

Exogenous Melatonin Enhances the Low Phosphorus Tolerance of Barley Roots of Different Genotypes

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Root Morphology and Phosphorus Concentration
2.3. Quantification of Melatonin by LC-MS
2.4. Determination of ROS Accumulation and Antioxidant Enzyme Activities
2.5. Metabolite Extraction and LC-MS/MS Analysis
2.6. Qualitative and Quantitative Analyses of Metabolites
2.7. Statistical Analysis
3. Results
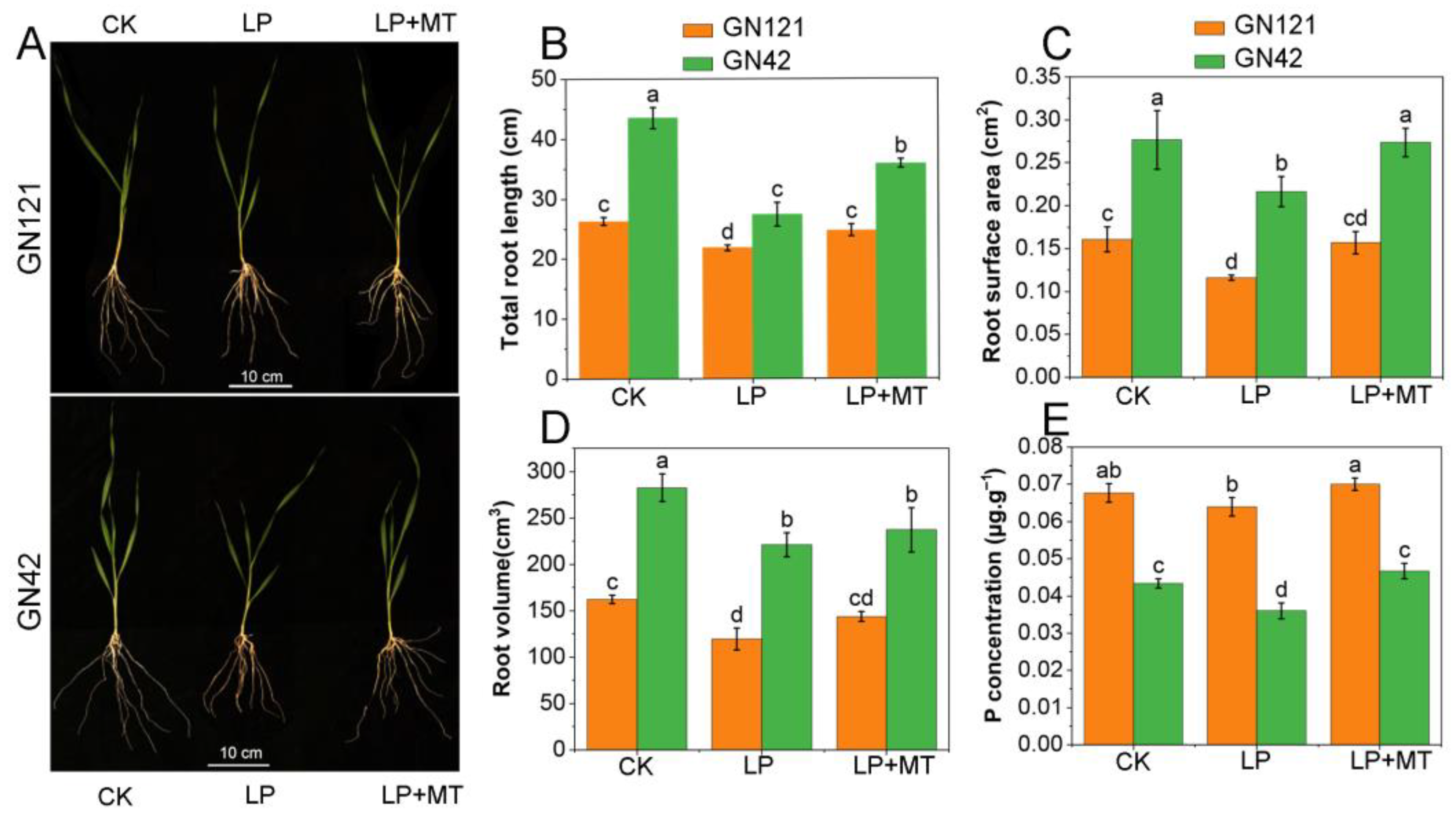
3.1. Exogenous Melatonin Improved the Primary Root Growth under LP Stress
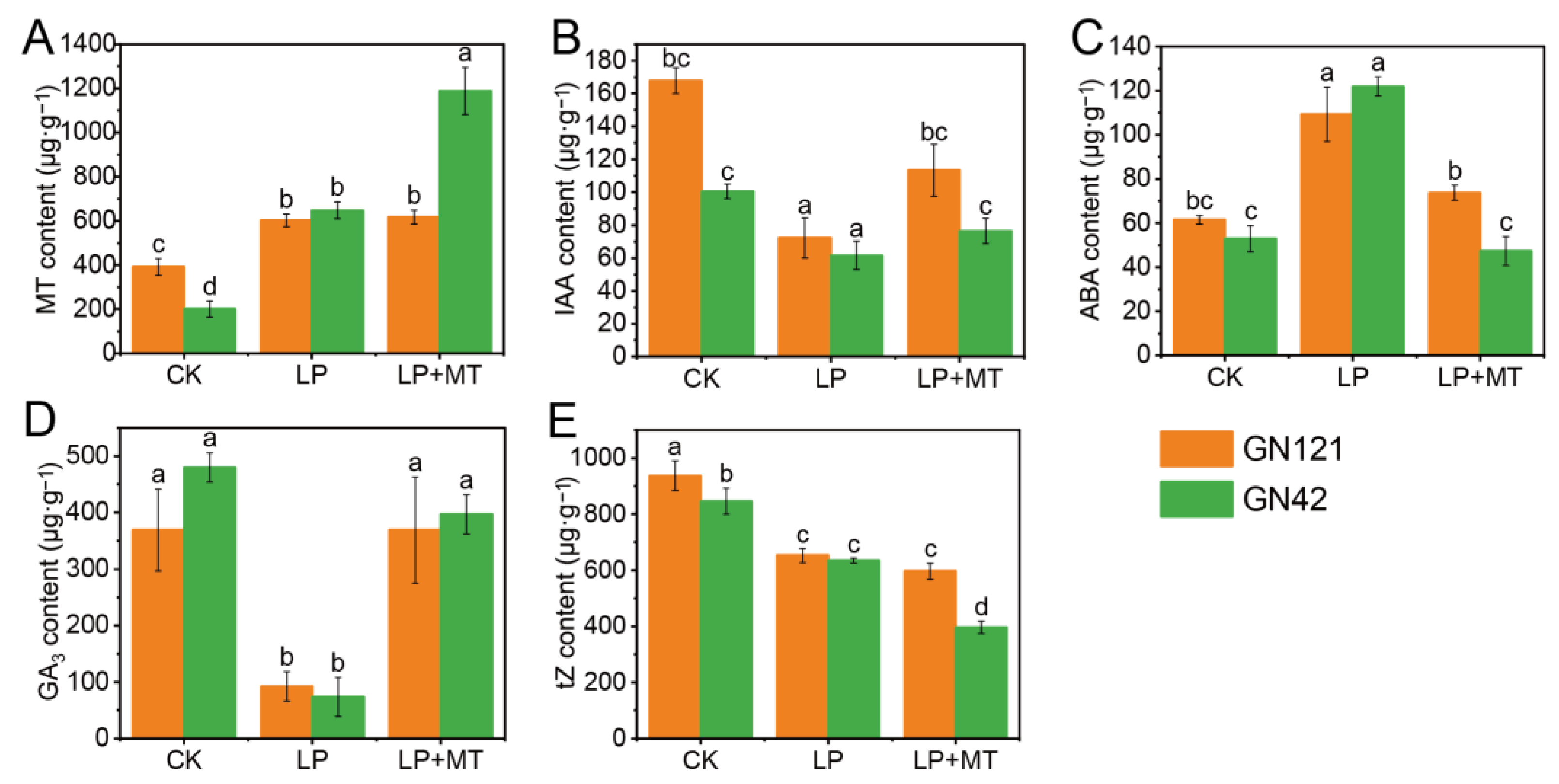
3.2. Exogenous Melatonin Regulates Changes in Endogenous Hormone Levels
3.3. Exogenous Melatonin Alleviated ROS Accumulation against LP-Induced Oxidative Stress
3.4. Data Quality Analysis
3.5. Qualitative and Quantitative Analyses of Metabolites
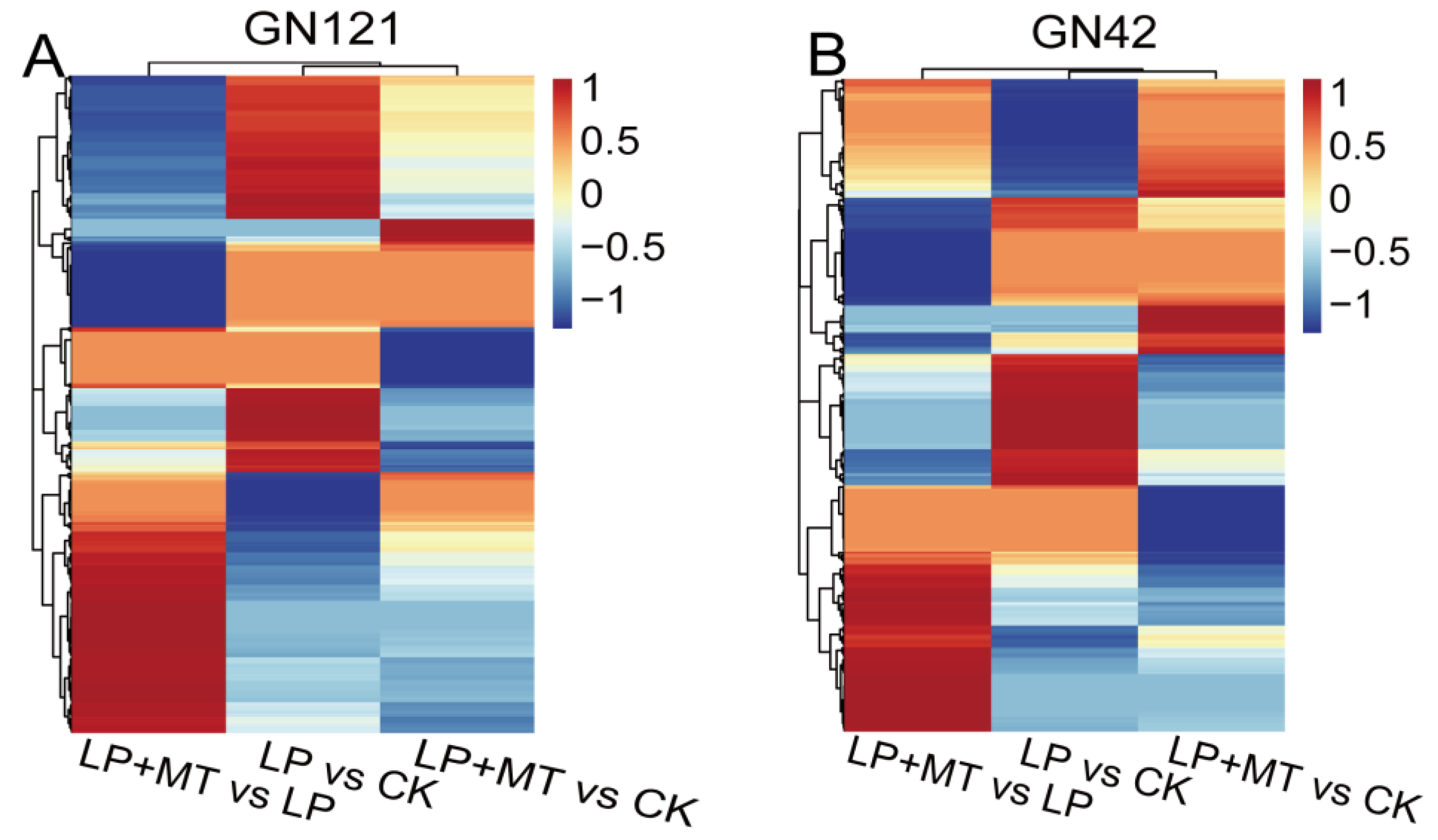
3.6. Global Metabolic Changes under LP Stress and Exogenous Melatonin Treatments
3.7. Changes in P-Containinglobal Metabolic Changes under LP Stress and Exogenous Melatonin Treatments
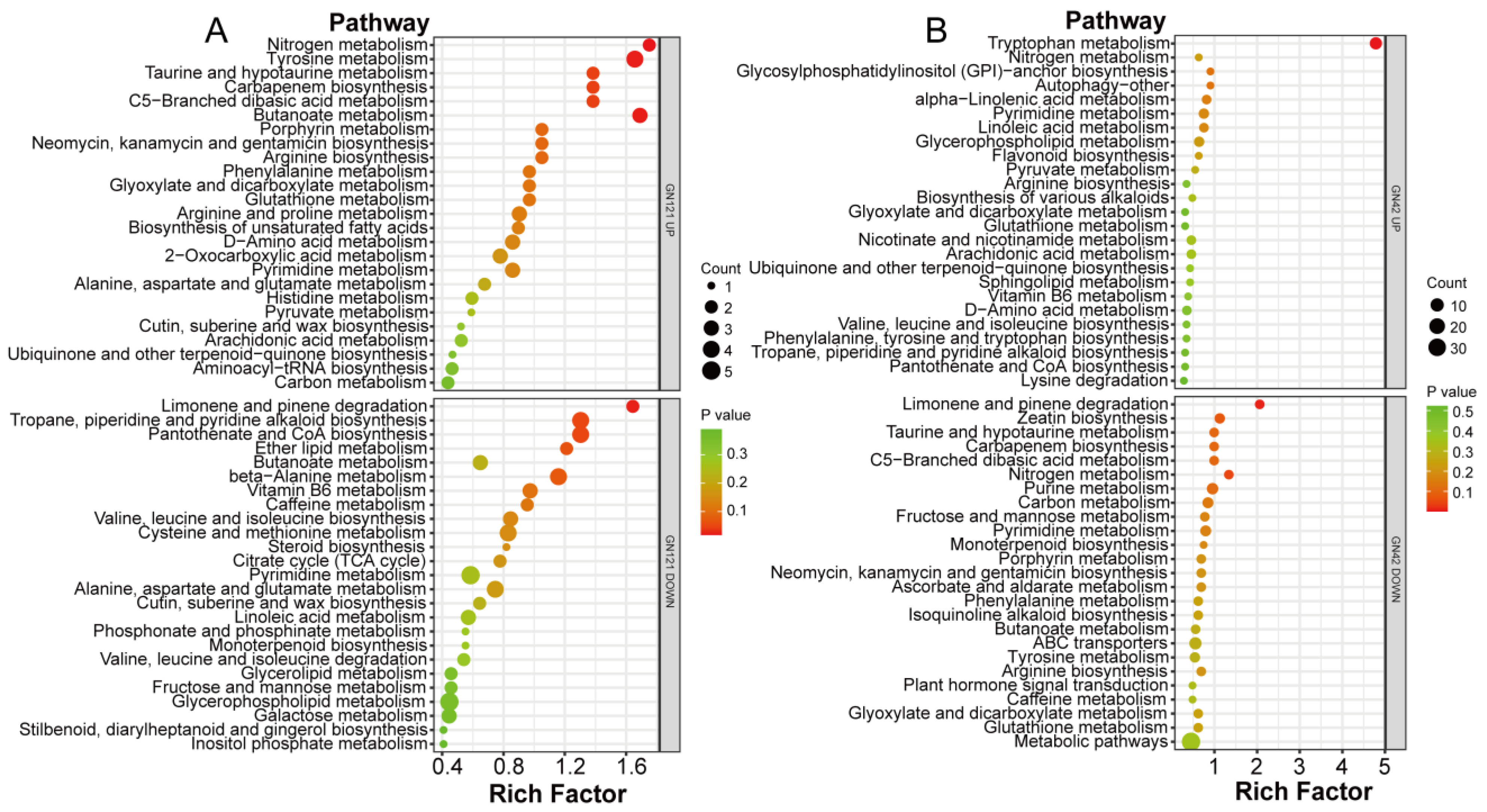
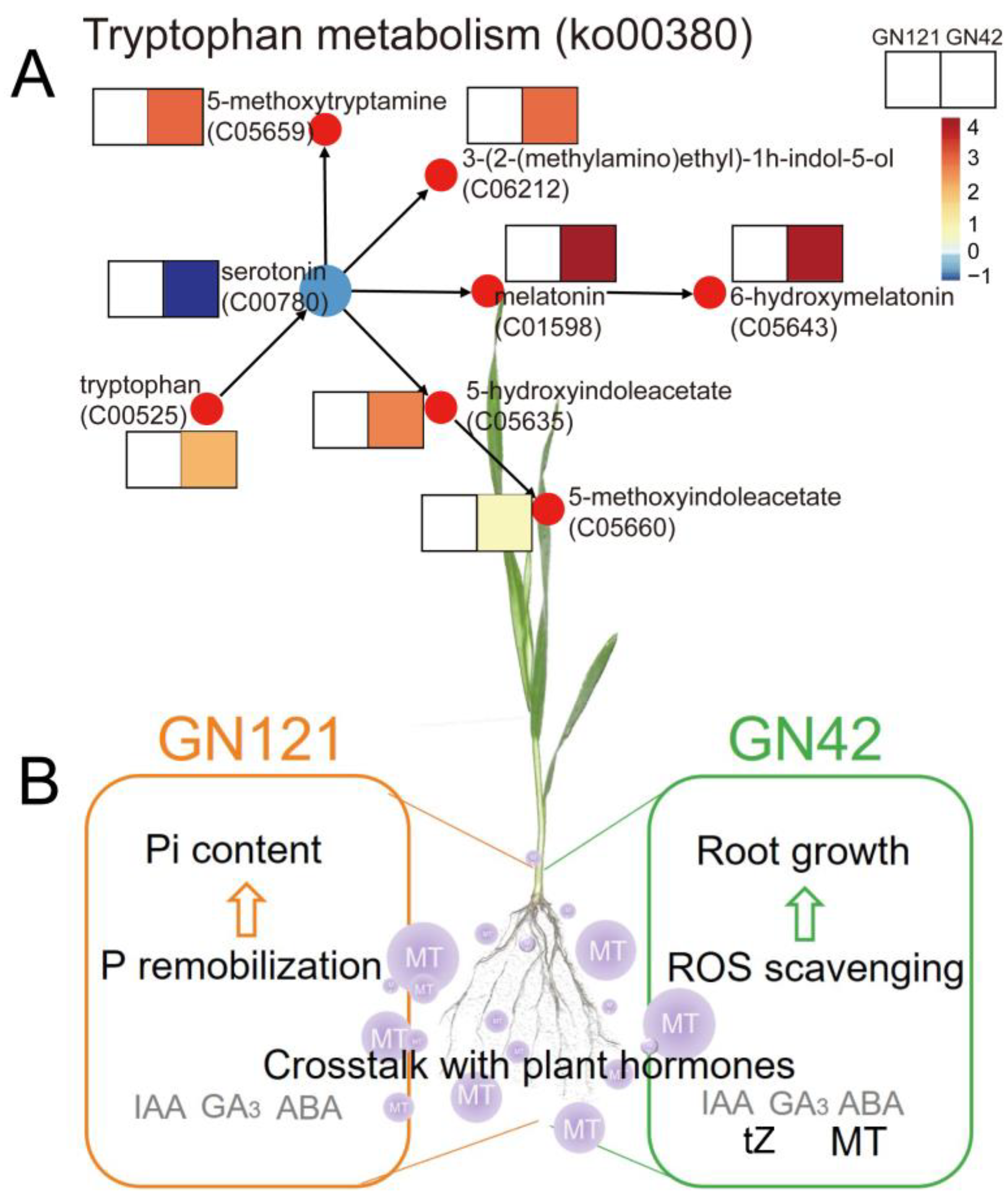
3.8. Metabolic Pathway Annotation of DAMs after Exogenous Melatonin Treatment
4. Discussion
4.1. Exogenous Melatonin and Its Crosstalk with Plant Hormones Regulates Barley Root Growth under LP Stress
4.2. Exogenous Melatonin Alleviated the Oxidative Stress of Barley Roots under LP Stress
4.3. Exogenous Melatonin Promoted the Root P Remobilization in LP-Tolerant Barley
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cong, W.F.; Suriyagoda, L.D.B.; Lambers, H. Tightening the phosphorus cycle through phosphorus-efficient crop genotypes. Trends Plant Sci. 2020, 25, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.; Goedde, V.; Niehaus, K.; Zoerb, C. Metabolic adaptations of white lupin roots and shoots under phosphorus deficiency. Front. Plant Sci. 2015, 6, 1014. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Shou, H.; Xu, G.; Lian, X. Improvement of phosphorus efficiency in rice on the basis of understanding phosphate signaling and homeostasis. Curr. Opin. Plant Biol. 2013, 16, 205–212. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, L.; Yu, D.; Xu, K.; Zhang, J.; Li, X.; Wang, P.; Chen, G.; Liu, Z.; Peng, C.; et al. Integrative analysis of the wheat PHT1 gene family reveals a novel member involved in arbuscular mycorrhizal phosphate transport and immunity. Cells 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Raghothama, K.G. Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef]
- Ragothama, K.G. Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Yang, J.; Xie, M.Y.; Yang, X.L.; Liu, B.H.; Lin, H.H. Phosphoproteomic profiling reveals the importance of CK2, MAPKs and CDPKs in response to phosphate starvation in rice. Plant Cell Physiol. 2019, 60, 2785–2796. [Google Scholar] [CrossRef]
