
Transcriptome Changes in Glioma Cells upon Infection with the Oncolytic Virus VV-GMCSF-Lact
 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Immortalized Cell Lines
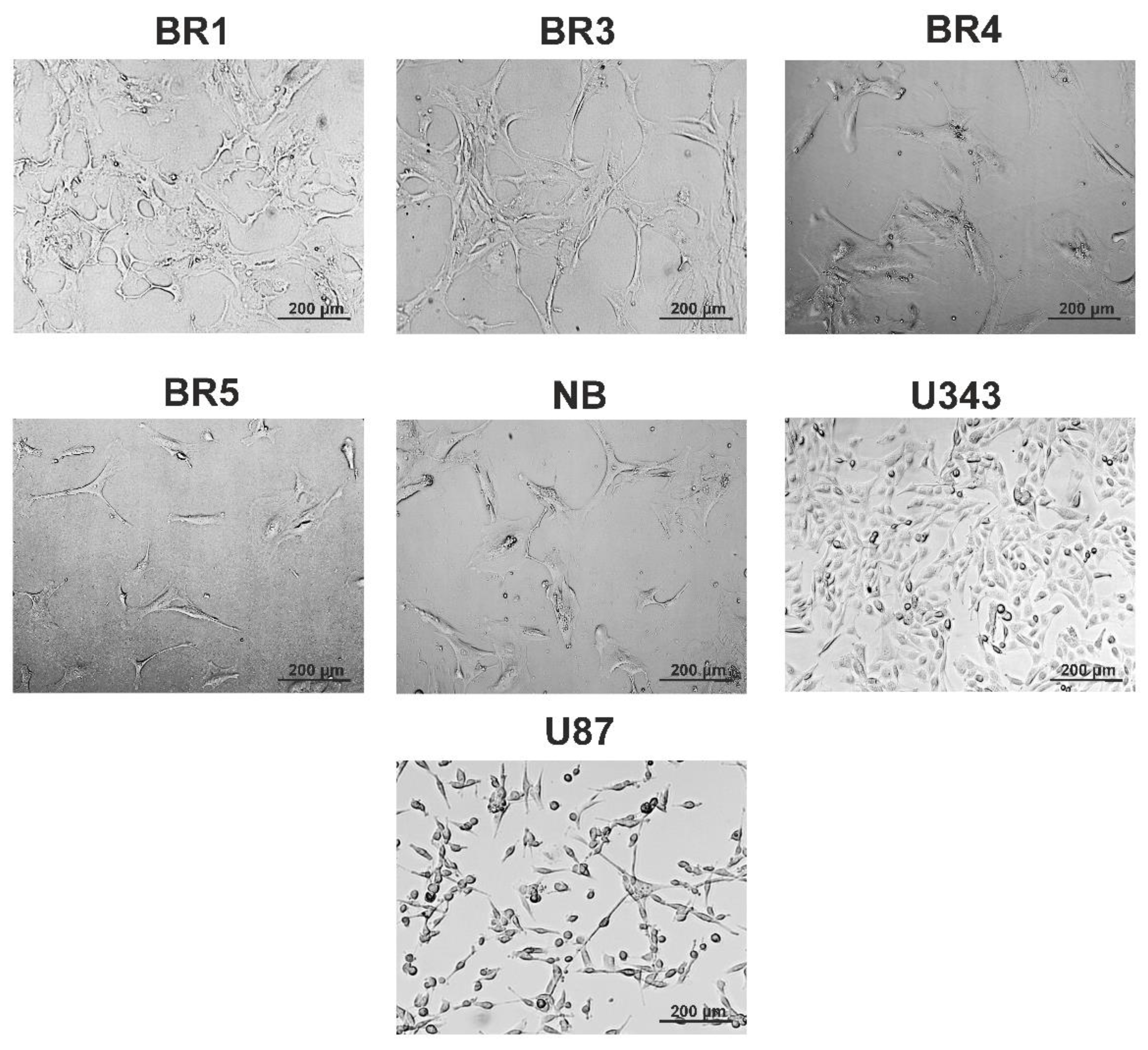
2.2. Patient-Derived Cell Cultures
2.3. Oncolytic Virus VV-GMCSF-Lact
2.4. Flow Cytometry
2.5. XCelligence Assay
2.6. Apoptosis Detection
2.7. Determination of the Cytotoxic Dose (CD50) of VV-GMCSF-Lact for Glioma and Normal Brain Cell Cultures
2.8. Treatment with VV-GMCSF-Lact for Next-Generation RNA Sequencing
2.9. RNA Isolation
2.10. RNA Sequencing
2.11. Transcriptome Analysis
3. Results
3.1. Cell Cultures and General NGS RNA-Seq Data
3.1.1. Cell Cultures
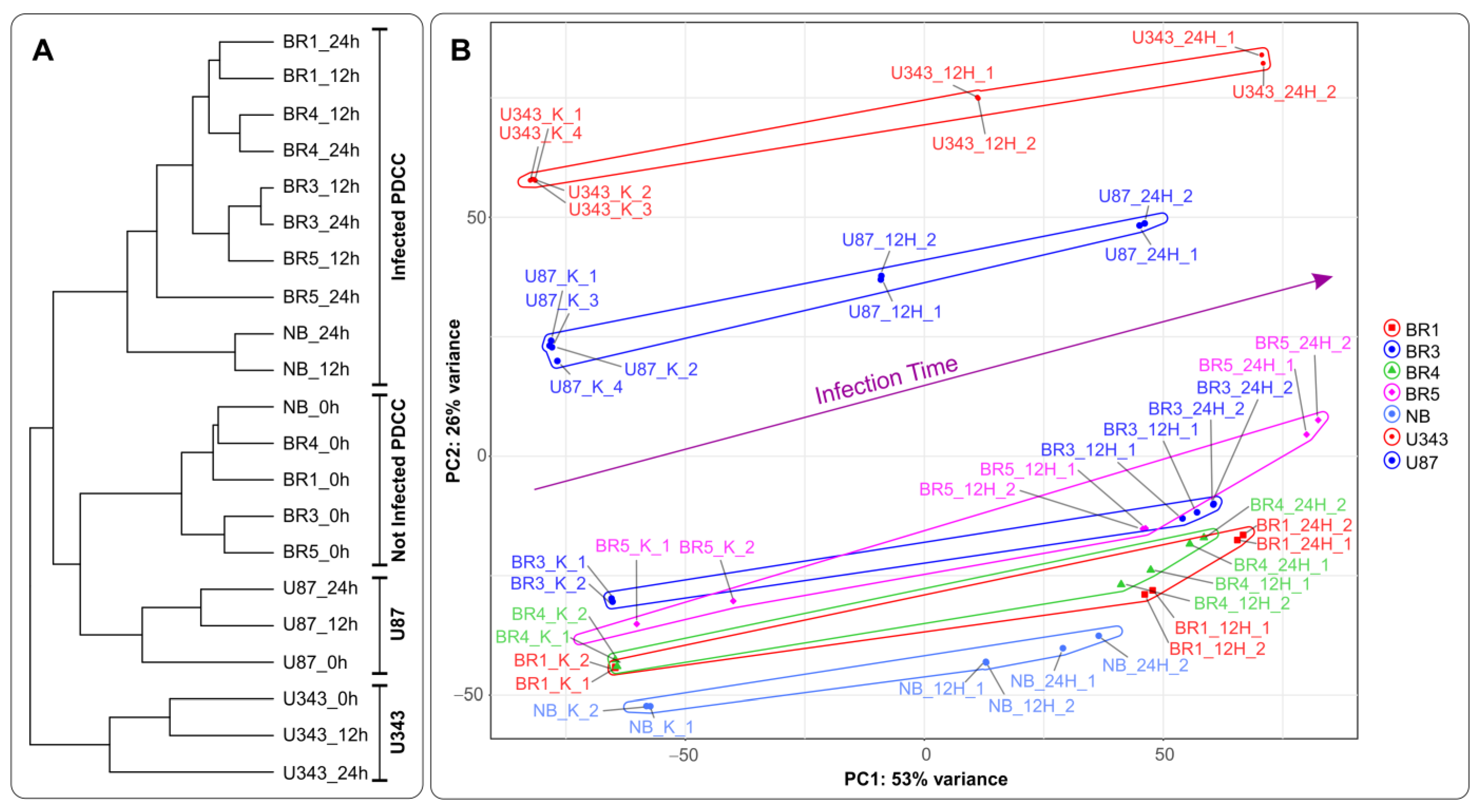
3.1.2. RNA-Seq Data
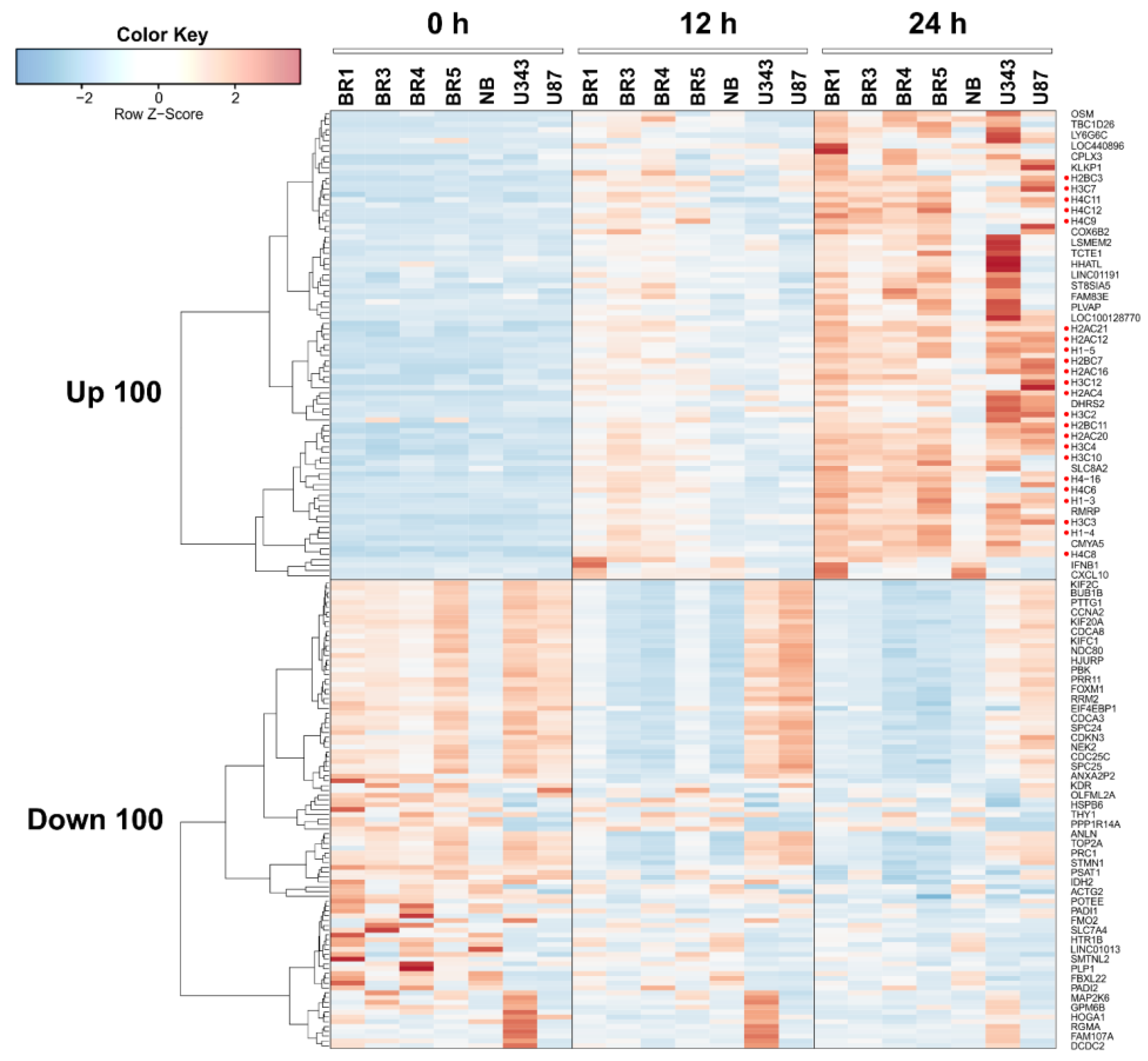
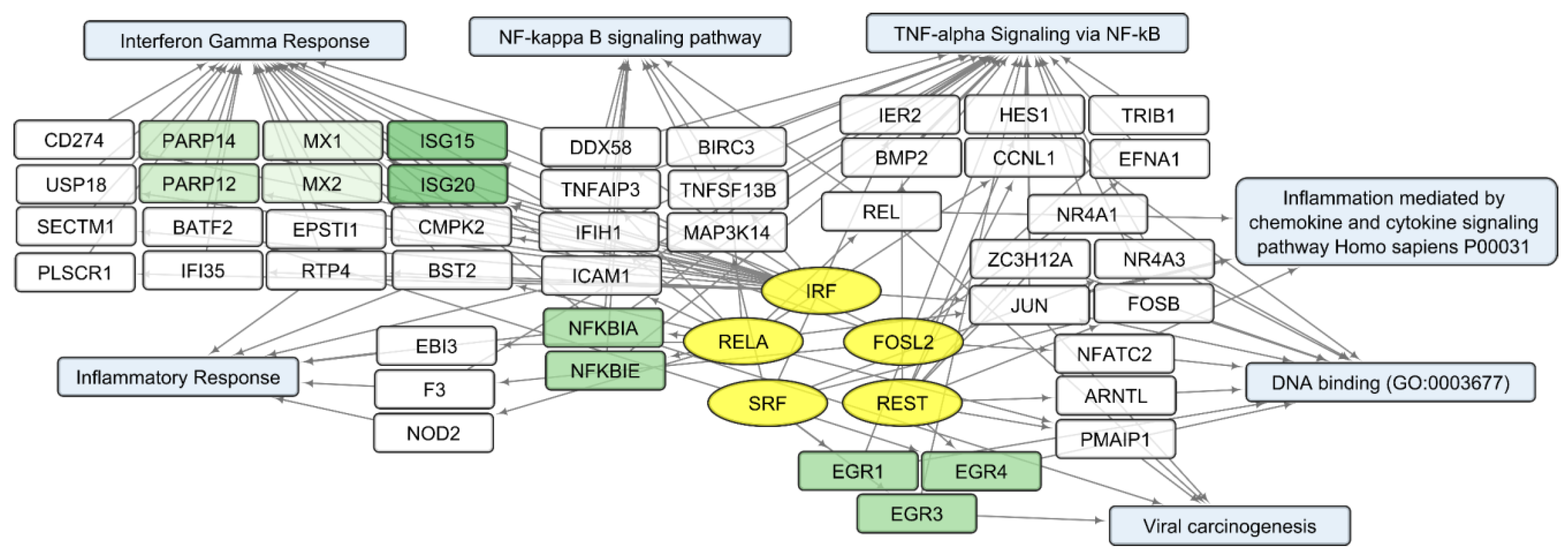
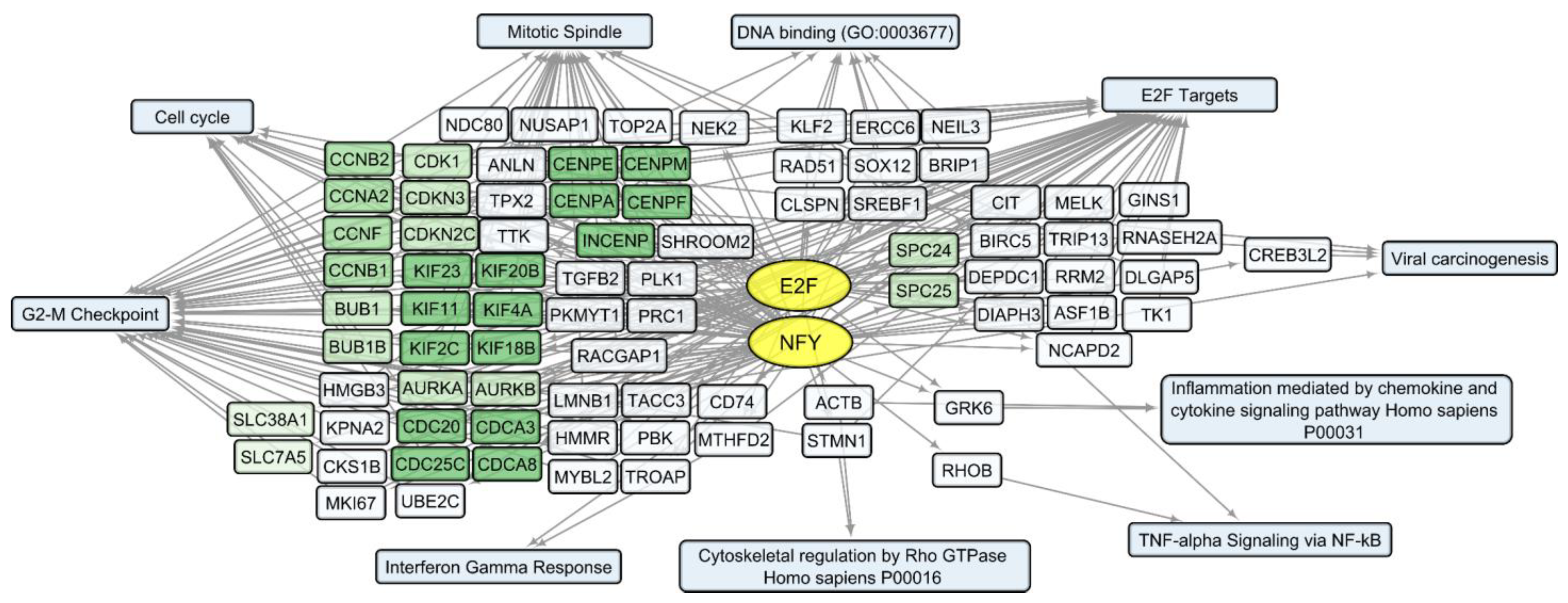
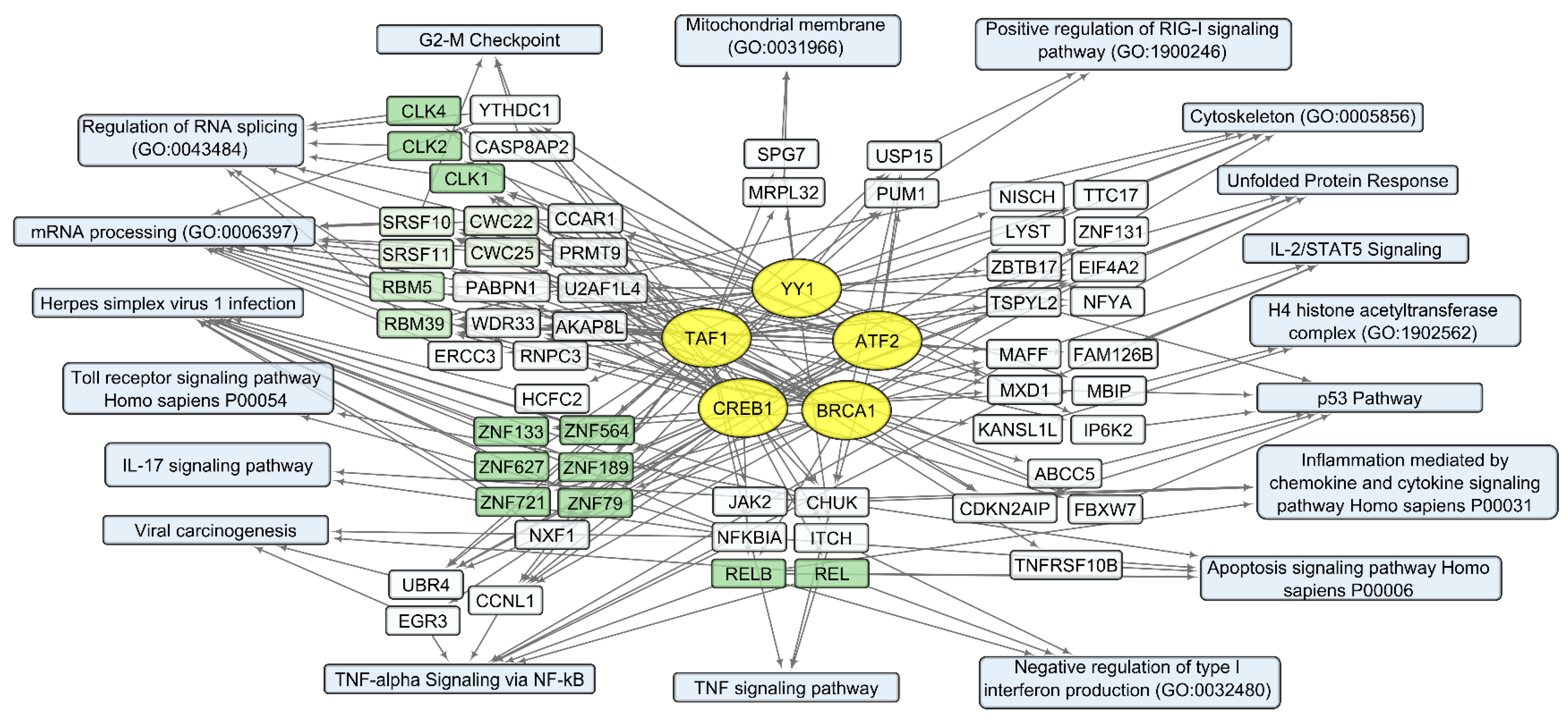
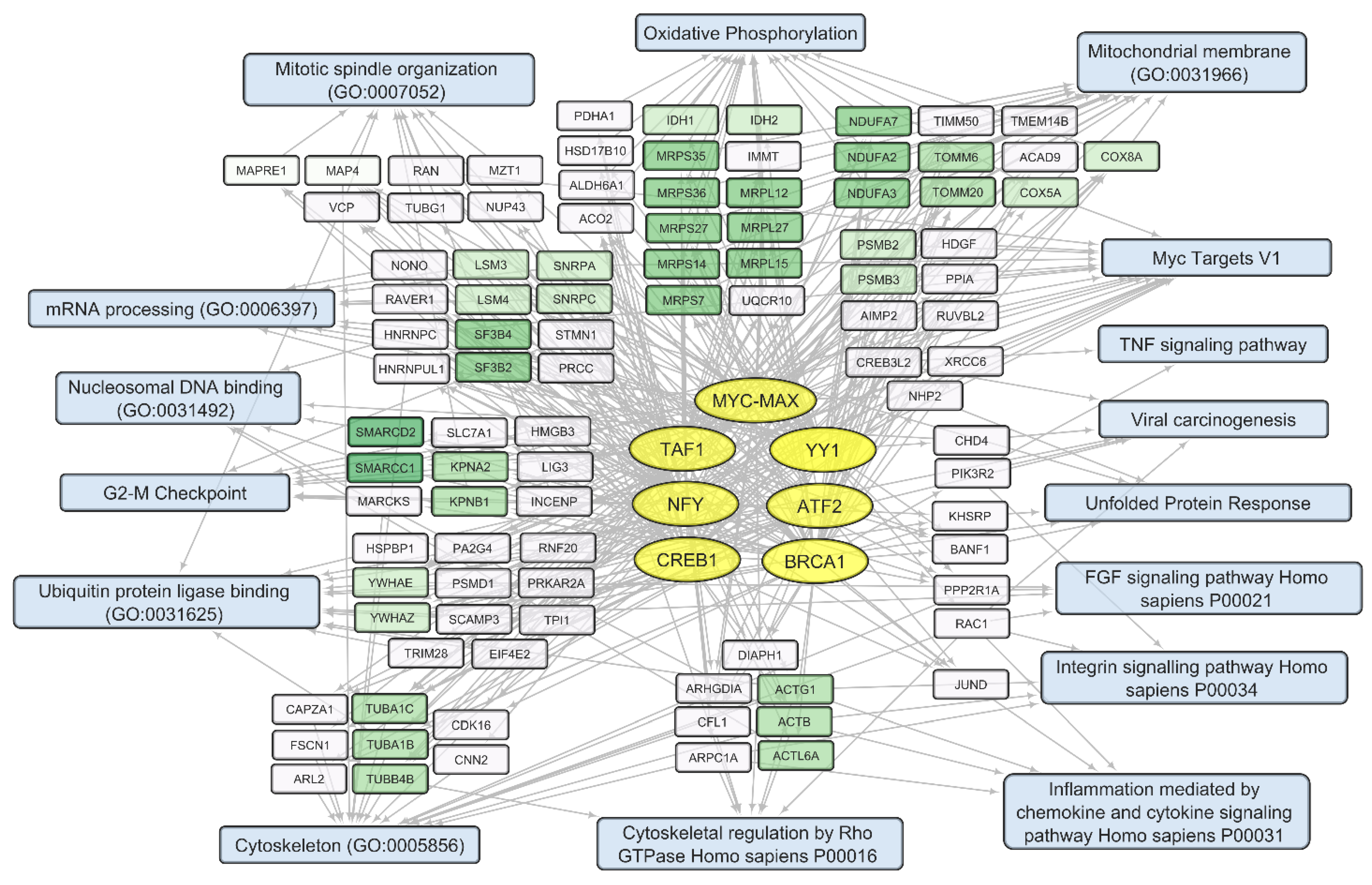
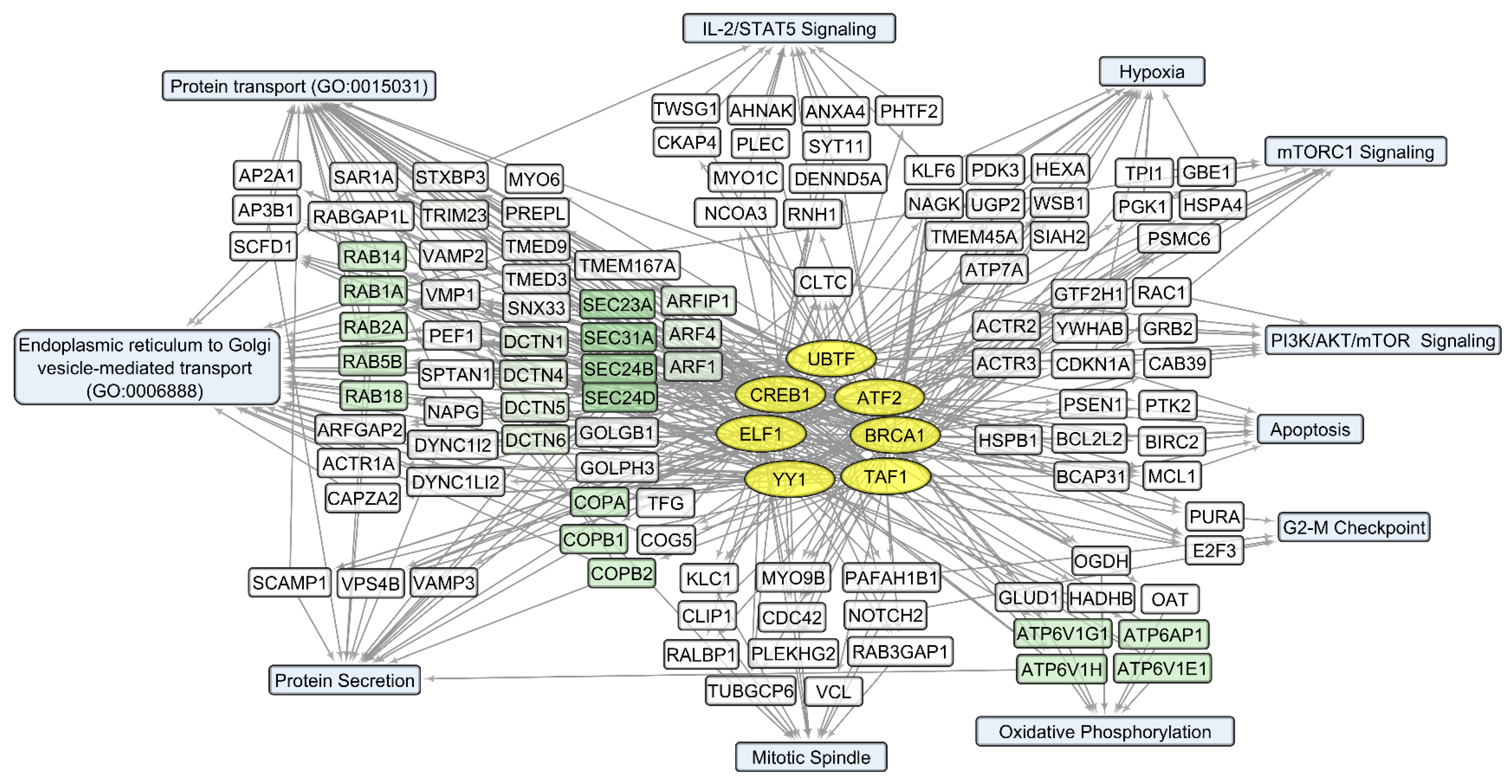
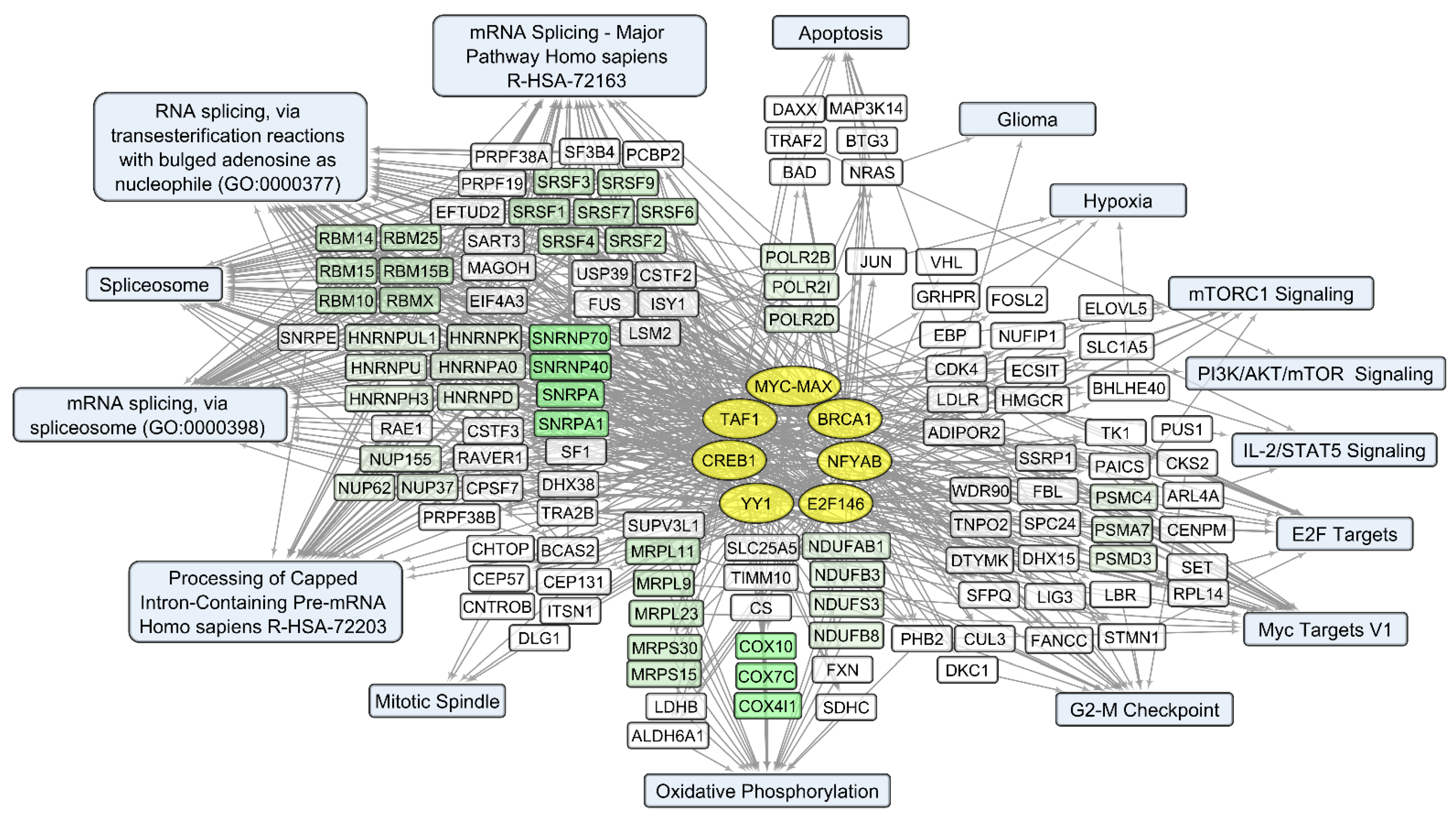
3.2. General Changes in Individual Transcript Levels, Transcription Factor Activity, Biological Processes, and Pathways Affected by VV-GMCSF-Lact Infection
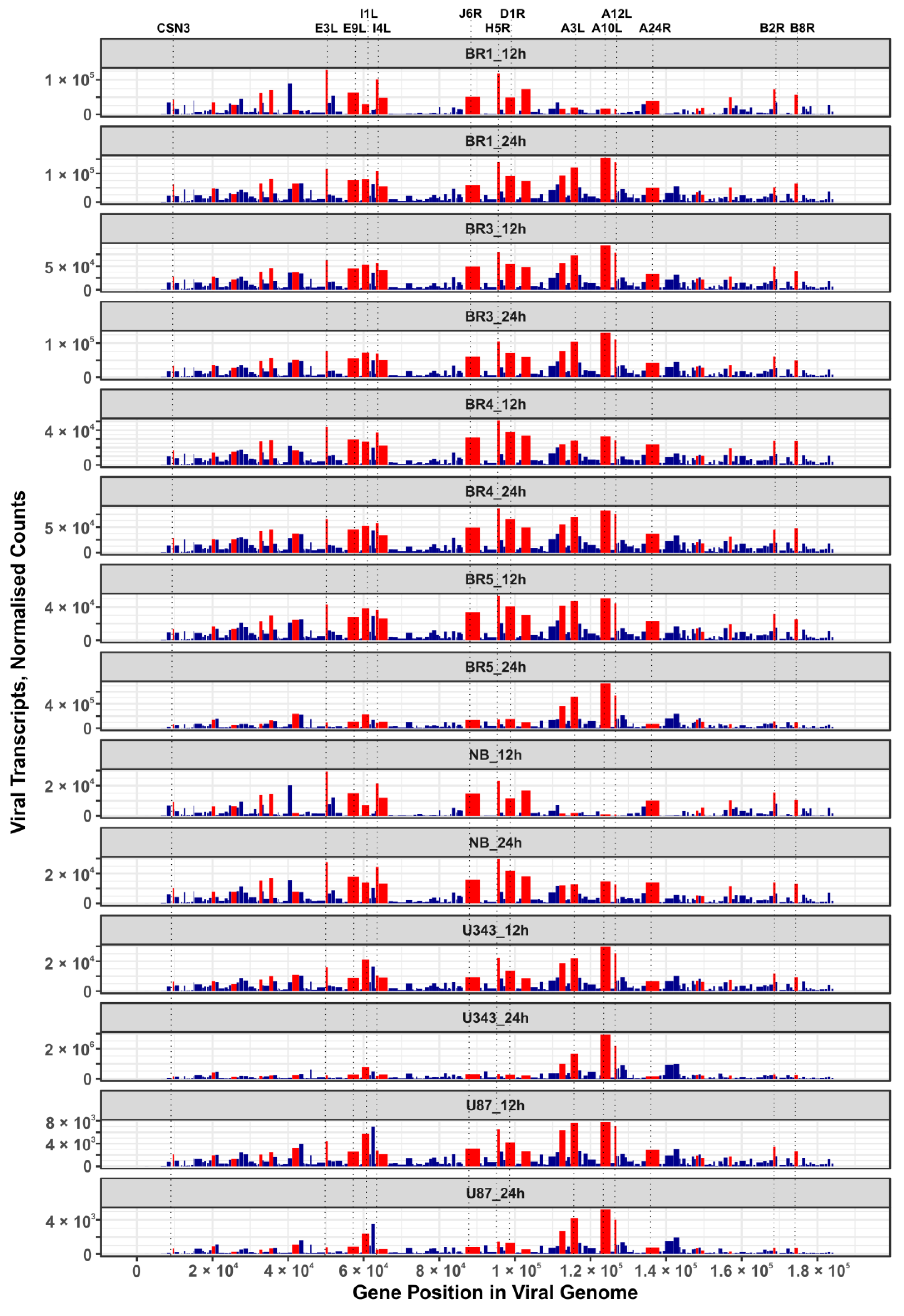
3.3. VV-GMCSF-Lact Transcripts in Infected Glioma and NB Cell Cultures
3.4. Human Transcripts That Correlate with the VV-GMCSF-Lact RNA in Glioma and NB Cells
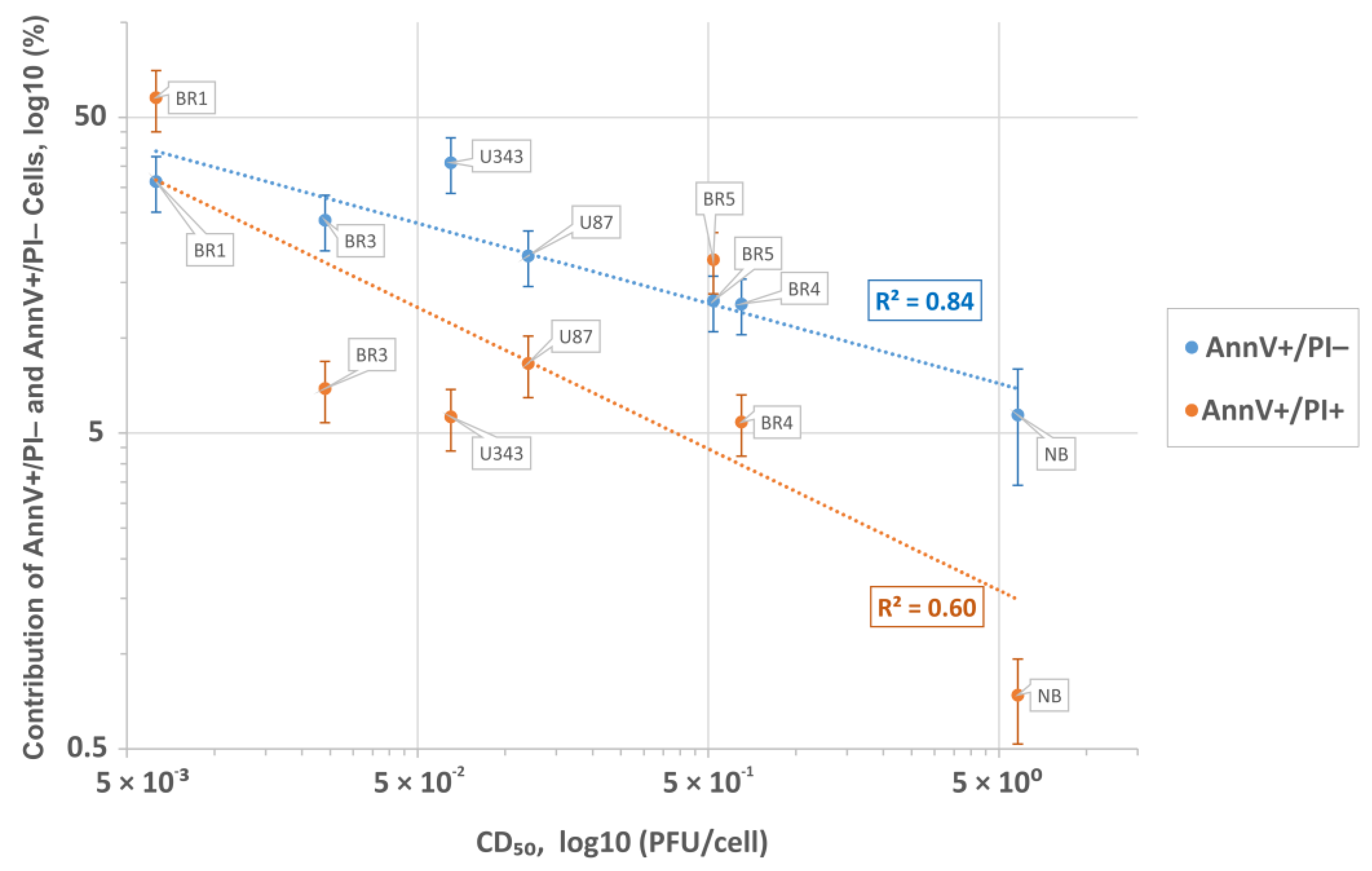
3.5. Human Transcripts That Correlate with the VV-GMCSF-Lact Cytotoxic Dose CD50 in Glioma and NB Cells
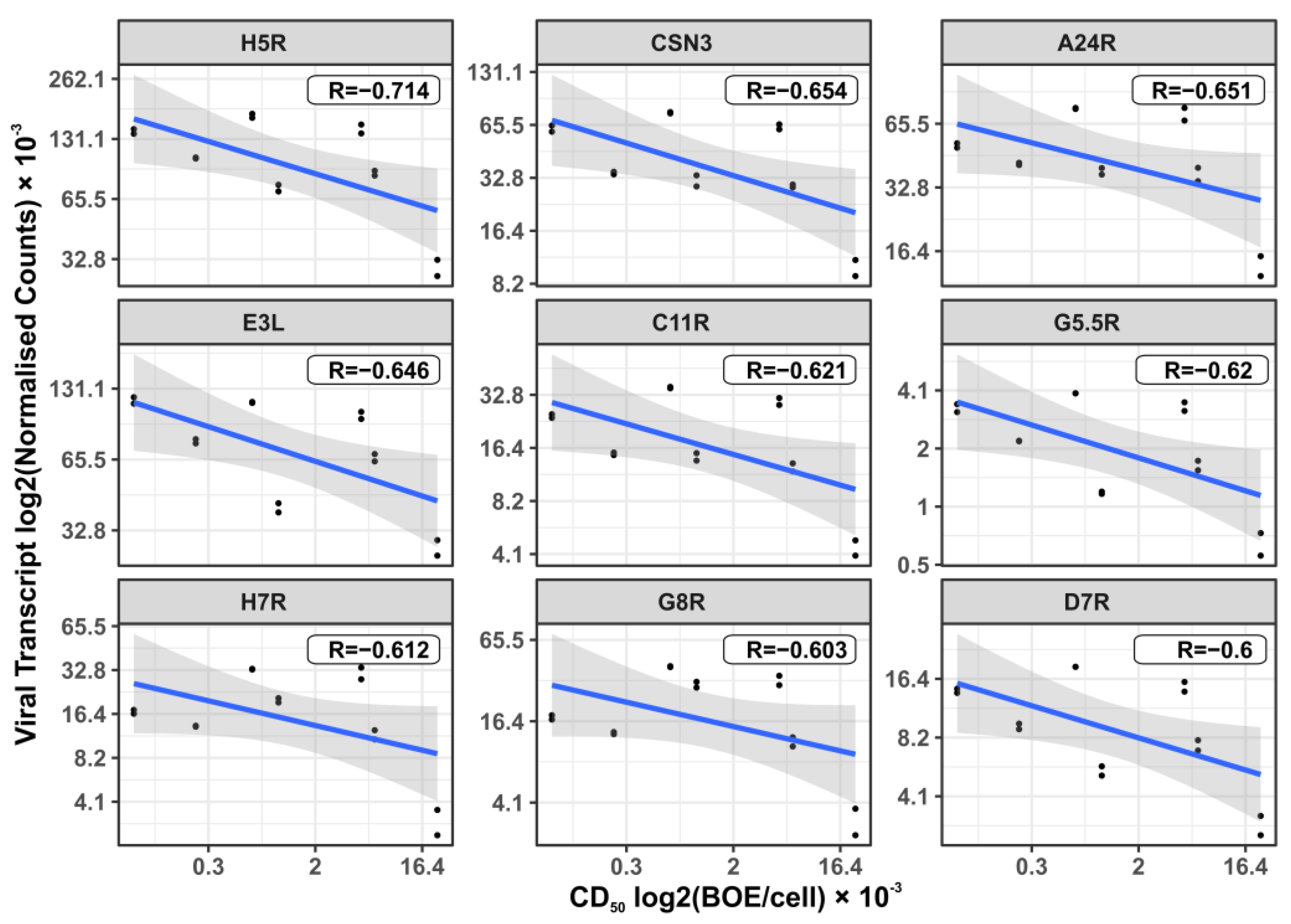
3.6. Viral Transcripts That Correlate with the Cytotoxic Dose of the Virus in Glioma and NB Cells Infected with VV-GMCSF-Lact
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CD50 | cytotoxic dose |
| CI | cell index |
| CSN2 | kappa-casein gene |
| DEG | differentially expressed gene |
| GBM | glioblastoma, previously referred to as “glioblastoma multiforme” |
| GM-CSF | granulocyte-macrophage colony-stimulating factor (product of CSF2 gene) |
| HC | hierarchical clustering |
| MOI | the multiplicity of infection |
| NB | normal brain tissue cell culture |
| PCA | principal component analysis |
| PFU | plaque-forming unit |
| tk | thymidine kinase |
| U343 | U343 MG cell culture |
| U87 | U87 MG cell culture |
| VACV | vaccinia virus |
| vgf | virus growth factor |
| VST | variance stabilizing transformation |
References
- Ostrom, Q.T.; Patil, N.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013–2017. Neuro. Oncol. 2020, 22, iv1–iv96. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, X.; Sander, M.; Zhang, H.; Yan, G.; Lin, Y. Oncolytic Viro-Immunotherapy: An Emerging Option in the Treatment of Gliomas. Front. Immunol. 2021, 12, 721830. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Núñez, F.J.; Haase, S.; McClellan, B.L.; Faisal, S.M.; Carney, S.V.; Yu, J.; Alghamri, M.S.; Asad, A.S.; Candia, A.J.N.; et al. Current Approaches for Glioma Gene Therapy and Virotherapy. Front. Mol. