
Integrated Analysis of Hormone Metabolomics and Transcriptomics Reveals the Differential Mechanism of Ginger (Zingiber officinale Roscoe) Rhizome Bulking




 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotypic Identification
2.3. Metabolome Analysis of Ginger Rhizome Hormones
2.4. Transcriptome Analysis and qRT-PCR Validation
2.5. Statistical Analysis
3. Results
3.1. Analysis of Growth Parameters and Ploidy Determination in Ginger
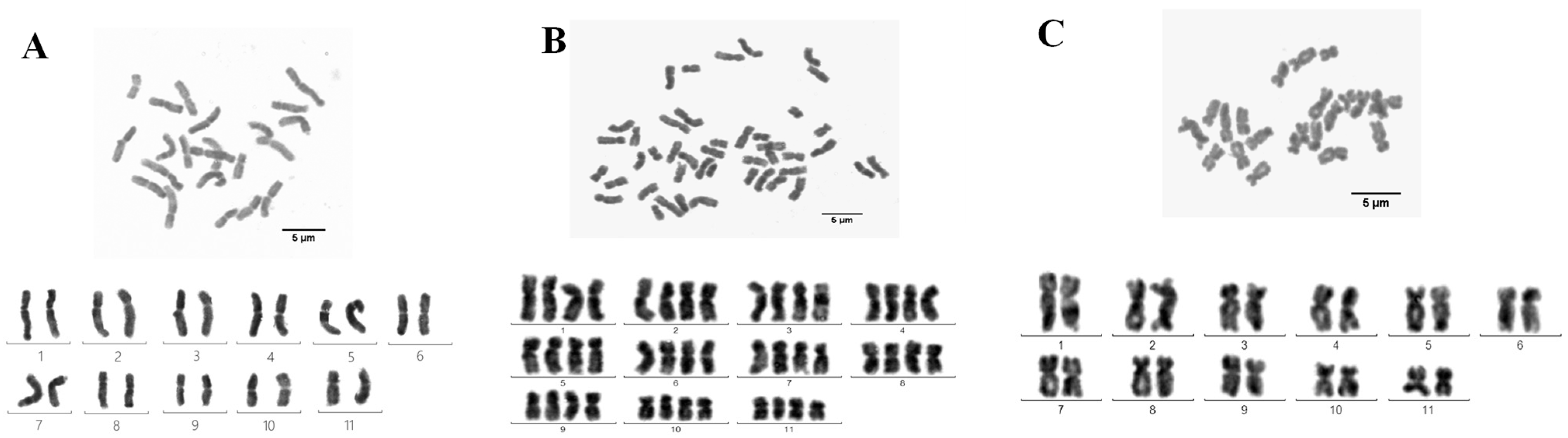
3.2. Karyotype Analysis of Ginger
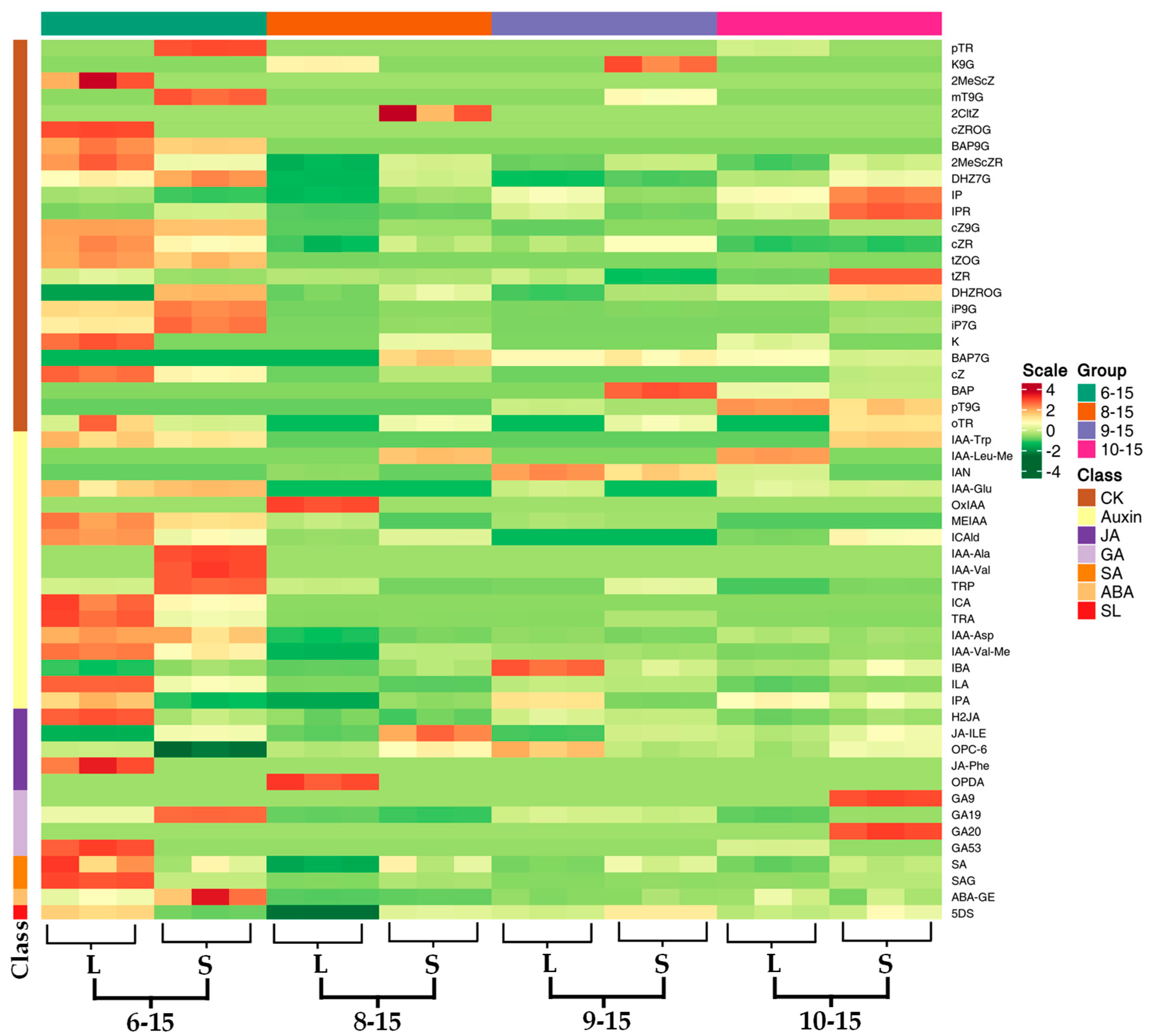
3.3. Analysis of Hormone Levels in Ginger Rhizomes
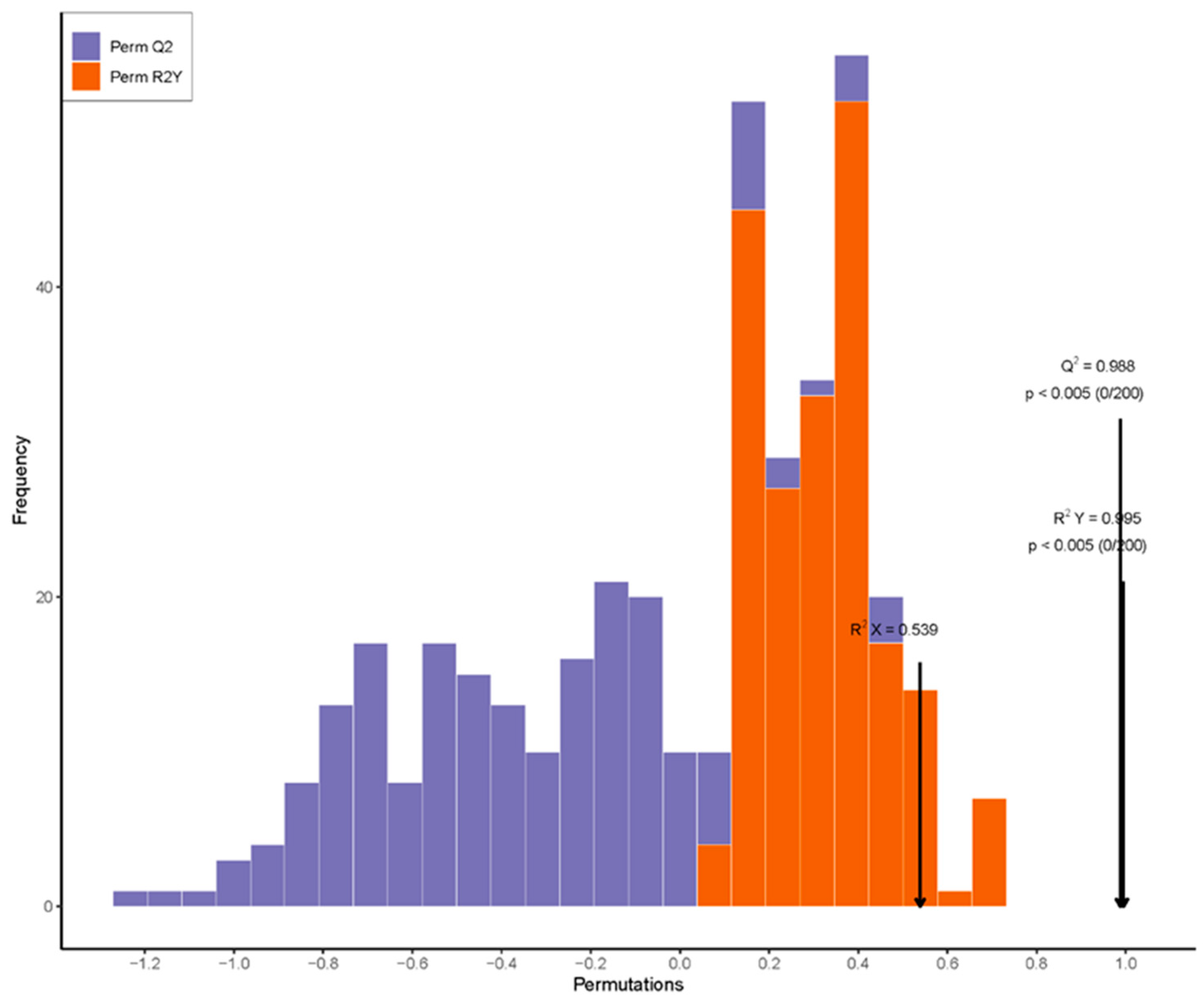
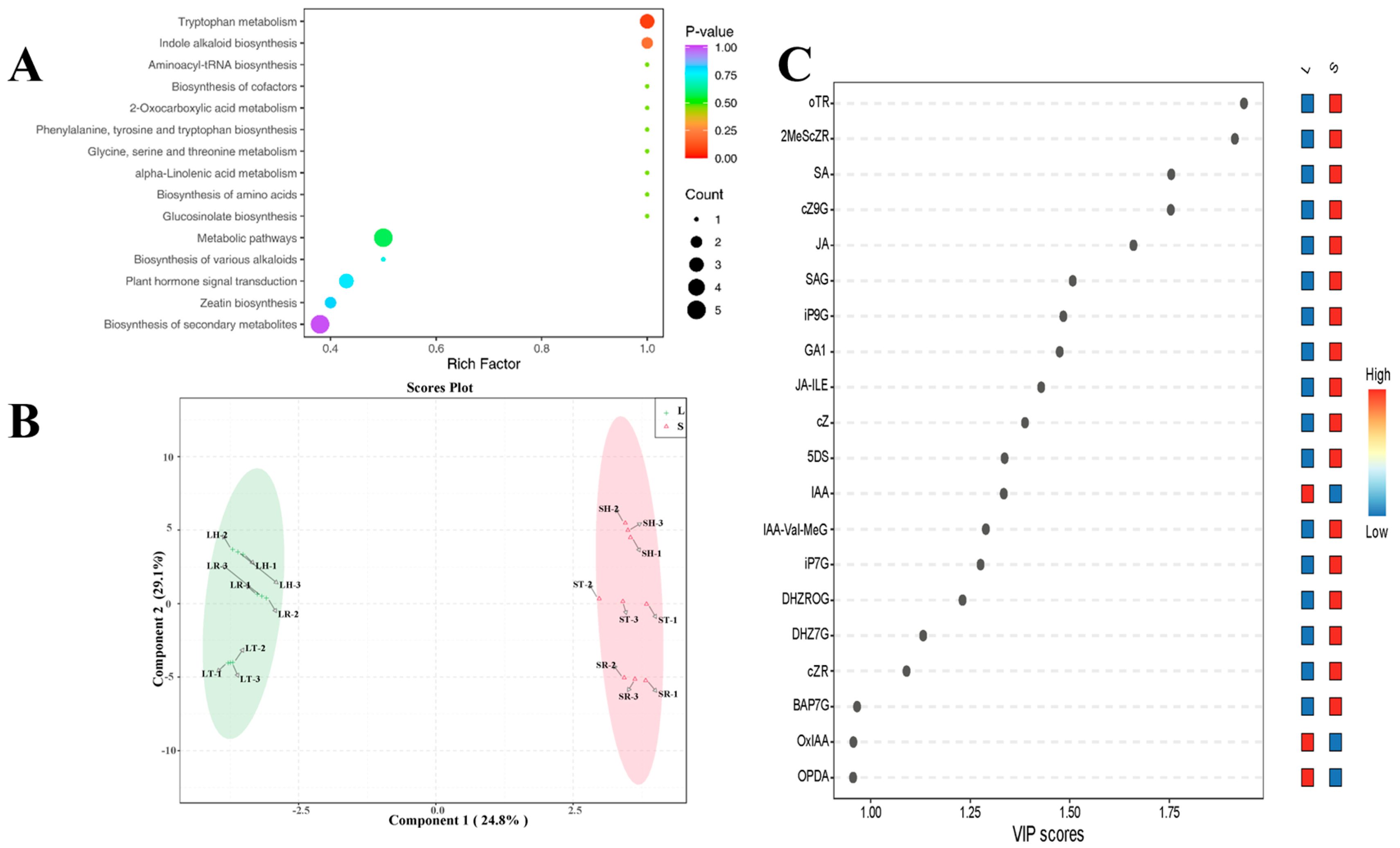
3.4. KEGG Functional Classification and OPLS-DA Analysis of Hormones in Ginger Rhizomes
3.5. Transcriptome Analysis of Ginger Rhizomes
3.6. Quantitative Real-Time PCR of DEGs
4. Discussion
4.1. The Influence of Chromosomal Ploidy Variation on Rhizome Bulking
4.2. Role of Auxin, Cytokinin, and Salicylic Acid in Rhizome Bulking
4.3. Potential Influence of ARF Genes on the Bulking of Ginger Rhizomes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ → 3′) | Tm (°C) |
|---|---|---|
| ZoRPII | F: CTGCTGATGGATACGAATG | 60.0 |
| R: CTGCCCAAGAGAATGAAAG | ||
| ZoARF7 | F: AACTGCGTGTTGGTGTCAGAGG | 61.0 |
| R: CCGTAGTGATTGCGTGGGATGC | ||
| ZoARF23 | F: CAGCGGAAATGCGAGGACAGAG | 60.9 |
| R: GATCGGAGAGAGCAGCGGAAATG | ||
| ZoARF17 | F: GTGGTGGAGGCATTCGGATTGG | 61.5 |
| R: ACTCTCGCTCCTACGGTCCAAC | ||
| ZoGH3.1 | F: TGTTCGTGAAGTCGGAGGTTTGC | 61.4 |
| R: TCGTCTGGGCGGTTGAGGAAG | ||
| ZoPIN1b | F: CCATCAAGAAGGTCGGGCAGAAC | 61.8 |
| R: CACTCGCAGGCGGCATCAAG | ||
| ZoARR9 | F: TCGGACATGACAAGGCTCAGACC | 61.4 |
| R: TTGCTATTGCTGCTGCCACTGG | ||
| ZoPR1 | F: GGATCGGGCGACTGCCAAC | 63.2 |
| R: TCCCACCCACAAGCCCACAG | ||
| ZoNPR1 | F: GAGCATCTCGCCTCCCTCCTC | 62.5 |
| R: ACATCGGTGGACACGGACAGAG | ||
| ZoSABP2 | F: CGCCGCTTCGTTGACATCCG | 62.8 |
| R: AGGCTGTGACCGACGAGGAC |
References
- Varakumar, S.; Umesh, K.V.; Singhal, R.S. Enhanced extraction of oleoresin from ginger (Zingiber officinale) rhizome powder using enzyme-assisted three phase partitioning. Food Chem. 2017, 216, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Fang, H.; Zhang, X.; Yan, Y.-M.; Liu, Y.; Miao, J.; Niu, H.; Feng, W.; Cheng, Y.-X.; Wang, Y. Renoprotective Glycoside Derivatives from Zingiber officinale (Ginger) Peels. J. Agric. Food Chem. 2023, 71, 15170–15185. [Google Scholar] [CrossRef] [PubMed]
- Paula, S.; Linn, B.; Kristina, H.; Jonathan, R.; Johan, J.; Hazem, k.; Cristiane, S.F.; Kristiina, O. Multifunctional Ginger Nanofiber Hydrogels with Tunable Absorption: The Potential for Advanced Wound Dressing Applications. Biomacromolecules 2021, 22, 3202–3215. [Google Scholar] [CrossRef]
