
Epichloë gansuensis Enhances Achnatherum inebrians Seedlings Growth and Antioxidant Capacity Under UV-B Stress



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Experimental Materials
2.2. Experimental Design
2.3. Measurements
2.4. Data Analysis
3. Results
3.1. Growth Parameters of E+ and E− A. inebrians Seedlings Under UV-B Radiation
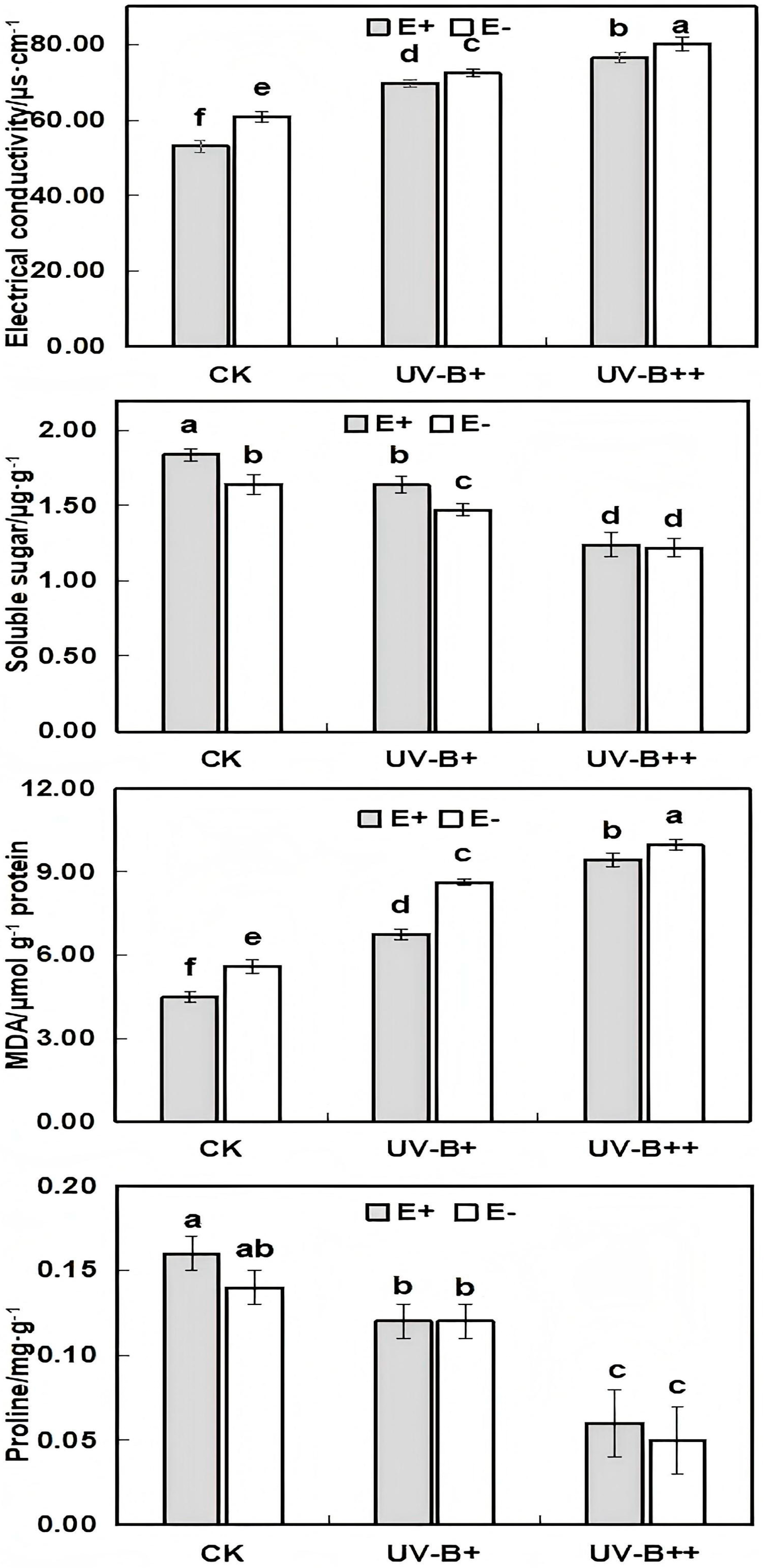
3.2. Membrane Lipid Peroxidation Levels in E+ and E− A. inebrians Seedlings Under UV-B RADIATION
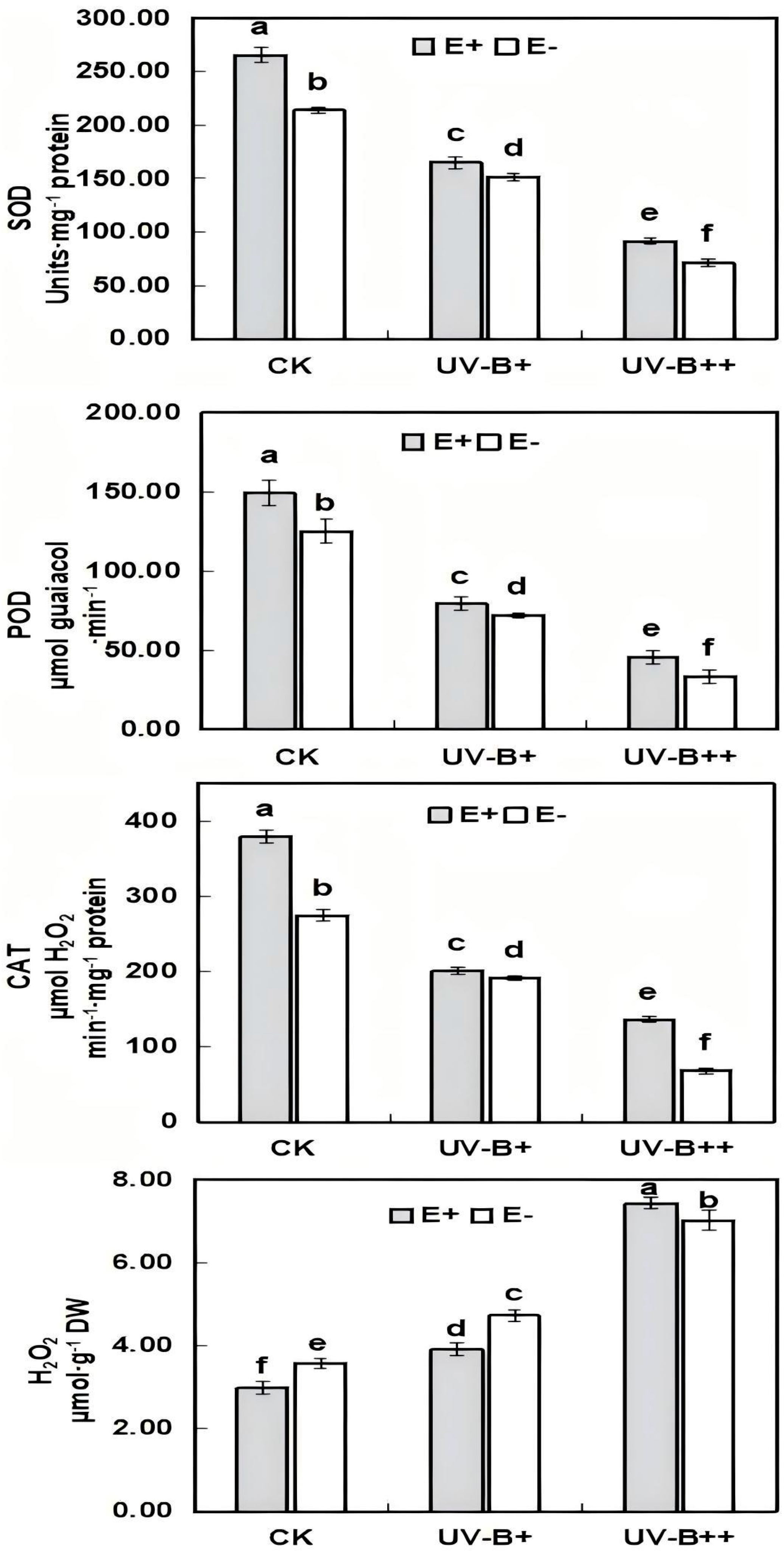
3.3. Activities of Antioxidant Enzymes in E+ and E− A. inebrians Seedlings Under UV-B Radiation
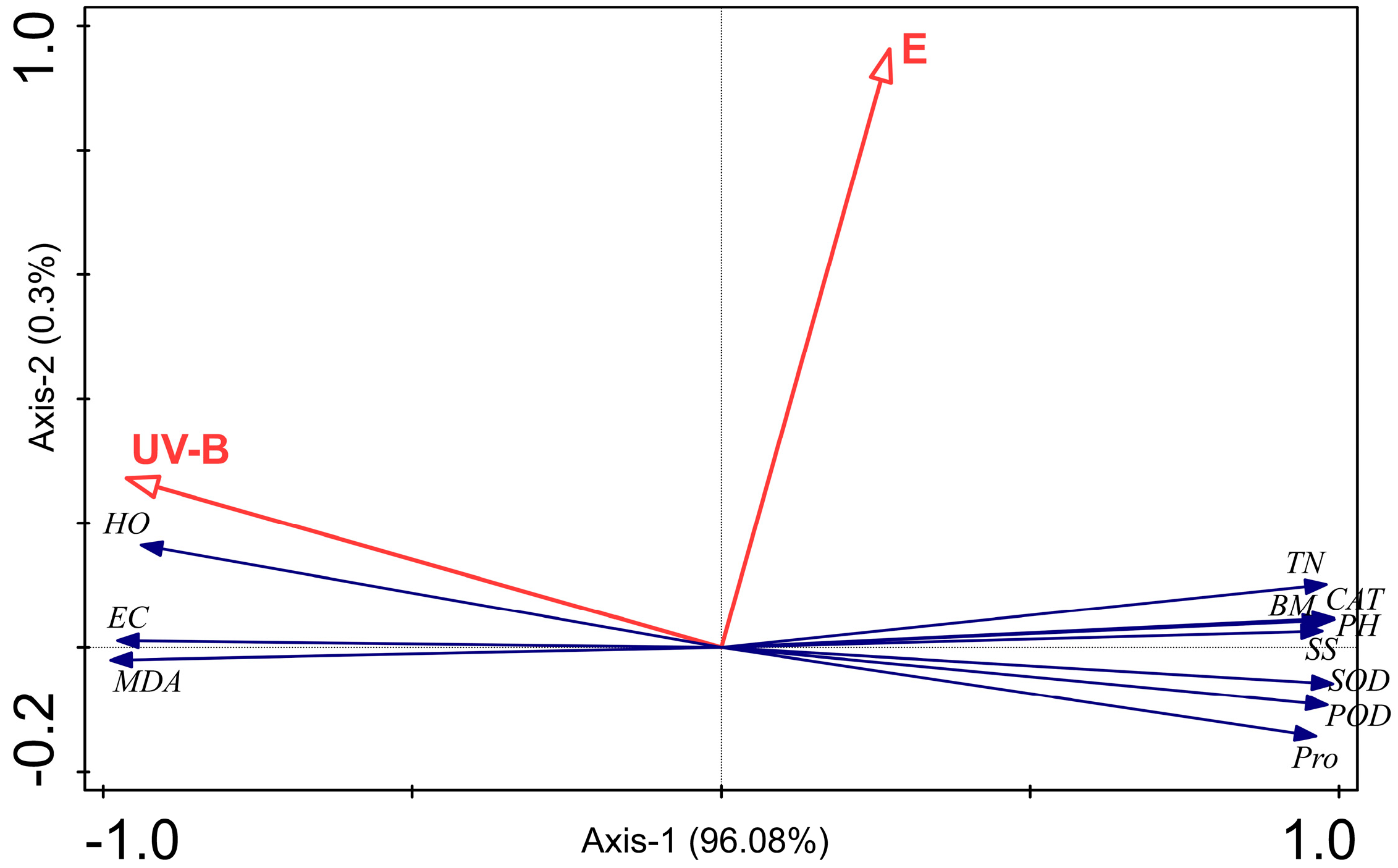
3.4. RDA Analysis Among E. gansuensis Treatment, UV-B Treatment, Plant Growth, Lipid Peroxidation, and Antioxidant Capacity
4. Discussion
4.1. Effect of UV-B Radiation on the Growth of E+ and E− A. inebrians Seedlings
4.2. Effect of UV-B Radiation on Membrane Lipid Peroxidation in E+ and E− A. inebrians Seedlings
4.3. Effect of UV-B Radiation on Antioxidant Enzyme Systems of E+ and E− A. inebrians Seedlings
4.4. Relationship Among E. gansuensis Treatment, UV-B Treatment, Plant Growth, Lipid Peroxidation, and Antioxidant Capacity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- National Aeronautics and Space Administration. NASA Ozone Watch: Annual Records. Available online: https://ozonewatch.gsfc.nasa.gov/statistics/annual_data.html (accessed on 3 June 2025).
- Grooss, J.U.; Müller, R. Simulation of record arctic stratospheric ozone depletion in 2020. J. Geophys. Res.-Atmos. 2021, 126, e2020JD033339. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, M.; Li, D.X.; You, M.H.; Yan, J.J.; Bai, S.Q. Metabolomic analysis of Elymus sibiricus exposed to UV-B radiation stress. Pratac. Sci. 2024, 29, 5133. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Qing, Z. Effect of enhanced UV-B radiation on plant flavonoids. Chin. J. Eco-Agric. 2007, 15, 191–194. [Google Scholar]
- Caldwell, M.M.; Bornman, J.F.; Ballaré, C.L.; Flint, S.D.; Kulandaivelu, G. Terrestrial ecosystems, increased solar ultraviolet radiation, and interactions with bother climate change factors. Photochem. Photobiol. Sci. 2007, 6, 252–266. [Google Scholar] [CrossRef]
- Dai, Q.J.; Yan, B.; Huang, S.B.; Liu, X.Z.; Peng, S.B.; Miranda, M.L.L.; Chavez, A.Q.; Vergara, B.S.; Olszyk, D.M. Response of oxidative stress defense systems in rice (Oryza sativa) leaves with supplemental UV-B radiation. Physiol. Plant. 1997, 101, 301–308. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Zhao, D.; Gao, W. Senescence and hyperspectral reflectance of cotton leaves exposed to ultraviolet-B radiation and carbon dioxide. Physiol. Plant. 2004, 121, 250–257. [Google Scholar] [CrossRef]
- Li, Y.S.; Jin, J.; Wang, G.H.; Liu, X.B. Preventive mechanisms to UV- B radiation damages in Crops. Soils Crops 2016, 5, 223–233. [Google Scholar]
- Rastogi, R.P.; Singh, S.P.; Incharoensakdi, A.; Häder, D.-P.; Sinha, R.P. Ultraviolet radiation-induced generation of reactive oxygen species, DNA damage and induction of UV-absorbing compounds in the cyanobacterium Rivularia sp. HKAR-4. S. Afr. J. Bot. 2014, 90, 163–169. [Google Scholar] [CrossRef]
