
Genetic Characterization and Fruit Quality Analysis of New Mandarin Hybrids

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Characterizations of Mandarin and Their Hybrids
2.2. Physical–Chemical Analyses
2.3. Response to Alternaria Alternata Inoculation
2.4. Molecular Characterization
3. Results and Discussion
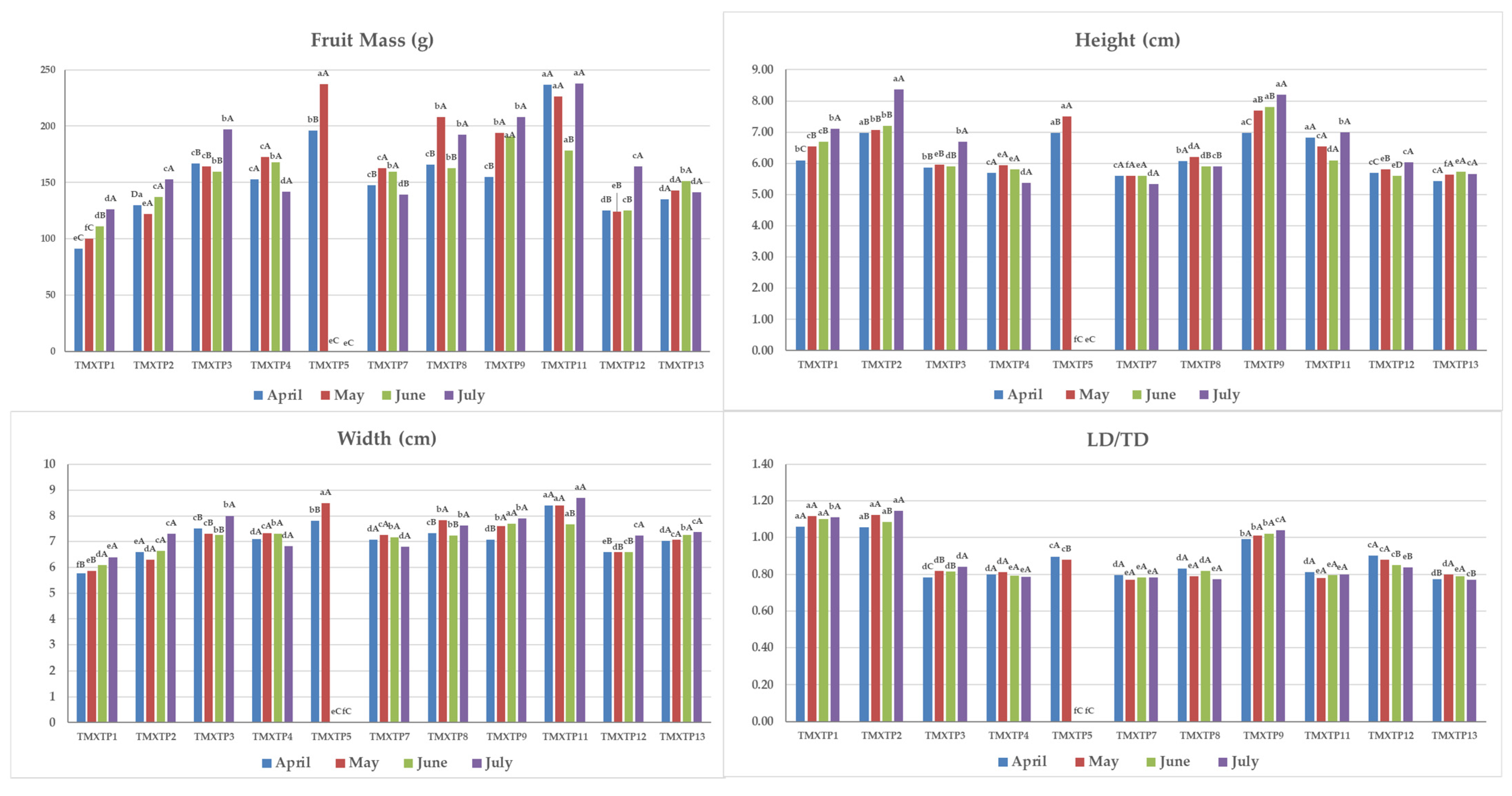
3.1. Morphological Characterization of Mandarins and Their Hybrids