- Cao, Z.; Tang, N.; Chen, Z.; Wu, P.; Zheng, J.; Ye, J.; Liu, Y.; Hu, Y.; Zhang, L.; Sun, X.; et al. Transcriptomic analysis reveals transcription factors involved in vascular bundle development and tissue maturation in ginger rhizomes (Zingiber officinale Roscoe). Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 13131. [Google Scholar] [CrossRef]
- Lehretz, G.G.; Sonnewald, S.; Hornyik, C.; Corral, J.M.; Sonnewald, U. Post-transcriptional Regulation of FLOWERING LOCUS T Modulates Heat-Dependent Source-Sink Development in Potato. Curr. Biol. 2019, 29, 1614–1624.e3. [Google Scholar] [CrossRef]
- Jabborova, D.; Choudhary, R.; Azimov, A.; Jabbarov, Z.; Selim, S.; Abu-Elghait, M.; Desouky, S.E.; Azab, I.H.E.; Alsuhaibani, A.M.; Khattab, A.; et al. Composition of Zingiber officinale Roscoe (Ginger), Soil Properties and Soil Enzyme Activities Grown in Different Concentration of Mineral Fertilizers. Horticulturae 2022, 8, 43. [Google Scholar] [CrossRef]
- Lv, X.; Gao, S.; Li, N.; Lv, Y.; Chen, Z.; Cao, B.; Xu, K. Comprehensive insights into the influence of supplemental green light on the photosynthesis of ginger (Zingiber officinale Roscoe). Protoplasma 2022, 259, 1477–1491. [Google Scholar] [CrossRef]
- Zierer, W.; Rüscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and Tuberous Root Development. Annu. Rev. Plant Biol. 2021, 72, 551–580. [Google Scholar] [CrossRef]
- Huang, H.; Wei, Y.; Huang, S.; Lu, S.; Su, H.; Ma, L.; Huang, W. Integrated metabolomic and transcriptomic analyses provide insights into regulation mechanisms during bulbous stem development in the Chinese medicinal herb plant, Stephania kwangsiensis. BMC Plant Biol. 2024, 24, 276. [Google Scholar] [CrossRef]
- Yao, S.; Lan, Z.; Huang, R.; Tan, Y.; Huang, D.; Gu, J.; Pan, C. Hormonal and transcriptional analyses provides new insights into the molecular mechanisms underlying root thickening and isoflavonoid biosynthesis in Callerya speciosa (Champ. ex Benth.) Schot. Sci. Rep. 2021, 11, 9. [Google Scholar] [CrossRef]
- Xue, L.; Wang, Y.; Fan, Y.; Jiang, Z.; Wei, Z.; Zhai, H.; He, S.; Zhang, H.; Yang, Y.; Zhao, N.; et al. IbNF-YA1 is a key factor in the storage root development of sweet potato. Plant J. 2024, 118, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.A.; Lee, H.-S.; Huh, E.J.; Huh, G.H.; Paek, K.-H.; Shin, J.S.; Bae, J.M. SRD1 is involved in the auxin-mediated initial thickening growth of storage root by enhancing proliferation of metaxylem and cambium cells in sweetpotato (Ipomoea batatas). J. Exp. Bot. 2010, 61, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Kondhare, K.R.; Kumar, A.; Patil, N.S.; Malankar, N.N.; Saha, K.; Banerjee, A.K. Development of aerial and belowground tubers in potato is governed by photoperiod and epigenetic mechanism. Plant Physiol. 