
Effects of Exogenous Selenium Species and Arbuscular Mycorrhizal Fungi on Selenium Uptake in Soybean

 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Pot Experiments
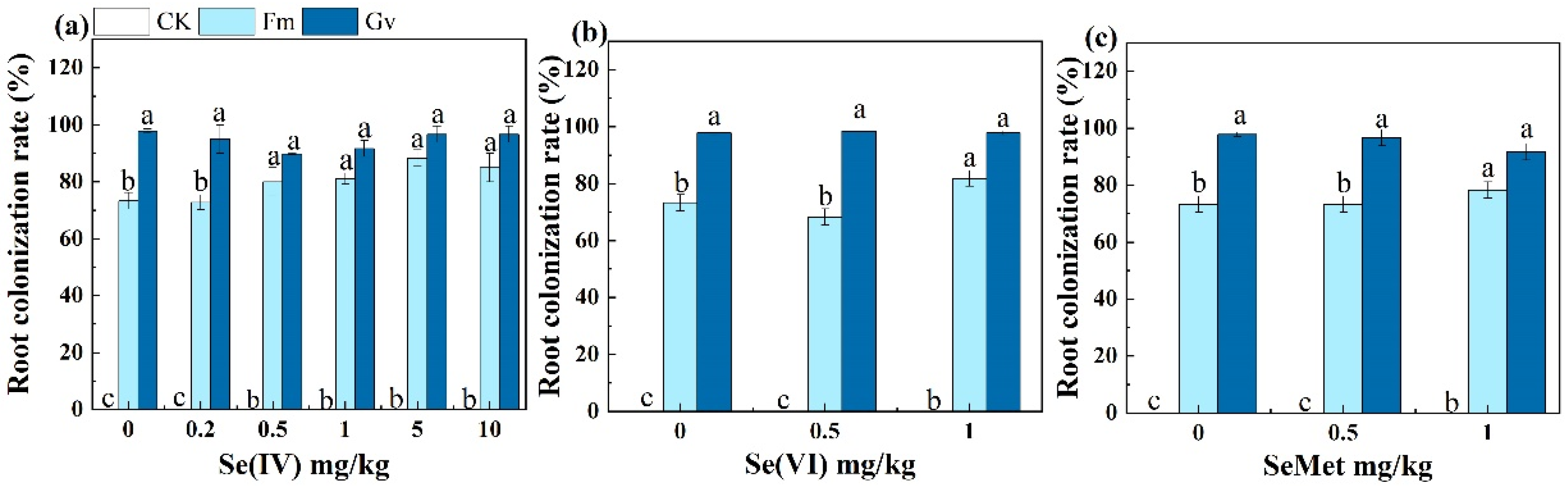
2.2. Determination of Mycorrhizal Colonization Rate in Roots of Soybean
2.3. Chemical Analysis
2.3.1. Determination of P Concentration in Soybean
2.3.2. Determination of Se Concentration in Soil and Plant
2.3.3. Analysis of Se Fractions in Soil
2.4. Quality Control
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effects of AMF on P Uptake and Growth of Soybean Grown in Soils Amended with Different Se Species
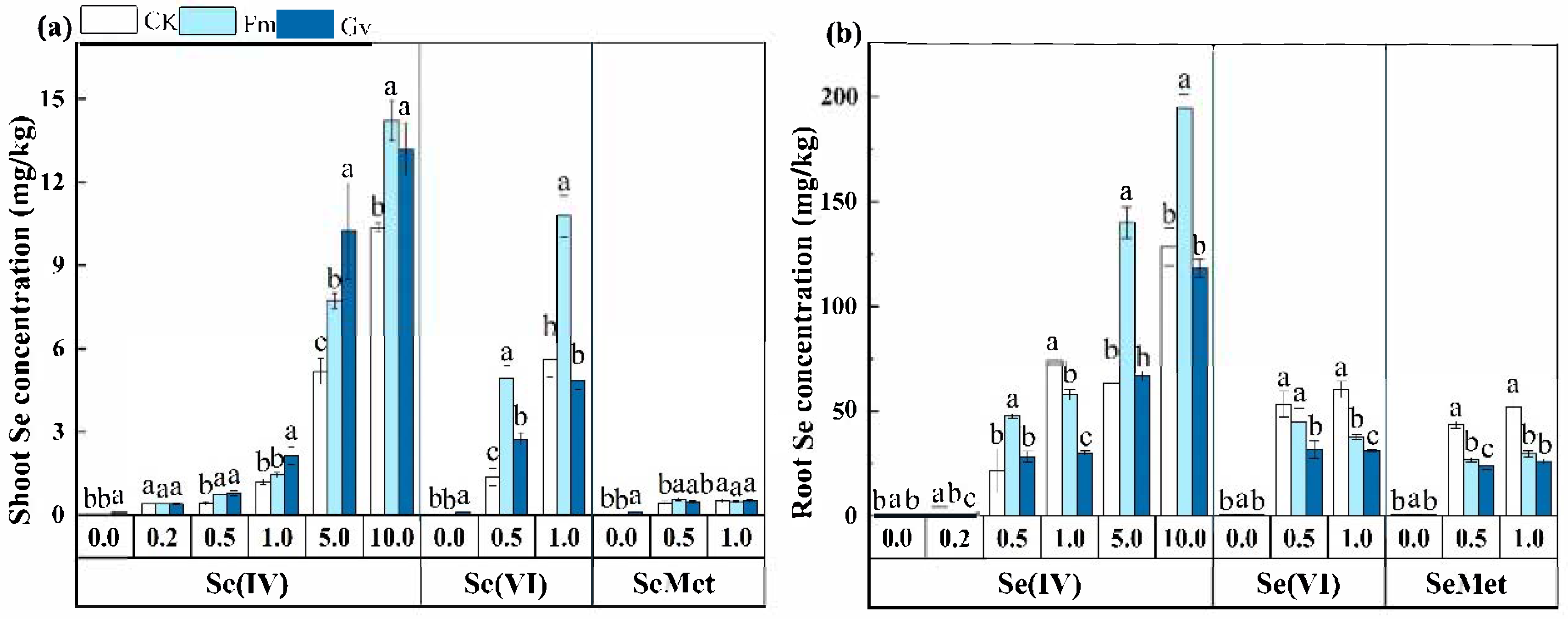
3.2. Effect of AMF on Se Uptake in Soybean Grown in Soils Amend with Different Se Species
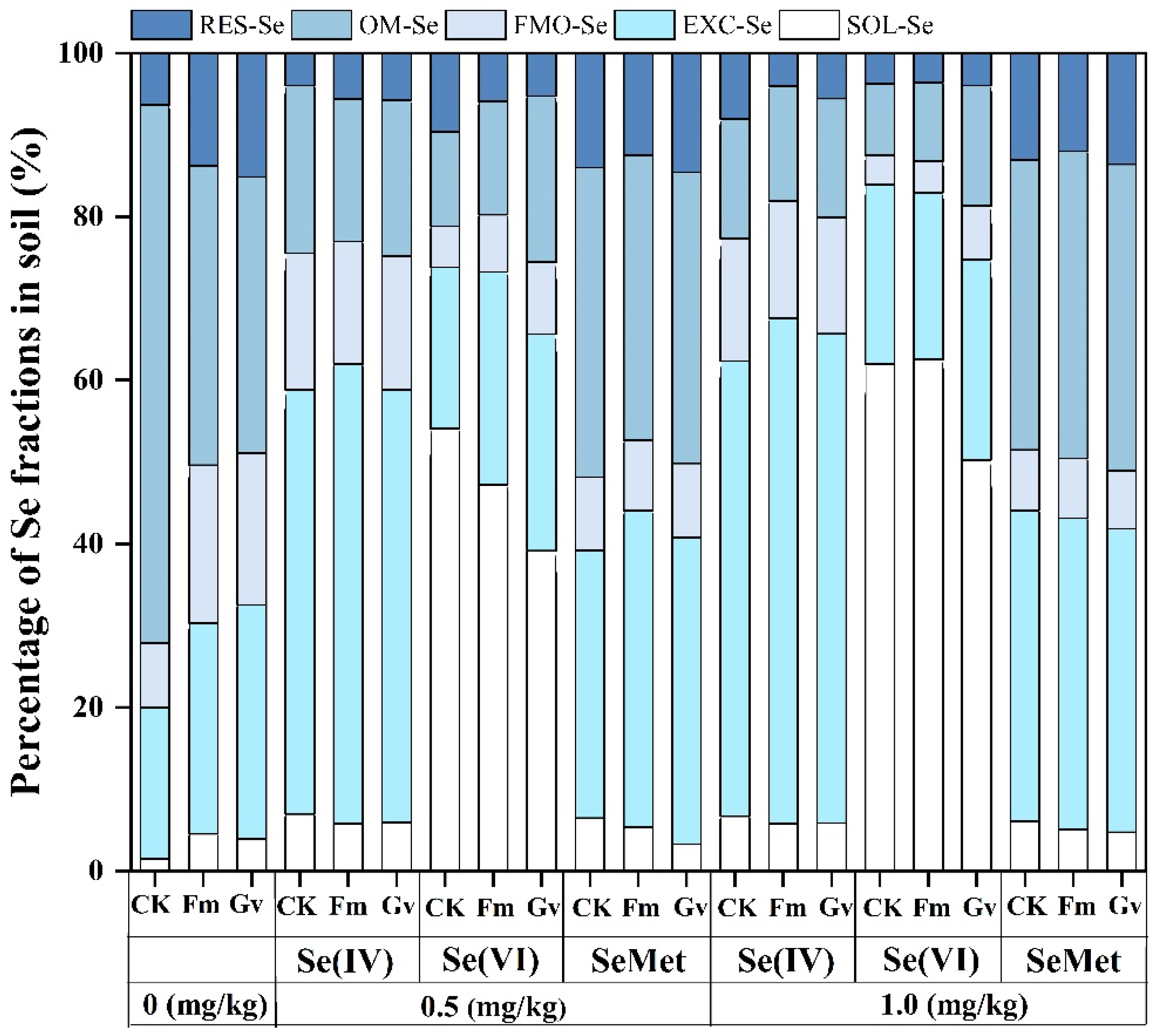
3.3. Effects of Different Se Species and AMF on Se Fractions in Rhizosphere Soil of Soybean
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schomburg, L. Dietary selenium and human health. Nutrients 2017, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Zhang, M.; Tang, S.; Li, M.; Wu, R.; Wan, S.; Chen, L.; Wei, X.; Feng, S. Effects and Impact of Selenium on Human Health, a review. Molecules 2024, 30, 50. [Google Scholar] [CrossRef] [PubMed]
- Altekin, E.; Çoker, C.; Şişman, A.R.; Önvural, B.; Kuralay, F.; Kırımlı, Ö. The relationship between trace elements and cardiac markers in acute coronary syndromes. J. Trace Elem. Med. Biol. 2005, 18, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Tan, L.C.; Nancharaiah, Y.V.; van Hullebusch, E.D.; Lens, P.N. Selenium: Environmental significance, pollution, and biological treatment technologies. In Anaerobic Treatment of Mine Wastewater for the Removal of Selenate and Its Co-Contaminants; CRC Press: Boca Raton, FL, USA, 2018; pp. 9–71. [Google Scholar]
- Dinh, Q.T.; Cui, Z.; Huang, J.; Tran, T.A.T.; Wang, D.; Yang, W.; Zhou, F.; Wang, M.; Yu, D.; Liang, D. Selenium distribution in the Chinese environment and its relationship with human health: A review. Environ. Int. 2018, 112, 294–309. [Google Scholar] [CrossRef]
- Wu, Z.; Bañuelos, G.S.; Lin, Z.-Q.; Liu, Y.; Yuan, L.; Yin, X.; Li, M. Biofortification and phytoremediation of selenium in China. Front. Plant Sci. 2015, 6, 136. [Google Scholar] [CrossRef]
