

The Role of Sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor Signaling in Psoriasis



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Sphingolipid Metabolism
3. Ceramide Roles in the Skin
4. S1P and S1P–S1PR Signaling in the Skin
5. Psoriasis
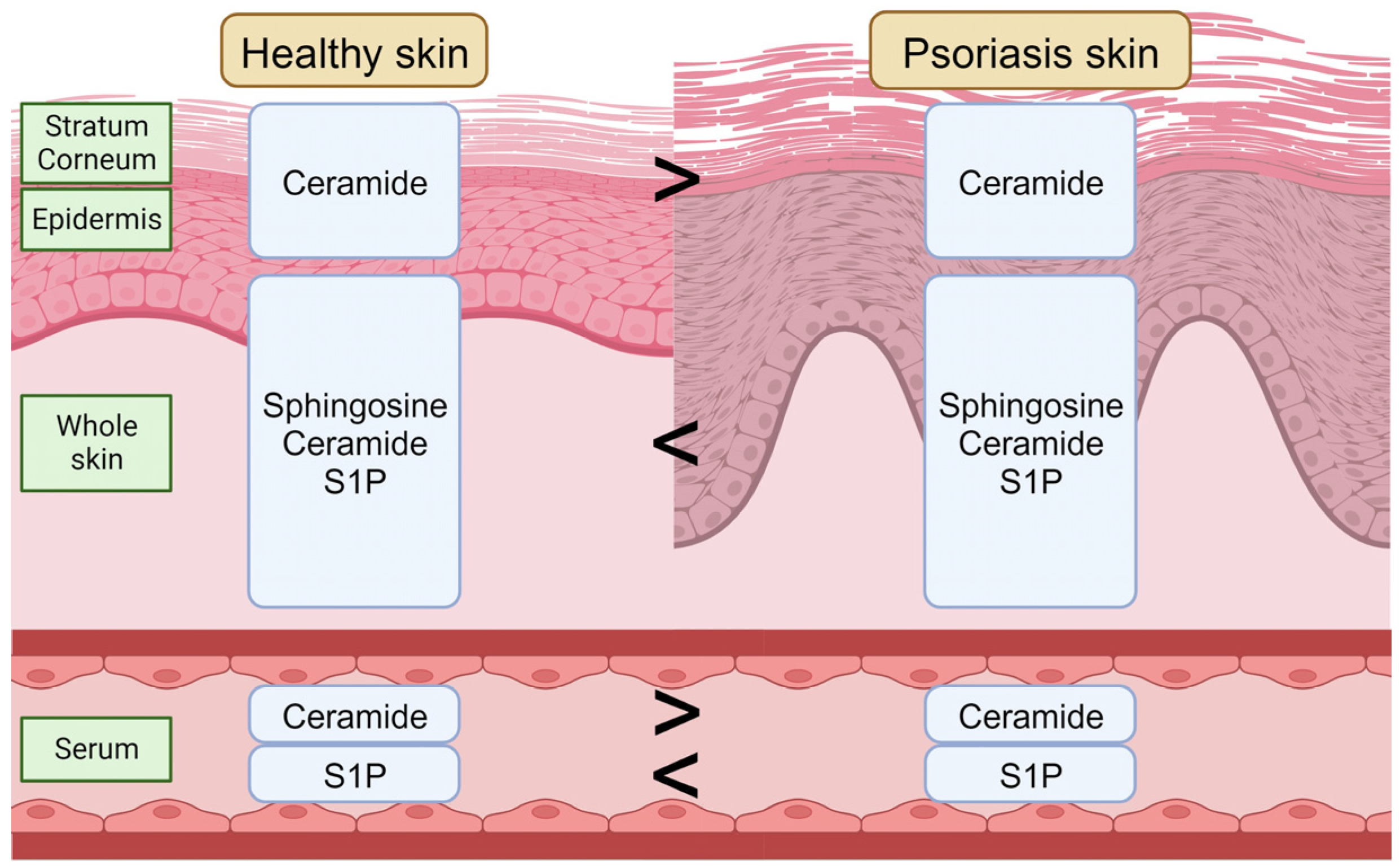
6. Sphingolipids in Psoriasis
7. S1P–S1PRs in Psoriasis
8. S1PR Modulators for Psoriasis
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, regional, and worldwide epidemiology of psoriasis: Systematic analysis and modelling study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Elmets, C.A.; Korman, N.J.; Prater, E.F.; Wong, E.B.; Rupani, R.N.; Kivelevitch, D.; Armstrong, A.W.; Connor, C.; Cordoro, K.M.; Davis, D.M.R.; et al. Joint AAD-NPF Guidelines of care for the management and treatment of psoriasis with topical therapy and alternative medicine modalities for psoriasis severity measures. J. Am. Acad. Dermatol. 2021, 84, 432–470. [Google Scholar] [CrossRef]
- Mahesh, P.; Haque, M.A.; Salman, B.I.; Belal, T.S.; Ibrahim, A.E.; El Deeb, S. Fast and Sensitive Bioanalytical Method for the Determination of Deucravacitinib in Human Plasma Using HPLC-MS/MS: Application and Greenness Evaluation. Molecules 2023, 28, 5471. [Google Scholar] [CrossRef] [PubMed]
- de Chaves, E.P.; Bussiere, M.; MacInnis, B.; Vance, D.E.; Campenot, R.B.; Vance, J.E. Ceramide inhibits axonal growth and nerve growth factor uptake without compromising the viability of sympathetic neurons. J. Biol. Chem. 2001, 276, 36207–36214. [Google Scholar] [CrossRef]
- Jin, Z.X.; Huang, C.R.; Dong, L.; Goda, S.; Kawanami, T.; Sawaki, T.; Sakai, T.; Tong, X.P.; Masaki, Y.; Fukushima, T.; et al. Impaired TCR signaling through dysfunction of lipid rafts in sphingomyelin synthase 1 (SMS1)-knockdown T cells. Int. Immunol. 2008, 20, 1427–1437. [Google Scholar] [CrossRef]
- Jin, Y.; Knudsen, E.; Wang, L.; Bryceson, Y.; Damaj, B.; Gessani, S.; Maghazachi, A.A. Sphingosine 1-phosphate is a novel inhibitor of T-cell proliferation. Blood 2003, 101, 4909–4915. [Google Scholar] [CrossRef]
- Brakch, N.; Dormond, O.; Bekri, S.; Golshayan, D.; Correvon, M.; Mazzolai, L.; Steinmann, B.; Barbey, F. Evidence for a role of sphingosine-1 phosphate in cardiovascular remodelling in Fabry disease. Eur. Heart J. 2010, 31, 67–76. [Google Scholar] [CrossRef]
- Czubowicz, K.; Jęśko, H.; Wencel, P.; Lukiw, W.J.; Strosznajder, R.P. The Role of Ceramide and Sphingosine-1-Phosphate in Alzheimer’s Disease and Other Neurodegenerative Disorders. Mol. Neurobiol. 2019, 56, 5436–5455. [Google Scholar] [CrossRef]
- Tringali, C.; Giussani, P. Ceramide and Sphingosine-1-Phosphate in Neurodegenerative Disorders and Their Potential Involvement in Therapy. Int. J. Mol. Sci. 2022, 23, 7806. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, I.; van Echten-Deckert, G. Sphingosine 1-phosphate—A double edged sword in the brain. Biochim. Biophys. Acta Biomembr. 2017, 1859 Pt B, 1573–1582. [Google Scholar] [CrossRef]
- Pyszko, J.; Strosznajder, J.B. Sphingosine kinase 1 and sphingosine-1-phosphate in oxidative stress evoked by 1-methyl-4-phenylpyridinium (MPP+) in human dopaminergic neuronal cells. Mol. Neurobiol. 2014, 50, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Takasugi, N.; Sasaki, T.; Suzuki, K.; Osawa, S.; Isshiki, H.; Hori, Y.; Shimada, N.; Higo, T.; Yokoshima, S.; Fukuyama, T.; et al. BACE1 activity is modulated by cell-associated sphingosine-1-phosphate. J. Neurosci. 2011, 31, 6850–6857. [Google Scholar] [CrossRef] [PubMed]
- Mehling, M.; Brinkmann, V.; Antel, J.; Bar-Or, A.; Goebels, N.; Vedrine, C.; Kristofic, C.; Kuhle, J.; Lindberg, R.L.; Kappos, L. FTY720 therapy exerts differential effects on T cell subsets in multiple sclerosis. Neurology 2008, 71, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S. Sphingosine 1-phosphate: A ligand for the EDG-1 family of G-protein-coupled receptors. Ann. N. Y. Acad. Sci. 2000, 905, 54–60. [Google Scholar] [CrossRef]
- Zhang, H.; Desai, N.N.; Olivera, A.; Seki, T.; Brooker, G.; Spiegel, S. Sphingosine-1-phosphate, a novel lipid, involved in cellular proliferation. J. Cell Biol. 1991, 114, 155–167. [Google Scholar] [CrossRef]