2021, 187, 1071–1086. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wen, J.; Ke, X.; Zhang, J.; Sun, X.; Wang, C.; Yang, Y. Gibberellin inhibition of taproot formation by modulation of DELLA-NAC complex activity in turnip (Brassica rapa var. rapa). Protoplasma 2021, 258, 925–934. [Google Scholar] [CrossRef]
- Ren, X.; Ma, W.; Xuan, S.; Li, D.; Wang, Y.; Xu, Y.; Feng, D.; Zhao, J.; Chen, X.; Luo, S.; et al. Hormones and carbohydrates synergistically regulate the formation of swollen roots in a Chinese cabbage translocation line. Hortic. Res. 2023, 10, uhad121. [Google Scholar] [CrossRef]
- Kloosterman, B.; Navarro, C.; Bijsterbosch, G.; Lange, T.; Prat, S.; Visser, R.G.F.; Bachem, C.W.B. StGA2ox1 is induced prior to stolon swelling and controls GA levels during potato tuber development. Plant J. 2007, 52, 362–373. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Y.-Q.; Li, G.-L.; Zhang, S.-Y. Genome-wide identification, characterization, and expression patterns of the BZR transcription factor family in sugar beet (Beta vulgaris L.). BMC Plant Biol. 2019, 19, 191. [Google Scholar] [CrossRef]
- Deng, R.; Huang, S.; Du, J.; Luo, D.; Liu, J.; Zhao, Y.; Zheng, C.; Lei, T.; Li, Q.; Zhang, S.; et al. The brassinosteroid receptor StBRI1 promotes tuber development by enhancing plasma membrane H+-ATPase activity in potato. Plant Cell 2024, 36, 3498–3520. [Google Scholar] [CrossRef]
- Jing, S.; Sun, X.; Yu, L.; Wang, E.; Cheng, Z.; Liu, H.; Jiang, P.; Qin, J.; Begum, S.; Song, B. Transcription factor StABI5-like 1 binding to the FLOWERING LOCUS T homologs promotes early maturity in potato. Plant Physiol. 2022, 189, 1677–1693. [Google Scholar] [CrossRef]
- Sun, Q.; Zhou, G.F.; Cai, Y.F.; Fan, Y.H.; Zhu, X.Y.; Liu, Y.H.; He, X.H.; Shen, J.J.; Jiang, H.Z.; Hu, D.W.; et al. Transcriptome analysis of stem development in the tumourous stem mustard Brassica juncea var. tumida Tsen et Lee by RNA sequencing. BMC Plant Biol. 2012, 12, 53. [Google Scholar] [CrossRef]
- Pasare, S.A.; Ducreux, L.J.M.; Morris, W.L.; Campbell, R.; Sharma, S.K.; Roumeliotis, E.; Kohlen, W.; van der Krol, S.; Bramley, P.M.; Roberts, A.G.; et al. The role of the potato (Solanum tuberosum) CCD8 gene in stolon and tuber development. New Phytol. 2013, 198, 1108–1120. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Li, W.B.; Li, Z.X.; Zhang, W.L.; Jue, D.W.; Xing, H.T.; Li, H.L.; Li, Q. Dynamic transcriptome profiling provides insights into rhizome enlargement in ginger (Zingiber officinale Rosc.). PLoS ONE 2023, 18, e0287969. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y. The Mechanism of Ginger Rhizome Formationand Enlargement. Doctor’s Dissertation, Shandong Agricultural University, Tai’an, China, 2021. [Google Scholar]