- McLeod, A.R.; Rey, A.; Newsham, K.K.; Lewis, G.C.; Wolferstan, P. Effects of elevated ultraviolet radiation and endophytic fungi on plant growth and insect feeding in Lolium perenne, Festuca rubra, F. arundinacea and F. pratensis. J. Photochem. Photobiol. B. 2001, 62, 97–107. [Google Scholar] [CrossRef]
- Caldwell, M.; Teramura, A.H.; Tevini, M.; Bornman, J.F.; Bjorn, L.O.; Kulandaivelu, G. Effects of increased solar ultraviolet-radiation on terrestrial plants. Ambio 1995, 24, 166–173. [Google Scholar]
- Liu, M.Y.; Sun, W.J.; Ma, Z.T.; Guo, C.C.; Chen, J.H.; Wu, Q.; Wang, X.Y.; Chen, H. Integrated network analyses identify MYB4R1 neofunctionalization in the UV-B adaptation of Tartary buckwheat. Plant Commun. 2022, 3, 100414. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liu, H.T. How plants protect themselves from ultraviolet-B radiation stress. Plant Physiol. 2021, 187, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jia, Z.P.; Wang, J.; Du, M.N.; Su, X. Response of antioxidant enzyme system and total flavonoid of woody plant Hippophae neurocarpa seedlings to enhanced UV-B radiation. Guihaia 2020, 40, 1595–1601. [Google Scholar]
- Shi, S.B.; Shang, Y.X.; Zhu, P.J.; Yang, L. Effects of short term enhanced UV-B radiation on the PS II photochemical efficiency of alpine plant Saussurea superba. Chin. J. Appl. Ecol. 2011, 22, 1147–1154. [Google Scholar] [CrossRef]
- Zhou, X.J.; Luo, C.; Li, W.L.; Shi, J.E. Changes of total ozone in whole China and its low contents center in Qing-Zang Plateau regions. Chin. Sci. Bull. 1995, 1396–1398. [Google Scholar]
- Liu, Y.; Li, W.L. Deepening of ozone valley over Tibetan Plateau and its possible influences. Acta Meteorol. Sin. 2001, 97–106. [Google Scholar]
- Zhang, P.P. Characteristic of Endophytic Fungi Isolated from Elymus and Their Effect on Host Resistances. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2013. [Google Scholar]
- Pennell, C.G.L.; Popay, A.J.; Rolston, M.P.; Townsend, R.J.; Lloyd-West, C.M.; Card, S.D. Avanex unique endophyte technology: Reduced insect food source at airports. Environ. Entomol. 2016, 45, 101–108. [Google Scholar] [CrossRef]
- Song, M.L.; Chai, Q.; Li, X.Z.; Yao, X.; Li, C.J.; Christensen, M.J.; Nan, Z.B. An asexual Epichloë endophyte modifies the nutrient stoichiometry of wild barley (Hordeum brevisubulatum) under salt stress. Plant Soil 2015, 387, 153–165. [Google Scholar] [CrossRef]
- Tian, P.; Nan, Z.B.; Li, C.J.; Spangenberg, G. Effect of the endophyte Neotyphodium lolii on susceptibility and host physiological response of perennial ryegrass to fungal pathogens. Eur. J. Plant Pathol. 2008, 122, 593–602. [Google Scholar] [CrossRef]
- Kannadan, S.; Rudgers, J.A. Endophyte symbiosis benefits a rare grass under low water availability. Funct. Ecol. 2008, 22, 706–713. [Google Scholar] [CrossRef]