- Sha, A.; Li, M.; Yang, P. Identification of phosphorus deficiency responsive proteins in a high phosphorus acquisition soybean (Glycine max) cultivar through proteomic analysis. Biochim. Biophys. Acta 2016, 1864, 427–434. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes1. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef]
- Byeon, Y.; Lee, H.Y.; Hwang, O.J.; Lee, H.J.; Lee, K.; Back, K. Coordinated regulation of melatonin synthesis and degradation genes in rice leaves in response to cadmium treatment. J. Pineal Res. 2015, 58, 470–478. [Google Scholar] [CrossRef]
- Hardeland, R. Taxon- and site-specific melatonin catabolism. Molecules 2017, 22, 2015. [Google Scholar] [CrossRef]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Saxena, P.K.; Murch, S.J. Melatonin in plant signalling and behaviour. Funct. Plant Biol. 2018, 45, 58–69. [Google Scholar] [CrossRef]
- Tan, D.-X. Melatonin and plants preface. J. Exp. Bot. 2015, 66, 625–626. [Google Scholar] [CrossRef]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Bai, Z.; et al. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar] [CrossRef]
- Tiryaki, I.; Keles, H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J. Pineal Res. 2012, 52, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, H.; Wang, B.; Zhang, Y.; Wang, J.; Cheng, C.; Huang, Y. Exogenous melatonin enhances cold resistance by improving antioxidant defense and cold-responsive genes’ expression in banana. Horticulturae 2022, 8, 260. [Google Scholar] [CrossRef]
- Li, C.; He, Q.; Zhang, F.; Yu, J.; Li, C.; Zhao, T.; Zhang, Y.; Xie, Q.; Su, B.; Mei, L.; et al. Melatonin enhances cotton immunity to Verticillium wilt via manipulating lignin and gossypol biosynthesis. Plant J. 2019, 100, 784–800. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liang, B.; Chang, C.; Wei, Z.; Zhou, S.; Ma, F. Exogenous melatonin improved potassium content in Malus under different stress conditions. J. Pineal Res. 2016, 61, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, Q.; Gong, B. Characterization of COMT1-mediated low phosphorus resistance mechanism by metabolomics in tomato plants. Environ. Exp. Bot. 2020, 179, 104187. [Google Scholar] [CrossRef]
- Chen, L.; Fan, J.; Hu, Z.; Huang, X.; Amombo, E.; Liu, A.; Bi, A.; Chen, K.; Xie, Y.; Fu, J. Melatonin is involved in regulation of bermudagrass growth and development and response to low K+ stress. Front. Plant Sci. 2017, 8, 2038. [Google Scholar] [CrossRef]
- Du, P.; Yin, B.; Cao, Y.; Han, R.; Ji, J.; He, X.; Liang, B.; Xu, J. Beneficial effects of exogenous melatonin and dopamine on low nitrate stress in Malus hupehensis. Front. Plant Sci. 2021, 12, 807472. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef]
