
HLA-Homozygous iPSC-Derived Mesenchymal Stem Cells Rescue Rotenone-Induced Experimental Leber’s Hereditary Optic Neuropathy-like Models In Vitro and In Vivo

 , ,
, ,  , ,
, ,  and
and
Abstract
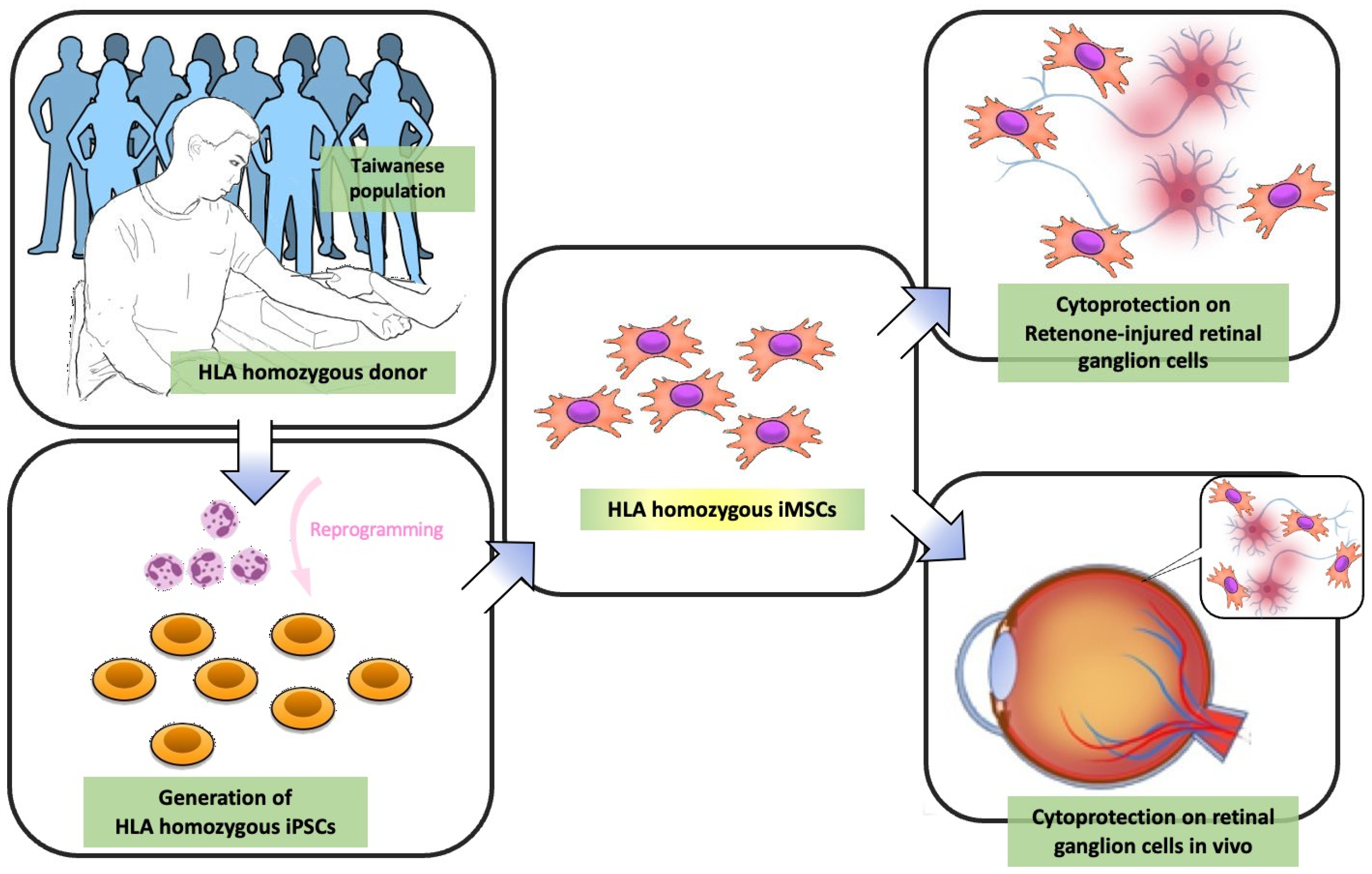
:1. Introduction
2. Materials and Methods
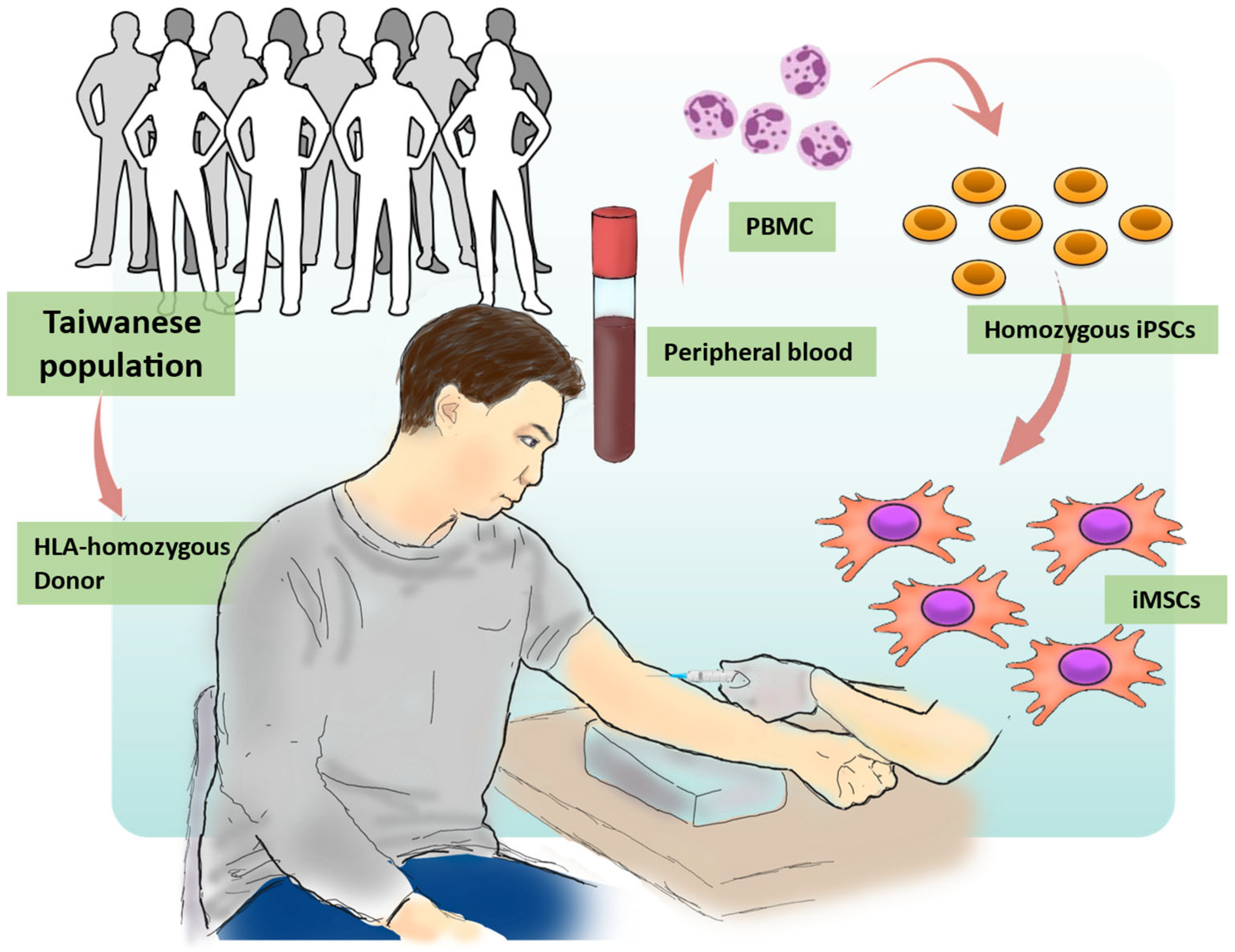
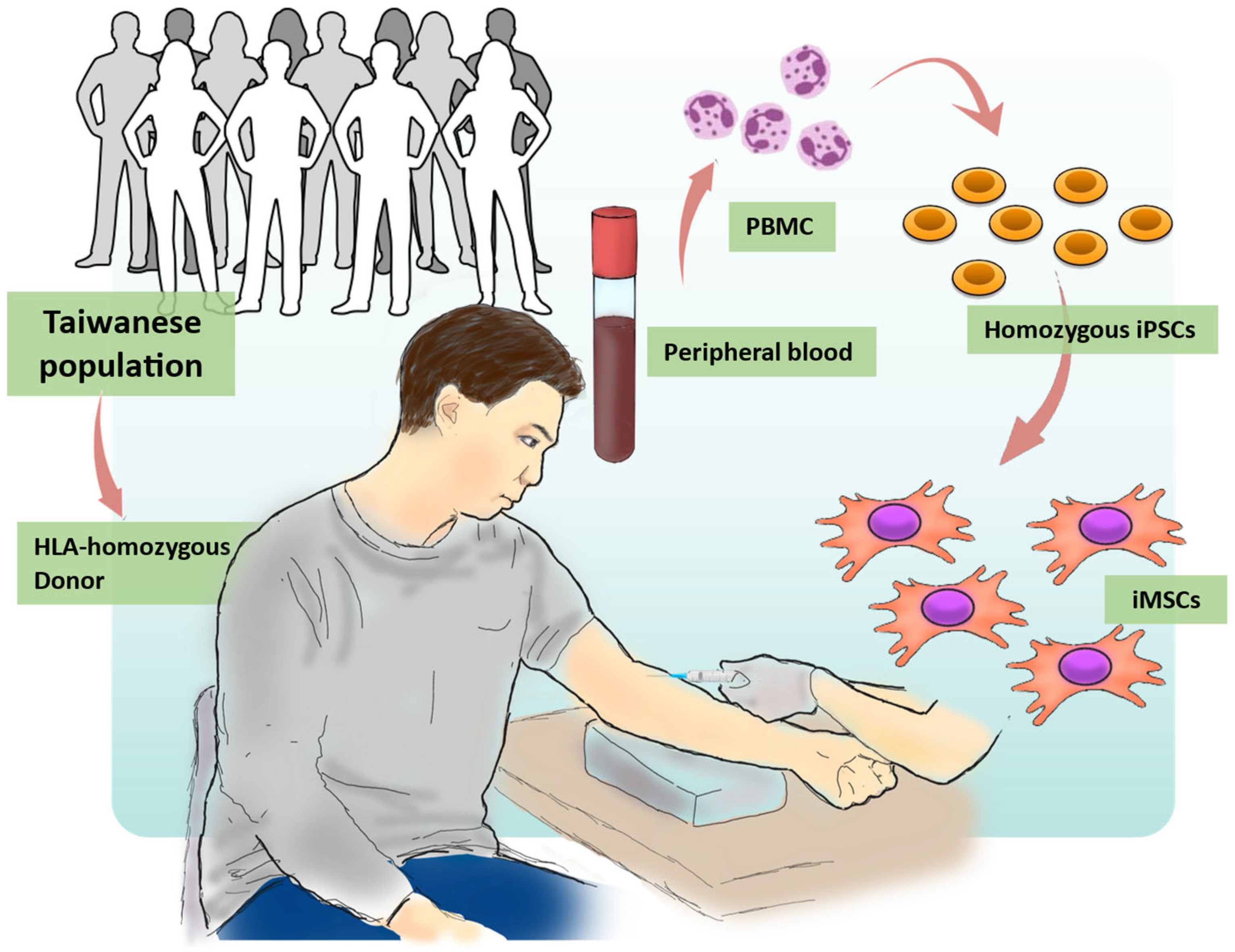
2.1. Generation of Human iPSCs from the HLA Homozygous Donor
2.2. Alkaline Phosphatase Staining
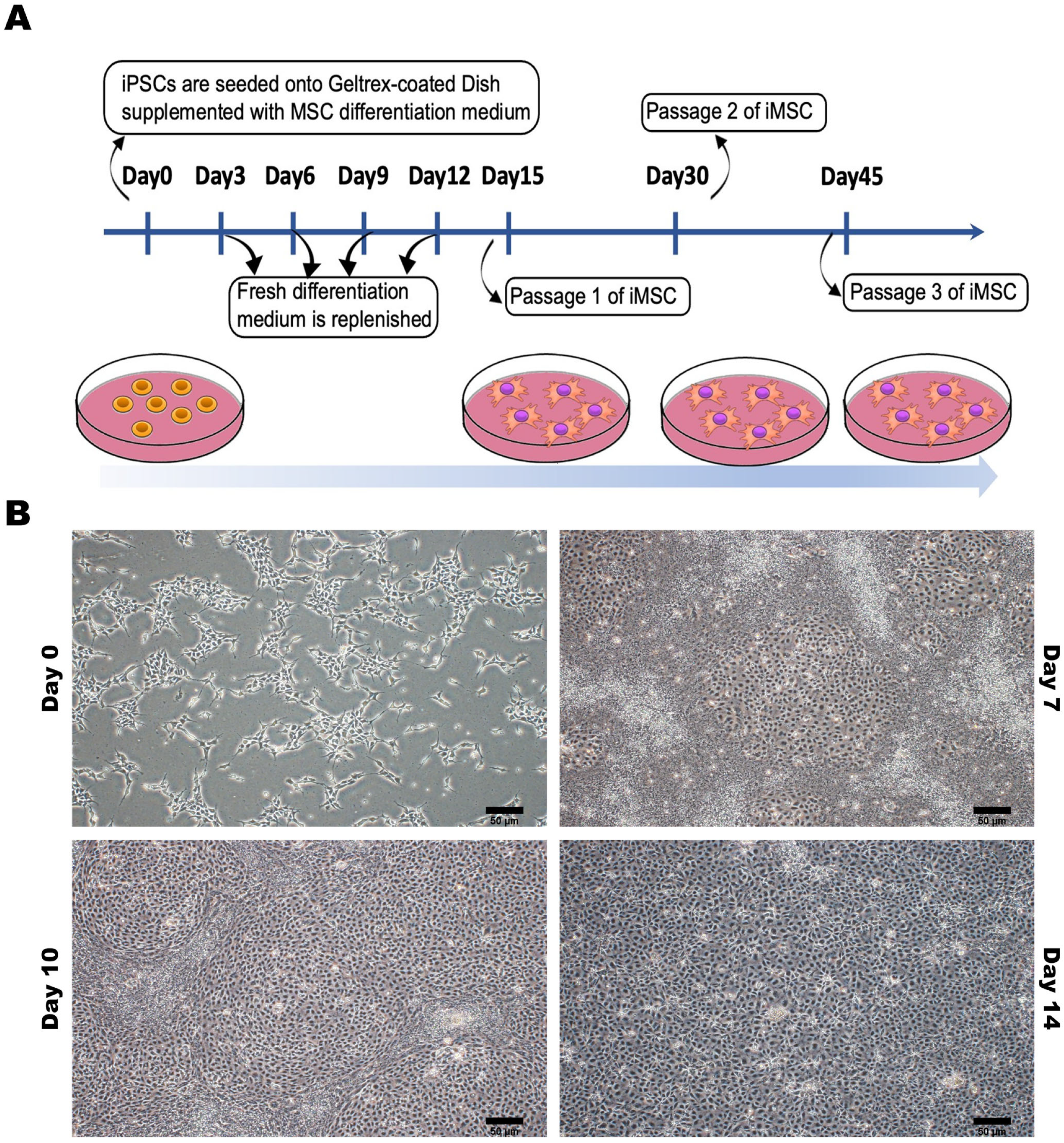
2.3. Human iPSC-Derived Mesenchymal Stem Cell Differentiation and Culture
2.4. In Vitro Treatment of Rotenone in Cultured Retinal Ganglion Cells
2.5. Measurement of Intracellular Reactive Oxygen Species
2.6. Mitochondrial Reactive Oxygen Species Assay
2.7. Immunofluorescence Staining
2.8. Experimental Animals and Intravitreal Injection of Rotenone
2.9. Spectral Domain OCT
2.10. Electroretinography (ERG)
2.11. Statistical Analysis
3. Results
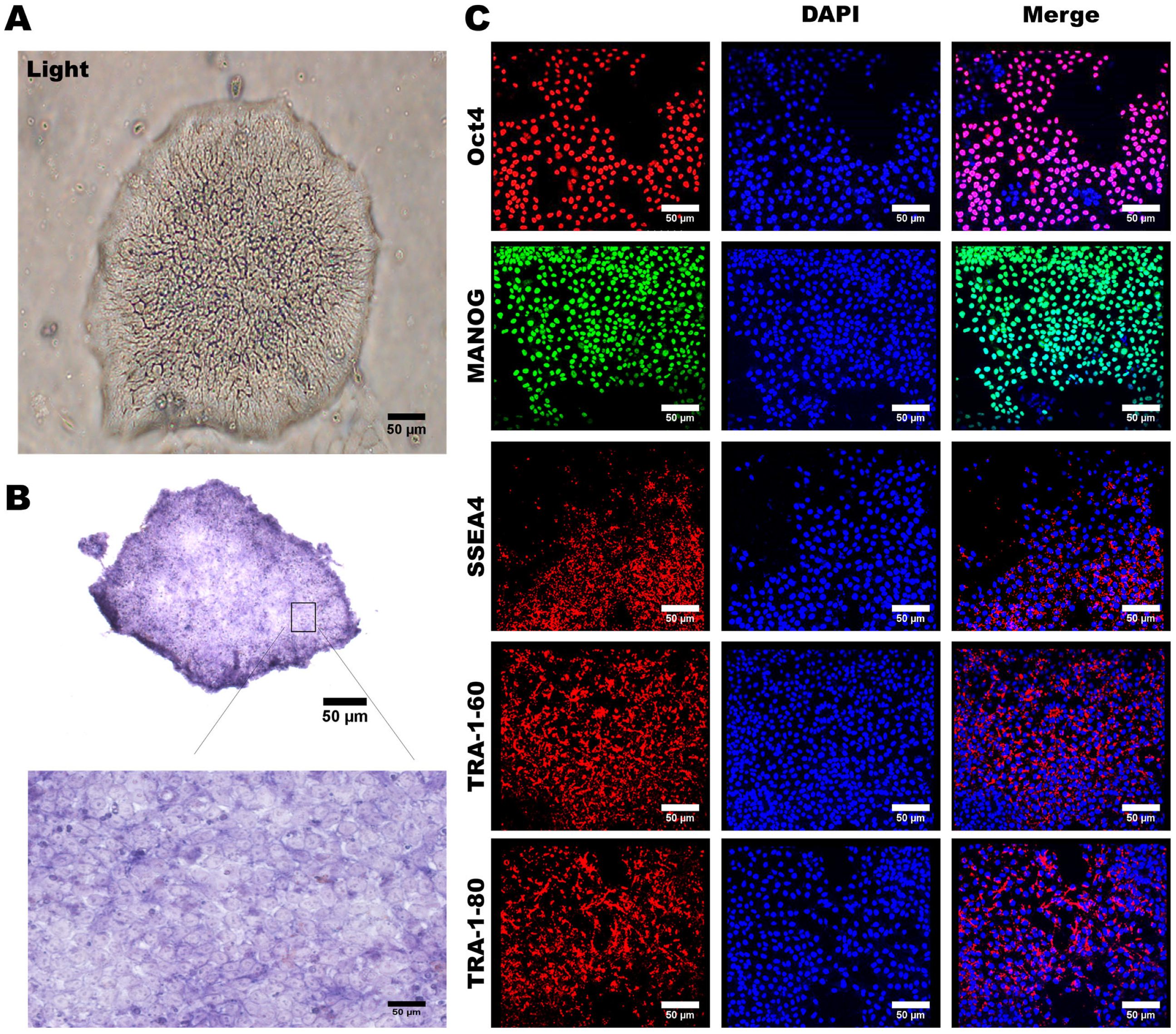
3.1. Generation of Human-Induced Pluripotent Stem Cells (hiPSCs)
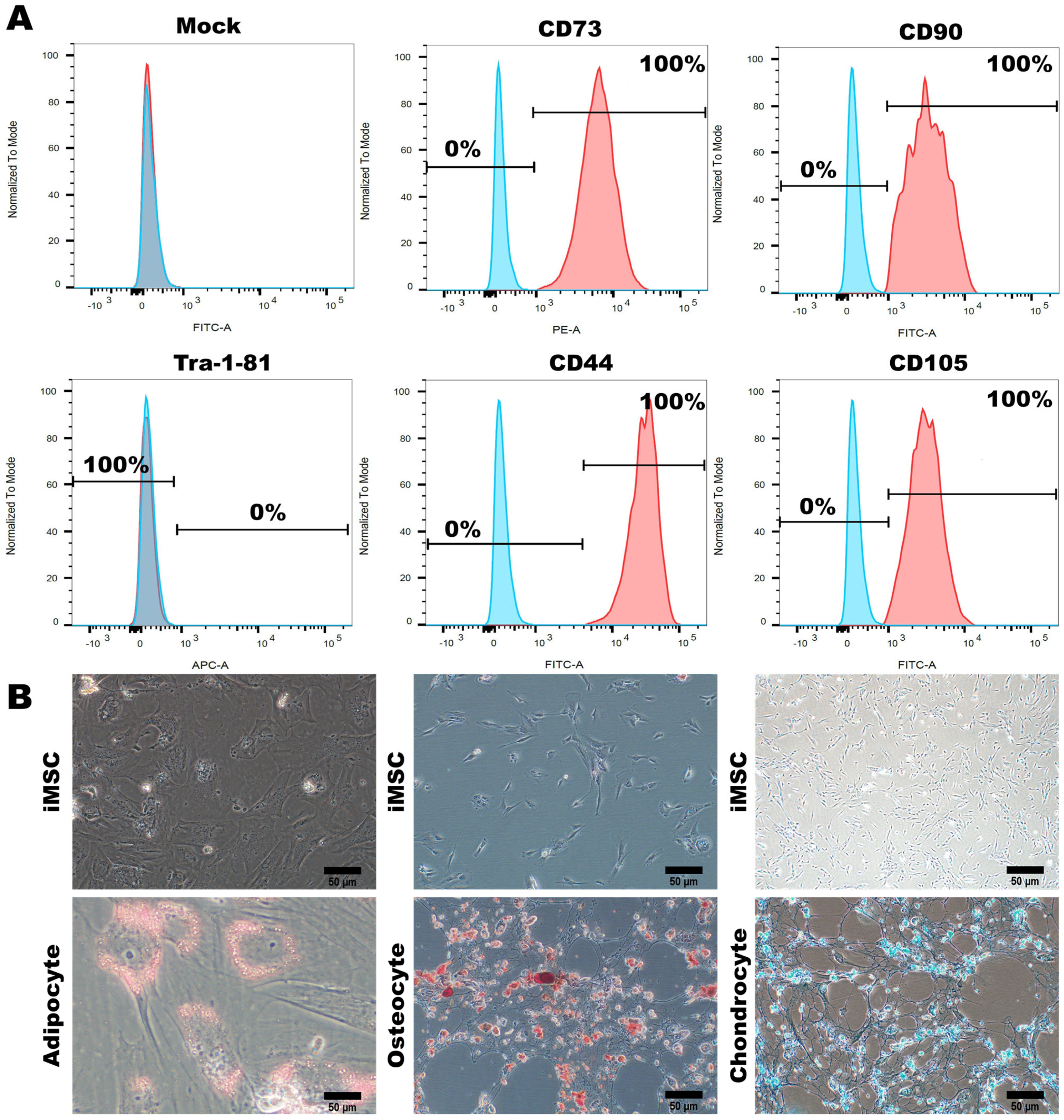
3.2. Characterization of MSC-Specific Features in HLA Homozygous iMSCs
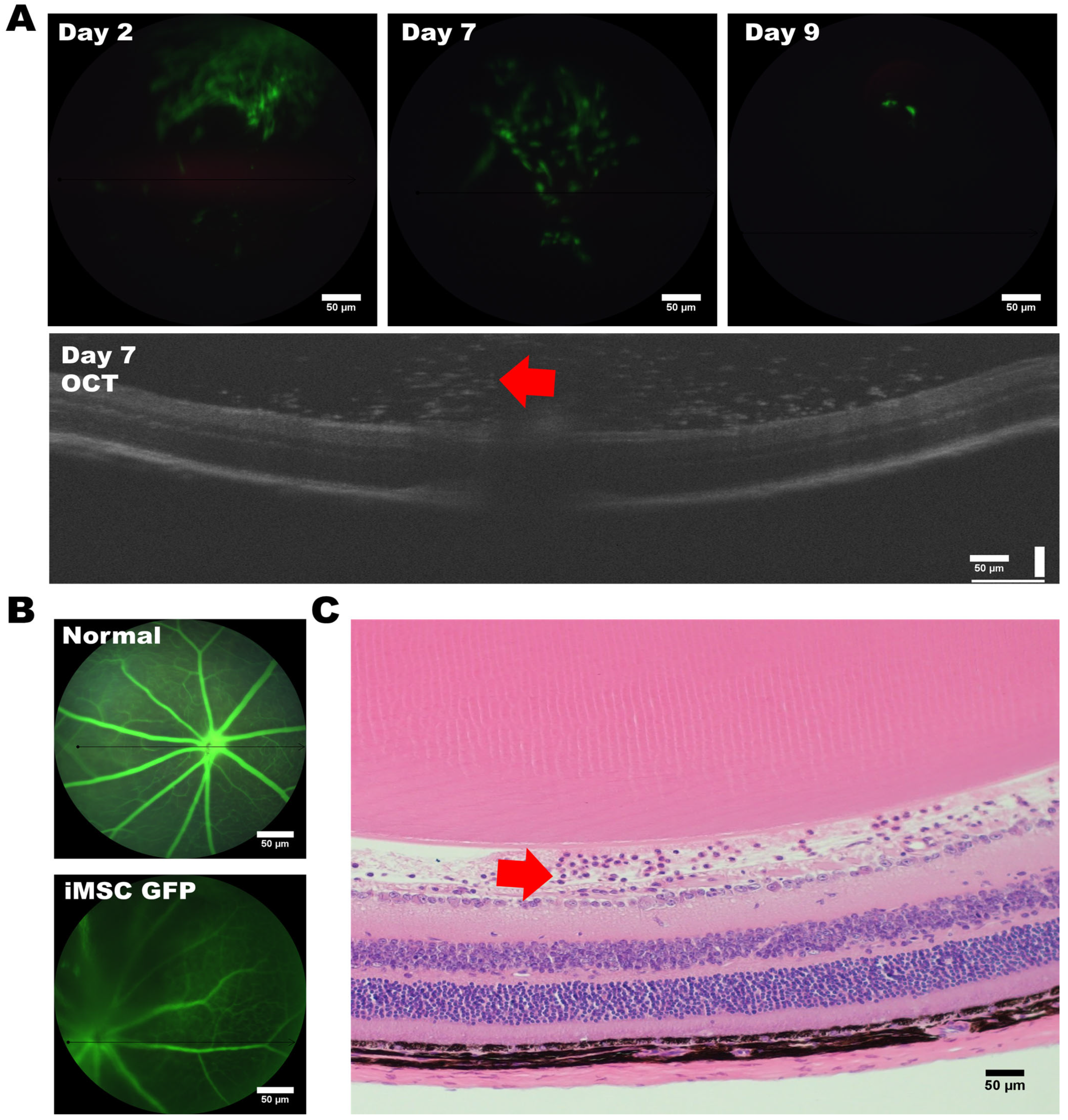
3.3. Assessing the Tumorigenic Risk and the Fate of HLA-Homozygous iMSCs in the Xenograft-Bearing Mice
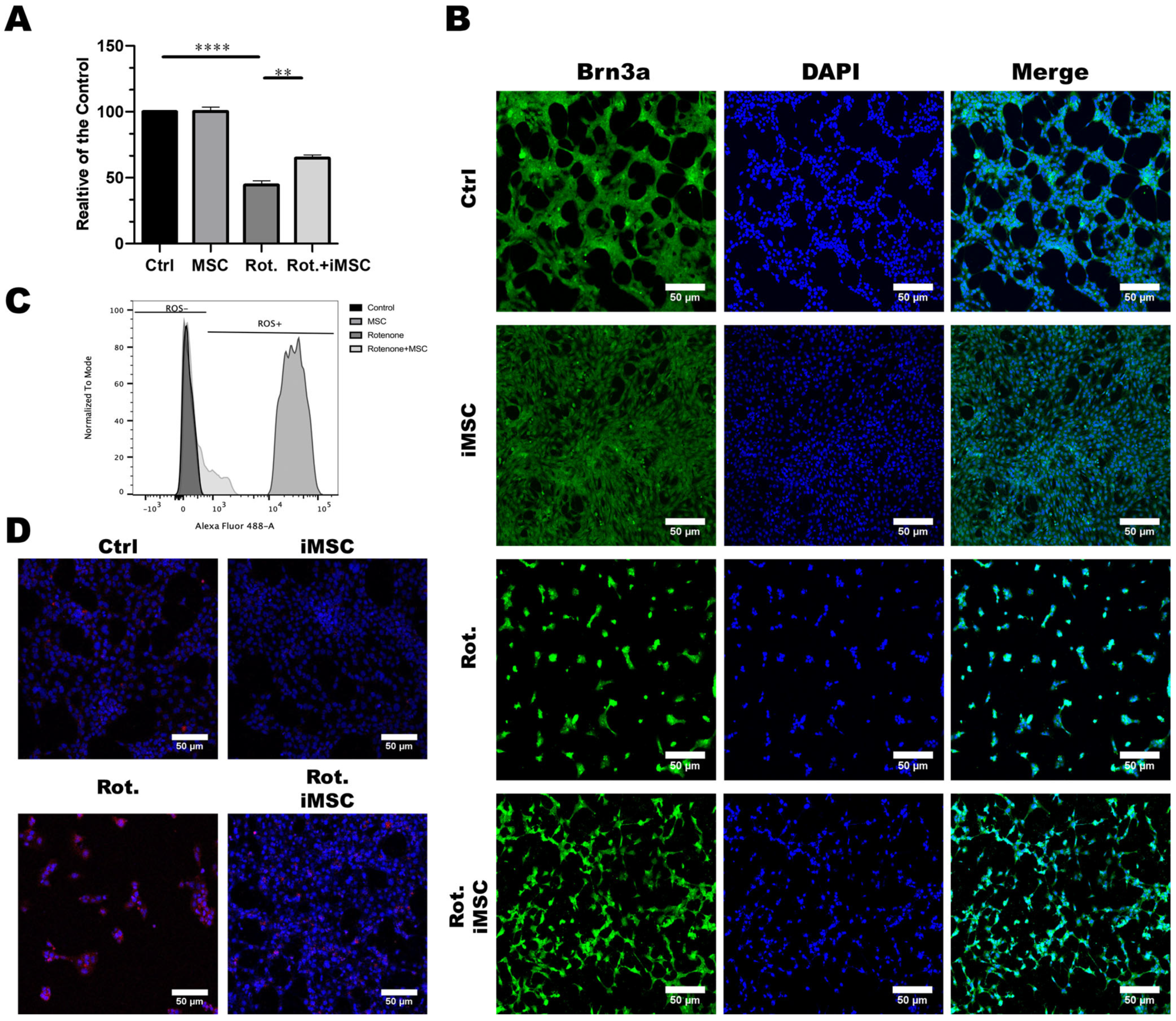
3.4. iPSC-MSCs Safeguard Retinal Ganglion Cells from Rotenone-Induced Damage In Vitro
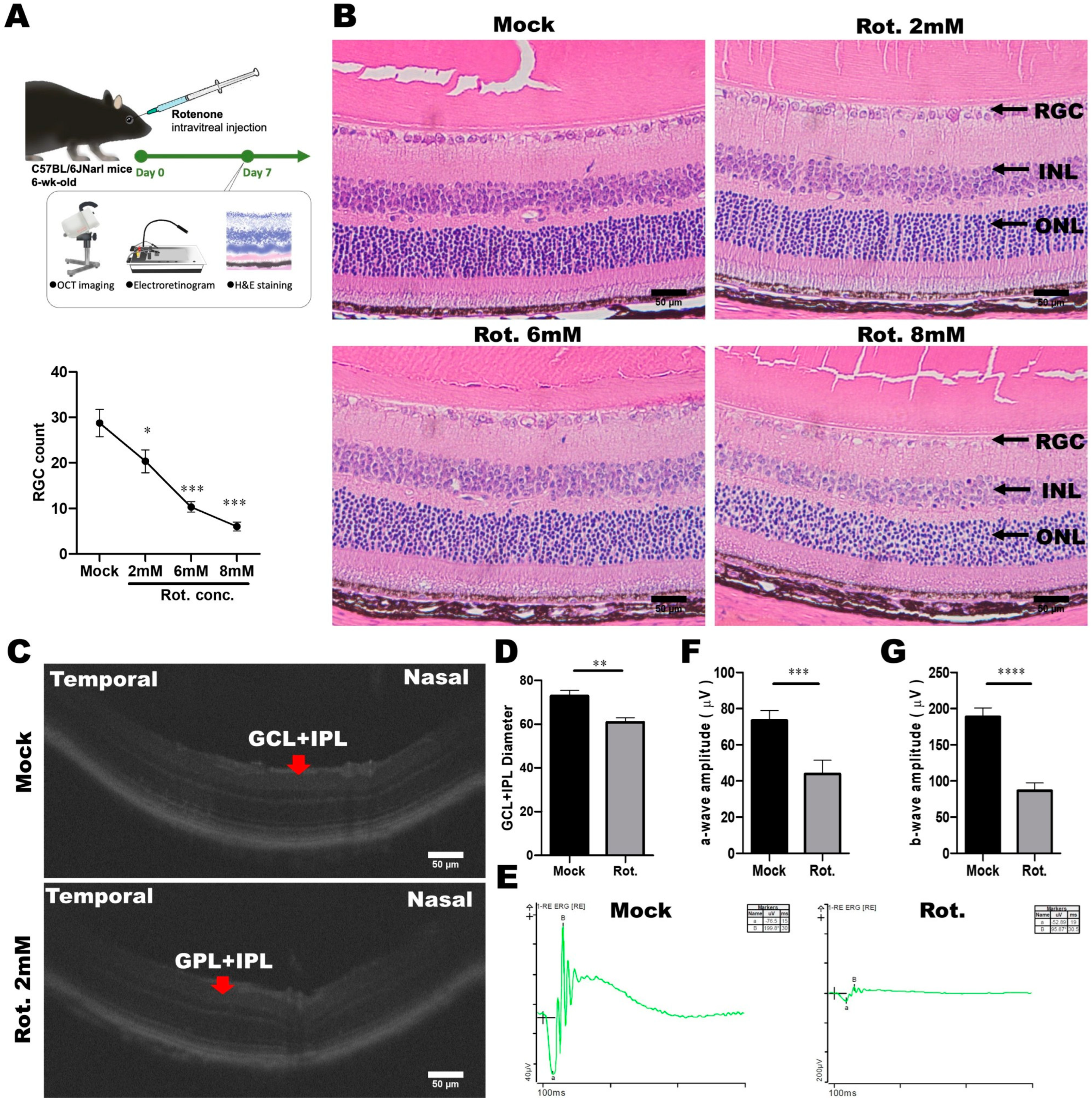
3.5. Establishment and Validation of the Rotenone-Induced LHON Mouse Model
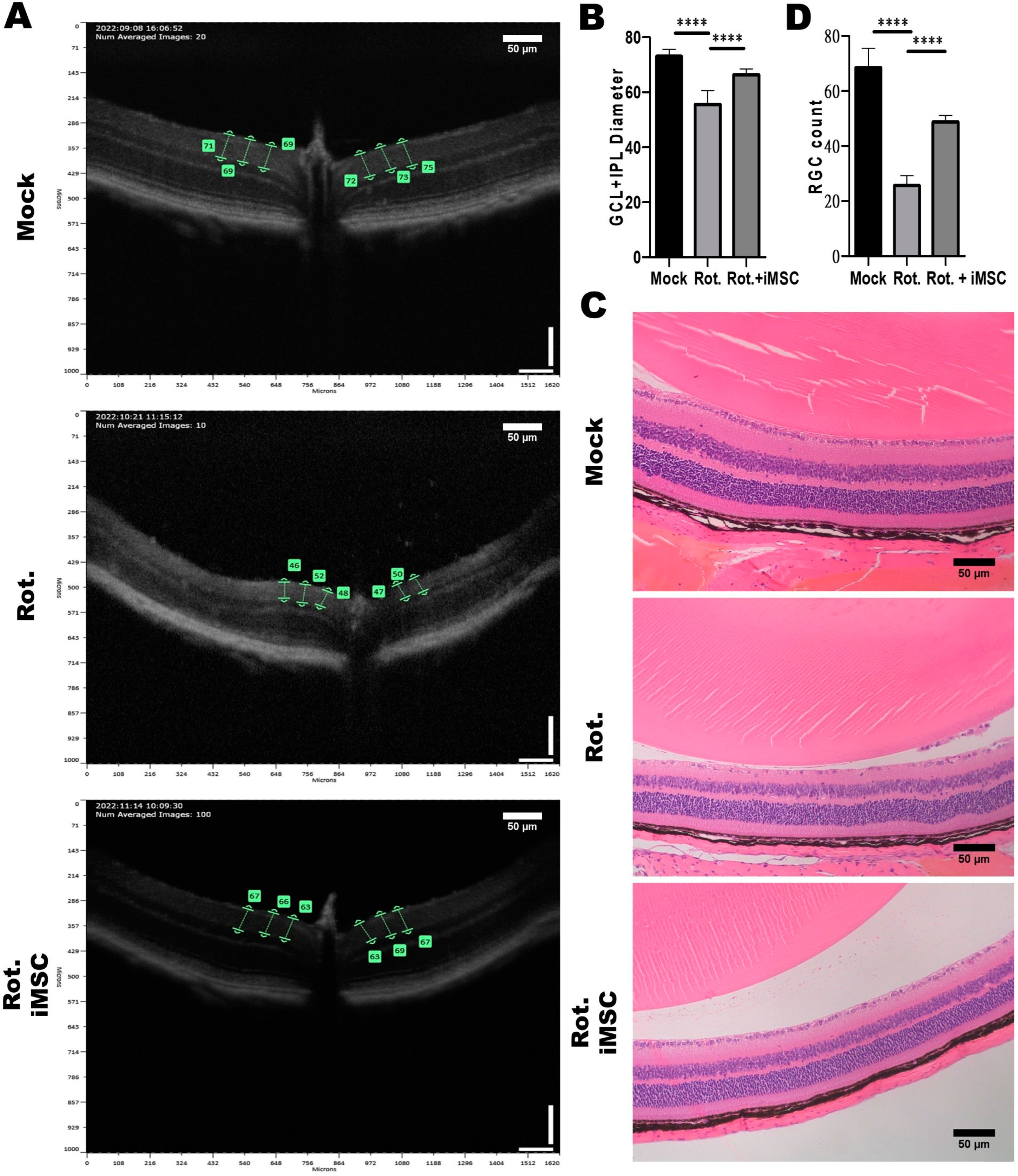
3.6. Evaluating the Treatment Efficacy of iMSC Transplantation in Rotenone-Injured Eyes Using Optical Coherence Tomography and a Histological Examination
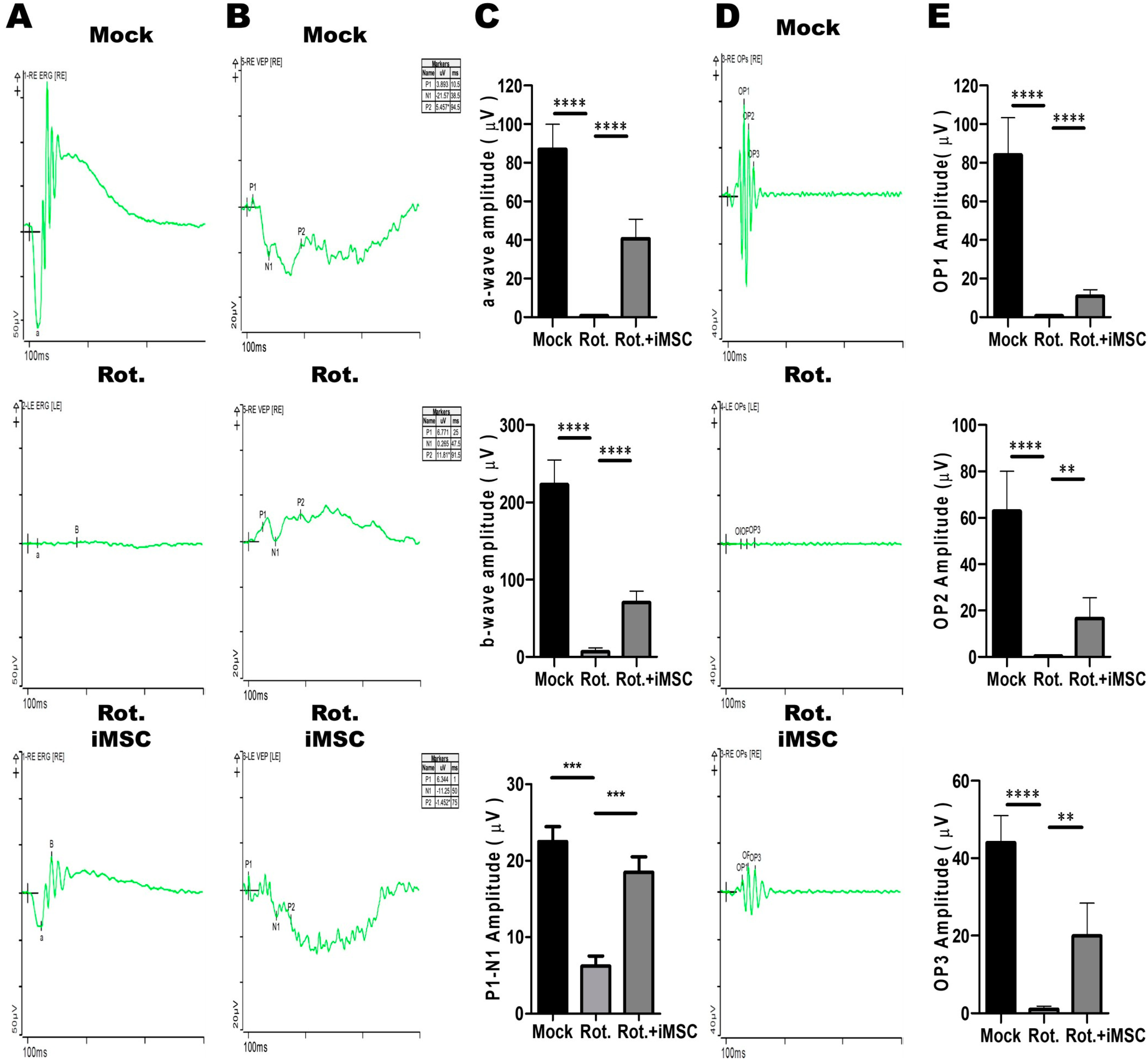
3.7. Evaluating the Treatment Efficacy of iMSC Transplantation in Rotenone-Injured Eyes Using the Electroretinogram
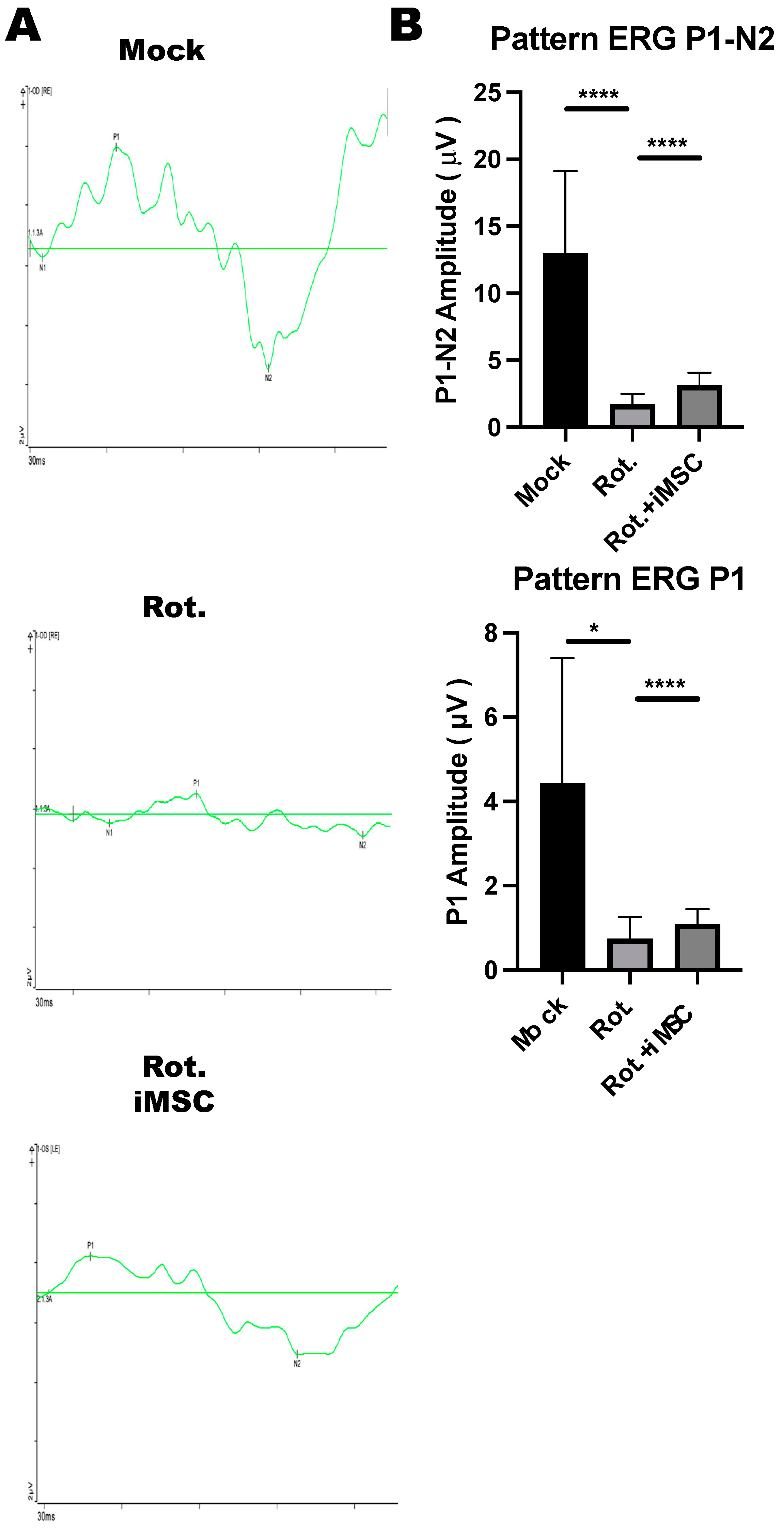
3.8. iMSC Transplantation Partially Rescued Retinal Ganglion Cell Functions in Rotenone-Injured Eyes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gimble, J.M.; Zvonic, S.; Floyd, Z.E.; Kassem, M.; Nuttall, M.E. Playing with bone and fat. J. Cell. Biochem. 2006, 98, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Dezawa, M.; Kanno, H.; Hoshino, M.; Cho, H.; Matsumoto, N.; Itokazu, Y.; Tajima, N.; Yamada, H.; Sawada, H.; Ishikawa, H.; et al. Specific induction of neuronal cells from bone marrow stromal cells and application for autologous transplantation. J. Clin. Investig. 2004, 113, 1701–1710. [Google Scholar] [CrossRef]