- Tabasinezhad, M.; Samadi, N.; Ghanbari, P.; Mohseni, M.; Saei, A.A.; Sharifi, S.; Saeedi, N.; Pourhassan, A. Sphingosin 1-phosphate contributes in tumor progression. J. Cancer Res. Ther. 2013, 9, 556–563. [Google Scholar]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef]
- Nădăban, A.; Rousel, J.; El Yachioui, D.; Gooris, G.S.; Beddoes, C.M.; Dalgliesh, R.M.; Malfois, M.; Rissmann, R.; Bouwstra, J.A. Effect of sphingosine and phytosphingosine ceramide ratio on lipid arrangement and barrier function in skin lipid models. J. Lipid Res. 2023, 64, 100400. [Google Scholar] [CrossRef]
- Masuda-Kuroki, K.; Di Nardo, A. Sphingosine 1-Phosphate Signaling at the Skin Barrier Interface. Biology 2022, 11, 809. [Google Scholar] [CrossRef] [PubMed]
- Berdyshev, E.; Goleva, E.; Bronova, I.; Bronoff, A.S.; Streib, J.E.; Vang, K.A.; Richers, B.N.; Taylor, P.; Beck, L.; Villarreal, M.; et al. Signaling sphingolipids are biomarkers for atopic dermatitis prone to disseminated viral infections. J. Allergy Clin. Immunol. 2022, 150, 640–648. [Google Scholar] [CrossRef]
- Masuda-Kuroki, K.; Alimohammadi, S.; Di Nardo, A.S. Epidermidis Rescues Allergic Contact Dermatitis in Sphingosine 1-Phosphate Receptor 2-Deficient Skin. Int. J. Mol. Sci. 2023, 24, 13190. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Y.; Li, J.; Zhang, N.; Jin, Q.; Qi, Y.; Song, P. Pathogenic sphingosine 1-phosphate pathway in psoriasis: A critical review of its pathogenic significance and potential as a therapeutic target. Lipids Health Dis. 2023, 22, 52. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.K.; Bian, J.; Gill, D.L. Intracellular calcium release mediated by sphingosine derivatives generated in cells. Science 1990, 248, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Dressler, K.A.; Kolesnick, R.N. Ceramide 1-phosphate, a novel phospholipid in human leukemia (HL-60) cells. Synthesis via ceramide from sphingomyelin. J. Biol. Chem. 1990, 265, 14917–14921. [Google Scholar] [CrossRef]
- Mitsutake, S.; Kita, K.; Okino, N.; Ito, M. [14C]ceramide synthesis by sphingolipid ceramide N-deacylase: New assay for ceramidase activity detection. Anal. Biochem. 1997, 247, 52–57. [Google Scholar] [CrossRef]
- Kanfer, J.N.; Young, O.M.; Shapiro, D.; Brady, R.O. The metabolism of sphingomyelin. I. Purification and properties of a sphingomyelin-cleaving enzyme from rat liver tissue. J. Biol. Chem. 1966, 241, 1081–1084. [Google Scholar] [CrossRef]
- Becker, K.P.; Kitatani, K.; Idkowiak-Baldys, J.; Bielawski, J.; Hannun, Y.A. Selective inhibition of juxtanuclear translocation of protein kinase C betaII by a negative feedback mechanism involving ceramide formed from the salvage pathway. J. Biol. Chem. 2005, 280, 2606–2612. [Google Scholar] [CrossRef]
- Merrill, A.H., Jr.; Jones, D.D. An update of the enzymology and regulation of sphingomyelin metabolism. Biochim. Biophys. Acta 1990, 1044, 1–12. [Google Scholar] [CrossRef]
- Wollny, T.; Wątek, M.; Durnaś, B.; Niemirowicz, K.; Piktel, E.; Żendzian-Piotrowska, M.; Góźdź, S.; Bucki, R. Sphingosine-1-Phosphate Metabolism and Its Role in the Development of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2017, 18, 741. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.; van Echten-Deckert, G. Conversion of dihydroceramide to ceramide occurs at the cytosolic face of the endoplasmic reticulum. FEBS Lett. 1997, 416, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Hampton, R.Y.; Morand, O.H. Sphingomyelin synthase and PKC activation. Science 1989, 246, 1050. [Google Scholar] [CrossRef] [PubMed]
- Postma, F.R.; Jalink, K.; Hengeveld, T.; Moolenaar, W.H. Sphingosine-1-phosphate rapidly induces Rho-dependent neurite retraction: Action through a specific cell surface receptor. Embo J. 1996, 15, 2388–2392. [Google Scholar] [CrossRef]
- Hisano, Y.; Kobayashi, N.; Yamaguchi, A.; Nishi, T. Mouse SPNS2 functions as a sphingosine-1-phosphate transporter in vascular endothelial cells. PLoS ONE 2012, 7, e38941. [Google Scholar] [CrossRef]
- Delgado, A.; Fabriàs, G.; Casas, J.; Abad, J.L. Natural products as platforms for the design of sphingolipid-related anticancer agents. Adv. Cancer Res. 2013, 117, 237–281. [Google Scholar]
- Matsuura, K.; Canfield, K.; Feng, W.; Kurokawa, M. Metabolic Regulation of Apoptosis in Cancer. Int. Rev. Cell Mol. Biol. 2016, 327, 43–87. [Google Scholar]
