
Identification of CTLA-4-Positive Cells in the Human Tonsil


Abstract
1. Introduction
2. Materials and Methods
2.1. Case Selection
2.2. Tissue Probe Stainings
2.3. Immunohistochemistry
2.4. Quantification of CTLA-4-Positive Cells in Tonsillar Compartments
2.5. Controls
2.6. Image Acquisition
2.7. Data Availability
3. Results
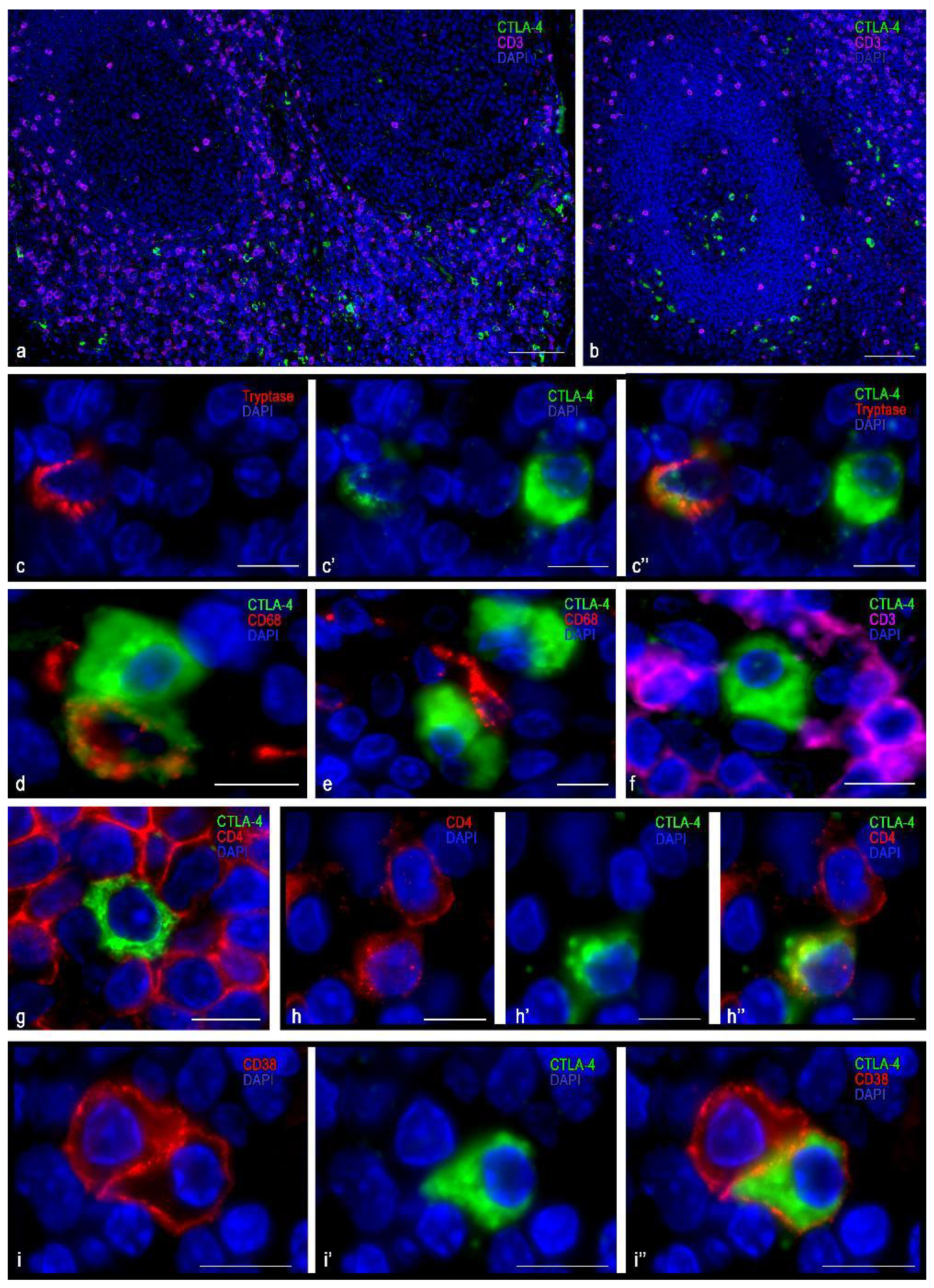
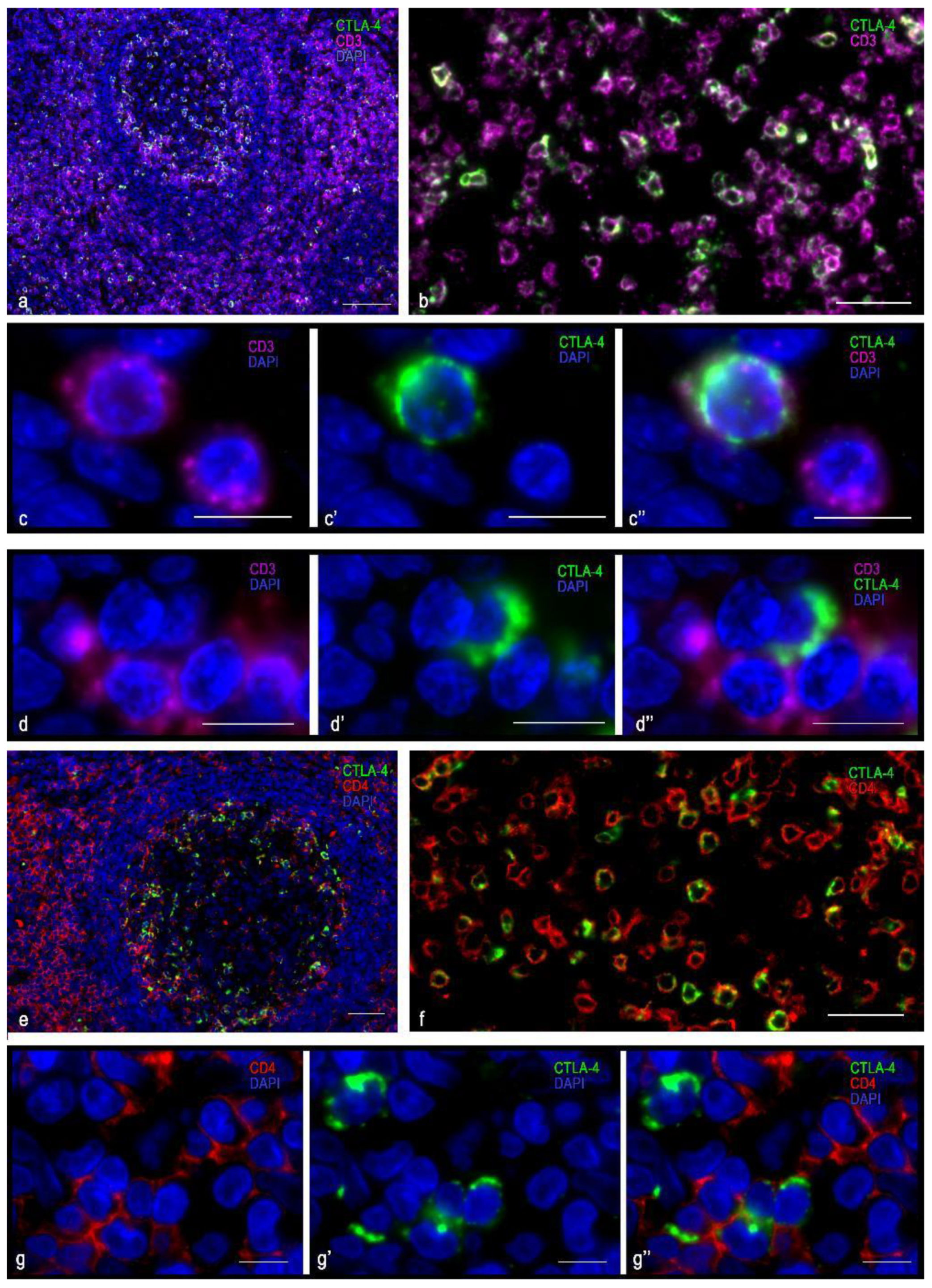
3.1. Immunomorphological and Histotopographic Characteristics of CTLA-4-Positive Cells Identified with Antibodies of the SP355 Clone
3.2. Immunomorphological and Histotopographic Characteristics of CTLA-4-Positive Cells Identified with Antibodies of the UMAB249 Clone
3.3. Immunomorphological and Histotopographic Characteristics of CTLA-4-Positive Cells Identified with Antibodies of the CAL49 Clone
3.4. Immunomorphological and Histotopographic Characteristics of CTLA-4-Positive Cells Identified with Antibodies of the AA 57-86 Clone
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PD-1 | Programmed death-1 and its ligand PD-L1 |
| CTLA-4 | Cytotoxic T-lymphocyte-associated protein 4 |
References
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A new member of the immunoglobulin superfamily—CTLA-4. Nature 1987, 328, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Peach, R.J.; Bajorath, J.; Naemura, J.; Leytze, G.; Greene, J.; Aruffo, A.; Linsley, P.S. Both extracellular immunoglobin-like domains of CD80 contain residues critical for binding T cell surface receptors CTLA-4 and CD28. J. Biol. Chem. 1995, 270, 21181–21187. [Google Scholar] [CrossRef]
- Linsley, P.S.; Nadler, S.G.; Bajorath, J.; Peach, R.; Leung, H.T.; Rogers, J.; Bradshaw, J.; Stebbins, M.; Leytze, G.; Brady, W.; et al. Binding stoichiometry of the cytotoxic T lymphocyte-associated molecule-4 (CTLA-4). A disulfide-linked homodimer binds two CD86 molecules. J. Biol. Chem. 1995, 270, 15417–15424. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.A.; Punt, J.; Stranford, S.A.; Jones, P.P.; Kuby, J. Kuby Immunology, 7th ed.; Freeman and Company: New York, NY, USA, 2013. [Google Scholar]