- Sun, C.; Lv, T.; Huang, L.; Liu, X.; Jin, C.; Lin, X. Melatonin ameliorates aluminum toxicity through enhancing aluminum exclusion and reestablishing redox homeostasis in roots of wheat. J. Pineal Res. 2020, 68, e12642. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S. Melatonin alleviates cold-induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, eaba0196. [Google Scholar] [CrossRef] [PubMed]
- Spoor, W. Zohary D, Hopf M. 2000. Domestication of plants in the Old World. 3rd edn. 316pp. New York: Oxford University Press. 19.95 (softback). Ann. Bot. 2001, 88, 666. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Z.; Li, C.; Ren, P.; Yao, L.; Li, B.; Meng, Y.; Ma, X.; Si, E.; Yang, K.; et al. Dynamic responses of barley root succinyl-proteome to short-term phosphate starvation and recovery. Front. Plant Sci. 2021, 12, 649147. [Google Scholar] [CrossRef]
- Ahn, H.R.; Kim, Y.J.; Lim, Y.J.; Duan, S.; Eom, S.H.; Jung, K.H. Key genes in the melatonin biosynthesis pathway with circadian rhythm are associated with various abiotic stresses. Plants 2021, 10, 129. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J.; Cano, A.; Reiter, R.J. Melatonin and carbohydrate metabolism in plant cells. Plants 2021, 10, 1917. [Google Scholar] [CrossRef]
- Bahcesular, B.; Yildirim, E.D.; Karacocuk, M.; Kulak, M.; Karaman, S. Seed priming with melatonin effects on growth, essential oil compounds and antioxidant activity of basil (Ocimum basilicum L.) under salinity stress. Ind. Crops Prod. 2020, 146, 112165. [Google Scholar] [CrossRef]
- Yu, Y.; Lv, Y.; Shi, Y.; Li, T.; Chen, Y.; Zhao, D.; Zhao, Z. The role of phyto-melatonin and related metabolites in response to stress. Molecules 2018, 23, 1887. [Google Scholar] [CrossRef]
- Ganie, A.H.; Ahmad, A.; Pandey, R.; Aref, I.M.; Yousuf, P.Y.; Ahmad, S.; Iqbal, M. Metabolite profiling of low-P tolerant and low-P sensitive maize genotypes under phosphorus starvation and restoration conditions. PLoS ONE 2015, 10, e0129520. [Google Scholar] [CrossRef]
- Ham, B.K.; Chen, J.; Yan, Y.; Lucas, W.J. Insights into plant phosphate sensing and signaling. Curr. Opin. Biotechnol. 2018, 49, 1–9. [Google Scholar] [CrossRef]
- Luo, B.; Ma, P.; Nie, Z.; Zhang, X.; He, X.; Ding, X.; Feng, X.; Lu, Q.; Ren, Z.; Lin, H.; et al. Metabolite profiling and genome-wide association studies reveal response mechanisms of phosphorus deficiency in maize seedling. Plant J. 2019, 97, 947–969. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Ma, X.; Li, B.; Meng, Y.; Wang, H. Identification and selection of low-phosphate-tolerant germplasm in barley (Hordeum vulgare L.). Soil Sci. Plant Nutr. 2016, 62, 471–480. [Google Scholar] [CrossRef]