- Herzog, E.L.; Chai, L.; Krause, D.S. Plasticity of marrow-derived stem cells. Blood 2003, 102, 3483–3493. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Prockop, D.J.; Spees, J.L. Non-hematopoietic bone marrow stem cells: Molecular control of expansion and differentiation. Exp. Cell Res. 2005, 306, 330–335. [Google Scholar] [CrossRef]
- Chen, Q.; Long, Y.; Yuan, X.; Zou, L.; Sun, J.; Chen, S.; Perez-Polo, J.R.; Yang, K. Protective effects of bone marrow stromal cell transplantation in injured rodent brain: Synthesis of neurotrophic factors. J. Neurosci. Res. 2005, 80, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Cancedda, R.; Bianchi, G.; Derubeis, A.; Quarto, R. Cell therapy for bone disease: A review of current status. Stem Cells 2003, 21, 610–619. [Google Scholar] [CrossRef]
- Liu, X.; Robbins, S.; Wang, X.; Virk, S.; Schuck, K.; Deveza, L.A.; Oo, W.M.; Carmichael, K.; Antony, B.; Eckstein, F.; et al. Efficacy and cost-effectiveness of Stem Cell injections for symptomatic relief and strUctural improvement in people with Tibiofemoral knee OsteoaRthritis: Protocol for a randomised placebo-controlled trial (the SCUlpTOR trial). BMJ Open 2021, 11, e056382. [Google Scholar] [CrossRef]
- Kelly, K.; Rasko, J.E.J. Mesenchymal Stromal Cells for the Treatment of Graft Versus Host Disease. Front. Immunol. 2021, 12, 761616. [Google Scholar] [CrossRef]
- Gnecchi, M.; Melo, L.G. Bone marrow-derived mesenchymal stem cells: Isolation, expansion, characterization, viral transduction, and production of conditioned medium. Methods Mol. Biol. 2009, 482, 281–294. [Google Scholar] [CrossRef]
- Beyer Nardi, N.; da Silva Meirelles, L. Mesenchymal stem cells: Isolation, in vitro expansion and characterization. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 249–282. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Huang, K.C.; Wang, M.L.; Chen, S.J.; Kuo, J.C.; Wang, W.J.; Nhi Nguyen, P.N.; Wahlin, K.J.; Lu, J.F.; Tran, A.A.; Shi, M.; et al. Morphological and Molecular Defects in Human Three-Dimensional Retinal Organoid Model of X-Linked Juvenile Retinoschisis. Stem Cell Rep. 2019, 13, 906–923. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.S.; Chien, Y.; Lin, Y.Y.; Tsai, P.H.; Chou, S.J.; Yarmishyn, A.A.; Rastegari, E.; Wang, T.X.; Leu, H.B.; Yang, Y.P.; et al. Dual DNA Transfection Using 1,6-Hexanedithiol-Conjugated Maleimide-Functionalized PU-PEI600 For Gene Correction in a Patient iPSC-Derived Fabry Cardiomyopathy Model. Front. Cell Dev. Biol. 2021, 9, 634190. [Google Scholar] [CrossRef]