- Song, M.L.; Li, X.Z.; Saikkonen, K.; Li, C.J.; Nan, Z.B. An asexual Epichloë endophyte enhances waterlogging tolerance of Hordeum brevisubulatum. Fungal Ecol. 2015, 13, 44–52. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Li, C.J.; Zhang, X.X.; Johnson, R.; Bao, G.S.; Yao, X.; Chai, Q. Effects of cold shocked Epichloë infected Festuca sinensis on ergot alkaloid accumulation. Fungal Ecol. 2015, 14, 99–104. [Google Scholar] [CrossRef]
- Jin, Y.Y.; SAMAN, B.; Tian, P.; Peng, Z.C.; Hou, F.J.; Chun, J.L. Research developments on the effects of grass—endophyte fungi symbiosis onsoil physical and chemical properties and microbes. Pratac. Sci. 2019, 36, 1292–1307. [Google Scholar]
- Ma, Y.L.; Li, C.J.; White, J.F. Effects of aqueous extracts of endophyte-infected grass Achnatherum inebrians on growth and development of pea aphid Acyrthosiphon pisum. Insects 2021, 12, 944. [Google Scholar] [CrossRef]
- Zhang, X.X.; Li, C.J.; Zhi, B.N. Neotyphodium endophyte increases Achnatherum inebrians (drunken horse grass) resistance to herbivores and seed predators. Weed Res. 2012, 52, 70–78. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F.; Tadych, M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Li, F. Biological and physiological characteristics of Neotyphodium gansuense symbiotic with Achnatherum inebrians. Microbiol. Res. 2008, 163, 431–440. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.Z.; Li, C.J.; Swoboda, G.A.; Young, C.A.; Sugawara, K.; Leuchtmann, A.; Schardl, C.L. Two distinct Epichloë species symbiotic with Achnatherum inebrians, drunken horse grass. Mycologia 2015, 107, 863–873. [Google Scholar] [CrossRef]
- Li, X.L.; Zheng, R.; Deng, M.H.; Wang, B.C.; Liu, R.G.; Wang, J.F. Effects of endophytic fungi on growth and nitrogen metabolism of Achnatherum inebrians under different nitrogen concentrations. Pratac. Sci. 2024, 66, 1–24. [Google Scholar]
- He, L. Effect of Inhanced UV-B Radiation on the Diversity of Arbuscular Mycorrhizal Fungi in Alpine Meadow Soil Ecosystem. Master’s Thesis, Lanzhou University, Lanzhou, China, 2009. [Google Scholar]
- Bao, G.S.; Zhang, X.X.; Li, X.Z.; Wei, X.X.; Li, C.J. Incidence and isolation of endophyte in native grasses of Qinghai Plateau. Pratac. Sci. 2015, 32, 1997–2007. [Google Scholar]
- Shabala, S. Plant Stress Physiology, 2nd ed.; Cabi: Wallingford, UK, 2017. [Google Scholar]
- Li, C.J.; Nan, Z.B.; Liu, Y.; Paul, V.H.; Peter, D. Studies on the detection of endophytes in drunken horse grass (Achnatherum inebrians). In Proceedings of the Annual Meeting of Chinese Society for Plant Pathology, Guangzhou, China, 18–20 July 2008; p. 4. [Google Scholar]
- Li, N.N.; Zhao, Y.F.; Xia, C.; Zhong, R.; Zhang, X.X. Effects of thiophanate methyl on seed borne Epichloë fungal endophyte of Achnatherum inebrians. Pratac. Sci. 2016, 33, 1306–1314. [Google Scholar]
- Lan, Y. Physiological Responses of Three Echinochloa Forages to Saline-Alkaline Stress and Comprehensive Evaluation for Their Saline-Alkaline Tolerance. Master’s Thesis, Ningxia University, Yinchuan, China, 2022. [Google Scholar]
