Pitler, A.; Whitehead, I.; Obar, J.J.; Durbin, J.E.; et al. Type III interferon is a critical regulator of innate antifungal immunity. Sci. Immunol. 2017, 2, eaan5357. [Google Scholar] [CrossRef]
- Maldonado, S.; Fitzgerald-Bocarsly, P. Antifungal activity of plasmacytoid dendritic cells and the impact of chronic HIV infection. Front. Immunol. 2017, 8, 1705. [Google Scholar] [CrossRef]
- Hassanzadeh-Kiabi, N.; Yáñez, A.; Dang, I.; Martins, G.A.; Underhill, D.M.; Goodridge, H.S. Autocrine type I IFN signaling in dendritic cells stimulated with fungal β-glucans or lipopolysaccharide promotes CD8 T cell activation. J. Immunol. 2017, 198, 375–382. [Google Scholar] [CrossRef]
- Wu, D.; Sanin, D.E.; Everts, B.; Chen, Q.; Qiu, J.; Buck, M.D.; Patterson, A.; Smith, A.M.; Chang, C.H.; Liu, Z.; et al. Type1 interferons induce changes in core metabolism that are critical for immune function. Immunity 2016, 44, 1325–1336. [Google Scholar] [CrossRef]
- Parlakgul, G.; Hotamisligil, G.S. Type I interferons interfere with liver glucose metabolism. Cell Metab. 2017, 25, 1211–1212. [Google Scholar] [CrossRef]
- Tian, Y.; Jennings, J.; Gong, Y.; Sang, Y. Viral infections and interferons in the development of obesity. Biomolecules 2019, 9, 726. [Google Scholar] [CrossRef]
- Forde, N.; Lonergan, P. Interferon-tau and fertility in ruminants. Reproduction 2017, 154, F33–F43. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).