- Lee, C.Y.; Huang, C.H.; Rastegari, E.; Rengganaten, V.; Liu, P.C.; Tsai, P.H.; Chin, Y.F.; Wu, J.R.; Chiou, S.H.; Teng, Y.C.; et al. Tumor Necrosis Factor-Alpha Exacerbates Viral Entry in SARS-CoV2-Infected iPSC-Derived Cardiomyocytes. Int. J. Mol. Sci. 2021, 22, 9869. [Google Scholar] [CrossRef]
- Lien, C.Y.; Chen, T.T.; Tsai, E.T.; Hsiao, Y.J.; Lee, N.; Gao, C.E.; Yang, Y.P.; Chen, S.J.; Yarmishyn, A.A.; Hwang, D.K.; et al. Recognizing the Differentiation Degree of Human Induced Pluripotent Stem Cell-Derived Retinal Pigment Epithelium Cells Using Machine Learning and Deep Learning-Based Approaches. Cells 2023, 12, 211. [Google Scholar] [CrossRef]
- Tsai, P.H.; Sun, J.R.; Chien, Y.; Chan, M.S.; Khor, W.; Yang, H.C.; Huang, C.H.; Hsiung, C.N.; Hwa, T.Y.; Lin, Y.Y.; et al. Modifications of lipid pathways restrict SARS-CoV-2 propagation in human induced pluripotent stem cell-derived 3D airway organoids. J. Adv. Res. 2023. [Google Scholar] [CrossRef]
- Zomer, H.D.; Vidane, A.S.; Goncalves, N.N.; Ambrosio, C.E. Mesenchymal and induced pluripotent stem cells: General insights and clinical perspectives. Stem Cells Cloning 2015, 8, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kimbrel, E.A.; Ijichi, K.; Paul, D.; Lazorchak, A.S.; Chu, J.; Kouris, N.A.; Yavanian, G.J.; Lu, S.J.; Pachter, J.S.; et al. Human ESC-Derived MSCs Outperform Bone Marrow MSCs in the Treatment of an EAE Model of Multiple Sclerosis. Stem Cell Rep. 2021, 16, 370–371. [Google Scholar] [CrossRef]
- Hawkins, K.E.; Corcelli, M.; Dowding, K.; Ranzoni, A.M.; Vlahova, F.; Hau, K.L.; Hunjan, A.; Peebles, D.; Gressens, P.; Hagberg, H.; et al. Embryonic Stem Cell-Derived Mesenchymal Stem Cells (MSCs) Have a Superior Neuroprotective Capacity Over Fetal MSCs in the Hypoxic-Ischemic Mouse Brain. Stem Cells Transl. Med. 2018, 7, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Soontararak, S.; Chow, L.; Johnson, V.; Coy, J.; Wheat, W.; Regan, D.; Dow, S. Mesenchymal Stem Cells (MSC) Derived from Induced Pluripotent Stem Cells (iPSC) Equivalent to Adipose-Derived MSC in Promoting Intestinal Healing and Microbiome Normalization in Mouse Inflammatory Bowel Disease Model. Stem Cells Transl. Med. 2018, 7, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Y.; Zhao, B.; Niu, X.; Hu, B.; Li, Q.; Zhang, J.; Ding, J.; Chen, Y.; Wang, Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res. Ther. 2017, 8, 64. [Google Scholar] [CrossRef]