- Peggs, K.S.; Quezada, S.A.; Chambers, C.A.; Korman, A.J.; Allison, J.P. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti–CTLA-4 antibodies. J. Exp. Med. 2009, 206, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 can function as a negative regulator of T cell activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Chambers, C.A.; Krummel, M.F.; Boitel, B.; Hurwitz, A.; Sullivan, T.J.; Fournier, S.; Cassell, D.; Brunner, M.; Allison, J.P. The role of CTLA-4 in the regulation and initiation of T-cell responses. Immunol. Rev. 1996, 153, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Leach, D.R.; Krummel, M.F.; Allison, J.P. Enhancement of antitumor immunity by CTLA-4 blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef]
- Oosterwegel, M.A.; Greenwald, R.J.; Mandelbrot, D.A.; Lorsbach, R.B.; Sharpe, A.H. CTLA-4 and T cell activation. Curr. Opin. Immunol. 1999, 11, 294–300. [Google Scholar] [CrossRef]
- Syn, N.L.; Teng, M.W.L.; Mok, T.S.K.; Soo, R.A. De-novo and acquired resistance to immune checkpoint targeting. Lancet Oncol. 2017, 18, e731–e741. [Google Scholar] [CrossRef]
- Hazarika, M.; Chuk, M.K.; Theoret, M.R.; Mushti, S.; He, K.; Weis, S.L.; Putman, A.H.; Helms, W.S.; Cao, X.; Li, H.; et al. U.S. FDA approval summary: Nivolumab for treatment of unresectable or metastatic melanoma following progression on ipilimumab. Clin. Cancer Res. 2017, 23, 3484–3488. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.B.; Feng, Z.; Bifulco, C.B.; Leidner, R.; Weinberg, A.; Fox, B.A. Immunotherapy. In Oral, Head and Neck Oncology and Reconstructive Surgery; Bell, R.B., Ferndandes, R., Andersen, P.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 314–340. [Google Scholar]
- Halpert, M.; Konduri, V.; Liang, D.; Chen, Y.; Wing, J.; Paust, S.; Levitt, J.; Decker, W. Dendritic cell-secreted cytotoxic T-lymphocyte-associated protein-4 regulates the T-cell response by downmodulating bystander surface B7. Stem Cells Dev. 2016, 25, 774–787. [Google Scholar] [CrossRef] [PubMed]
- Contardi, E.; Palmisano, G.L.; Tazzari, P.L.; Martelli, A.M.; Fala, F.; Fabbi, M.; Kato, T.; Lucarelli, E.; Donati, D.; Polito, L.; et al. CTLA-4 is constitutively expressed on tumor cells and can trigger apoptosis upon ligand interaction. Int. J. Cancer 2005, 117, 538–550. [Google Scholar] [CrossRef]
- Gao, Y.H.; Wang, P.; Takagi, K.; Shimozato, O.; Yagita, H.; Okigaki, T.; Matasumura, M. Expression of a soluble form of CTLA4 on macrophage and its biological activity. Cell Res. 1999, 9, 189–199. [Google Scholar] [CrossRef]