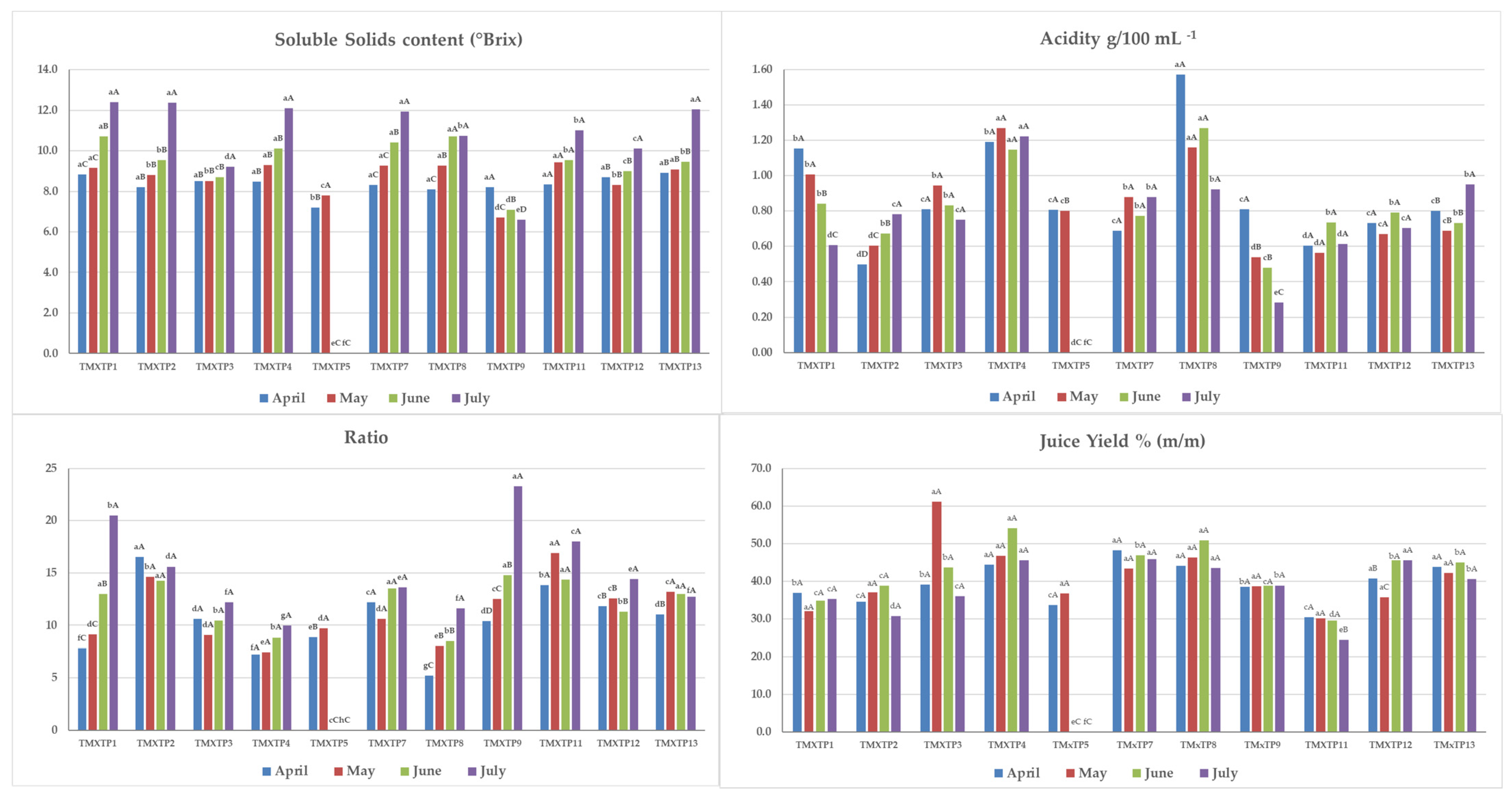
3.2. Physical–Chemical Analyses of Fruits
3.3. Reaction of Genotypes to Inoculation with Alternaria alternata
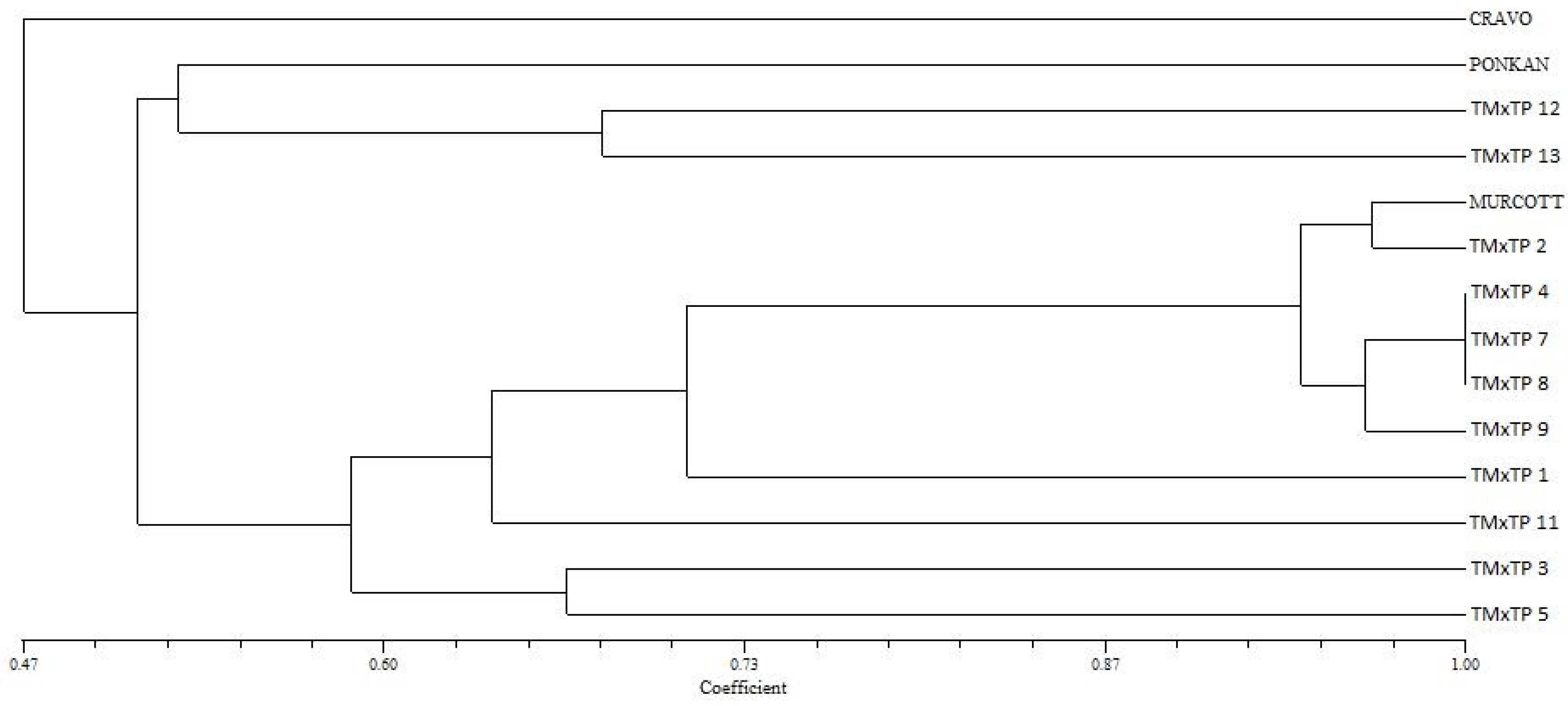
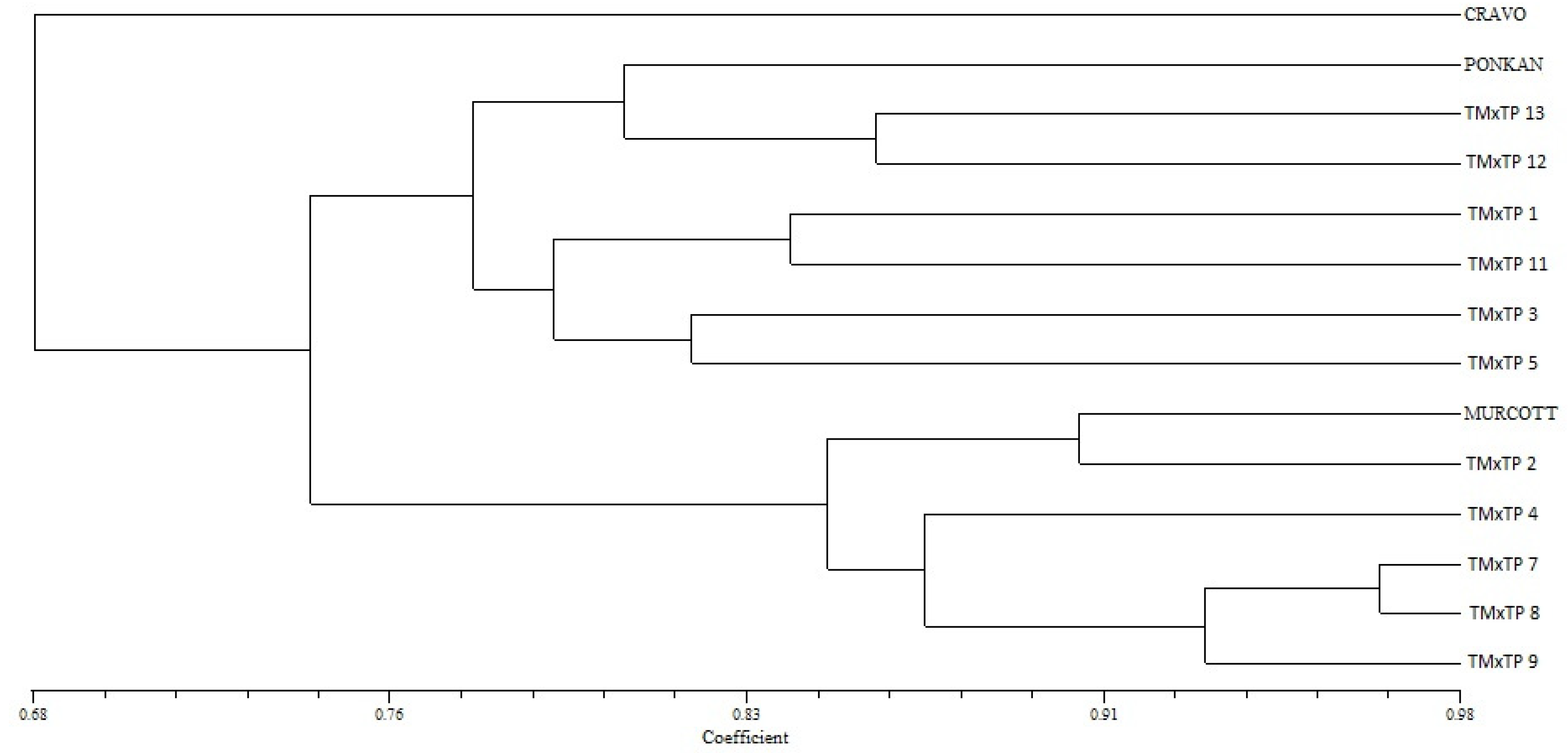
3.4. Molecular Characterization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Instituto Brasileiro de Geografia e Estatística. IBGE. 2021. Available online: https://www.ibge.gov.br (accessed on 10 March 2024).