- Ozay, E.I.; Vijayaraghavan, J.; Gonzalez-Perez, G.; Shanthalingam, S.; Sherman, H.L.; Garrigan, D.T., Jr.; Chandiran, K.; Torres, J.A.; Osborne, B.A.; Tew, G.N.; et al. Cymerus iPSC-MSCs significantly prolong survival in a pre-clinical, humanized mouse model of Graft-vs-host disease. Stem Cell Res. 2019, 35, 101401. [Google Scholar] [CrossRef]
- Sabapathy, V.; Kumar, S. hiPSC-derived iMSCs: NextGen MSCs as an advanced therapeutically active cell resource for regenerative medicine. J. Cell. Mol. Med. 2016, 20, 1571–1588. [Google Scholar] [CrossRef]
- Fraser, J.A.; Biousse, V.; Newman, N.J. The neuro-ophthalmology of mitochondrial disease. Surv. Ophthalmol. 2010, 55, 299–334. [Google Scholar] [CrossRef]
- Carelli, V.; La Morgia, C.; Valentino, M.L.; Rizzo, G.; Carbonelli, M.; De Negri, A.M.; Sadun, F.; Carta, A.; Guerriero, S.; Simonelli, F.; et al. Idebenone treatment in Leber’s hereditary optic neuropathy. Brain 2011, 134, e188. [Google Scholar] [CrossRef]
- Johnson, T.V.; Bull, N.D.; Hunt, D.P.; Marina, N.; Tomarev, S.I.; Martin, K.R. Neuroprotective effects of intravitreal mesenchymal stem cell transplantation in experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2051–2059. [Google Scholar] [CrossRef]
- Beretta, S.; Wood, J.P.; Derham, B.; Sala, G.; Tremolizzo, L.; Ferrarese, C.; Osborne, N.N. Partial mitochondrial complex I inhibition induces oxidative damage and perturbs glutamate transport in primary retinal cultures. Relevance to Leber Hereditary Optic Neuropathy (LHON). Neurobiol. Dis. 2006, 24, 308–317. [Google Scholar] [CrossRef]
- Finnegan, L.K.; Chadderton, N.; Kenna, P.F.; Palfi, A.; Carty, M.; Bowie, A.G.; Millington-Ward, S.; Farrar, G.J. SARM1 Ablation Is Protective and Preserves Spatial Vision in an In Vivo Mouse Model of Retinal Ganglion Cell Degeneration. Int. J. Mol. Sci. 2022, 23, 1606. [Google Scholar] [CrossRef] [PubMed]
- Mansergh, F.C.; Chadderton, N.; Kenna, P.F.; Gobbo, O.L.; Farrar, G.J. Cell therapy using retinal progenitor cells shows therapeutic effect in a chemically-induced rotenone mouse model of Leber hereditary optic neuropathy. Eur. J. Hum. Genet. 2014, 22, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, Y.; Inagaki, S.; Aoshima, K.; Iwata, Y.; Nakamura, S.; Hara, H.; Shimazawa, M. Involvement of endoplasmic reticulum stress in rotenone-induced leber hereditary optic neuropathy model and the discovery of new therapeutic agents. J. Pharmacol. Sci. 2021, 147, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, L.; Philip, A.L.; Martinez, J.C.; Guttierez, J.C.; Marella, M.; Patki, G.; Matsuno-Yagi, A.; Yagi, T.; Thomas, B.B. Long-term evaluation of Leber’s hereditary optic neuropathy-like symptoms in rotenone administered rats. Neurosci. Lett. 2015, 585, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Hynes, K.; Menicanin, D.; Mrozik, K.; Gronthos, S.; Bartold, P.M. Generation of functional mesenchymal stem cells from different induced pluripotent stem cell lines. Stem Cells Dev. 2014, 23, 1084–1096. [Google Scholar] [CrossRef]
- Nakagawa, M.; Takizawa, N.; Narita, M.; Ichisaka, T.; Yamanaka, S. Promotion of direct reprogramming by transformation-deficient Myc. Proc. Natl. Acad. Sci. USA 2010, 107, 14152–14157. [Google Scholar] [CrossRef]
- Singh, U.; Quintanilla, R.H.; Grecian, S.; Gee, K.R.; Rao, M.S.; Lakshmipathy, U. Novel live alkaline phosphatase substrate for identification of pluripotent stem cells. Stem Cell Rev. Rep. 2012, 8, 1021–1029. [Google Scholar] [CrossRef]
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced pluripotent stem cell technology: A decade of progress. Nat. Rev. Drug Discov. 2017, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Paik, D.T.; Chandy, M.; Wu, J.C. Patient and disease–specific induced pluripotent stem cells for discovery of personalized cardiovascular drugs and therapeutics. Pharmacol. Rev. 2020, 72, 320–342. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.C.; Opelz, G.; McGarvey, C.J.; Weil, E.J.; Chakkera, H.A. The risk of transplant failure with HLA mismatch in first adult kidney allografts from deceased donors. Transplantation 2016, 100, 1094. [Google Scholar] [CrossRef]
- Aron Badin, R.; Bugi, A.; Williams, S.; Vadori, M.; Michael, M.; Jan, C.; Nassi, A.; Lecourtois, S.; Blancher, A.; Cozzi, E. MHC matching fails to prevent long-term rejection of iPSC-derived neurons in non-human primates. Nat. Commun. 2019, 10, 4357. [Google Scholar] [CrossRef]
- Maeda, T.; Sugita, S.; Kurimoto, Y.; Takahashi, M. Trends of stem cell therapies in age-related macular degeneration. J. Clin. Med. 2021, 10, 1785. [Google Scholar] [CrossRef]
- Sugita, S.; Mandai, M.; Hirami, Y.; Takagi, S.; Maeda, T.; Fujihara, M.; Matsuzaki, M.; Yamamoto, M.; Iseki, K.; Hayashi, N. HLA-matched allogeneic iPS cells-derived RPE transplantation for macular degeneration. J. Clin. Med. 2020, 9, 2217. [Google Scholar] [CrossRef] [PubMed]
- Manickam, A.H.; Michael, M.J.; Ramasamy, S. Mitochondrial genetics and therapeutic overview of Leber’s hereditary optic neuropathy. Indian J. Ophthalmol. 2017, 65, 1087. [Google Scholar]