- Bodnar, M.; Marzena, S.; Piotr, K.; Namiesnik, J. Methods of selenium supplementation: Bioavailability and determination of selenium compounds. Crit. Rev. Food Sci. Nutr. 2016, 56, 36–55. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Agriculture: The real nexus for enhancing bioavailable micronutrients in food crops. J. Trace Elem. Med. Biol. 2005, 18, 299–307. [Google Scholar] [CrossRef]
- Wei, H. Comparison of the bioavailability of soybean selenoprotein and sodium selenite. Soybean Sci. 2010, 29, 534–536. [Google Scholar]
- Dai, H.; Wei, S.; Twardowska, I. Biofortification of soybean (Glycine max L.) with Se and Zn, and enhancing its physiological functions by spiking these elements to soil during flowering phase. Sci. Total Environ. 2020, 740, 139648. [Google Scholar] [CrossRef]
- Ros, G.; Van Rotterdam, A.; Bussink, D.; Bindraban, P.S. Selenium fertilization strategies for bio-fortification of food: An agro-ecosystem approach. Plant Soil 2016, 404, 99–112. [Google Scholar] [CrossRef]
- Deng, X.; Liu, K.; Li, M.; Zhang, W.; Zhao, X.; Zhao, Z.; Liu, X. Difference of selenium uptake and distribution in the plant and selenium form in the grains of rice with foliar spray of selenite or selenate at different stages. Field Crops Res. 2017, 211, 165–171. [Google Scholar] [CrossRef]
- Terry, N.; Zayed, A.; De Souza, M.; Tarun, A. Selenium in higher plants. Annu. Rev. Plant Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Ikram, S.; Li, Y.; Lin, C.; Yi, D.; Heng, W.; Li, Q.; Tao, L.; Hongjun, Y.; Weijie, J. Selenium in plants: A nexus of growth, antioxidants, and phytohormones. J. Plant Physiol. 2024, 296, 154237. [Google Scholar] [CrossRef]
- Poblaciones, M.J.; Rodrigo, S.; Santamaría, O.; Chen, Y.; McGrath, S. Agronomic selenium biofortification in Triticum durum under Mediterranean conditions: From grain to cooked pasta. Food Chem. 2014, 146, 378–384. [Google Scholar] [CrossRef]
- Dai, Z.; Imtiaz, M.; Rizwan, M.; Yuan, Y.; Huang, H.; Tu, S. Dynamics of selenium uptake, speciation, and antioxidant response in rice at different panicle initiation stages. Sci. Total Environ. 2019, 691, 827–834. [Google Scholar] [CrossRef]
- Qi, M.; Wang, D.; Zhai, H.; Zhou, F.; Wu, H.; Zhao, W.; Ren, R.; Shi, J.; Liang, D. Effects of straw amendment on the bioavailability of selenite in soil and its mechanisms. Ecotoxicol. Environ. Saf. 2025, 290, 117578. [Google Scholar] [CrossRef]