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Todo, T.; Ito, H.; Ino, Y.; Ohtsu, H.; Ota, Y.; Shibahara, J.; Tanaka, M. Intratumoral Oncolytic Herpes Virus G47∆ for Residual or Recurrent Glioblastoma: A Phase 2 Trial. Nat. Med. 2022, 28, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Thorne, S.; Bartlett, D.; Kirn, D. The Use of Oncolytic Vaccinia Viruses in the Treatment of Cancer: A New Role for an Old Ally? Curr. Gene Ther. 2005, 5, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Beerli, C.; Yakimovich, A.; Kilcher, S.; Reynoso, G.V.; Fläschner, G.; Müller, D.J.; Hickman, H.D.; Mercer, J. Vaccinia Virus Hijacks EGFR Signalling to Enhance Virus Spread through Rapid and Directed Infected Cell Motility. Nat. Microbiol. 2019, 4, 216–225. [Google Scholar] [CrossRef]
- Parato, K.A.; Breitbach, C.J.; Le Boeuf, F.; Wang, J.; Storbeck, C.; Ilkow, C.; Diallo, J.S.; Falls, T.; Burns, J.; Garcia, V.; et al. The Oncolytic Poxvirus JX-594 Selectively Replicates in and Destroys Cancer Cells Driven by Genetic Pathways Commonly Activated in Cancers. Mol. Ther. 2012, 20, 749–758. [Google Scholar] [CrossRef]
- Hengstschläger, M.; Knöfler, M.; Müllner, E.W.; Ogris, E.; Wintersberger, E.; Wawra, E. Different Regulation of Thymidine Kinase during the Cell Cycle of Normal versus DNA Tumor Virus-Transformed Cells. J. Biol. Chem. 1994, 269, 13836–13842. [Google Scholar] [CrossRef]
- Thorne, S.H.; Hwang, T.-H.H.; O’Gorman, W.E.; Bartlett, D.L.; Sei, S.; Kanji, F.; Brown, C.; Werier, J.; Cho, J.-H.; Lee, D.-E.; et al. Rational Strain Selection and Engineering Creates a Broad-Spectrum, Systemically Effective Oncolytic Poxvirus, JX-963. J. Clin. Investig. 2007, 117, 3350–3358. [Google Scholar] [CrossRef]
- Kochneva, G.; Sivolobova, G.; Tkacheva, A.; Grazhdantseva, A.; Troitskaya, O.; Nushtaeva, A.; Tkachenko, A.; Kuligina, E.; Richter, V.; Koval, O. Engineering of Double Recombinant Vaccinia Virus with Enhanced Oncolytic Potential for Solid Tumor Virotherapy. Oncotarget 2016, 7, 74171–74188. [Google Scholar] [CrossRef]
- Koval, O.A.; Tkachenko, A.V.; Fomin, A.S.; Semenov, D.V.; Nushtaeva, A.A.; Kuligina, E.V.; Zavjalov, E.L.; Richter, V.A. Lactaptin Induces P53-Independent Cell Death Associated with Features of Apoptosis and Autophagy and Delays Growth of Breast Cancer Cells in Mouse Xenografts. PLoS ONE 2014, 9, e93921. [Google Scholar] [CrossRef]
- Vasileva, N.; Ageenko, A.; Dmitrieva, M.; Nushtaeva, A.; Mishinov, S.; Kochneva, G.; Richter, V.; Kuligina, E. Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma. Life 2021, 11, 1084. [Google Scholar] [CrossRef]
- Hughes, L.; Wilkins, K.; Goldsmith, C.S.; Smith, S.; Hudson, P.; Patel, N.; Karem, K.; Damon, I.; Li, Y.; Olson, V.A.; et al. A Rapid Orthopoxvirus Purification Protocol Suitable for High-Containment Laboratories. J. Virol. Methods 2017, 243, 68–73. [Google Scholar] [CrossRef]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase Update, a Database of Eukaryotic Repetitive Elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Hartley, S.W.; Mullikin, J.C. QoRTs: A Comprehensive Toolset for Quality Control and Data Processing of RNA-Seq Experiments. BMC Bioinform. 2015, 16. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Caragher, S.; Chalmers, A.J.; Gomez-Roman, N. Glioblastoma’s Next Top Model: Novel Culture Systems for Brain Cancer Radiotherapy Research. Cancers 2019, 11, 44. [Google Scholar] [CrossRef]