- Pistillo, M.P.; Tazzari, P.L.; Palmisano, G.L.; Pierri, I.; Bolognesi, A.; Ferlito, F.; Capanni, P.; Polito, L.; Ratta, M.; Pileri, S.; et al. CTLA-4 is not restricted to the lymphoid cell lineage and can function as a target molecule for apoptosis induction of leukemic cells. Blood 2003, 101, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Oyewole-Said, D.; Konduri, V.; Vazquez-Perez, J.; Weldon, S.A.; Levitt, J.M.; Decker, W.K. Beyond T-cells: Functional characterization of CTLA-4 expression in immune and non-immune cell types. Front. Immunol. 2020, 11, 608024. [Google Scholar] [CrossRef] [PubMed]
- Postow, M.A.; Callahan, M.K.; Wolchok, J.D. Immune checkpoint blockade in cancer therapy. J. Clin. Oncol. 2015, 33, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 pathways: Similarities, differences, and implications of their inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Buchwalow, I.B.; Boecker, W. Immunohistochemistry: Basics and Methods, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Buchwalow, I.; Samoilova, V.; Boecker, W.; Tiemann, M. Non-specific binding of antibodies in immunohistochemistry: Fallacies and facts. Sci. Rep. 2011, 1, 28. [Google Scholar] [CrossRef]
- Buchwalow, I.; Atiakshin, D.; Samoilova, V.; Boecker, W.; Tiemann, M. Identification of autofluorescent cells in human angioimmunoblastic T-cell lymphoma. Histochem. Cell Biol. 2018, 149, 169–177. [Google Scholar] [CrossRef]
- Buchwalow, I.; Samoilova, V.; Boecker, W.; Tiemann, M. Multiple immunolabeling with antibodies from the same host species in combination with tyramide signal amplification. Acta Histochem. 2018, 120, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Hubbard, A.; Jones, T.; Racolta, A.; Bhaumik, S.; Cummins, N.; Zhang, L.; Garsha, K.; Ventura, F.; Lefever, M.R.; et al. Fully automated 5-plex fluorescent immunohistochemistry with tyramide signal amplification and same species antibodies. Lab. Investig. 2017, 97, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Sekhavati, F.; Cardenes, R.; Windmueller, C.; Dacosta, K.; Rodriguez-Canales, J.; Steele, K.E. CTLA-4 immunohistochemistry and quantitative image analysis for profiling of human cancers. J. Histochem. Cytochem. 2019, 67, 901–918. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies/Clone | Host | Source | Dilution |
|---|---|---|---|