- Wang, L.; Gao, F.S.; Xu, K.; Li, X. Natural occurrence of mixploid ginger (Zingiber officinale Rosc.) in China and its morphological variations. Sci. Hortic. 2014, 172, 54–60. [Google Scholar] [CrossRef]
- Liu, M.; Wang, G.; Liu, G.; Ma, F.; Bao, Z. Biostimulants Promote the Sedimentation of Salts to Restore Tomato Plant Growth Under Salt Stress. J. Soil Sci. Plant Nutr. 2023, 23, 1830–1844. [Google Scholar] [CrossRef]
- Xin, R. Research on Crossing Compatibility and Karyotype Analysis in Paeonia. Master’s Dissertation, Shenyang Agricultural University, Shenyang, China, 2020. [Google Scholar]
- Sheng, C.; Song, S.; Zhou, W.; Dossou, S.S.K.; Zhou, R.; Zhang, Y.; Li, D.; You, J.; Wang, L. Integrating transcriptome and phytohormones analysis provided insights into plant height development in sesame. Plant Physiol. Biochem. 2023, 198, 107695. [Google Scholar] [CrossRef]
- Lv, Y.; Li, Y.; Liu, X.; Xu, K. Identification of Ginger (Zingiber officinale Roscoe) Reference Genes for Gene Expression Analysis. Front. Genet. 2020, 11, 586098. [Google Scholar] [CrossRef]
- Bozan, I.; Achakkagari, S.R.; Anglin, N.L.; Ellis, D.; Tai, H.H.; Strömvik, M.V. Pangenome analyses reveal impact of transposable elements and ploidy on the evolution of potato species. Proc. Natl. Acad. Sci. USA 2023, 120, 11. [Google Scholar] [CrossRef]
- Guo, H.; Zhou, M.; Zhang, G.; He, L.; Yan, C.; Wan, M.; Hu, J.; He, W.; Zeng, D.; Zhu, B.; et al. Development of homozygous tetraploid potato and whole genome doubling-induced the enrichment of H3K27ac and potentially enhanced resistance to cold-induced sweetening in tubers. Hortic. Res. 2023, 10, uhad017. [Google Scholar] [CrossRef]
- Mahmud, E.; Zhu, H.J.; Kaseb, M.O.; Sajjad, M.Z.; He, N.; Lu, X.Q.; Liu, W.G. Polyploidization Impact on Plant Architecture of Watermelon (Citrullus lanatus). Horticulturae 2024, 10, 24. [Google Scholar] [CrossRef]
- Pu, T.; Wang, Y.; Han, W.; Li, H.; Sun, P.; Suo, Y.; Fu, J. Physiological Characteristics and Transcriptional Differences of Growth Traits of Persimmon with Different Ploidy. Horticulturae 2024, 10, 207. [Google Scholar] [CrossRef]