- Da Hora, C.C.; Schweiger, M.W.; Wurdinger, T.; Tannous, B.A. Patient-Derived Glioma Models: From Patients to Dish to Animals. Cells 2019, 8. [Google Scholar] [CrossRef]
- Mooney, K.L.; Choy, W.; Sidhu, S.; Pelargos, P.; Bui, T.T.; Voth, B.; Barnette, N.; Yang, I. The Role of CD44 in Glioblastoma Multiforme. J. Clin. Neurosci. 2016, 34, 1–5. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, K.; Wang, Z.; Zhao, M.; Deng, Y.; Ji, W.; Zou, Y.; Qian, C.; Liu, Y.; Xiao, H.; et al. CD44-Mediated Poor Prognosis in Glioma Is Associated With M2-Polarization of Tumor-Associated Macrophages and Immunosuppression. Front. Surg. 2021, 8, 775194. [Google Scholar] [CrossRef]
- Zeng, J.; Xi, S.-Y.; Wang, F.; Liao, H.-D.; Yang, Y.-Z.; Hu, W.-M. L1CAM High Expression Associates with Poor Prognosis in Glioma but Does Not Correlate with C11orf95-RELA Fusion. Biomed Res. Int. 2020, 2020, 1353284. [Google Scholar] [CrossRef]
- Wachowiak, R.; Krause, M.; Mayer, S.; Peukert, N.; Suttkus, A.; Müller, W.C.; Lacher, M.; Meixensberger, J.; Nestler, U. Increased L1CAM (CD171) Levels Are Associated with Glioblastoma and Metastatic Brain Tumors. Medicine 2018, 97, e12396. [Google Scholar] [CrossRef]
- Maness, P.F.; Schachner, M. Neural Recognition Molecules of the Immunoglobulin Superfamily: Signaling Transducers of Axon Guidance and Neuronal Migration. Nat. Neurosci. 2007, 10, 19–26. [Google Scholar] [CrossRef]
- Dzwonek, J.; Wilczynski, G.M. CD44: Molecular Interactions, Signaling and Functions in the Nervous System. Front. Cell. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Pathogenesis of Viral Infections and Diseases. In Fenner’s Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 47–78. ISBN 9780123751584. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Lehman, C.W.; Smith, A.; Kelly, J.; Jacobs, J.L.; Dinman, J.D.; Kehn-Hall, K. EGR1 Upregulation during Encephalitic Viral Infections Contributes to Inflammation and Cell Death. Viruses 2022, 14, 1210. [Google Scholar] [CrossRef]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Greseth, M.D.; Traktman, P. The Life Cycle of the Vaccinia Virus Genome. Annu. Rev. Virol. 2022, 9, 239–259. [Google Scholar] [CrossRef]
- Sodeik, B.; Doms, R.W.; Ericsson, M.; Hiller, G.; Machamer, C.E.; Van’t Hof, W.; Van Meer, G.; Moss, B.; Griffiths, G. Assembly of Vaccinia Virus: Role of the Intermediate Compartment between the Endoplasmic Reticulum and the Golgi Stacks. J. Cell Biol. 1993, 121, 521–542. [Google Scholar] [CrossRef] [PubMed]