- Haug, A.; Graham, R.D.; Christophersen, O.A.; Lyons, G.H. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228. [Google Scholar]
- Zhang, F.; Liu, M.; Li, Y.; Che, Y.; Xiao, Y. Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Sci. Total Environ. 2019, 655, 1150–1158. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, L.; Meng, S.; Song, X.; Long, R.; Huang, H.; Tang, Y.; Zhou, X.; Li, M.; Sun, B.; et al. The physiological responses of celery (Apium graveolens L.) and its ability to accumulate selenium when inoculated with Funneliformis mosseae. Sci. Hortic. 2024, 326, 112752. [Google Scholar] [CrossRef]
- Wang, J.; Pi, Y.; Li, Y.; Wang, H.; Huang, K.; Wang, X.; Xia, H.; Zhang, X.; Liang, D.; Lv, X.; et al. Transcriptome and metabolome analyses reveal the promoting effects of arbuscular mycorrhizal fungi on selenium uptake in grapevines. Plant Physiol. Biochem. 2025, 219, 109456. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Sun, J.; Liu, D.; Li, B.; Zhang, A.; Li, Z.; Tong, Y. Characterization of the promoter of phosphate transporter TaPHT1. 2 differentially expressed in wheat varieties. J. Genet. Genom. 2009, 36, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Pochon, N.; Ayadi, A.; Nakanishi, T.M.; Thibaud, M.-C. Phosphate import in plants: Focus on the PHT1 transporters. Front. Plant Sci. 2011, 2, 83. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, B.; Zheng, S.; Zhang, X.; Wang, X.; Dong, W.; Xie, Q.; Wang, G.; Xiao, Y.; Chen, F. A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell 2021, 184, 5527–5540. [Google Scholar] [CrossRef]
- Klugh-Stewart, K.; Cumming, J.R. Organic acid exudation by mycorrhizal Andropogon virginicus L. (broomsedge) roots in response to aluminum. Soil Biol. Biochem. 2009, 41, 367–373. [Google Scholar] [CrossRef]
- Lindblom, S.D.; Valdez-Barillas, J.R.; Fakra, S.C.; Marcus, M.A.; Wangeline, A.L.; Pilon-Smits, E.A.H. Influence of microbial associations on selenium localization and speciation in roots of Astragalus and Stanleya hyperaccumulators. Environ. Exp. Bot. 2013, 88, 33–42. [Google Scholar] [CrossRef]
- Golubkina, N.; Amagova, Z.; Matsadze, V.; Zamana, S.; Tallarita, A.; Caruso, G. Effects of arbuscular mycorrhizal fungi on yield, biochemical characteristics, and elemental composition of garlic and onion under selenium supply. Plants 2020, 9, 84. [Google Scholar] [CrossRef]