- Kondhare, K.R.; Patil, A.B.; Giri, A.P. Auxin: An emerging regulator of tuber and storage root development. Plant Sci. 2021, 306, 110854. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, Y.; Tanaka, M.; Utsumi, C.; Takahashi, S.; Matsui, A.; Fukushima, A.; Kobayashi, M.; Sasaki, R.; Oikawa, A.; Kusano, M.; et al. Integrative omics approaches revealed a crosstalk among phytohormones during tuberous root development in cassava. Plant Mol. Biol. 2022, 109, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Chakrabarti, S.K.; Makeshkumar, T.; Saravanan, R. Molecular Regulation of Storage Root Formation and Development in Sweet Potato. In Horticultural Reviews; John Wiley & Sons: Hoboken, NJ, USA, 2014; Volume 42. [Google Scholar]
- Abts, W.; Vandenbussche, B.; De Proft, M.P.; Van de Poel, B. The Role of Auxin-Ethylene Crosstalk in Orchestrating Primary Root Elongation in Sugar Beet. Front. Plant Sci. 2017, 8, 444. [Google Scholar] [CrossRef]
- Roumeliotis, E.; Visser, R.G.F.; Bachem, C.W.B. A crosstalk of auxin and GA during tuber development. Plant Signal. Behav. 2014, 7, 1360–1363. [Google Scholar] [CrossRef]
- Hartmann, A.; Senning, M.; Hedden, P.; Sonnewald, U.; Sonnewald, S. Reactivation of Meristem Activity and Sprout Growth in Potato Tubers Require Both Cytokinin and Gibberellin. Plant Physiol. 2011, 155, 776–796. [Google Scholar] [CrossRef]
- Borzenkova, R.A.; Borovkova, M.P. Developmental Patterns of Phytohormone Content in the Cortex and Pith of Potato Tubers as Related to Their Growth and Starch Content. Russ. J. Plant Physiol. 2003, 50, 119–124. [Google Scholar]
- Dong, T.; Zhu, M.; Yu, J.; Han, R.; Tang, C.; Xu, T.; Liu, J.; Li, Z. RNA-Seq and iTRAQ reveal multiple pathways involved in storage root formation and development in sweet potato (Ipomoea batatas L.). BMC Plant Biol. 2019, 19, 136. [Google Scholar] [CrossRef]
- Tao, G.Q.; Stuart, D.; Yong, J.W.H.; Zhang, K.; Farquhar, G.D. Promotion of shoot development and tuberisation in potato by expression of a chimaeric cytokinin synthesis gene at normal and elevated CO2 levels. Funct. Plant Biol. 2010, 37, 43–54. [Google Scholar]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benková, E.; Mähönen, A.P.; Helariutta, Y. A Mutually Inhibitory Interaction between Auxin and Cytokinin Specifies Vascular Pattern in Roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, H.; Ruan, W.; Deng, M.; Wang, F.; Peng, J.; Luo, J.; Chen, Z.; Yi, K. ABNORMAL INFLORESCENCE MERISTEM1 Functions in Salicylic Acid Biosynthesis to Maintain Proper Reactive Oxygen Species Levels for Root Meristem Activity in Rice. Plant Cell 2017, 29, 560–574. [Google Scholar] [CrossRef]