- Dueck, K.J.; Hu, Y.; Chen, P.; Deschambault, Y.; Lee, J.; Varga, J.; Cao, J. Mutational Analysis of Vaccinia Virus E3 Protein: The Biological Functions Do Not Correlate with Its Biochemical Capacity to Bind Double-Stranded RNA. J. Virol. 2015, 89, 5382–5394. [Google Scholar] [CrossRef] [PubMed]
- Bersch, B.; Tarbouriech, N.; Burmeister, W.P.; Iseni, F. Solution Structure of the C-Terminal Domain of A20, the Missing Brick for the Characterization of the Interface between Vaccinia Virus DNA Polymerase and Its Processivity Factor. J. Mol. Biol. 2021, 433, 167009. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.L.; Desaulniers, M.A.; Noyce, R.S.; Evans, D.H. The Acidic C-Terminus of Vaccinia Virus I3 Single-Strand Binding Protein Promotes Proper Assembly of DNA-Protein Complexes. Virology 2016, 489, 212–222. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gammon, D.B.; Gowrishankar, B.; Duraffour, S.; Andrei, G.; Upton, C.; Evans, D.H. Vaccinia Virus-Encoded Ribonucleotide Reductase Subunits Are Differentially Required for Replication and Pathogenesis. PLoS Pathog. 2010, 6, e1000984. [Google Scholar] [CrossRef]
- Mossman, K.; Upton, C.; Buller, R.M.; McFadden, G. Species Specificity of Ectromelia Virus and Vaccinia Virus Interferon-Gamma Binding Proteins. Virology 1995, 208, 762–769. [Google Scholar] [CrossRef]
- D’Costa, S.M.; Bainbridge, T.W.; Kato, S.E.; Prins, C.; Kelley, K.; Condit, R.C. Vaccinia H5 Is a Multifunctional Protein Involved in Viral DNA Replication, Postreplicative Gene Transcription, and Virion Morphogenesis. Virology 2010, 401, 49–60. [Google Scholar] [CrossRef]
- Schweneker, M.; Lukassen, S.; Späth, M.; Wolferstätter, M.; Babel, E.; Brinkmann, K.; Wielert, U.; Chaplin, P.; Suter, M.; Hausmann, J. The Vaccinia Virus O1 Protein Is Required for Sustained Activation of Extracellular Signal-Regulated Kinase 1/2 and Promotes Viral Virulence. J. Virol. 2012, 86, 2323–2336. [Google Scholar] [CrossRef]
- Tate, J.; Gollnick, P. The Role of Vaccinia Termination Factor and Cis-Acting Elements in Vaccinia Virus Early Gene Transcription Termination. Virology 2015, 485, 179–188. [Google Scholar] [CrossRef]
- Condit, R.C.; Moussatche, N.; Traktman, P. In a Nutshell: Structure and Assembly of the Vaccinia Virion. Adv. Virus Res. 2006, 66, 31–124. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Hruby, D.E. Vaccinia Virus A12L Protein and Its AG/A Proteolysis Play an Important Role in Viral Morphogenic Transition. Virol. J. 2007, 4, 73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ember, S.W.J.; Ren, H.; Ferguson, B.J.; Smith, G.L. Vaccinia Virus Protein C4 Inhibits NF-ΚB Activation and Promotes Virus Virulence. J. Gen. Virol. 2012, 93, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- Engelstad, M.; Smith, G.L. The Vaccinia Virus 42-KDa Envelope Protein Is Required for the Envelopment and Egress of Extracellular Virus and for Virus Virulence. Virology 1993, 194, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Gerlic, M.; Faustin, B.; Postigo, A.; Yu, E.C.-W.; Proell, M.; Gombosuren, N.; Krajewska, M.; Flynn, R.; Croft, M.; Way, M.; et al. Vaccinia Virus F1L Protein Promotes Virulence by Inhibiting Inflammasome Activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7808–7813. [Google Scholar] [CrossRef]