- Jin, L.; Zhu, J.; Yao, L.; Shen, G.; Xue, B.X.; Tao, W. Targeting SphK1/2 by SKI-178 inhibits prostate cancer cell growth. Cell Death Dis. 2023, 14, 537. [Google Scholar] [CrossRef]
- Ghosh, S.; Juin, S.K.; Bhattacharyya Majumdar, S.; Majumdar, S. Crucial role of glucosylceramide synthase in the regulation of stem cell-like cancer cells in B16F10 murine melanoma. Mol. Carcinog. 2021, 60, 840–858. [Google Scholar] [CrossRef]
- Hawkins, C.C.; Jones, A.B.; Gordon, E.R.; Williford, S.E.; Harsh, Y.; Ziebro, J.K.; Landis, C.J.; Gc, S.; Crossman, D.K.; Cooper, S.J.; et al. Targeting Acid Ceramidase Inhibits Glioblastoma Cell Migration through Decreased AKT Signaling. Cells 2022, 11, 1873. [Google Scholar] [CrossRef]
- Kendall, A.C.; Pilkington, S.M.; Wray, J.R.; Newton, V.L.; Griffiths, C.E.M.; Bell, M.; Watson, R.E.B.; Nicolaou, A. Menopause induces changes to the stratum corneum ceramide profile, which are prevented by hormone replacement therapy. Sci. Rep. 2022, 12, 21715. [Google Scholar] [CrossRef] [PubMed]
- Rabionet, M.; Gorgas, K.; Sandhoff, R. Ceramide synthesis in the epidermis. Biochim. Biophys. Acta 2014, 1841, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Kawana, M.; Miyamoto, M.; Ohno, Y.; Kihara, A. Comparative profiling and comprehensive quantification of stratum corneum ceramides in humans and mice by LC/MS/MS. J. Lipid Res. 2020, 61, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Hankemeier, T.; Kezic, S.; Wolterbeek, R.; et al. Increase in short-chain ceramides correlates with an altered lipid organization and decreased barrier function in atopic eczema patients. J. Lipid Res. 2012, 53, 2755–2766. [Google Scholar] [CrossRef]
- Mojumdar, E.H.; Kariman, Z.; van Kerckhove, L.; Gooris, G.S.; Bouwstra, J.A. The role of ceramide chain length distribution on the barrier properties of the skin lipid membranes. Biochim. Biophys. Acta 2014, 1838, 2473–2483. [Google Scholar] [CrossRef]
- Uche, L.E.; Gooris, G.S.; Bouwstra, J.A.; Beddoes, C.M. Increased Levels of Short-Chain Ceramides Modify the Lipid Organization and Reduce the Lipid Barrier of Skin Model Membranes. Langmuir 2021, 37, 9478–9489. [Google Scholar] [CrossRef]
- Draelos, Z.D.; Baalbaki, N.; Colon, G.; Dreno, B. Ceramide-Containing Adjunctive Skin Care for Skin Barrier Restoration During Acne Vulgaris Treatment. J. Drugs Dermatol. 2023, 22, 554–558. [Google Scholar] [CrossRef]
- Bzioueche, H.; Tamelghaghet, M.; Chignon-Sicard, B.; Bazile, N.; Hauchecorne, P.; Barbero Calderón, M.; Meunier, P.; Rocchi, S.; Passeron, T.; Tulic, M.K. Ceramide AD™ Restores Skin Integrity and Function following Exposure to House Dust Mite. Int. J. Mol. Sci. 2023, 24, 9234. [Google Scholar] [CrossRef]
- Nugroho, W.T.; Sawitri, S.; Astindari, A.; Utomo, B.; Listiawan, M.Y.; Ervianti, E.; Astari, L. The Efficacy of Moisturisers Containing Ceramide Compared with Other Moisturisers in the Management of Atopic Dermatitis: A Systematic Literature Review and Meta-Analysis. Indian J. Dermatol. 2023, 68, 53–58. [Google Scholar]
- Schachner, L.; Alexis, A.; Andriessen, A.; Baldwin, H.; Cork, M.; Kirsner, R.; Woolery-Lloyd, H. Supplement Individual Article: The Importance of a Healthy Skin Barrier From the Cradle to the Grave Using Ceramide-Containing Cleansers and Moisturizers: A Review and Consensus. J. Drugs Dermatol. 2023, 22, SF344607s3–SF344607s14. [Google Scholar]
- Kirsner, R.; Andriessen, A. An International Evaluation of a Ceramide-Containing Hydrating Cleanser and Moisturizing Cream for the Improvement of Diabetes Mellitus-Related Xerosis. J. Drugs Dermatol. 2023, 22, 65–73. [Google Scholar] [CrossRef] [PubMed]
- van Smeden, J.; Al-Khakany, H.; Wang, Y.; Visscher, D.; Stephens, N.; Absalah, S.; Overkleeft, H.S.; Aerts, J.; Hovnanian, A.; Bouwstra, J.A. Skin barrier lipid enzyme activity in Netherton patients is associated with protease activity and ceramide abnormalities. J. Lipid Res. 2020, 61, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Fölster-Holst, R.; Naß, C.; Dähnhardt-Pfeiffer, S.; Freitag-Wolf, S. Analysis of the structure and function of the epidermal barrier in patients with ichthyoses-clinical and electron microscopical investigations. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Kircik, L.; Alexis, A.F.; Andriessen, A.; Blattner, C.; Glick, B.P.; Lynde, C.W.; Gold, L.S. Psoriasis and Skin Barrier Dysfunction: The Role of Gentle Cleansers and Moisturizers in Treating Psoriasis. J. Drugs Dermatol. 2023, 22, 773–778. [Google Scholar] [CrossRef]