- Huang, X.W.; Zhang, Y.J.; Feng, Q.; Chen, W.X.; Zhang, W.H.; Li, B.Y. Analysis of wheat seeding leaf proteome after UV-B radiation. J. China Agric. Univ. 2012, 17, 31–36. [Google Scholar]
- Zhao, Z.R.; Kou, M.Z.; Zhong, R.; Xia, C.; Christensen, M.J.; Zhang, X.X. Transcriptome analysis revealed plant hormone biosynthesis and response pathway modification by Epichloë gansuensis in Achnatherum inebrians under different soil moisture availability. J. Fungi 2021, 7, 640. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Han, F.; Yue, X.G.; Shi, S.B.; Wang, X.Y. Effects of long-term intensified UV-B radiation on the photosynthetic rates and antioxidative systems of three plants in alpine meadows. Acta Bot. Boreali-Occident. Sin. 2005, 67, 2010–2016. [Google Scholar]
- Bao, L.L. Comparative Analyses of the Physiological Responses of Different Rice Cultivarsto Cold Stress. Master’s Thesis, Chongqing Normal University, Chongqing, China, 2016. [Google Scholar]
- Wang, L.X.; Fang, C.; Wang, K. Physiological responses of Leymus Chinensis to long-term salt, alkali and mixed salt-alkali stresses. J. Plant Nutr. 2015, 38, 526–540. [Google Scholar] [CrossRef]
- Liu, Y. Study on the Effect of Salt Stress on the Seed Germination and Seedling Growth of Qinyin No3 Naked Oats. Master’s Thesis, Qinghai University, Xining, China, 2013. [Google Scholar]
- Shi, Q.; Simpson, W.R.; Li, Y.L.; Xu, C.T.; De, K.; Li, X.Z. Epichloë bromicola enhances Elymus dahucirus plant growth and antioxidant capacity under cadmium stress. Agronomy 2024, 14, 365. [Google Scholar] [CrossRef]
- Li, Z.Q. Physiological Responses of Cotton to Salt Stress After Adaptation Under Low Salinity in Seeding Stage. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2017. [Google Scholar]
- Wang, W.L.; Wang, Z.; Wang, J.Y. Optimization of determination method of peroxidase activity in plant. Res. Explor. Lab. 2010, 29, 21–23. [Google Scholar]
- Li, X.N.; Yang, Y.L.; Jia, L.Y.; Chen, H.J.; Wei, X. Zinc-induced oxidative damage, antioxidant enzyme response and proline metabolism in roots and leaves of wheat plants. Ecotoxicol. Environ. Saf. 2013, 89, 150–157. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants—Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Lou, Y.S.; Zhang, Z.; Wu, J. Crop growth, yield and quality as affected by ultraviolet-B (UV-B) radiation elevating. J. Agro-Environ. Sci. 2020, 39, 812–821. [Google Scholar]
- Badridze, G.; Kacharava, N.; Chkhubianishvili, E.; Rapava, L.; Kikvidze, M.; Chanishvili, S.; Chigladze, L. Influence of ultraviolet irradiation and acid precipitations on the content of antioxidants in wheat leaves. Appl. Ecol. Env. Res. 2015, 13, 993–1013. [Google Scholar] [CrossRef]
- Lou, Y.S.; Wu, L.; Ren, L.X.; Yan, M.; Zhao, S.D.; Zhu, H.W.; Zhang, Y.W. Effects of silicon application on diurnal variations of physiological properties of rice leaves of plants at the heading stage under elevated UV-B radiation. Int. J. Biometeorol. 2016, 60, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Rudnoy, S.; Majlath, I.; Pal, M.; Paldi, K.; Racz, I.; Janda, T. Interactions of S-methylmethionine and UV-B can modify the defence mechanisms induced in maize. Acta Physiol. Plant. 2015, 37, 148. [Google Scholar] [CrossRef]