| CD1a (EP362R) | rabbit monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD3 (EPS1) | mouse monoclonal Ab | AbCam, United Kingdom | 1:50 |
| CD3 (E2GV6) | rabbit monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD4 (SP35) | rabbit monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD8 (ESP57) | rabbit monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD14 (EPR3653) | rabbit monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD15 (MMA) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD16 (SP175) | rabbit monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD20 (L26) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD34 (QBEnd/10) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD38 (SP149) | rabbit monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD45 (LCA) | mouse monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD57 (NK-1) | mouse monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD68 (EKP-1) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CD117 (A4502) | rabbit monoclonal Ab | DAKO/Leica | Ready-to-use |
| CD138 (B-A38) | mouse monoclonal Ab | Cell Marque, USA | Ready-to-use |
| CD163 (MRQ-26) | mouse monoclonal Ab | Cell Marque, USA | Ready-to-use |
| Tryptase (AA1) | mouse monoclonal Ab | AbCam, UK | 1:3000 |
| Podoplanin (D2-40) | mouse monoclonal Ab | Cell Marque, USA | Ready-to-use |
| Vimentin (V9) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| Pan Cytokeratin (AE1/AE3/PCK26) | mouse monoclonal Ab | Ventana/Roche, Germany | Ready-to-use |
| CTLA-4 (UMAB249) | mouse monoclonal Ab | Origene Technologies | 1:50 |
| CTLA-4 (AA 57-86) | rabbit polyclonal Ab | antikoerper-online, Germany | 1:200 |
| CTLA-4 (SP355) | rabbit monoclonal Ab | AbCam, United Kingdom | 1:100 |
| CTLA-4 (CAL49) | rabbit monoclonal Ab | AbCam, United Kingdom | 1:300 |
| Antibodies and Other Reagents | Source | Dilution | Label |
|---|---|---|---|
| Goat anti-mouse IgG Ab (#A21236) | Invitrogen, Darmstadt, Germany | 1/200 | Alexa Fluor 647 |
| Goat anti-rabbit IgG Ab (#A21245) | Invitrogen, Darmstadt, Germany | 1/200 | Alexa Fluor 647 |
| Goat anti-mouse IgG Ab (#115-165-166) | Jackson ImmunoResearch | 1/200 | Cy3 |
| Goat anti-rabbit IgG Ab (#A-11034) | Invitrogen Darmstadt, Germany | 1/200 | Cy3 |
| Goat anti-mouse IgG Ab (#A-11029 | Invitrogen Darmstadt, Germany | 1/200 | Alexa Fluor 488 |