- Li, J.; Awasthi, M.K.; Xing, W.; Liu, R.; Bao, H.; Wang, X.; Wang, J.; Wu, F. Arbuscular mycorrhizal fungi increase the bioavailability and wheat (Triticum aestivum L.) uptake of selenium in soil. Ind. Crops Prod. 2020, 150, 112383. [Google Scholar] [CrossRef]
- Zhou, F.; Yang, W.; Wang, M.; Miao, Y.; Cui, Z.; Li, Z.; Liang, D. Effects of selenium application on Se content and speciation in Lentinula edodes. Food Chem. 2018, 265, 182–188. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Z.; Gu, M.; Li, H.; Shohag, M.J.I.; Shen, F.; Wang, X.; Wei, Y. Combined use of arbuscular mycorrhizal fungus and selenium fertilizer shapes microbial community structure and enhances organic selenium accumulation in rice grain. Sci. Total Environ. 2020, 748, 141166. [Google Scholar] [CrossRef]
- Bao, S. Analytical methods of soil agrochemistry. In Soil Science Society of China Beijing, 3rd ed.; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Luo, W.; Li, J.; Ma, X.; Niu, H.; Hou, S.; Wu, F. Effect of arbuscular mycorrhizal fungi on uptake of selenate, selenite, and selenomethionine by roots of winter wheat. Plant Soil 2019, 438, 71–83. [Google Scholar] [CrossRef]
- Wu, F.; Ye, Z.; Wong, M.H. Intraspecific differences of arbuscular mycorrhizal fungi in their impacts on arsenic accumulation by Pteris vittata L. Chemosphere 2009, 76, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ali, F.; Wang, M.; Dinh, Q.T.; Zhou, F.; Bañuelos, G.S.; Liang, D. Understanding boosting selenium accumulation in Wheat (Triticum aestivum L.) following foliar selenium application at different stages, forms, and doses. Environ. Sci. Pollut. Res. 2020, 27, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cui, Z.; Xue, M.; Peng, Q.; Zhou, F.; Wang, D.; Dinh, Q.T.; Liu, Y.; Liang, D. Assessing the uptake of selenium from naturally enriched soils by maize (Zea mays L.) using diffusive gradients in thin-films technique (DGT) and traditional extractions. Sci. Total Environ. 2019, 689, 1–9. [Google Scholar] [CrossRef]
- Öpik, M.; Zobel, M.; Cantero, J.J.; Davison, J.; Facelli, J.M.; Hiiesalu, I.; Jairus, T.; Kalwij, J.M.; Koorem, K.; Leal, M.E.; et al. Global sampling of plant roots expands the described molecular diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2013, 23, 411–430. [Google Scholar] [CrossRef]