- Ren, P.; Meng, Y.; Li, B.; Ma, X.; Si, E.; Lai, Y.; Wang, J.; Yao, L.; Yang, K.; Shang, X.; et al. Molecular mechanisms of acclimatization to phosphorus starvation and recovery underlying full-length transcriptome profiling in barley (Hordeum vulgare L.). Front. Plant Sci. 2018, 9, 500. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D.; Pratt, P.F. Methods of analysis for soils, plants and waters. Soil Sci. 1962, 93, 68. [Google Scholar] [CrossRef]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef]
- Wang, C.F.; Huang, L.L.; Zhang, H.C.; Han, Q.M.; Buchenauer, H.; Kang, Z.S. Cytochemical localization of reactive oxygen species (O2− and H2O2) and peroxidase in the incompatible and compatible interaction of wheat—Puccinia striiformis f. sp. tritici. Physiol. Mol. Plant Pathol. 2010, 74, 221–229. [Google Scholar] [CrossRef]
- Zeng, F.; Wu, X.; Qiu, B.; Wu, F.; Jiang, L.; Zhang, G. Physiological and proteomic alterations in rice (Oryza sativa L.) seedlings under hexavalent chromium stress. Planta 2014, 240, 291–308. [Google Scholar] [CrossRef]
- Wang, J.; Yang, K.; Yao, L.; Ma, Z.; Li, C.; Si, E.; Li, B.; Meng, Y.; Ma, X.; Shang, X.; et al. Metabolomics analyses provide insights into nutritional value and abiotic stress tolerance in halophyte Halogeton glomeratus. Front. Plant Sci. 2021, 12, 703255. [Google Scholar] [CrossRef]
- Liu, D.; Qin, S.; Su, D.; Wang, K.; Huang, Y.; Huang, Y.; Pang, Y. Metabolic reprogramming of the right ventricle and pulmonary arteries in a flow-associated pulmonary arterial hypertension rat model. ACS Omega 2022, 7, 1273–1287. [Google Scholar] [CrossRef]
- Liu, Y.; Kong, Z.; Liu, J.; Zhang, P.; Wang, Q.; Huan, X.; Li, L.; Qin, P. Non-targeted metabolomics of quinoa seed filling period based on liquid chromatography-mass spectrometry. Food Res. Int. 2020, 137, 109743. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Y.; Zhao, X.; Jin, X.; Hou, L.; Shi, Y.; Ahammed, G.J. Silicon compensates phosphorus deficit-Induced growth inhibition by improving photosynthetic capacity, antioxidant potential, and nutrient homeostasis in tomato. Agronomy 2019, 9, 733. [Google Scholar] [CrossRef]
- Janas, K.M.; Posmyk, M.M. Melatonin, an underestimated natural substance with great potential for agricultural application. Acta Physiol. Plant. 2013, 35, 3285–3292. [Google Scholar] [CrossRef]
- Byeon, Y.; Park, S.; Lee, H.Y.; Kim, Y.S.; Back, K. Elevated production of melatonin in transgenic rice seeds expressing rice tryptophan decarboxylase. J. Pineal Res. 2014, 56, 275–282. [Google Scholar] [CrossRef]
- Murch, S.J.; Erland, L.A.E. A systematic review of melatonin in plants: An example of evolution of literature. Front. Plant Sci. 2021, 12, 683047. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, A.; Aftab, T. Phytohormones, plant growth regulators and signaling molecules: Cross-talk and stress responses. Plant Cell Rep. 2021, 40, 1301–1303. [Google Scholar] [CrossRef]