- Kondo, T.; Kitano, T.; Iwai, K.; Watanabe, M.; Taguchi, Y.; Yabu, T.; Umehara, H.; Domae, N.; Uchiyama, T.; Okazaki, T. Control of ceramide-induced apoptosis by IGF-1: Involvement of PI-3 kinase, caspase-3 and catalase. Cell Death Differ. 2002, 9, 682–692. [Google Scholar] [CrossRef]
- Yabu, T.; Shiba, H.; Shibasaki, Y.; Nakanishi, T.; Imamura, S.; Touhata, K.; Yamashita, M. Stress-induced ceramide generation and apoptosis via the phosphorylation and activation of nSMase1 by JNK signaling. Cell Death Differ. 2015, 22, 258–273. [Google Scholar] [CrossRef]
- Ravid, T.; Tsaba, A.; Gee, P.; Rasooly, R.; Medina, E.A.; Goldkorn, T. Ceramide accumulation precedes caspase-3 activation during apoptosis of A549 human lung adenocarcinoma cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 284, L1082–L1092. [Google Scholar] [CrossRef]
- Zundel, W.; Giaccia, A. Inhibition of the anti-apoptotic PI(3)K/Akt/Bad pathway by stress. Genes Dev. 1998, 12, 1941–1946. [Google Scholar] [CrossRef]
- Bourbon, N.A.; Sandirasegarane, L.; Kester, M. Ceramide-induced inhibition of Akt is mediated through protein kinase Czeta: Implications for growth arrest. J. Biol. Chem. 2002, 277, 3286–3292. [Google Scholar] [CrossRef]
- Arboleda, G.; Morales, L.C.; Benítez, B.; Arboleda, H. Regulation of ceramide-induced neuronal death: Cell metabolism meets neurodegeneration. Brain Res. Rev. 2009, 59, 333–346. [Google Scholar] [CrossRef]
- Monick, M.M.; Mallampalli, R.K.; Carter, A.B.; Flaherty, D.M.; McCoy, D.; Robeff, P.K.; Peterson, M.W.; Hunninghake, G.W. Ceramide regulates lipopolysaccharide-induced phosphatidylinositol 3-kinase and Akt activity in human alveolar macrophages. J. Immunol. 2001, 167, 5977–5985. [Google Scholar] [CrossRef] [PubMed]
- Kihara, A.; Mitsutake, S.; Mizutani, Y.; Igarashi, Y. Metabolism and biological functions of two phosphorylated sphingolipids, sphingosine 1-phosphate and ceramide 1-phosphate. Prog. Lipid Res. 2007, 46, 126–144. [Google Scholar] [CrossRef]
- Spiegel, S.; Maczis, M.A.; Maceyka, M.; Milstien, S. New insights into functions of the sphingosine-1-phosphate transporter SPNS2. J. Lipid Res. 2019, 60, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Strub, G.M.; Maceyka, M.; Hait, N.C.; Milstien, S.; Spiegel, S. Extracellular and intracellular actions of sphingosine-1-phosphate. Adv. Exp. Med. Biol. 2010, 688, 141–155. [Google Scholar] [PubMed]
- Ye, L.; Song, J.; Zheng, Y.; Zhong, M.; Liu, J.; Zhu, D.; Hu, S. New mechanism for mesenchymal stem cell microvesicle to restore lung permeability: Intracellular S1P signaling pathway independent of S1P receptor-1. Stem Cell Res. Ther. 2022, 13, 496. [Google Scholar] [CrossRef]
- Usatyuk, P.V.; He, D.; Bindokas, V.; Gorshkova, I.A.; Berdyshev, E.V.; Garcia, J.G.; Natarajan, V. Photolysis of caged sphingosine-1-phosphate induces barrier enhancement and intracellular activation of lung endothelial cell signaling pathways. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 300, L840–L850. [Google Scholar] [CrossRef]
- Billich, A.; Urtz, N.; Reuschel, R.; Baumruker, T. Sphingosine kinase 1 is essential for proteinase-activated receptor-1 signalling in epithelial and endothelial cells. Int. J. Biochem. Cell Biol. 2009, 41, 1547–1555. [Google Scholar] [CrossRef]
- Xia, P.; Wang, L.; Moretti, P.A.; Albanese, N.; Chai, F.; Pitson, S.M.; D’Andrea, R.J.; Gamble, J.R.; Vadas, M.A. Sphingosine kinase interacts with TRAF2 and dissects tumor necrosis factor-alpha signaling. J. Biol. Chem. 2002, 277, 7996–8003. [Google Scholar] [CrossRef]
- Vann, L.R.; Payne, S.G.; Edsall, L.C.; Twitty, S.; Spiegel, S.; Milstien, S. Involvement of sphingosine kinase in TNF-alpha-stimulated tetrahydrobiopterin biosynthesis in C6 glioma cells. J. Biol. Chem. 2002, 277, 12649–12656. [Google Scholar] [CrossRef]
- Etemadi, N.; Chopin, M.; Anderton, H.; Tanzer, M.C.; Rickard, J.A.; Abeysekera, W.; Hall, C.; Spall, S.K.; Wang, B.; Xiong, Y.; et al. Correction: TRAF2 regulates TNF and NF-κB signalling to suppress apoptosis and skin inflammation independently of Sphingosine kinase 1. Elife 2017, 6, e10592. [Google Scholar] [CrossRef]