- Yang, Q.; Ravnskov, S.; Neumann Andersen, M. Nutrient uptake and growth of potato: Arbuscular mycorrhiza symbiosis interacts with quality and quantity of amended biochars. J. Plant Nutr. Soil Sci. 2020, 183, 220–232. [Google Scholar] [CrossRef]
- Tran, B.T.T.; Watts-Williams, S.J.; Cavagnaro, T.R. Impact of an arbuscular mycorrhizal fungus on the growth and nutrition of fifteen crop and pasture plant species. Funct. Plant Biol. 2019, 46, 732–742. [Google Scholar] [CrossRef]
- Chandrasekaran, M. Arbuscular mycorrhizal fungi mediated enhanced biomass, root morphological traits and nutrient uptake under drought stress: A meta-analysis. J. Fungi 2022, 8, 660. [Google Scholar] [CrossRef]
- Hammer, E.C.; Balogh-Brunstad, Z.; Jakobsen, I.; Olsson, P.A.; Stipp, S.L.; Rillig, M.C. A mycorrhizal fungus grows on biochar and captures phosphorus from its surfaces. Soil Biol. Biochem. 2014, 77, 252–260. [Google Scholar] [CrossRef]
- Li, S.; Chi, S.; Lin, C.; Cai, C.; Yang, L.; Peng, K.; Huang, X.; Liu, J. Combination of biochar and AMF promotes phosphorus utilization by stimulating rhizosphere microbial co-occurrence networks and lipid metabolites of Phragmites. Sci. Total Environ. 2022, 845, 157339. [Google Scholar] [CrossRef]
- Paszkowski, U.; Kroken, S.; Roux, C.; Briggs, S.P. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 13324–13329. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Yu, T.; Qi, H.; Li, F.; Yang, Z. Analysis of phosphorus and sulfur effect on soil selenium bioavailability based on diffusive gradients in thin films technique and sequential extraction. Chemosphere 2022, 302, 134831. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Wu, M.; Zhang, Z.; Su, R.; He, H.; Zhang, X. The interaction of arbuscular mycorrhizal fungi and phosphorus inputs on selenium uptake by alfalfa (Medicago sativa L.) and selenium fraction transformation in soil. Front. Plant Sci. 2020, 11, 966. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, N.; Liang, X.; Zheng, L.; Zhang, C.; Li, Y.-F.; Zhang, Z.; Gao, Y.; Zhao, J. A comparative study on the accumulation, translocation and transformation of selenite, selenate, and SeNPs in a hydroponic-plant system. Ecotoxicol. Environ. Saf. 2020, 189, 109955. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets–iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Radawiec, A.; Rutkowska, B.; Tidaback, J.A.; Gozdowski, D.; Knapowski, T.; Szulc, W. The impact of selenium fertilization on the quality characteristics of spring wheat grain. Agronomy 2021, 11, 2100. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef]