| Goat anti-rabbit IgG Ab (#A-11034) | Invitrogen Darmstadt, Germany | 1/200 | Alexa Fluor 488 |
| AmpliStain anti-Mouse 1-Step HRP (#AS-M1-HRP) | SDT GmbH, Baesweiler, Germany | ready-to-use | HRP |
| AmpliStain anti-Rabbit 1-Step HRP (#AS-R1-HRP) | SDT GmbH, Baesweiler, Germany | ready-to-use | HRP |
| 4′,6-diamidino-2-phenylindole (DAPI, #D9542-5MG) | Sigma, Hamburg, Germany | 5 µg/ml | w/o |
| VECTASHIELD Mounting Medium (#H-1000) | Vector Laboratories, Burlingame, CA, USA | ready-to-use | w/o |
| CC2 solution (# 950-223) | Ventana | ready-to-use | w/o |
| TSA Plus Fluorescein (#NEL741E001K) | PerkinElmer, Rodgau Germany | 1/200 | Fluorescein |
| TSA Plus Cyanine 3 (#NEL744E001KT) | PerkinElmer, Rodgau Germany | 1/200 | Cyanine 3 |
| TSA Plus Cyanine 5 (#NEL745E001KT) | PerkinElmer, Rodgau Germany | 1/200 | Cyanine 5 |
| CTLA4 Clones | UMAB249 | AA 57-86 | SP355 | CAL49 | ||
|---|---|---|---|---|---|---|
| Overall count of CTLA-4-positive cells in the human tonsil | 29.4/mm2 | 3.0/mm2 | 10.5/mm2 | 127.4/mm2 | ||
| Comparative distribution of CTLA-4-positive cells in tonsillar compartments | ||||||
| Reticulated crypt epithelium (lymphoepithelium) | +++ | ++ | * | * | ||
| Stratified squamous non-keratinized epithelium | +++ | + | * | * | ||
| Lymphoid follicle | Mantle zone | − | − | * | +++ | |
| Germinal center | Dark zone | * | − | + | +++ | |
| Apical light zone | − | * | + | ++ | ||
| Basal light zone | − | * | + | ++ | ||
| Extrafollicular region | +++ | +++ | +++ | +++ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiemann, M.; Atiakshin, D.; Samoilova, V.; Buchwalow, I. Identification of CTLA-4-Positive Cells in the Human Tonsil. Cells 2021, 10, 1027. https://doi.org/10.3390/cells10051027
Tiemann M, Atiakshin D, Samoilova V, Buchwalow I. Identification of CTLA-4-Positive Cells in the Human Tonsil. Cells. 2021; 10(5):1027. https://doi.org/10.3390/cells10051027
Chicago/Turabian StyleTiemann, Markus, Dmitri Atiakshin, Vera Samoilova, and Igor Buchwalow. 2021. "Identification of CTLA-4-Positive Cells in the Human Tonsil" Cells 10, no. 5: 1027. https://doi.org/10.3390/cells10051027
APA StyleTiemann, M., Atiakshin, D., Samoilova, V., & Buchwalow, I. (2021). Identification of CTLA-4-Positive Cells in the Human Tonsil. Cells, 10(5), 1027. https://doi.org/10.3390/cells10051027