- Blaho, V.A.; Hla, T. An update on the biology of sphingosine 1-phosphate receptors. J. Lipid Res. 2014, 55, 1596–1608. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Hla, T.; Lynch, K.R.; Spiegel, S.; Moolenaar, W.H. International Union of Basic and Clinical Pharmacology. LXXVIII. Lysophospholipid receptor nomenclature. Pharmacol. Rev. 2010, 62, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Igawa, S.; Choi, J.E.; Wang, Z.; Chang, Y.L.; Wu, C.C.; Werbel, T.; Ishida-Yamamoto, A.; Di Nardo, A. Human Keratinocytes Use Sphingosine 1-Phosphate and its Receptors to Communicate Staphylococcus aureus Invasion and Activate Host Defense. J. Investig. Dermatol. 2019, 139, 1743–1752.e5. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, G.C.; Gilmour, J.; Giannoudaki, E.; Kirby, J.A.; Sheerin, N.S.; Ali, S. Dissecting the Therapeutic Mechanisms of Sphingosine-1-Phosphate Receptor Agonism during Ischaemia and Reperfusion. Int. J. Mol. Sci. 2023, 24, 11192. [Google Scholar] [CrossRef]
- Gray, N.; Limberg, M.M.; Wiebe, D.; Weihrauch, T.; Langner, A.; Brandt, N.; Bräuer, A.U.; Raap, U. Differential Upregulation and Functional Activity of S1PR1 in Human Peripheral Blood Basophils of Atopic Patients. Int. J. Mol. Sci. 2022, 23, 16117. [Google Scholar] [CrossRef]
- Jo, H.; Shim, K.; Jeoung, D. The Crosstalk between FcεRI and Sphingosine Signaling in Allergic Inflammation. Int. J. Mol. Sci. 2022, 23, 13892. [Google Scholar] [CrossRef]
- Karuppuchamy, T.; Behrens, E.H.; González-Cabrera, P.; Sarkisyan, G.; Gima, L.; Boyer, J.D.; Bamias, G.; Jedlicka, P.; Veny, M.; Clark, D.; et al. Sphingosine-1-phosphate receptor-1 (S1P(1)) is expressed by lymphocytes, dendritic cells, and endothelium and modulated during inflammatory bowel disease. Mucosal Immunol. 2017, 10, 162–171. [Google Scholar] [CrossRef]
- Bock, S.; Pfalzgraff, A.; Weindl, G. Sphingosine 1-phospate differentially modulates maturation and function of human Langerhans-like cells. J. Dermatol. Sci. 2016, 82, 9–17. [Google Scholar] [CrossRef]
- Landeen, L.K.; Aroonsakool, N.; Haga, J.H.; Hu, B.S.; Giles, W.R. Sphingosine-1-phosphate receptor expression in cardiac fibroblasts is modulated by in vitro culture conditions. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2698–H2711. [Google Scholar] [CrossRef][Green Version]
- Chen, T.; Song, P.; He, M.; Rui, S.; Duan, X.; Ma, Y.; Armstrong, D.G.; Deng, W. Sphingosine-1-phosphate derived from PRP-Exos promotes angiogenesis in diabetic wound healing via the S1PR1/AKT/FN1 signalling pathway. Burn. Trauma. 2023, 11, tkad003. [Google Scholar] [CrossRef]
- Igawa, S.; Ohzono, A.; Pham, P.; Wang, Z.; Nakatsuji, T.; Dokoshi, T.; Di Nardo, A. Sphingosine 1-Phosphate Receptor 2 Is Central to Maintaining Epidermal Barrier Homeostasis. J. Investig. Dermatol. 2021, 141, 1188–1197.e5. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Im, D.S. Blockage of sphingosine-1-phosphate receptor 2 attenuates 2,4-dinitrochlorobenzene-induced atopic dermatitis in mice. Acta Pharmacol. Sin. 2020, 41, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, K.; Liu, C.H.; Faraco, G.; Galvani, S.; Smith, H.K.; Burg, N.; Anrather, J.; Sanchez, T.; Iadecola, C.; Hla, T. Size-selective opening of the blood-brain barrier by targeting endothelial sphingosine 1-phosphate receptor 1. Proc. Natl. Acad. Sci. USA 2017, 114, 4531–4536. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Nakamura, K.; Kaneko, F.; Nakagawa, H.; Iizuka, H. Analysis of psoriasis patients registered with the Japanese Society for Psoriasis Research from 2002–2008. J. Dermatol. 2011, 38, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Takahashi, A.; Taniguchi, Y.; Terada, Y.; Sano, S. Detection of asymptomatic enthesitis in psoriasis patients: An onset of psoriatic arthritis? J. Dermatol. 2016, 43, 650–654. [Google Scholar] [CrossRef]
- Nestle, F.O.; Di Meglio, P.; Qin, J.Z.; Nickoloff, B.J. Skin immune sentinels in health and disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef]
- Fenton, C.; Plosker, G.L. Calcipotriol/betamethasone dipropionate: A review of its use in the treatment of psoriasis vulgaris. Am. J. Clin. Dermatol. 2004, 5, 463–478. [Google Scholar] [CrossRef]