- Hernandez-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin: A growth-stimulating compound present in lupin tissues. Planta 2004, 220, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Qi, W.B.; Reiter, R.J.; Wei, W.; Wang, B.M. Exogenously applied melatonin stimulates root growth and raises endogenous indoleacetic acid in roots of etiolated seedlings of Brassica juncea. J. Plant Physiol. 2009, 166, 324–328. [Google Scholar] [CrossRef]
- Gao, T.; Liu, X.; Tan, K.; Zhang, D.; Zhu, B.; Ma, F.; Li, C. Introducing melatonin to the horticultural industry: Physiological roles, potential applications, and challenges. Hortic. Res. 2022, 9, uhac094. [Google Scholar] [CrossRef]
- Li, X.; Tan, D.X.; Jiang, D.; Liu, F. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat-induced leaf senescence involves changes in abscisic acid and cytokinin biosynthesis and signaling pathways in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar] [CrossRef]
- Chen, L.; Lu, B.; Liu, L.; Duan, W.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Li, C.; et al. Melatonin promotes seed germination under salt stress by regulating ABA and GA3 in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2021, 162, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Prerostova, S.; Kramna, B.; Dobrev, P.I.; Gaudinova, A.; Marsik, P.; Fiala, R.; Knirsch, V.; Vanek, T.; Kuresova, G.; Vankova, R. Organ-specific hormonal cross-talk in phosphate deficiency. Environ. Exp. Bot. 2018, 153, 198–208. [Google Scholar] [CrossRef]
- Silva-Navas, J.; Conesa, C.M.; Saez, A.; Navarro-Neila, S.; Garcia-Mina, J.M.; Zamarreno, A.M.; Baigorri, R.; Swarup, R.; Del Pozo, J.C. Role of cis-zeatin in root responses to phosphate starvation. New Phytol. 2019, 224, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Yang, L.; Chan, Z. Melatonin antagonizes cytokinin responses to stimulate root growth in Arabidopsis. J. Plant Growth Regul. 2023, 42, 1833–1845. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhang, M.; Xu, H.; Ning, K.; Wang, B.; Chen, M. Melatonin increases growth and salt tolerance of Limonium bicolor by improving photosynthetic and antioxidant capacity. BMC Plant Biol. 2022, 22, 16. [Google Scholar] [CrossRef]
- Wang, P.; Yin, L.; Liang, D.; Li, C.; Ma, F.; Yue, Z. Delayed senescence of apple leaves by exogenous melatonin treatment: Toward regulating the ascorbate-glutathione cycle. J. Pineal Res. 2012, 53, 11–20. [Google Scholar] [CrossRef]