- Deng, X.; Liao, J.; Zhao, Z.; Qin, Y.; Liu, X. Distribution and speciation of selenium in soybean proteins and its effect on protein structure and functionality. Food Chem. 2022, 370, 130982. [Google Scholar] [CrossRef]
- Peng, Q.; Guo, L.; Ali, F.; Li, J.; Qin, S.; Feng, P.; Liang, D. Influence of pak choi plant cultivation on Se distribution, speciation and bioavailability in soil. Plant Soil 2016, 403, 331–342. [Google Scholar] [CrossRef]
- Lyu, C.; Chen, J.; Li, L.; Zhao, Z.; Liu, X. Characteristics of Se in water-soil-plant system and threshold of soil Se in seleniferous areas in Enshi, China. Sci. Total Environ. 2022, 827, 154372. [Google Scholar] [CrossRef]
- Li, Z.; Liang, D.; Peng, Q.; Cui, Z.; Huang, J.; Lin, Z. Interaction between selenium and soil organic matter and its impact on soil selenium bioavailability: A review. Geoderma 2017, 295, 69–79. [Google Scholar] [CrossRef]
- Guo, H.; Chen, J.; Zhong, B.; Liu, C.; Wu, J.; He, L.; Ye, Z.; Liu, D. Heavy metal concentration, enzyme activity, and physical and chemical properties of rhizosphere and non-rhizosphere soils containing Moso bamboo. Acta Ecol. Sin. 2017, 27, 6149–6156. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Se Species | Soil Se Level (mg/kg) | Shoot (g/pot, DW) | Root (g/pot, DW) | ||||
|---|---|---|---|---|---|---|---|
| CK | Fm | Gv | CK | Fm | Gv | ||
| Se(Ⅳ) | 0 | 6.62 ± 0.01 Aa | 9.03 ± 0.01 ABa | 7.16 ± 0.02 Ca | 0.34 ± 0.03 Aab | 0.66 ± 0.09 ABa | 0.33 ± 0.13 Bb |
| 0.2 | 6.22 ± 0.09 Ab | 9.44 ± 0.05 ABa | 7.39 ± 0.03 BCab | 0.38 ± 0.04 Aa | 0.67 ± 0.14 ABa | 0.71 ± 0.19 ABa | |
| 0.5 | 6.63 ± 0.06 Aa | 10.5 ± 0.13 ABa | 9.15 ± 0.08 Aa | 0.37 ± 0.03 Ab | 0.73 ± 0.13 ABa | 0.46 ± 0.07 Bab | |
| 1.0 | 5.90 ± 0.10 Ab | 11.5 ± 0.10 Aa | 9.46 ± 0.31 Aab | 0.38 ± 0.06 Ab | 0.78 ± 0.15 Aa | 0.66 ± 0.05 ABa | |
| 5.0 | 5.42 ± 0.47 Ab | 12.4 ± 0.27 Aa | 7.84 ± 1.74 ABCb | 0.40 ± 0.09 Ac | 1.00 ± 0.06 Aa | 0.74 ± 0.00 ABb | |
| 10.0 | 6.85 ± 0.73 Aab | 5.79 ± 0.73 Bb | 8.99 ± 0.94 ABa | 0.37 ± 0.02 Ab | 0.39 ± 0.14 Bb | 0.91 ± 0.10 Aa | |
| Se(Ⅵ) | 0 | 6.62 ± 0.01 Aa | 9.03 ± 0.01 Aa | 7.16 ± 0.02 Ba | 0.34 ± 0.03 Aab | 0.66 ± 0.09 Aa | 0.33 ± 0.13 Ab |