- Stein, L.R.; Imai, S.-I. The dynamic regulation of NAD metabolism in mitochondria. Trends Endocrinol. Metab. 2012, 23, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Meyerson, C.; Van Stavern, G.; McClelland, C. Leber hereditary optic neuropathy: Current perspectives. Clin. Ophthalmol. 2015, 9, 1165–1176. [Google Scholar]
- Limoli, P.G.; Vingolo, E.M.; Morales, M.U.; Nebbioso, M.; Limoli, C. Preliminary study on electrophysiological changes after cellular autograft in age-related macular degeneration. Medicine 2014, 93, e355. [Google Scholar] [CrossRef]
- Oner, A.; Gonen, Z.B.; Sevim, D.G.; Smim Kahraman, N.; Unlu, M. Suprachoroidal Adipose Tissue-Derived Mesenchymal Stem Cell Implantation in Patients with Dry-Type Age-Related Macular Degeneration and Stargardt’s Macular Dystrophy: 6-Month Follow-Up Results of a Phase 2 Study. Cell. Reprogram. 2018, 20, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.N.; Levy, S. Stem Cell Ophthalmology Treatment Study: Bone marrow derived stem cells in the treatment of Retinitis Pigmentosa. Stem Cell Investig. 2018, 5, 18. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Kong, J.; Dong, M.; Duan, H.; Chen, S. Therapeutic efficacy of neural stem cells originating from umbilical cord-derived mesenchymal stem cells in diabetic retinopathy. Sci. Rep. 2017, 7, 408. [Google Scholar] [CrossRef]
- Mead, B.; Hill, L.J.; Blanch, R.J.; Ward, K.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Mesenchymal stromal cell-mediated neuroprotection and functional preservation of retinal ganglion cells in a rodent model of glaucoma. Cytotherapy 2016, 18, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Nagai, A.; Kim, W.K.; Lee, H.J.; Jeong, H.S.; Kim, K.S.; Hong, S.H.; Park, I.H.; Kim, S.U. Multilineage potential of stable human mesenchymal stem cell line derived from fetal marrow. PLoS ONE 2007, 2, e1272. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.W.; Kim, S.S.; Lee, S.Y.; Lee, H.S.; Kim, H.S.; Lee, Y.D.; Suh-Kim, H. Mesenchymal stem cells promote proliferation of endogenous neural stem cells and survival of newborn cells in a rat stroke model. Exp. Mol. Med. 2008, 40, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Maltman, D.J.; Hardy, S.A.; Przyborski, S.A. Role of mesenchymal stem cells in neurogenesis and nervous system repair. Neurochem. Int. 2011, 59, 347–356. [Google Scholar] [CrossRef]