- Chang, T.; Xi, Q.; Wei, X.; Xu, L.; Wang, Q.; Fu, J.; Ling, C.; Zuo, Y.; Zhao, Y.; He, H.; et al. Rhythmical redox homeostasis can be restored by exogenous melatonin in hulless barley (Hordeum vulgare L.var. nudum) under cold stress. Environ. Exp. Bot. 2022, 194, 104756. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, J.; Wang, W.; Wang, Y.; Xu, J.; Li, Z.; Zhao, X.; Fu, B. Integrated analysis of the transcriptome and metabolome revealed the molecular mechanisms underlying the enhanced salt tolerance of rice due to the application of exogenous melatonin. Front. Plant Sci. 2020, 11, 618680. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Duan, W.; Li, W.; Wang, Q.; Li, J.; Song, H.; Xu, X. Melatonin maintained higher contents of unsaturated fatty acid and cell membrane structure integrity in banana peel and alleviated postharvest chilling injury. Food Chem. 2022, 397, 133836. [Google Scholar] [CrossRef]
- Hayat, F.; Sun, Z.; Ni, Z.; Iqbal, S.; Xu, W.; Gao, Z.; Qiao, Y.; Tufail, M.A.; Jahan, M.S.; Khan, U.; et al. Exogenous melatonin improves cold tolerance of strawberry (Fragaria x ananassa Duch.) through modulation of DREB/CBF-COR pathway and antioxidant defense system. Horticulturae 2022, 8, 194. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Xu, B.; Li, J.; Huang, B. Exogenous melatonin suppresses dark-induced leaf senescence by activating the superoxide dismutase-catalase antioxidant pathway and down-regulating chlorophyll degradation in excised leaves of perennial ryegrass (Lolium perenne L.). Front. Plant Sci. 2016, 7, 1500. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prasad, M.N.V.; Achary, V.M.M.; Panda, B.B. Elucidation of lead-induced oxidative stress in Talinum triangulare roots by analysis of antioxidant responses and DNA damage at cellular level. Environ. Sci. Pollut. Res. 2013, 20, 4551–4561. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, G.; Liu, X. Amounts of malondialdehyde do not accurately represent the real oxidative level of all vegetable oils: A kinetic study of malondialdehyde formation. Int. J. Food Sci. Technol. 2019, 54, 412–423. [Google Scholar] [CrossRef]
- Meng, X.; Chen, W.W.; Wang, Y.Y.; Huang, Z.R.; Ye, X.; Chen, L.S.; Yang, L.T. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef]
- Chen, H.; Quan, W.; Liu, H.; Ding, G. Effects of Suillus luteus and S. bovinus on the physiological response and nutrient absorption of Pinus massoniana seedlings under phosphorus deficiency. Plant Soil 2022, 471, 577–590. [Google Scholar] [CrossRef]
- Murakami, M.; Kudo, I. Recent advances in molecular biology and physiology of the prostaglandin E2-biosynthetic pathway. Prog. Lipid Res. 2004, 43, 3–35. [Google Scholar] [CrossRef]
- Yang, S.Y.; Huang, T.K.; Kuo, H.F.; Chiou, T.J. Role of vacuoles in phosphorus storage and remobilization. J. Exp. Bot. 2017, 68, 3045–3055. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Dissanayaka, D.; Plaxton, W.C.; Lambers, H.; Siebers, M.; Marambe, B.; Wasaki, J. Molecular mechanisms underpinning phosphorus-use efficiency in rice. Plant Cell Environ. 2018, 41, 1483–1496. [Google Scholar] [CrossRef]