- Ali, I.; Wang, X.; Tareen, M.J.; Wattoo, F.M.; Qayyum, A.; Hassan, M.U.; Shafique, M.; Liaquat, M.; Asghar, S.; Hussain, T.; et al. Foliar Application of Salicylic Acid at Different Phenological Stages of Peach Fruit CV. ‘Flordaking’ Improves Harvest Quality and Reduces Chilling Injury during Low Temperature Storage. Plants 2021, 10, 1981. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, Y.; Yuan, D.; Duan, M.; Liu, Y.; Shen, Z.; Yang, C.; Qiu, Z.; Liu, D.; Wen, P.; et al. An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice. Proc. Natl. Acad. Sci. USA 2019, 117, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, S.; Li, F.; Zhang, H.; Sun, R.; Li, G.; Zhang, S. Cloning and Functional Analysis of BraTSD2 Associated with Root Swelling in Turnip (Brassica rapa L.). Horticulturae 2025, 11, 33. [Google Scholar] [CrossRef]
- Zhou, Y.; Yao, L.; Xie, Y.; Huang, B.; Li, Y.; Huang, X.; Yu, L.; Pan, C. Metabolic and transcriptional analysis of tuber expansion in Curcuma kwangsiensis. Sci. Rep. 2025, 15, 1588. [Google Scholar] [CrossRef]
- Wang, D.; Pei, K.; Fu, Y.; Sun, Z.; Li, S.; Liu, H.; Tang, K.; Han, B.; Tao, Y. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene 2007, 394, 13–24. [Google Scholar] [CrossRef]
- Yang, C. Functional Analysis of Auxin-responsive Genes StARF2 and StARF5 in Potato Tuber Development. Doctor’s Dissertation, Northwest A&F University, Xianyang, China, 2022. [Google Scholar]
- Shen, C.; Wang, S.; Bai, Y.; Wu, Y.; Zhang, S.; Chen, M.; Guilfoyle, T.J.; Wu, P.; Qi, Y. Functional analysis of the structural domain of ARF proteins in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 3971–3981. [Google Scholar] [CrossRef]
- Sun, C.Y.; Fan, K.; Wang, X.; Liu, H.H.; Guo, N.P.; Liu, W.Y.; Ye, G.X.; Lin, W.W.; Lin, W.X.; Li, Z.W. The involvement of auxin response factor OsARF7 in positively regulating root development by mediating the expression of OsCRL1 in rice (Oryza sativa L.). Plant Mol. Biol. 2025, 115, 38. [Google Scholar] [CrossRef]
- Li, G.; Liang, W.; Zhang, X.; Ren, H.; Hu, J.; Bennett, M.J.; Zhang, D. Rice actin-binding protein RMD is a key link in the auxin–actin regulatory loop that controls cell growth. Proc. Natl. Acad. Sci. USA 2014, 111, 10377–10382. [Google Scholar] [CrossRef]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1andAUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef]
- Li, Y.; Yang, S.; Shi, M.; Zhang, S.; Wu, S.; Chen, Y.; Li, W.; Tian, W.-M. HbARF2 and HbARF16.3 function as negative regulators for the radial trunk growth of rubber tree. Ind. Crops Prod. 2020, 158, 112978. [Google Scholar] [CrossRef]



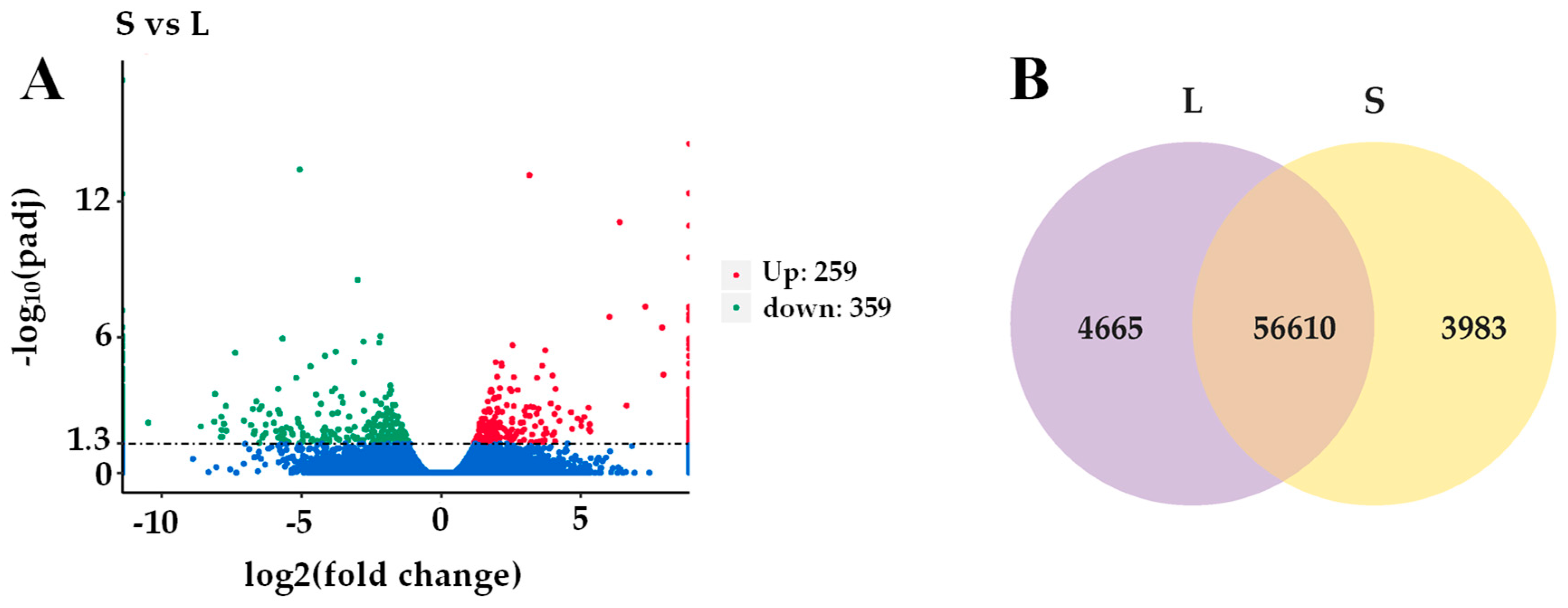
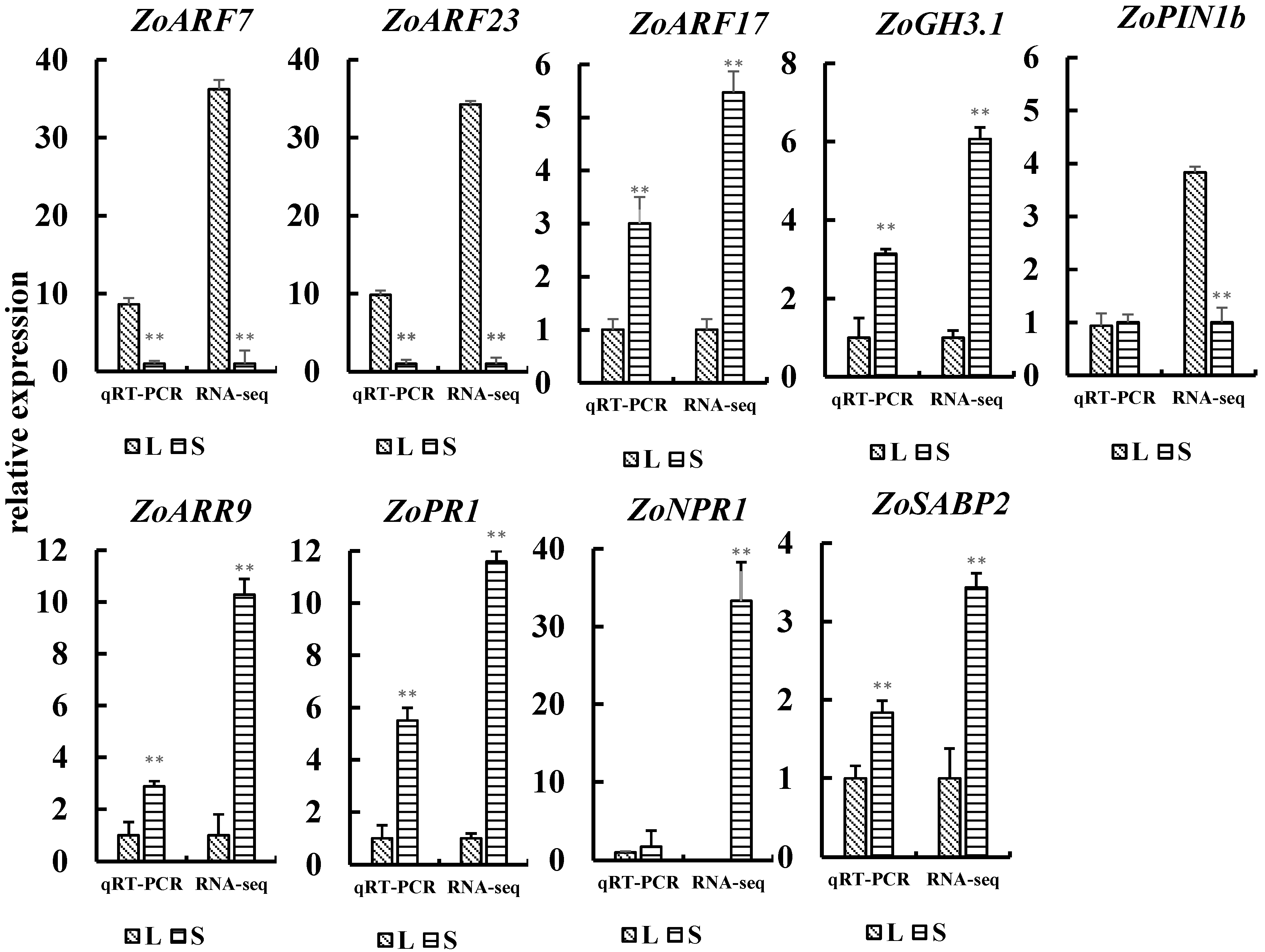

| Generation | Plant Height | Stem Diameter | Shoot Number | Root FW | Stem FW | Leaf FW | Rhizome FW | Leaf Number of the Main Stem | |
|---|---|---|---|---|---|---|---|---|---|
| (cm) | (mm) | (g/Plant) | (g/Plant) | (g/Plant) | (g/Plant) | ||||
| VM1 | L | 94.8 ± 3.9 | 12.5 ± 0.4 | 16.7 ± 0.6 | 116.0 ± 6.2 | 580.4 ± 48.6 | 226.8 ± 17.5 | 1014.9 ± 27.8 | 27.3 ± 1.2 |