| 0.5 | 6.38 ± 0.32 Ab | 7.57 ± 0.46 Ab | 10.2 ± 0.23 Aa | 0.39 ± 0.10 Aa | 0.50 ± 0.20 Aa | 0.57 ± 0.24 Aa | |
| 1.0 | 7.29 ± 0.62 Aab | 6.25 ± 0.75 Ab | 7.89 ± 0.32 Ba | 0.40 ± 0.05 Aa | 0.47 ± 0.04 Aa | 0.45 ± 0.11 Aa | |
| SeMet | 0 | 6.62 ± 0.01 Aa | 9.03 ± 0.01 Aa | 7.16 ± 0.02 Ba | 0.34 ± 0.03 Aab | 0.66 ± 0.09 Aa | 0.33 ± 0.13 Ab |
| 0.5 | 5.95 ± 0.07 Ab | 7.75 ± 0.06 Aab | 10.2 ± 0.04 Aa | 0.46 ± 0.04 Aa | 0.55 ± 0.18 Aa | 0.66 ± 0.14 Aa | |
| 1.0 | 6.68 ± 0.07 Ab | 8.03 ± 0.03 Ab | 11.0 ± 0.06 Aa | 0.37 ± 0.08 Ab | 0.49 ± 0.10 Aab | 0.58 ± 0.19 Aa | |
| Se Species | Soil Se Level (mg/kg) | P Concentration in Shoot (g/kg) | P Concentration in Root (g/kg) | ||||
|---|---|---|---|---|---|---|---|
| CK | Fm | Gv | CK | Fm | Gv | ||
| Se(Ⅳ) | 0 | 1.09 ± 0.07 Ac | 1.95 ± 0.03 Eb | 2.39 ± 0.03 Aa | 1.48 ± 0.30 Ab | 1.77 ± 0.37 Bb | 4.68 ± 0.26 Aa |
| 0.2 | 1.12 ± 0.05 Ac | 2.48 ± 0.02 Aa | 1.97 ± 0.04 Bb | 1.25 ± 0.25 ABc | 1.74 ± 0.05 Bb | 2.73 ± 0.08 Ca | |
| 0.5 | 1.07 ± 0.03 Ac | 2.16 ± 0.07 Ca | 1.67 ± 0.01 Cb | 1.01 ± 0.14 Cc | 2.66 ± 0.13 Ab | 3.22 ± 0.15 Ba | |
| 1.0 | 1.07 ± 0.05 Ab | 2.07 ± 0.00 Da | 2.08 ± 0.05 Ba | 1.35 ± 0.18 ABc | 2.64 ± 0.13 Ab | 3.11 ± 0.24 Ba | |
| 5.0 | 1.08 ± 0.01 Ac | 2.34 ± 0.02 Ba | 1.93 ± 0.08 Bb | 1.57 ± 0.29 Ab | 2.56 ± 0.36 Aa | 2.27 ± 0.09 Da | |
| 10.0 | 1.12 ± 0.04 Ab | 1.63 ± 0.05 Fa | 1.52 ± 0.26 Ca | 1.25 ± 0.20 ABb | 2.70 ± 0.84 Aa | 1.95 ± 0.04 Eb | |
| Se(Ⅵ) | 0 | 1.09 ± 0.07 Ac | 1.95 ± 0.03 Cb | 2.39 ± 0.03 Aa | 1.48 ± 0.30 Ab | 1.77 ± 0.37 Ab | 4.68 ± 0.26 Aa |
| 0.5 | 1.07 ± 0.05 Ab | 2.25 ± 0.03 Ba | 2.08 ± 0.27 ABa | 1.20 ± 0.04 Ab | 2.59 ± 0.44 Ba | 2.62 ± 0.47 Ca | |
| 1.0 | 1.14 ± 0.02 Ac | 2.46 ± 0.15 Aa | 1.96 ± 0.12 Bb | 1.12 ± 0.30 Ab | 3.42 ± 0.29 Ca | 3.50 ± 0.32 Ba | |
| SeMet | 0 | 1.09 ± 0.07 Ac | 1.95 ± 0.03 Bb | 2.39 ± 0.03 Aa | 1.48 ± 0.30 Ab | 1.77 ± 0.37 Bb | 4.68 ± 0.26 Aa |
| 0.5 | 0.93 ± 0.04 Bc | 2.19 ± 0.07 ABa | 1.85 ± 0.16 Cb | 1.12 ± 0.13 Ab | 2.56 ± 0.33 Aa | 2.51 ± 0.57 Ba | |
| 1.0 | 1.14 ± 0.07 Ab | 2.42 ± 0.20 Aa | 2.23 ± 0.07 Ba | 1.35 ± 0.33 Ab | 2.68 ± 0.31 Aa | 2.75 ± 0.08 Ba | |
| Treatment | Se (mg/kg) | BCFshoot/soil | BCFroot/soil | TF |
|---|---|---|---|---|
| CK | 0 | 0.04 c | 0.04 b | 0.020 b |
| Fm | 0.21 b | 6.20 a | 0.036 b | |
| Gv | 0.85 a | 3.65 b | 0.233 a | |
| Se(Ⅳ)-CK | 0.5 | 0.69 b | 35.95 a | 0.023 a |