- Morikawa, S.; Sakiyama, T.; Hasegawa, H.; Saijo, M.; Maeda, A.; Kurane, I.; Maeno, G.; Kimura, J.; Hirama, C.; Yoshida, T.; et al. An Attenuated LC16m8 Smallpox Vaccine: Analysis of Full-Genome Sequence and Induction of Immune Protection. J. Virol. 2005, 79, 11873–11891. [Google Scholar] [CrossRef]
- Jesus, D.M.; Moussatche, N.; McFadden, B.B.D.; Nielsen, C.P.; D’Costa, S.M.; Condit, R.C. Vaccinia Virus Protein A3 Is Required for the Production of Normal Immature Virions and for the Encapsidation of the Nucleocapsid Protein L4. Virology 2015, 481, 1–12. [Google Scholar] [CrossRef]
- De Silva, F.S.; Paran, N.; Moss, B. Products and Substrate/Template Usage of Vaccinia Virus DNA Primase. Virology 2009, 383, 136–141. [Google Scholar] [CrossRef]
- Van Meir, E.; Wittek, R. Fine Structure of the Vaccinia Virus Gene Encoding the Precursor of the Major Core Protein 4 A. Arch. Virol. 1988, 102, 19–27. [Google Scholar] [CrossRef]
- Blasco, R.; Sisler, J.R.; Moss, B. Dissociation of Progeny Vaccinia Virus from the Cell Membrane Is Regulated by a Viral Envelope Glycoprotein: Effect of a Point Mutation in the Lectin Homology Domain of the A34R Gene. J. Virol. 1993, 67, 3319–3325. [Google Scholar] [CrossRef]
- Brennan, G.; Kitzman, J.O.; Shendure, J.; Geballe, A.P. Experimental Evolution Identifies Vaccinia Virus Mutations in A24R and A35R That Antagonize the Protein Kinase R Pathway and Accompany Collapse of an Extragenic Gene Amplification. J. Virol. 2015, 89, 9986–9997. [Google Scholar] [CrossRef]
- Lynn, H.; Howell, L.M.; Diefenbach, R.J.; Newsome, T.P. Phototracking Vaccinia Virus Transport Reveals Dynamics of Cytoplasmic Dispersal and a Requirement for A36R and F12L for Exit from the Site of Wrapping. Viruses 2018, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Fedosyuk, S.; Grishkovskaya, I.; de Almeida Ribeiro, E.; Skern, T. Characterization and Structure of the Vaccinia Virus NF-ΚB Antagonist A46. J. Biol. Chem. 2014, 289, 3749–3762. [Google Scholar] [CrossRef]
- Rehm, K.E.; Connor, R.F.; Jones, G.J.B.; Yimbu, K.; Roper, R.L. Vaccinia Virus A35R Inhibits MHC Class II Antigen Presentation. Virology 2010, 397, 176–186. [Google Scholar] [CrossRef]
- Ryerson, M.R.; Richards, M.M.; Kvansakul, M.; Hawkins, C.J.; Shisler, J.L. Vaccinia Virus Encodes a Novel Inhibitor of Apoptosis That Associates with the Apoptosome. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.; Kumar, N.D.; Reggiori, F.; Khan, G. How Viruses Hijack and Modify the Secretory Transport Pathway. Cells 2021, 10, 2535. [Google Scholar] [CrossRef]
- Lin, D.; Shen, Y.; Liang, T. Oncolytic Virotherapy: Basic Principles, Recent Advances and Future Directions. Signal Transduct. Target. Ther. 2023, 8, 156. [Google Scholar] [CrossRef]