- Xu, K. Studies on the Growth and Photosynthetic Responses of Two Hybid Rice Cultivars to Enhanced U-VB Radiation. Master’s Thesis, Central China Normal University, Wuhan, China, 2006. [Google Scholar]
- Xu, K.; Qiu, B.S. Responses of superhigh-yield hybrid rice Liangyoupeijiu to enhancement of ultraviolet-B radiation. Plant Sci. 2006, 172, 139–149. [Google Scholar] [CrossRef]
- Zhang, Y.F.; He, P.; Zhang, C.P. Influences of enhanced UV-B radiation and drought stress on biomass accumulation and allocation of Fagopyrum dibotrys. China J. Chin. Mater. Med. 2011, 36, 2032–2037. [Google Scholar]
- Liu, L.L.; Zhang, W.H.; Fan, Y.L.; Lu, Y.W.; Miao, X.L. Effects of different dose UV-B radiation on the morphological and physiological indices of winter wheat seedlings. Chin. J. Ecol. 2010, 29, 314–318. [Google Scholar] [CrossRef]
- Bacelar, E.; Moutinho-Pereira, J.; Ferreira, H.; Correia, C. Enhanced ultraviolet-B radiation affect growth, yield and physiological processes on triticale plants. Procedia Environ. Sci. 2015, 29, 219–220. [Google Scholar] [CrossRef]
- Decunta, F.A.; Perez, L.I.; Malinowski, D.P.; Molina-Montenegro, M.A.; Gundel, P.E. A systematic review on the effects of Epichloë fungal endophytes on drought tolerance in cool-season grasses. Front. Plant Sci. 2021, 12, 644731. [Google Scholar] [CrossRef]
- Wang, Z.F.; Li, C.J.; White, J. Effects of Epichloë endophyte infection on growth, physiological properties and seed germination of wild barley under saline conditions. J. Agron. Crop Sci. 2020, 206, 43–51. [Google Scholar] [CrossRef]
- Yang, Y. Neotyphodium Endophyte in festuca sinensis and Effect on Cold Tolerance to Host. Master’s Thesis, Lanzhou University, Lanzhou, China, 2010. [Google Scholar]
- Zhang, X.X.; Fan, X.M.; Li, C.J.; Nan, Z.B. Effects of cadmium stress on seed germination, seedling growth and antioxidative enzymes in Achnatherum inebrians plants infected with a Neotyphodium endophyte. Plant Growth Regul. 2010, 60, 91–97. [Google Scholar] [CrossRef]
- Li, Y.S.; Liu, X.B.; Henson, J.F. Advances in crop responses to enhanced UV-B radiation. Appl. Ecol. Environ. Res. 2016, 14, 339–367. [Google Scholar] [CrossRef]
- Ros, J.; Tevini, M. Interaction of UV-radiation and IAA during growth of seedlings and hypocotyl segments of sunflower. J. Plant Physiol. 1995, 146, 295–302. [Google Scholar] [CrossRef]
- Schmid, J.; Day, R.; Zhang, N.X.; Dupont, P.Y.; Cox, M.P.; Schardl, C.L.; Minards, N.; Truglio, M.; Moore, N.; Harris, D.R.; et al. Host tissue environment directs activities of an Epichloë Endophyte, While it induces systemic hormone and defense responses in its native Perennial Ryegrass host. Mol. Plant-Microbe Interact. 2017, 30, 138–149. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Gundel, P.E.; Helander, M.; Saikkonen, K. Endophytic mediation of reactive oxygen species and antioxidant activity in plants: A review. Fungal Divers. 2012, 54, 1–10. [Google Scholar] [CrossRef]