- Bastianel, M.; Martinelli, R.; Devite, F.T.; Cristofani-Yaly, M.; Ferreira, R.d.V.; Stuchi, E.S.; Azevedo, F.A.d. Reaction of mandarins to the Alternaria Brown spot and huanglongbing: Identification of potential varieties for these diseases to be managed in the field. Horticulturae 2023, 9, 641. [Google Scholar] [CrossRef]
- Timmer, L.W.; Darhower, H.M.; Zitko, S.E.; Peever, T.L.; Ibanez, A.M.; Bushong, P.M. Environmental factors affecting the severity of Alternaria brown spot of citrus and their potential use in timing fungicide applications. Plant Dis. 2000, 84, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Vicent, A.; Badal, J.; Asensi, M.J.; Sanz, N.; Armengol, J.; Garc, J. Laboratory evaluation of citrus cultivars susceptibility and influence of fruit size on Fortune mandarin to infection by Alternaria alternata pv. citri. Eur. J. Plant Pathol. 2004, 110, 245–251. [Google Scholar] [CrossRef]
- Cuenca, J.; Navarro, L.; Aleza, P. Advances in mandarin breeding. In Achieving Sustainable Cultivation of Tropical Fruits; Burleigh Dodds Science Publishing: London, UK, 2019; Volume 1, pp. 183–212. [Google Scholar]
- Stitou, M.; Fadli1, A.; Chetto, O.; Talha, A.; Benkirane, R.; Benyahia, H. Fruit quality analysis in four new mandarin hybrids during maturation period. Annu. Res. Rev. Biol. 2017, 18, 1–10. [Google Scholar] [CrossRef]
- Tarancón, P.; Giménez-Sanchis, A.; Aleza, P.; Besada, C. Selection of new lateseason mandarin cultivars based on sensory changes and consumer acceptance after fruit cold storage. Agronomy 2021, 11, 116. [Google Scholar] [CrossRef]
- .Yu, Q.; Dai, F.; Russo, R.; Guha, A.; Pierre, M.; Zhuo, X.; Wang, Y.Z.; Vincent, C.; Gmitter, F.G., Jr. Phenotypic and Genetic Variation in Morphophysiological Traits in Huanglongbing-Affected Mandarin Hybrid Populations. Plants 2023, 12, 42. [Google Scholar] [CrossRef]
- Deng, H.; Achor, D.; Exteberria, E.; Yu, Q.; Du, D.; Stanton, D.; Liang, G.; Gmitter, F.G., Jr. Phloem regeneration is a mechanism for Huanglongbing-tolerance of “Bearss” lemon and “LB8-9” Sugar Belle® mandarin. Front. Plant Sci. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Peever, T.L.; Canihos, Y.; Olsen, L.; Ibañez, A.; Liu, Y.C.; Timmer, L.W. Population Genetic Structure and Host Specificity of Alternaria spp. Causing Brown Spot of Minneola Tangelo and Rough Lemon in Florida. Phytopathology 1999, 89, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Martelli, I.B.; Pacheco, C.A.; Bastianel, M.; Schinor, E.H.; Conceição, P.M.; Azevedo, F.A. Diagrammatic scale for assessing foliar symptoms of alternaria brown spot in citrus. Agron. Sci. Biotechnol. 2016, 2, 56–61. [Google Scholar] [CrossRef]
- Althaus, R.A.; Canteri, M.G.; Giglioti, E.A. Tecnologia da Informação Aplicada ao Agronegócio e CIÊNCIAS ambientais: Sistema para Análise e Separação de Médias Pelos Métodos de Duncan, Tukey e Scott-Knott; Parte 1; Anais do X Encontro Anual de Iniciação Científica: Ponta Grossa, Brazil, 2001; pp. 280–281. [Google Scholar]
- Scott, A.; Knott, M. A Cluster Analysis Method for Grouping Means in the Analysis of Variance. Biometrics 1974, 30, 507. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acid Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Dezotti, A.L.; Soratto, T.A.T.; Bastianel, M.; Novelli, V.M.; Cristofani-Yaly, M. Seleção de marcadores específicos para caracterização de variedades do grupo das tangerinas e híbridos. UNAR Rev. Científica Cent. Univ. Araras “Dr. Edmundo Ulson” 2017, 14, 21–39. [Google Scholar] [CrossRef]
- Palmieri, D.A.; Novelli, V.M.; Bastianel, M.; Cristofani-Yaly, M.; Astúa-Monge, G.; Carlos, E.F.; Oliveira, A.C.; Machado, M.A. Frequency and distribution of microsatellites from ESTs of citrus. CitrusEST Genet. Mol. Biol. 2007, 30, 1009–1018. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYSpc Numerical Taxonomy and Multivariate Analysis System Version 2.0 User Guide; Applied Biostatistics Inc.: Port Jefferson, NY, USA, 1998. [Google Scholar]
- Marshall, T.C.; Slate, J.; Kruk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Farooq, N. Planilha RAPD data for PIC.; University of the Punjab, Institute of Agricultural Sciences: Lahore, Pakistan, 2017; Available online: https://www.researchgate.net/post/How_can_we_find_PIC_IB_RP_MI_SI_index_EMR_and_H_of_RAPD_data (accessed on 1 November 2013).