- Townsley, A.C.; Weisberg, A.S.; Wagenaar, T.R.; Moss, B. Vaccinia Virus Entry into Cells via a Low-PH-Dependent Endosomal Pathway. J. Virol. 2006, 80, 8899–8908. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bruno, D.P.; Martens, C.A.; Porcella, S.F.; Moss, B. Simultaneous High-Resolution Analysis of Vaccinia Virus and Host Cell Transcriptomes by Deep RNA Sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 11513–11518. [Google Scholar] [CrossRef] [PubMed]
- Alkhalil, A.; Hammamieh, R.; Hardick, J.; Ichou, M.A.; Jett, M.; Ibrahim, S. Gene Expression Profiling of Monkeypox Virus-Infected Cells Reveals Novel Interfaces for Host-Virus Interactions. Virol. J. 2010, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Xuan, D.T.M.; Yeh, I.-J.; Wu, C.-C.; Su, C.-Y.; Liu, H.-L.; Chiao, C.-C.; Ku, S.-C.; Jiang, J.-Z.; Sun, Z.; Ta, H.D.K.; et al. Comparison of Transcriptomic Signatures between Monkeypox-Infected Monkey and Human Cell Lines. J. Immunol. Res. 2022, 2022, 3883822. [Google Scholar] [CrossRef]
- Tolonen, N.; Doglio, L.; Schleich, S.; Krijnse Locker, J. Vaccinia Virus DNA Replication Occurs in Endoplasmic Reticulum-Enclosed Cytoplasmic Mini-Nuclei. Mol. Biol. Cell 2001, 12, 2031–2046. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.J.; Benfield, C.T.O.; Ren, H.; Lee, V.H.; Frazer, G.L.; Strnadova, P.; Sumner, R.P.; Smith, G.L. Vaccinia Virus Protein N2 Is a Nuclear IRF3 Inhibitor That Promotes Virulence. J. Gen. Virol. 2013, 94, 2070–2081. [Google Scholar] [CrossRef]
- Teferi, W.M.; Desaulniers, M.A.; Noyce, R.S.; Shenouda, M.; Umer, B.; Evans, D.H. The Vaccinia Virus K7 Protein Promotes Histone Methylation Associated with Heterochromatin Formation. PLoS ONE 2017, 12, e0173056. [Google Scholar] [CrossRef] [PubMed]
- Soday, L.; Lu, Y.; Albarnaz, J.D.; Davies, C.T.R.; Antrobus, R.; Smith, G.L.; Weekes, M.P. Quantitative Temporal Proteomic Analysis of Vaccinia Virus Infection Reveals Regulation of Histone Deacetylases by an Interferon Antagonist. Cell Rep. 2019, 27, 1920–1933.e7. [Google Scholar] [CrossRef] [PubMed]
- Novoa, R.R.; Calderita, G.; Arranz, R.; Fontana, J.; Granzow, H.; Risco, C. Virus Factories: Associations of Cell Organelles for Viral Replication and Morphogenesis. Biol. Cell 2005, 97, 147–172. [Google Scholar] [CrossRef]
- Grimm, C.; Hillen, H.S.; Bedenk, K.; Bartuli, J.; Neyer, S.; Zhang, Q.; Hüttenhofer, A.; Erlacher, M.; Dienemann, C.; Schlosser, A.; et al. Structural Basis of Poxvirus Transcription: Vaccinia RNA Polymerase Complexes. Cell 2019, 179, 1537–1550.e19. [Google Scholar] [CrossRef]
- Ly, M.; Burgess, H.M.; Shah, S.B.; Mohr, I.; Glaunsinger, B.A. Vaccinia Virus D10 Has Broad Decapping Activity That Is Regulated by MRNA Splicing. PLoS Pathog. 2022, 18, e1010099. [Google Scholar] [CrossRef]
- Huang, T.; Nilsson, C.E.; Punga, T.; Akusjarvi, G. Functional Inactivation of the SR Family of Splicing Factors during a Vaccinia Virus Infection. EMBO Rep. 2002, 3, 1088–1093. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Culture | Diagnosis (1) | Tumor Grade (1) | Cell Markers (%) | Cell Index (2) | CD50 (3) (PFU/Cell) | Apoptotic Markers (4) (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CD133+ | CD44+ | CD133+/CD44+ | CD171+ | CD15+ | AnnV+/PI- | AnnV+/PI+ | |||||