- Luo, J.; Liu, Y.; Zhang, H.; Wang, J.; Chen, Z.; Luo, L.; Liu, G.; Liu, P. Metabolic alterations provide insights into Stylosanthes roots responding to phosphorus deficiency. BMC Plant Biol. 2020, 20, 85. [Google Scholar] [CrossRef]
- Pant, B.D.; Pant, P.; Erban, A.; Huhman, D.; Kopka, J.; Scheible, W.R. Identification of primary and secondary metabolites with phosphorus status-dependent abundance in Arabidopsis, and of the transcription factor PHR1 as a major regulator of metabolic changes during phosphorus limitation. Plant Cell Environ. 2015, 38, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Roessner, U.; Eickmeier, I.; Genc, Y.; Callahan, D.L.; Shirley, N.; Langridge, P.; Bacic, A. Metabolite profiling reveals distinct changes in carbon and nitrogen metabolism in phosphate-deficient barley plants (Hordeum vulgare L.). Plant Cell Physiol. 2008, 49, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, M.M.; Carr, J.M.; Williamson, C.L.; Slocum, R.D. Effects of phosphate limitation on expression of genes involved in pyrimidine synthesis and salvaging in Arabidopsis. Plant Physiol. Biochem. 2005, 43, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Mehra, P.; Pandey, B.K.; Giri, J. Comparative morphophysiological analyses and molecular profiling reveal Pi-efficient strategies of a traditional rice genotype. Front. Plant Sci. 2015, 6, 1184. [Google Scholar] [CrossRef]
- Miao, N.; Zhou, J.; Li, M.; Zhang, J.; Hu, Y.; Guo, J.; Zhang, T.; Shi, L. Remodeling and protecting the membrane system to resist phosphorus deficiency in wild soybean (Glycine soja) seedling leaves. Planta 2022, 255, 53. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Index | Log2_FC * | MS2_Name |
|---|---|---|---|
| GN121 | M403T35_NEG | 0.8825 | 1-o-(9z-octadecenyl)-sn-glycero-2,3-cyclic-phosphate |
| M811T39_POS | 0.3398 | 1-octadecanoyl-2-octadecenoyl-sn-glycero-3-phosphocholine | |
| M715T38_1_NEG | −0.4421 | 2-linoleoyl-1-palmitoyl-sn-glycero-3-phosphoethanolamine | |
| M431T245_NEG | −0.6881 | 1-Palmitoyl Lysophosphatidic Acid | |
| M618T36_NEG | −0.719 | N-palmitoyl-d-erythro-dihydroceramide-1-phosphate | |
| M752T151_2_NEG | −0.7461 | Pe (16:0/8-hepe) | |
| M757T146_1_POS | −0.9683 | PC (16:0/16:0) | |
| M860T193_NEG | −1.0082 | Pi 36:3 | |
| M409T252_NEG | −1.1 | 1-hexadecanoyl-2-sn-glycero-3-phosphate | |
| M509T145_NEG | −1.1835 | 1-oleoyl-2-hydroxy-sn-glycero-3-phospho-(1′-rac-glycerol) | |
| M241T395_NEG | −1.2173 | alpha.-d-galactose 1-phosphate | |
| M322T432_NEG | −1.2436 | Cytidine 3′-monophosphate | |
| M850T198_POS | −1.2817 | 1,2-dipalmitoyl-sn-glycero-3-phospho-(1′-myo-inositol) | |
| M774T71_NEG | −1.3111 | 1,2-distearoyl-sn-glycero-3-phospho-l-serine | |
| M328T265_NEG | −1.3342 | Adenosine 3′,5′-cyclic monophosphate | |
| M323T412_1_NEG | −1.4557 | Uridine 5′-monophosphate (UMP) | |
| M304T319_NEG | −1.482 | Cytidine 2′,3′-cyclic phosphate | |