- Kemp, K.; Hares, K.; Mallam, E.; Heesom, K.J.; Scolding, N.; Wilkins, A. Mesenchymal stem cell-secreted superoxide dismutase promotes cerebellar neuronal survival. J. Neurochem. 2010, 114, 1569–1580. [Google Scholar] [CrossRef]
- Chen, Y.T.; Sun, C.K.; Lin, Y.C.; Chang, L.T.; Chen, Y.L.; Tsai, T.H.; Chung, S.Y.; Chua, S.; Kao, Y.H.; Yen, C.H.; et al. Adipose-derived mesenchymal stem cell protects kidneys against ischemia-reperfusion injury through suppressing oxidative stress and inflammatory reaction. J. Transl. Med. 2011, 9, 51. [Google Scholar] [CrossRef]
- Kim, H.S.; Choi, D.Y.; Yun, S.J.; Choi, S.M.; Kang, J.W.; Jung, J.W.; Hwang, D.; Kim, K.P.; Kim, D.W. Proteomic analysis of microvesicles derived from human mesenchymal stem cells. J. Proteome Res. 2012, 11, 839–849. [Google Scholar] [CrossRef]
- Tomasoni, S.; Longaretti, L.; Rota, C.; Morigi, M.; Conti, S.; Gotti, E.; Capelli, C.; Introna, M.; Remuzzi, G.; Benigni, A. Transfer of growth factor receptor mRNA via exosomes unravels the regenerative effect of mesenchymal stem cells. Stem Cells Dev. 2013, 22, 772–780. [Google Scholar] [CrossRef]
- Wang, S.P.; Wang, Z.H.; Peng, D.Y.; Li, S.M.; Wang, H.; Wang, X.H. Therapeutic effect of mesenchymal stem cells in rats with intracerebral hemorrhage: Reduced apoptosis and enhanced neuroprotection. Mol. Med. Rep. 2012, 6, 848–854. [Google Scholar] [CrossRef]
- Budoni, M.; Fierabracci, A.; Luciano, R.; Petrini, S.; Di Ciommo, V.; Muraca, M. The immunosuppressive effect of mesenchymal stromal cells on B lymphocytes is mediated by membrane vesicles. Cell Transplant. 2013, 22, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Hawryluk, G.W.; Mothe, A.; Wang, J.; Wang, S.; Tator, C.; Fehlings, M.G. An in vivo characterization of trophic factor production following neural precursor cell or bone marrow stromal cell transplantation for spinal cord injury. Stem Cells Dev. 2012, 21, 2222–2238. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.; Jimenez-Luna, C.; Perales-Adan, J.; Perazzoli, G.; Melguizo, C.; Prados, J. Differentiation of Human Mesenchymal Stem Cells towards Neuronal Lineage: Clinical Trials in Nervous System Disorders. Biomol. Ther. 2020, 28, 34–44. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Haplotypes | Frequency (%) | Sample Size | Population |
|---|---|---|---|---|
| 1 | A*33:03~B*58:01~DRB1*03:01 | 8.3610 | 46,628 | Taiwan |
| 2 | A*02:07~B*46:01~DRB1*09:01 | 4.4300 | 46,628 | Taiwan |
| 3 | A*11:01~B*15:02~DRB1*12:02 | 1.5000 | 46,628 | Taiwan |
| 4 | A*02:03~B*38:02~DRB1*16:02 | 1.4950 | 46,628 | Taiwan |
| 5 | A*30:01~B*13:02~DRB1*07:01 | 1.3310 | 46,628 | Taiwan |