| BR1 | GBM | IV | 15.1 | 67.7 | 14.2 | 24.03 | 0 | 11.12 | 6.3 × 10−3 | 31.3 | 57.7 |
| BR3 | GBM | IV | 3.83 | 90.9 | 3.69 | 0 | 0 | 3.43 | 2.4 × 10−2 | 23.6 | 6.92 |
| BR4 | GBM | IV | 9.92 | 78.9 | 9.56 | 20.4 | 1.95 | 0.94 | 6.5 × 10−1 | 12.8 | 5.42 |
| BR5 | Anaplastic astrocytoma | III | 0.57 | 34.7 | 0.37 | 0 | 0 | 3.45 | 5.2 × 10−1 | 13.1 | 17.7 |
| NB | Normal brain | -- | 0 | 99.7 | 0 | 44.1 | 0 | 1.07 | 5.8 × 100 | 5.7 | 0.74 |
| U87 | GBM | IV | 0 | 7.47 | 0 | 0 | 50.1 | 1.39 | 1.2 × 10−1 | 18.2 | 8.31 |
| U343 | GBM | IV | 2.48 | 15.9 | 0.59 | 0 | 6.04 | 2.59 | 6.5 × 10−2 | 35.9 | 5.63 |
| Infection Time, Regulation | Number of DEGs | DEGs * | |
|---|---|---|---|
| 12 h | Up | 5 | H2BC5; H4C5; H2BC8; H4C8; H4C2 |
| Down | 1 | MARCKS | |
| 24 h | Up | 85 | AHSA2P; AOC2; ARID4B; AVIL; AXIN2; B3GNT2; BAZ2A; BRD1; CCDC17; CCNL1; CDKN2AIP; CLCN6; CLDN15; CLK4; CMYA5; CWC22; CWC25; DNAJA1; DNHD1; FGFR1OP2; FLCN; FNBP4; H1-2; H1-3; H1-4; H2AC17; H2AC4; H2AC6; H2AC8; H2BC11; H2BC15; H2BC18; H2BC21; H2BC4; H2BC5; H2BC8; H2BC9; H3C3; H3C4; H4C2; H4C3; H4C5; H4C8; HSPA1B; HSPA6; JMJD1C; KDM6A; KHDC4; LIME1; LOC102724814; LOC284454; LOC729603; LSMEM1; LTB4R; MAPK8IP3; MBIP; MORC3; NFKBIZ; NXF1; PAXBP1; PPTC7; PTGS2; PUM2; QRICH2; RBBP6; RBM33; RBM5; RNU4-2; RPPH1; RSRC2; SCARNA2; SERTAD1; SLC25A25; SLC8A2; SMPD4BP; SYNGAP1; TENT4B; TMEM259; UBE2B; WAC; YOD1; ZFYVE27; ZNF160; ZNF211; ZNF451 |
| Down | 54 | ACTB; ACTR1A; AP2B1; ARF3; ARPC4; ATP5MC2; C11orf68; CALD1; CAPNS1; CAPRIN1; CAPZB; CAVIN1; CDC42; CDK4; CFL2; CLTA; CNN2; COPZ1; CSRP1; DAZAP2; EHD2; EI24; EPN1; FAM98A; FSCN1; IMMT; LASP1; LOXL1; LRRC59; MAP1A; MAP4; MARCKS; NONO; PABPC4; PARVA; PLIN3; RHOA; RNF20; RSU1; RTL8C; S100A11; SH3BGRL3; SKI; SMARCC1; SNX12; STMP1; TLN1; TRAPPC1; TXN2; UQCR10; UROS; USP22; VIM; WFS1 | |
| Viral Gene | Average Rank (1) | Temporal Expression (2) | Preferred Name (2) Annotation | Ref |
|---|---|---|---|---|
| E3L | 30.1 | Early | Double-strand RNA-binding protein. Host range function and inhibition of innate immune responses. | [36] |
| E9L | 33.3 | Early | DNA polymerase. The catalytic subunit, a family B DNA polymerase. | [37] |
| I3L | 34.1 | Early Intermediate Late | Single-stranded DNA-binding protein. | [38] |
| I4L | 34.8 | Early | Ribonucleotide reductase large subunit. | [39] |
| B8R | 35.1 | Early | Soluble interferon-gamma receptor-like protein. Host defense modulator. | [40] |
| H5R | 35.9 | Early Late | Late transcription factor VLTF-4. DNA synthesis, postreplicative gene transcription, and virion morphogenesis. | [41] |
| O1L | 37.5 | -- | Hypothetical protein. Positive regulator of the ERK1/2 pathway downstream of the EGFR. | [42] |
| D1R | 38.6 | Early | Large subunit of mRNA capping enzyme. Transcription termination factor. | [43] |