| M428T93_POS | −1.5782 | Arachidonoyl ethanolamide phosphate | |
| M330T266_POS | −1.6632 | Adenosine 2′,3′-cyclic monophosphate | |
| M520T186_2_POS | −1.9729 | Lpc 18:2 | |
| M184T486_POS | −2.1167 | 1-o-hexadecyl-2-deoxy-2-thio-s-acetyl-sn-glyceryl-3-phosphorylcholine | |
| M214T390_NEG | −2.2073 | sn-Glycerol 3-phosphoethanolamine | |
| M258T384_POS | −2.3738 | Glycerophosphocholine | |
| M171T430_NEG | −2.4824 | Glycerophosphate (2) | |
| M243T470_POS | −3.0013 | glucose 6-phosphate | |
| M259T468_NEG | −3.9351 | mannose 6-phosphate | |
| GN42 | M743T183_NEG | 2.6019 | 1-stearoyl-2-linoleoyl-sn-glycero-3-phosphoethanolamine |
| M753T183_POS | 1.9118 | 1,2-dipalmitoleoyl-sn-glycero-3-phosphocholine | |
| M767T183_POS | 1.8359 | 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine | |
| M821T200_1_NEG | 1.4875 | 1-stearoyl-2-linoleoyl-sn-glycero-3-phosphocholine | |
| M508T186_POS | 1.2428 | 1-(1z-octadecenyl)-sn-glycero-3-phosphocholine | |
| M520T186_2_POS | 1.0943 | Lpc 18:2 | |
| M835T39_1_POS | 0.3304 | 1-stearoyl-2-docosahexaenoyl-sn-glycero-3-phosphocholine | |
| M452T200_NEG | 0.3301 | 1-palmitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamine | |
| M811T39_POS | 0.2905 | 1-octadecanoyl-2-octadecenoyl-sn-glycero-3-phosphocholine | |
| M279T109_NEG | −0.3926 | 3-Deoxy-2-keto-6-phosphogluconic acid | |
| M774T71_NEG | −0.4553 | 1,2-distearoyl-sn-glycero-3-phospho-l-serine | |
| M715T38_1_NEG | −0.5803 | 2-linoleoyl-1-palmitoyl-sn-glycero-3-phosphoethanolamine | |
| M841T144_NEG | −0.767 | Pc (16:0e/8-hepe) | |
| M817T146_NEG | −1.0013 | Pc (16:1e/9-hode) | |
| M362T437_NEG | −1.5738 | Guanosine 5′-monophosphate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Yang, K.; Wang, J.; Ma, J.; Yao, L.; Si, E.; Li, B.; Ma, X.; Shang, X.; Meng, Y.; et al. Exogenous Melatonin Enhances the Low Phosphorus Tolerance of Barley Roots of Different Genotypes. Cells 2023, 12, 1397. https://doi.org/10.3390/cells12101397
Ma Z, Yang K, Wang J, Ma J, Yao L, Si E, Li B, Ma X, Shang X, Meng Y, et al. Exogenous Melatonin Enhances the Low Phosphorus Tolerance of Barley Roots of Different Genotypes. Cells. 2023; 12(10):1397. https://doi.org/10.3390/cells12101397
Chicago/Turabian StyleMa, Zengke, Ke Yang, Juncheng Wang, Jingwei Ma, Lirong Yao, Erjing Si, Baochun Li, Xiaole Ma, Xunwu Shang, Yaxiong Meng, and et al. 2023. "Exogenous Melatonin Enhances the Low Phosphorus Tolerance of Barley Roots of Different Genotypes" Cells 12, no. 10: 1397. https://doi.org/10.3390/cells12101397
APA StyleMa, Z., Yang, K., Wang, J., Ma, J., Yao, L., Si, E., Li, B., Ma, X., Shang, X., Meng, Y., & Wang, H. (2023). Exogenous Melatonin Enhances the Low Phosphorus Tolerance of Barley Roots of Different Genotypes. Cells, 12(10), 1397. https://doi.org/10.3390/cells12101397