| 6 | A*02:07~B*46:01~DRB1*08:03 | 1.2620 | 46,628 | Taiwan |
| 7 | A*02:01~B*40:01~DRB1*11:01 | 1.1050 | 46,628 | Taiwan |
| 8 | A*11:02~B*27:04~DRB1*12:02 | 0.9990 | 46,628 | Taiwan |
| 9 | A*11:01~B*40:01~DRB1*09:01 | 0.9290 | 46,628 | Taiwan |
| 10 | A*11:01~B*13:01~DRB1*15:01 | 0.9270 | 46,628 | Taiwan |
| 11 | A*11:01~B*46:01~DRB1*09:01 | 0.8700 | 46,628 | Taiwan |
| 12 | A*11:01~B*15:01~DRB1*04:06 | 0.8110 | 46,628 | Taiwan |
| 13 | A*11:01~B*40:01~DRB1*08:03 | 0.7670 | 46,628 | Taiwan |
| 14 | A*11:01~B*40:01~DRB1*11:01 | 0.7560 | 46,628 | Taiwan |
| 15 | A*11:01~B*40:01~DRB1*04:05 | 0.6770 | 46,628 | Taiwan |
| 16 | A*02:03~B*38:02~DRB1*08:03 | 0.6620 | 46,628 | Taiwan |
| 17 | A*11:01~B*13:01~DRB1*16:02 | 0.6360 | 46,628 | Taiwan |
| 18 | A*11:01~B*40:01~DRB1*12:01 | 0.6340 | 46,628 | Taiwan |
| 19 | A*24:02~B*40:01~DRB1*15:01 | 0.6330 | 46,628 | Taiwan |
| 20 | A*24:02~B*40:01~DRB1*09:01 | 0.5750 | 46,628 | Taiwan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, E.-T.; Peng, S.-Y.; Wu, Y.-R.; Lin, T.-C.; Chen, C.-Y.; Liu, Y.-H.; Tseng, Y.-H.; Hsiao, Y.-J.; Tseng, H.-C.; Lai, W.-Y.; et al. HLA-Homozygous iPSC-Derived Mesenchymal Stem Cells Rescue Rotenone-Induced Experimental Leber’s Hereditary Optic Neuropathy-like Models In Vitro and In Vivo. Cells 2023, 12, 2617. https://doi.org/10.3390/cells12222617
Tsai E-T, Peng S-Y, Wu Y-R, Lin T-C, Chen C-Y, Liu Y-H, Tseng Y-H, Hsiao Y-J, Tseng H-C, Lai W-Y, et al. HLA-Homozygous iPSC-Derived Mesenchymal Stem Cells Rescue Rotenone-Induced Experimental Leber’s Hereditary Optic Neuropathy-like Models In Vitro and In Vivo. Cells. 2023; 12(22):2617. https://doi.org/10.3390/cells12222617
Chicago/Turabian StyleTsai, En-Tung, Shih-Yuan Peng, You-Ren Wu, Tai-Chi Lin, Chih-Ying Chen, Yu-Hao Liu, Yu-Hsin Tseng, Yu-Jer Hsiao, Huan-Chin Tseng, Wei-Yi Lai, and et al. 2023. "HLA-Homozygous iPSC-Derived Mesenchymal Stem Cells Rescue Rotenone-Induced Experimental Leber’s Hereditary Optic Neuropathy-like Models In Vitro and In Vivo" Cells 12, no. 22: 2617. https://doi.org/10.3390/cells12222617
APA StyleTsai, E.-T., Peng, S.-Y., Wu, Y.-R., Lin, T.-C., Chen, C.-Y., Liu, Y.-H., Tseng, Y.-H., Hsiao, Y.-J., Tseng, H.-C., Lai, W.-Y., Lin, Y.-Y., Yang, Y.-P., Chiou, S.-H., Chen, S.-P., & Chien, Y. (2023). HLA-Homozygous iPSC-Derived Mesenchymal Stem Cells Rescue Rotenone-Induced Experimental Leber’s Hereditary Optic Neuropathy-like Models In Vitro and In Vivo. Cells, 12(22), 2617. https://doi.org/10.3390/cells12222617