| F4L | 39.6 | Early | Ribonucleotide reductase small subunit. | [39] |
| J6R | 39.7 | Early Late | DNA-dependent RNA polymerase subunit rpo147. | [44] |
| A12L | 40.2 | Late | Core protein. | [45] |
| C4L | 41.2 | -- | Inhibits NF-kB activation and promotes virus virulence | [46] |
| B2R | 42.9 | -- | Hypothetical protein. Forms part of the envelope of the extracellular virus. | [47] |
| CSN3 | 43.4 | -- | Lactaptin. Proapoptotic fragment of the human kappa-casein gene, inserted in the VACV genome (3) | [10] |
| F1L | 44.2 | Early | Hypothetical protein. Suppressor of NLR family proteins involved in IL-1β activation. | [48] |
| D13L | 44.4 | Late | Rifampicin target. Rifampicin resistance. | [49] |
| A3L | 44.6 | Late | P4b precursor of core protein 4b. Critical for proper formation of the core wall and nucleocapsid. | [50] |
| D5R | 44.6 | Early Late | NTPase. DNA primase. | [51] |
| A10L | 46.0 | Late | Precursor p4a of core protein 4a. | [52] |
| A37R | 46.5 | -- | Hypothetical protein. Regulate the release of cell-associated virions. | [53] |
| A24R | 47.9 | Early Late | DNA-dependent RNA polymerase subunit rpo132. | [54] |
| F12L | 48.3 | Early | EEV maturation protein. Facilitates the transport of intracellular viral particles to the cell membrane. | [55] |
| A46R | 48.4 | -- | Toll/IL1-receptor. Antagonists of the activation of the proinflammatory transcription factor NF-κB. | [56] |
| A35R | 49.4 | -- | Hypothetical protein. Inhibitor of MHC class II-restricted antigen presentation. | [57] |
| M1L | 50.2 | Early | Ankyrin-like protein. Apoptosis inhibitor | [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, D.V.; Vasileva, N.S.; Dymova, M.A.; Mishinov, S.V.; Savinovskaya, Y.I.; Ageenko, A.B.; Dome, A.S.; Zinchenko, N.D.; Stepanov, G.A.; Kochneva, G.V.; et al. Transcriptome Changes in Glioma Cells upon Infection with the Oncolytic Virus VV-GMCSF-Lact. Cells 2023, 12, 2616. https://doi.org/10.3390/cells12222616
Semenov DV, Vasileva NS, Dymova MA, Mishinov SV, Savinovskaya YI, Ageenko AB, Dome AS, Zinchenko ND, Stepanov GA, Kochneva GV, et al. Transcriptome Changes in Glioma Cells upon Infection with the Oncolytic Virus VV-GMCSF-Lact. Cells. 2023; 12(22):2616. https://doi.org/10.3390/cells12222616
Chicago/Turabian StyleSemenov, Dmitriy V., Natalia S. Vasileva, Maya A. Dymova, Sergey V. Mishinov, Yulya I. Savinovskaya, Alisa B. Ageenko, Anton S. Dome, Nikita D. Zinchenko, Grigory A. Stepanov, Galina V. Kochneva, and et al. 2023. "Transcriptome Changes in Glioma Cells upon Infection with the Oncolytic Virus VV-GMCSF-Lact" Cells 12, no. 22: 2616. https://doi.org/10.3390/cells12222616
APA StyleSemenov, D. V., Vasileva, N. S., Dymova, M. A., Mishinov, S. V., Savinovskaya, Y. I., Ageenko, A. B., Dome, A. S., Zinchenko, N. D., Stepanov, G. A., Kochneva, G. V., Richter, V. A., & Kuligina, E. V. (2023). Transcriptome Changes in Glioma Cells upon Infection with the Oncolytic Virus VV-GMCSF-Lact. Cells, 12(22), 2616. https://doi.org/10.3390/cells12222616