- Bagel, J. Treat to Target in Psoriasis: A Real-World Experience With Biologics and Adjunctive Topical Therapy. J. Drugs Dermatol. 2018, 17, 918. [Google Scholar]
- Brownstone, N.D.; Hong, J.; Mosca, M.; Hadeler, E.; Liao, W.; Bhutani, T.; Koo, J. Biologic Treatments of Psoriasis: An Update for the Clinician. Biologics 2021, 15, 39–51. [Google Scholar] [CrossRef]
- Gonciarz, M.; Pawlak-Buś, K.; Leszczyński, P.; Owczarek, W. TYK2 as a therapeutic target in the treatment of autoimmune and inflammatory diseases. Immunotherapy 2021, 13, 1135–1150. [Google Scholar] [CrossRef]
- Crispino, N.; Ciccia, F. JAK/STAT pathway and nociceptive cytokine signalling in rheumatoid arthritis and psoriatic arthritis. Clin. Exp. Rheumatol. 2021, 39, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Słuczanowska-Głąbowska, S.; Ziegler-Krawczyk, A.; Szumilas, K.; Pawlik, A. Role of Janus Kinase Inhibitors in Therapy of Psoriasis. J. Clin. Med. 2021, 10, 4307. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chen, X.; Zhang, H.; Xie, T.; Ye, X.Y. Selective Tyk2 inhibitors as potential therapeutic agents: A patent review (2015–2018). Expert. Opin. Ther. Pat. 2019, 29, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Saeki, H.; Mabuchi, T.; Asahina, A.; Abe, M.; Igarashi, A.; Imafuku, S.; Okubo, Y.; Komine, M.; Takahashi, K.; Torii, H.; et al. English version of Japanese guidance for the use of oral Janus kinase inhibitors (JAK1 and TYK2 inhibitors) in the treatments of psoriasis. J. Dermatol. 2023, 50, e138–e150. [Google Scholar] [CrossRef]
- Ortiz-Lopez, L.I.; Choudhary, V.; Bollag, W.B. Updated Perspectives on Keratinocytes and Psoriasis: Keratinocytes are More Than Innocent Bystanders. Psoriasis 2022, 12, 73–87. [Google Scholar] [CrossRef]
- Nowowiejska, J.; Baran, A.; Flisiak, I. Aberrations in Lipid Expression and Metabolism in Psoriasis. Int. J. Mol. Sci. 2021, 22, 6561. [Google Scholar] [CrossRef]
- Matwiejuk, M.; Mysliwiec, H.; Lukaszuk, B.; Lewoc, M.; Malla, H.; Mysliwiec, P.; Dadan, J.; Chabowski, A.; Flisiak, I. The Interplay between Bioactive Sphingolipids in the Psoriatic Skin and the Severity of the Disease. Int. J. Mol. Sci. 2023, 24, 11336. [Google Scholar] [CrossRef]
- Moon, S.H.; Kim, J.Y.; Song, E.H.; Shin, M.K.; Cho, Y.H.; Kim, N.I. Altered levels of sphingosine and sphinganine in psoriatic epidermis. Ann. Dermatol. 2013, 25, 321–326. [Google Scholar] [CrossRef]
- Liu, P.; Hou, G.; Kuang, Y.; Li, L.; Chen, C.; Yan, B.; Zhu, W.; Li, J.; Chen, M.; Su, J.; et al. Lipidomic profiling reveals metabolic signatures in psoriatic skin lesions. Clin. Immunol. 2023, 246, 109212. [Google Scholar] [CrossRef]
- Moskot, M.; Bocheńska, K.; Jakóbkiewicz-Banecka, J.; Banecki, B.; Gabig-Cimińska, M. Abnormal Sphingolipid World in Inflammation Specific for Lysosomal Storage Diseases and Skin Disorders. Int. J. Mol. Sci. 2018, 19, 247. [Google Scholar] [CrossRef]
- Nakajima, K.; Terao, M.; Takaishi, M.; Kataoka, S.; Goto-Inoue, N.; Setou, M.; Horie, K.; Sakamoto, F.; Ito, M.; Azukizawa, H.; et al. Barrier abnormality due to ceramide deficiency leads to psoriasiform inflammation in a mouse model. J. Investig. Dermatol. 2013, 133, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Uchino, T.; Kamiya, D.; Yagi, H.; Fujino-Shimaya, H.; Hatta, I.; Fujimori, S.; Miyazaki, Y.; Kirishita, Y.; Sano, Y.; Mizuno, H.; et al. Comparative analysis of intercellular lipid organization and composition between psoriatic and healthy stratum corneum. Chem. Phys. Lipids 2023, 254, 105305. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Shon, J.C.; Seo, H.S.; Liu, K.H.; Lee, J.W.; Ahn, S.K.; Hong, S.P. Decrease of ceramides with long-chain fatty acids in psoriasis: Possible inhibitory effect of interferon gamma on chain elongation. Exp. Dermatol. 2022, 31, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, W.; Wroński, A.; Domingues, P.; Domingues, M.R.; Skrzydlewska, E. Lipidomic Analysis Reveals Specific Differences between Fibroblast and Keratinocyte Ceramide Profile of Patients with Psoriasis Vulgaris. Molecules 2020, 25, 630. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Monti, M.; Sesana, S.; Caputo, R.; Carelli, S.; Ghidoni, R. Ceramide composition of the psoriatic scale. Biochim. Biophys. Acta 1993, 1182, 147–151. [Google Scholar] [CrossRef]