- Malencic, D.; Kiprovski, B.; Popovic, M.; Prvulovic, D.; Miladinovic, J.; Djordjevic, V. Changes in antioxidant systems in soybean as affected by Sclerotinia sclerotiorum (Lib.) de Bary. Plant Physiol. Biochem. 2010, 48, 903–908. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Wu, C.H.; Xie, B.; Liu, Y.; Cui, J.; Chen, G.; Zhang, Y.W. Model analysing the antioxidant responses of leaves and roots of switchgrass to NaCl-salinity stress. Plant Physiol. Biochem. 2012, 58, 288–296. [Google Scholar] [CrossRef]
- Yildiztugay, E.; Sekmen, A.H.; Turkan, I.; Kucukoduk, M. Elucidation of physiological and biochemical mechanisms of an endemic halophyte Centaurea tuzgoluensis under salt stress. Plant Physiol. Biochem. 2011, 49, 816–824. [Google Scholar] [CrossRef]
- Huff, A. Peroxidase-catalysed oxidation of chlorophyll by hydrogen peroxide. Phytochemistry 1982, 21, 261–265. [Google Scholar] [CrossRef]
- Catala, A. Lipid peroxidation of membrane phospholipids generates hydroxy-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem. Phys. Lipids 2009, 157, 1–11. [Google Scholar] [CrossRef]
- Delauney, A.J.; Verma, D.P.S. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Hsu, Y.T.; Kao, C.H. The effect of polyethylene glycol on proline accumulation in rice leaves. Biol. Plant. 2003, 46, 73–78. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Wang, C.; Chen, J.B.; Pang, Z.L.; Li, N.N.; Dong, H.H.; Li, D.D. The response and screening of germplasm tolerant to mixed saline-alkali stress in sweet sorghum. Crops 2016, 56–61. [Google Scholar] [CrossRef]
- He, J.M.; She, X.P.; Wang, R.B.; Liu, C. Physiological and ecological effects of enhanced UV-B radiation on wheat seedling under NaCl stress condition. Acta Bot. Boreali-Occident. Sin. 2004, 24, 1810–1815. [Google Scholar]
- Kirova, E.; Moskova, I.; Manova, V.; Koycheva, Y.; Tsekova, Z.; Borisova, D.; Nikolov, H.; Dimitrov, V.; Sergiev, I.; Kocheva, K. Exogenous cytokinin 4PU-30 modulates the response of wheat and einkorn seedlings to ultraviolet B radiation. Plants 2024, 13, 1401. [Google Scholar] [CrossRef]
- Lee, K.; Missaoui, A.; Mahmud, K.; Presley, H.; Lonnee, M. Interaction between grasses and Epichloë endophytes and its significance to biotic and abiotic stress tolerance and the rhizosphere. Microorganisms 2021, 9, 2186. [Google Scholar] [CrossRef]
- Nie, X.M.; Zhao, Z.R.; Zhang, X.X.; Bastias, D.A.; Nan, Z.B.; Li, C.J. Endophytes alleviate drought-derived oxidative damage in Achnatherum inebrians plants through increasing antioxidants and regulating host stress responses. Microb. Ecol. 2024, 87, 73. [Google Scholar] [CrossRef]
- Zhao, Z.R. The Mechanism of Response of Achnatherum inebrians-Epichloë Endophyte Symbiont to Drought Stress. Master’s Thesis, Lanzhou University, Lanzhou, China, 2023. [Google Scholar]
- Gou, X.Y. Effects of Neotyphodium Endophyte on Salt Tolerance to Drunken Horse Grass (Achnatherum inebrians). Master’s Thesis, Lanzhou University, Lanzhou, China, 2007. [Google Scholar]