| S | 84.4 ± 1.1 ** | 9.4 ± 0.5 ** | 50.3 ± 1.5 ** | 223.5 ± 14.6 ** | 824.3 ± 55.5 ** | 319.2 ± 36.7 ** | 513.7 ± 25.4 ** | 28.0 ± 2.0 | |
| VM2 | L | 99.8 ± 4.1 | 13.2 ± 0.2 | 24.7 ± 2.1 | 131.9 ± 6.7 | 604.2 ± 44.7 | 262.9 ± 23.8 | 1033.3 ± 36.9 | 27.3 ± 1.2 |
| S | 86.5 ± 3.6 ** | 9.3 ± 0.5 ** | 59.7 ± 3.1 ** | 249.6 ± 12.2 ** | 859.5 ± 36.9 ** | 349.8 ± 33.7 ** | 553.8 ± 34.9 ** | 29.3 ± 1.2 | |
| VM3 | L | 109.8 ± 7.8 | 14.3 ± 1.3 | 26.0 ± 1.7 | 129.6 ± 6.2 | 593.6 ± 53.1 | 241.9 ± 16.6 | 1101.4 ± 100.8 | 25.3 ± 2.3 |
| S | 86.8 ± 3.7 ** | 10.7 ± 0.9 ** | 62.3 ± 1.5 ** | 254.1 ± 19.3 ** | 855.3 ± 55.5 ** | 346.5 ± 35.3 ** | 530.8 ± 25.4 ** | 25.3 ± 4.2 | |
| Variety | No. | Relative Lengths (%) | Arm Ratio | Centromere Index (%) | Chromosomal Type | No. | Relative Lengths (%) | Arm Ratio | Centromere Index (%) | Chromosomal Type | As.K. (%) | Karyotypic Type | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Short Arm | Long Arm | Total Length | Short Arm | Long Arm | Total Length | ||||||||||||
| L | Diploid cells | 1 | 4.22 | 7.52 | 11.74 | 1.78 | 35.95 | sm | 7 | 3.30 | 5.48 | 8.78 | 1.66 | 37.59 | m | 63.56 | 2A |
| 2 | 4.16 | 6.53 | 10.69 | 1.57 | 38.91 | m | 8 | 3.50 | 5.08 | 8.58 | 1.45 | 40.79 | m | ||||
| 3 | 3.56 | 6.40 | 9.96 | 1.80 | 35.74 | sm | 9 | 2.71 | 5.54 | 8.25 | 2.05 | 32.85 | sm | ||||
| 4 | 3.50 | 6.07 | 9.57 | 1.74 | 36.57 | sm | 10 | 2.64 | 4.62 | 7.26 | 1.75 | 36.36 | sm | ||||
| 5 | 3.17 | 6.34 | 9.51 | 2.00 | 33.33 | sm | 11 | 2.71 | 3.7 | 6.41 | 1.37 | 42.28 | m | ||||
| 6 | 2.97 | 6.27 | 9.24 | 2.11 | 32.14 | sm | |||||||||||
| Tetraploid cells | 1 | 4.23 | 7.36 | 11.59 | 1.74 | 36.50 | sm | 7 | 3.01 | 5.88 | 8.90 | 1.95 | 33.87 | sm | 62.90 | 2A | |
| 2 | 4.29 | 6.41 | 10.70 | 1.49 | 40.11 | m | 8 | 3.36 | 5.16 | 8.52 | 1.54 | 39.46 | m | ||||
| 3 | 3.01 | 7.42 | 10.43 | 2.46 | 28.89 | sm | 9 | 3.48 | 4.52 | 8.00 | 1.30 | 43.48 | m | ||||
| 4 | 3.57 | 6.35 | 9.91 | 1.78 | 35.96 | sm | 10 | 2.52 | 4.20 | 6.73 | 1.67 | 37.50 | m | ||||
| 5 | 3.13 | 6.67 | 9.80 | 2.13 | 31.95 | sm | 11 | 2.61 | 3.71 | 6.32 | 1.42 | 41.29 | m | ||||
| 6 | 3.88 | 5.22 | 9.10 | 1.34 | 42.68 | m | |||||||||||
| S | Diploid cells | 1 | 4.21 | 7.64 | 11.85 | 1.82 | 35.51 | sm | 7 | 3.65 | 4.98 | 8.63 | 1.37 | 42.28 | m | 64.45 | 2A |
| 2 | 4.28 | 6.66 | 10.94 | 1.56 | 39.10 | m | 8 | 2.38 | 6.17 | 8.56 | 2.59 | 27.87 | sm | ||||
| 3 | 3.44 | 6.52 | 9.96 | 1.90 | 34.51 | sm | 9 | 3.16 | 5.19 | 8.35 | 1.64 | 37.82 | m | ||||
| 4 | 3.44 | 6.17 | 9.61 | 1.80 | 35.77 | sm | 10 | 2.38 | 4.49 | 6.87 | 1.88 | 34.69 | sm | ||||
| 5 | 3.09 | 6.45 | 9.54 | 2.09 | 32.36 | sm | 11 | 2.67 | 3.79 | 6.45 | 1.42 | 41.31 | m | ||||
| 6 | 2.88 | 6.38 | 9.26 | 2.22 | 31.06 | sm | |||||||||||