- Tawada, C.; Kanoh, H.; Nakamura, M.; Mizutani, Y.; Fujisawa, T.; Banno, Y.; Seishima, M. Interferon-γ decreases ceramides with long-chain fatty acids: Possible involvement in atopic dermatitis and psoriasis. J. Investig. Dermatol. 2014, 134, 712–718. [Google Scholar] [CrossRef]
- Myśliwiec, H.; Baran, A.; Harasim-Symbor, E.; Choromańska, B.; Myśliwiec, P.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Increase in circulating sphingosine-1-phosphate and decrease in ceramide levels in psoriatic patients. Arch. Dermatol. Res. 2017, 309, 79–86. [Google Scholar] [CrossRef]
- Checa, A.; Xu, N.; Sar, D.G.; Haeggström, J.Z.; Ståhle, M.; Wheelock, C.E. Circulating levels of sphingosine-1-phosphate are elevated in severe, but not mild psoriasis and are unresponsive to anti-TNF-α treatment. Sci. Rep. 2015, 5, 12017. [Google Scholar] [CrossRef]
- Nada, H.A.; Elshabrawy, M.M.; Ismail, N.I.; Hassan, E.T.; Jafferany, M.; Elsaie, M.L. Therapeutic implications and role of serum sphingolipids on psoriasis severity after narrow band ultraviolet B treatment: A cross sectional controlled study. Dermatol. Ther. 2020, 33, e13988. [Google Scholar] [CrossRef]
- Kozłowska, D.; Harasim-Symbor, E.; Myśliwiec, H.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Serum sphingolipid level in psoriatic patients with obesity. Postep. Dermatol. Alergol. 2019, 36, 714–721. [Google Scholar] [CrossRef]
- Schaper, K.; Dickhaut, J.; Japtok, L.; Kietzmann, M.; Mischke, R.; Kleuser, B.; Baumer, W. Sphingosine-1-phosphate exhibits anti-proliferative and anti-inflammatory effects in mouse models of psoriasis. J. Dermatol. Sci. 2013, 71, 29–36. [Google Scholar] [CrossRef]
- Chen, F.; Ni, C.; Wang, X.; Cheng, R.; Pan, C.; Wang, Y.; Liang, J.; Zhang, J.; Cheng, J.; Chin, Y.E.; et al. S1P defects cause a new entity of cataract, alopecia, oral mucosal disorder, and psoriasis-like syndrome. EMBO Mol. Med. 2022, 14, e14904. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, G.; Zhang, H.; Li, L. Xiaoyin Jiedu Granules may alleviate psoriasis-like skin diseases in mice by regulating sphingosine 1-phosphate receptor expression and reducing Th17 cells. Heliyon 2023, 9, e19109. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.N.; Raue, R.; Weigert, A.; von Knethen, A.; Brüne, B. Macrophage S1PR1 Signaling Alters Angiogenesis and Lymphangiogenesis During Skin Inflammation. Cells 2019, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Xue, N.; Lai, F.; Zhang, X.; Zhang, S.; Wang, Y.; Jin, J.; Chen, X. Validating a Selective S1P(1) Receptor Modulator Syl930 for Psoriasis Treatment. Biol. Pharm. Bull. 2018, 41, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Vaclavkova, A.; Chimenti, S.; Arenberger, P.; Holló, P.; Sator, P.G.; Burcklen, M.; Stefani, M.; D’Ambrosio, D. Oral ponesimod in patients with chronic plaque psoriasis: A randomised, double-blind, placebo-controlled phase 2 trial. Lancet 2014, 384, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Megna, M.; Noto, M.; Fabbrocini, G.; Fornaro, L. A case of psoriasis and multiple sclerosis succesfully treated with concomitant fingolimod and secukinumab. Australas. J. Dermatol. 2023, 64, e100–e102. [Google Scholar] [CrossRef] [PubMed]
- Ikumi, N.; Fujita, H. Generalized pustular psoriasis occurring in a patient with multiple sclerosis during treatment with fingolimod. J. Dermatol. 2023, 50, 16802. [Google Scholar] [CrossRef]
- Chen, Y.J.; Kyles, A.E.; Gregory, C.R. In vitro evaluation of the effect of a novel immunosuppressive agent, FTY720, on the function of feline neutrophils. Am. J. Vet. Res. 2006, 67, 588–592. [Google Scholar] [CrossRef]
- Bell, M.; Foley, D.; Naylor, C.; Robinson, C.; Riley, J.; Epemolu, O.; Scullion, P.; Shishikura, Y.; Katz, E.; McLean, W.H.I.; et al. Discovery of super soft-drug modulators of sphingosine-1-phosphate receptor 1. Bioorg. Med. Chem. Lett. 2018, 28, 3255–3259. [Google Scholar] [CrossRef]