| Se(Ⅳ)-Fm | 0.98 b | 64.68 a | 0.015 a | |
| Se(Ⅳ)-Gv | 1.44 a | 51.80 a | 0.028 a | |
| Se(Ⅵ)-CK | 2.24 c | 86.94 a | 0.026 b | |
| Se(Ⅵ)-Fm | 9.63 a | 87.55 a | 0.113 a | |
| Se(Ⅵ)-Gv | 5.67 b | 66.02 a | 0.087 a | |
| SeMet-CK | 0.92 a | 93.84 a | 0.010 b | |
| SeMet-Fm | 1.14 a | 52.91 b | 0.021 a | |
| SeMet-Gv | 0.89 a | 45.01 b | 0.020 a | |
| Se(Ⅳ)-CK | 1.0 | 1.12 b | 68.66 a | 0.016 b |
| Se(Ⅳ)-Fm | 1.28 ab | 51.61 b | 0.025 b | |
| Se(Ⅳ)-Gv | 1.78 a | 24.97 c | 0.072 a | |
| Se(Ⅵ)-CK | 8.59 a | 92.76 a | 0.093 c | |
| Se(Ⅵ)-Fm | 12.33 a | 43.24 b | 0.285 a | |
| Se(Ⅵ)-Gv | 6.98 a | 44.51 b | 0.157 b | |
| SeMet-CK | 0.66 a | 66.89 a | 0.010 b | |
| SeMet-Fm | 0.54 a | 32.72 b | 0.016 a | |
| SeMet-Gv | 0.63 a | 32.72 b | 0.019 a | |
| ANOVA (F Value) | ||||
| Fm | ns | ns | ** | |
| Gv | ns | ** | ** | |
| Se species (Se) | *** | ** | *** | |
| Fm × Se | ns | * | *** | |
| Gv × Se | ns | ns | ns | |
| Standardization Factor | R2 | p |
|---|---|---|
| Y1 (Shoot Se concentration) = 0.877X1 | 0.766 | <0.001 |
| Y2 (Root Se concentration) = 0.516X2 | 0.254 | <0.001 |
| Y2 (Root Se concentration) = 0.563X2 + 0.432X1 | 0.432 | <0.001 |
| Y2 (Root Se concentration) = 0.409X2 + 0.501X1 + 0.366X5 | 0.531 | <0.001 |
| Y2 (Root Se concentration) = 0.421X2 + 0.434X1 + 0.569X5 − 0.322X4 | 0.561 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Xing, W.; Chen, Q.; Ma, Y.; Yang, J.; Hao, S.; Wu, F. Effects of Exogenous Selenium Species and Arbuscular Mycorrhizal Fungi on Selenium Uptake in Soybean. Agronomy 2025, 15, 1498. https://doi.org/10.3390/agronomy15071498
Zhu H, Xing W, Chen Q, Ma Y, Yang J, Hao S, Wu F. Effects of Exogenous Selenium Species and Arbuscular Mycorrhizal Fungi on Selenium Uptake in Soybean. Agronomy. 2025; 15(7):1498. https://doi.org/10.3390/agronomy15071498
Chicago/Turabian StyleZhu, Haode, Wenjing Xing, Qimin Chen, Yuanzhe Ma, Jing Yang, Shangyan Hao, and Fuyong Wu. 2025. "Effects of Exogenous Selenium Species and Arbuscular Mycorrhizal Fungi on Selenium Uptake in Soybean" Agronomy 15, no. 7: 1498. https://doi.org/10.3390/agronomy15071498
APA StyleZhu, H., Xing, W., Chen, Q., Ma, Y., Yang, J., Hao, S., & Wu, F. (2025). Effects of Exogenous Selenium Species and Arbuscular Mycorrhizal Fungi on Selenium Uptake in Soybean. Agronomy, 15(7), 1498. https://doi.org/10.3390/agronomy15071498