| Unigene ID | Annotation | log2FoldChange | p-Value |
|---|---|---|---|
| Auxin-Related Genes | |||
| i3_LQ_ZR_c21855/f1p1/3608 | auxin response factor 7 | 5.1786 | 4.64 × 10−8 |
| i2_LQ_ZR_c19192/f1p0/2780 | auxin response factor 23 | Inf | 5.02 × 10−6 |
| i2_LQ_ZR_c59231/f1p0/2020 | indole-3-acetic acid-amido synthetase GH3.1 | 2.1061 | 6.73 × 10−6 |
| i2_LQ_ZR_c53301/f1p0/2680 | auxin efflux carrier component 1b | 1.9388 | 1.03 × 10−5 |
| i3_HQ_ZR_c81940/f26p0/3334 | auxin response factor 7 | 10.473 | 1.81 × 10−5 |
| i6_LQ_ZR_c2184/f1p0/6466 | auxin response factor 23 | 5.0987 | 1.82 × 10−5 |
| i3_LQ_ZR_c59597/f1p1/3518 | auxin response factor 17 | −2.4521 | 7.97 × 10−5 |
| i3_HQ_ZR_c35172/f2p2/3385 | auxin response factor 7 | 1.6362 | 0.000132 |
| Cytokinin-Related Genes | |||
| i2_LQ_ZR_c20879/f1p8/2289 | two-component response regulator ARR9 | −2.6022 | 1.73 × 10−6 |
| i0_LQ_ZR_c6058/f1p7/795 | two-component response regulator ARR9 | −3.3633 | 2.96 × 10−5 |
| i1_LQ_ZR_c33483/f1p0/1060 | two-component response regulator ARR9 | −2.1816 | 3.86 × 10−6 |
| i1_LQ_ZR_c27791/f1p0/1034 | two-component response regulator ARR9 | −2.3556 | 1.49 × 10−6 |
| Salicylic Acid-Related Genes | |||
| i0_LQ_ZR_c12160/f1p0/670 | pathogenesis-related protein 1 | −3.5348 | 0.000257 |
| i3_LQ_ZR_c19736/f1p0/3757 | ankyrin repeat-containing protein NPR1 | −Inf | 1.55 × 10−6 |
| i1_LQ_ZR_c22124/f1p3/1057 | salicylic acid-binding protein 2 | −1.7817 | 2.64 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Lv, Y.; Gao, S.; Kong, Y.; Liu, M.; Chen, Z.; Xu, K. Integrated Analysis of Hormone Metabolomics and Transcriptomics Reveals the Differential Mechanism of Ginger (Zingiber officinale Roscoe) Rhizome Bulking. Agronomy 2025, 15, 779. https://doi.org/10.3390/agronomy15040779
Wang K, Lv Y, Gao S, Kong Y, Liu M, Chen Z, Xu K. Integrated Analysis of Hormone Metabolomics and Transcriptomics Reveals the Differential Mechanism of Ginger (Zingiber officinale Roscoe) Rhizome Bulking. Agronomy. 2025; 15(4):779. https://doi.org/10.3390/agronomy15040779
Chicago/Turabian StyleWang, Kai, Yao Lv, Song Gao, Yuwen Kong, Miaohong Liu, Zijing Chen, and Kun Xu. 2025. "Integrated Analysis of Hormone Metabolomics and Transcriptomics Reveals the Differential Mechanism of Ginger (Zingiber officinale Roscoe) Rhizome Bulking" Agronomy 15, no. 4: 779. https://doi.org/10.3390/agronomy15040779
APA StyleWang, K., Lv, Y., Gao, S., Kong, Y., Liu, M., Chen, Z., & Xu, K. (2025). Integrated Analysis of Hormone Metabolomics and Transcriptomics Reveals the Differential Mechanism of Ginger (Zingiber officinale Roscoe) Rhizome Bulking. Agronomy, 15(4), 779. https://doi.org/10.3390/agronomy15040779