- Jena, P.K.; Sheng, L.; McNeil, K.; Chau, T.Q.; Yu, S.; Kiuru, M.; Fung, M.A.; Hwang, S.T.; Wan, Y.Y. Long-term Western diet intake leads to dysregulated bile acid signaling and dermatitis with Th2 and Th17 pathway features in mice. J. Dermatol. Sci. 2019, 95, 13–20. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yin, X.; Yan, J.; Li, X.; Sun, Q. The lncRNA H19/miR-766-3p/S1PR3 Axis Contributes to the Hyperproliferation of Keratinocytes and Skin Inflammation in Psoriasis via the AKT/mTOR Pathway. Mediat. Inflamm. 2021, 2021, 9991175. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.; Golfier, S.; Tabeling, C.; Räbel, K.; Gräler, M.H.; Witzenrath, M.; Lipp, M. Sphingosine-1-phospate receptor 4 (S1P₄) deficiency profoundly affects dendritic cell function and TH17-cell differentiation in a murine model. Faseb. J. 2011, 25, 4024–4036. [Google Scholar] [CrossRef] [PubMed]
- Dillmann, C.; Ringel, C.; Ringleb, J.; Mora, J.; Olesch, C.; Fink, A.F.; Roberts, E.; Brüne, B.; Weigert, A. S1PR4 Signaling Attenuates ILT 7 Internalization To Limit IFN-α Production by Human Plasmacytoid Dendritic Cells. J. Immunol. 2016, 196, 1579–1590. [Google Scholar] [CrossRef]
- Schuster, C.; Huard, A.; Sirait-Fischer, E.; Dillmann, C.; Brüne, B.; Weigert, A. S1PR4-dependent CCL2 production promotes macrophage recruitment in a murine psoriasis model. Eur. J. Immunol. 2020, 50, 839–845. [Google Scholar] [CrossRef]
- Okura, I.; Kamata, M.; Asano, Y.; Mitsui, A.; Shimizu, T.; Sato, S.; Tada, Y. Fingolimod ameliorates imiquimod-induced psoriasiform dermatitis by sequestrating interleukin-17-producing gd T cells in secondary lymph nodes. J. Dermatol. Sci. 2021, 102, 116–125. [Google Scholar] [CrossRef]
- Pérez-Jeldres, T.; Alvarez-Lobos, M.; Rivera-Nieves, J. Targeting Sphingosine-1-Phosphate Signaling in Immune-Mediated Diseases: Beyond Multiple Sclerosis. Drugs 2021, 81, 985–1002. [Google Scholar] [CrossRef]
- Kappos, L.; Radue, E.W.; O’Connor, P.; Polman, C.; Hohlfeld, R.; Calabresi, P.; Selmaj, K.; Agoropoulou, C.; Leyk, M.; Zhang-Auberson, L.; et al. A placebo-controlled trial of oral fingolimod in relapsing multiple sclerosis. N. Engl. J. Med. 2010, 362, 387–401. [Google Scholar] [CrossRef]
- Choden, T.; Cohen, N.A.; Rubin, D.T. Sphingosine-1 Phosphate Receptor Modulators: The Next Wave of Oral Therapies in Inflammatory Bowel Disease. Gastroenterol. Hepatol. 2022, 18, 265–271. [Google Scholar]
- Dertschnig, S.; Passweg, J.; Bucher, C.; Medinger, M.; Tzankov, A. Mocravimod, a S1P receptor modulator, increases T cell counts in bone marrow biopsies from patients undergoing allogeneic hematopoietic stem cell transplantation. Cell Immunol. 2023, 388–389, 104719. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Bissonnette, R.; Kircik, L.; Murrell, D.F.; Selfridge, A.; Liu, K.; Ahluwalia, G.; Guttman-Yassky, E. Efficacy and safety of etrasimod, a sphingosine 1-phosphate receptor modulator, in adults with moderate-to-severe atopic dermatitis (ADVISE). J. Eur. Acad. Dermatol. Venereol. 2023, 37, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.; D’Ambrosio, D.; Dingemanse, J. Modeling clinical efficacy of the S1P receptor modulator ponesimod in psoriasis. J. Dermatol. Sci. 2018, 89, 136–145. [Google Scholar] [CrossRef]
- Paik, J. Ozanimod: A Review in Ulcerative Colitis. Drugs 2022, 82, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, J.; Li, H.J.; Zhang, S.; Jin, M.Z.; Chen, S.T.; Sun, X.Y.; Zhou, Y.Q.; Lu, Y.; Yang, D.; et al. Sphingosine-1-Phosphate and Its Signal Modulators Alleviate Psoriasis-Like Dermatitis: Preclinical and Clinical Evidence and Possible Mechanisms. Front. Immunol. 2021, 12, 759276. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda-Kuroki, K.; Alimohammadi, S.; Di Nardo, A. The Role of Sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor Signaling in Psoriasis. Cells 2023, 12, 2352. https://doi.org/10.3390/cells12192352
Masuda-Kuroki K, Alimohammadi S, Di Nardo A. The Role of Sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor Signaling in Psoriasis. Cells. 2023; 12(19):2352. https://doi.org/10.3390/cells12192352
Chicago/Turabian StyleMasuda-Kuroki, Kana, Shahrzad Alimohammadi, and Anna Di Nardo. 2023. "The Role of Sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor Signaling in Psoriasis" Cells 12, no. 19: 2352. https://doi.org/10.3390/cells12192352
APA StyleMasuda-Kuroki, K., Alimohammadi, S., & Di Nardo, A. (2023). The Role of Sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor Signaling in Psoriasis. Cells, 12(19), 2352. https://doi.org/10.3390/cells12192352