- Xu, L.X.; Li, X.S.; Han, L.B.; Li, D.Y.; Song, G.L. Epichloë endophyte infection improved drought and heat tolerance of tall fescue through altered antioxidant enzyme activity. Eur. J. Hortic. Sci. 2017, 82, 90–97. [Google Scholar] [CrossRef]
- Xie, Y.; Sun, X.Y.; Feng, Q.J.; Luo, H.J.; Wassie, M.; Amee, M.; Amombo, E.; Chen, L. Comparative physiological and metabolomic analyses reveal mechanisms of Aspergillus aculeatus-mediated abiotic stress tolerance in tall fescue. Plant Physiol. Biochem. 2019, 142, 342–350. [Google Scholar] [CrossRef]
- Chandra, A.; Dubey, A. Effect of ploidy levels on the activities of Δ1-pyrroline-5-carboxylate synthetase, superoxide dismutase and peroxidase in Cenchrus species grown under water stress. Plant Physiol. Biochem. 2010, 48, 27–34. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Srivastava, S.; Penna, S.; D’Souza, S.F. Thiourea orchestrates regulation of redox state and antioxidant responses to reduce the NaCl-induced oxidative damage in Indian mustard (Brassica juncea (L.) Czern.). Plant Physiol. Biochem. 2011, 49, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Kumari, G.J.; Reddy, A.M.; Naik, S.T.; Kumar, S.G.; Prasanthi, J.; Sriranganayakulu, G.; Reddy, P.C.; Sudhakar, C. Jasmonic acid induced changes in protein pattern, antioxidative enzyme activities and peroxidase isozymes in peanut seedlings. Biol. Plant. 2006, 50, 219–226. [Google Scholar] [CrossRef]
- Singh, R.; Singh, S.; Tripathi, R.; Agrawal, S.B. Supplemental UV-B radiation induced changes in growth, pigments and antioxidant pool of bean (Dolichos lablab) under field conditions. J. Environ. Biol. 2011, 32, 139–145. [Google Scholar] [PubMed]
- Yang, S.H.; Wang, L.J.; Li, S.H.; Duan, W.; Loescher, W.; Liang, Z.C. The effects of UV-B radiation on photosynthesis in relation to Photosystem II photochemistry, thermal dissipation and antioxidant defenses in winter wheat (Triticum aestivum L.) seedlings at different growth temperatures. Funct. Plant Biol. 2007, 34, 907–917. [Google Scholar] [CrossRef]
- Amirikhah, R.; Etemadi, N.; Sabzalian, M.R.; Nikbakht, A.; Eskandari, A. Gamma radiation negatively impacted seed germination, seedling growth and antioxidant enzymes activities in tall fescue infected with Epichloë endophyte. Ecotoxicol. Environ. Saf. 2021, 216, 112169. [Google Scholar] [CrossRef]
- Barabás, K.N.; Szegletes, Z.; Pestenácz, A.; Fülöp, K.; Erdei, L. Effects of excess UV-B irradiation on the antioxidant defence mechanisms in wheat (Triticum aestivum L.) seedlings. J. Plant Physiol. 1998, 153, 146–153. [Google Scholar] [CrossRef]
- Chen, Z.J.; Ma, Y.; Weng, Y.; Yang, R.Q.; Gu, Z.X.; Wang, P. Effects of UV-B radiation on phenolic accumulation, antioxidant activity and physiological changes in wheat (Triticum aestivum L.) seedlings. Food Biosci. 2019, 30, 100409. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Bauerle, T.L. A new currency for mutualism? Fungal endophytes alter antioxidant activity in hosts responding to drought. Fungal Divers. 2012, 54, 39–49. [Google Scholar] [CrossRef]
- Chen, Y.Q. The Saline-Alkali Tolerance of Seed of Epichloë Endophytic Fungi-Achnatherum inebrians Symbiont and Effects of Nitrogen Applicationon Seed Yield. Master’s Thesis, Lanzhou University, Lanzhou, China, 2022. [Google Scholar]
- Zhang, X.X. Response of Achnatherum inebrians/Neotyphodium gansuense Symbiont to Stresses and Secondary Metabolites Activities. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2012. [Google Scholar]
- Oberhofer, M.; Güsewell, S.; Leuchtmann, A. Effects of natural hybrid and non-hybrid Epichloë endophytes on the response of Hordelymus europaeus to drought stress. New Phytol. 2014, 201, 242–253. [Google Scholar] [CrossRef]
- Soleimani, M.; Hajabbasi, M.A.; Afyuni, M.; Mirlohi, A.; Borggaard, O.K.; Holm, P.E. Effect of endophytic fungi on cadmium tolerance and bioaccumulation by Festuca Arundinacea and Festuca Pratensis. Int. J. Phytorem. 2010, 12, 535–549. [Google Scholar] [CrossRef]
- Hideg, É.; Jansen, M.A.K.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Suesslin, C.; Frohnmeyer, H. An Arabidopsis mutant defective in UV-B light-mediated responses. Plant J. 2003, 33, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Binder, B.Y.K.; Peebles, C.A.M.; Shanks, J.V.; San, K.Y. The effects of UV-B Stress on the production of terpenoid Indole alkaloids in Catharanthus roseus hairy roots. Biotechnol. Prog. 2009, 25, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.W.; Zhao, X.Q.; Zhang, X.L.; Wang, X.G.; Geng, Y.Y.; Hu, L.Y.; Zhao, N.; Mao, S.J.; Liu, H.J.; Kang, S.P.; et al. Sustainable development of ecological grass-based livestock husbandry in Qinghai-Tibet Plateau alpine area: Principle, technology and practice. Acta Ecol. Sin. 2020, 40, 6324–6337. [Google Scholar]
- Wu, G.L.; Du, G.Z. Germination is related to seed mass in grasses (Poaceae) of the eastern Qinghai-Tibetan Plateau, China. Nord. J. Bot. 2007, 25, 361–365. [Google Scholar] [CrossRef]
- Zhou, Q.P.; Ji, Y.J.; Bruijn, K.D.; Liang, G.L.; Yan, H.B. Preliminary evaluation of native grasses collected from alpine rangelands in Qinghai Province, China, as materials for breeding grazing-tolerant fine herbage. Grassl. Sci. 2009, 55, 41–45. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, C.; Li, X.; Shi, Q. Epichloë gansuensis Enhances Achnatherum inebrians Seedlings Growth and Antioxidant Capacity Under UV-B Stress. Agronomy 2025, 15, 1546. https://doi.org/10.3390/agronomy15071546
Wan C, Li X, Shi Q. Epichloë gansuensis Enhances Achnatherum inebrians Seedlings Growth and Antioxidant Capacity Under UV-B Stress. Agronomy. 2025; 15(7):1546. https://doi.org/10.3390/agronomy15071546
Chicago/Turabian StyleWan, Cuiling, Xiuzhang Li, and Qian Shi. 2025. "Epichloë gansuensis Enhances Achnatherum inebrians Seedlings Growth and Antioxidant Capacity Under UV-B Stress" Agronomy 15, no. 7: 1546. https://doi.org/10.3390/agronomy15071546
APA StyleWan, C., Li, X., & Shi, Q. (2025). Epichloë gansuensis Enhances Achnatherum inebrians Seedlings Growth and Antioxidant Capacity Under UV-B Stress. Agronomy, 15(7), 1546. https://doi.org/10.3390/agronomy15071546