- CEAGESP. Normas de Classificação de Citros de Mesa; CEAGESP: São Paulo, Brazil, 2011; pp. 1–12. [Google Scholar]
- Pio, R.M.; Minami, K. Caracterização dos frutos das variedades de tangerinas Sul da África e Thomas. Laranja 2002, 23, 473–488. [Google Scholar]
- Azevedo, F.A.; Polydoro, D.A.; Bastianel, M.; Kupper, K.C.; Stuart, R.M.; Costa, F.P.; Pio, R.M. Resposta de diferentes genótipos de tangerinas e seus híbridos à inoculação in vitro e in vivo de Alternaria alternata. Rev. Bras. Frutic. 2010, 32, 944–951. [Google Scholar] [CrossRef]
- Gulsen, O.; Uzun, A.; Canan, I.; Seday, U.; Canihos, E. A new citrus linkage map based on SRAP, SSR, ISSR, POGP, RGA and RAPD markers. Euphytica 2010, 173, 265–277. [Google Scholar] [CrossRef]
- Cuenca, J.; Aleza, P.; Vicent, A.; Brunel, D.; Ollitrault, P.; Navarro, L. Genetically Based Location from Triploid Populations and Gene Ontology of a 3.3-Mb Genome Region Linked to Alternaria Brown Spot Resistance in Citrus Reveal Clusters of Resistance Genes. PLoS ONE 2013, 8, e76755. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest Month | Number of Seeds | ||||||
|---|---|---|---|---|---|---|---|
| April | May | June | July | Polyembrionics | Aborted Seeds | Total | |
| Ponkan like | |||||||
| TMxTP 1 | 13.2 a | 6.8 a | 20.0 a | ||||
| TMxTP 2 | 4.6 b | 8.4 a | 13.0 a | ||||
| TMxTP 3 | 19.6 a | 3.0 b | 22.6 a | ||||
| TMxTP 9 | 14.0 a | 3.4 b | 17.4 a | ||||
| TMxTP 11 | 10.6 a | 1.8 b | 12.4 a | ||||
| TMxTP 13 | 13.0 a | 1.4 b | 14.4 a | ||||
| Murcott like | |||||||
| TMxTP 4 | 12.8 a | 4.2 a | 17.0 a | ||||
| TMxTP 7 | 15.2 a | 4.8 a | 20.0 a | ||||
| TMxTP 8 | 8.8 a | 2.8 b | 11.6 a | ||||
| TMxTP 12 | 12.6 a | 3.6 b | 16.2 a | ||||
| Hours After Inoculation | AUDPC | |||||||
|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||||||
| TMxTP 1 | 0.00 | dA | 0.00 | eA | 0.00 | dA | 0.00 | e |
| TMxTP 3 | 0.00 | dA | 0.00 | eA | 0.00 | dA | 0.00 | e |
| TMxTP 11 | 0.00 | dA | 0.00 | eA | 0.00 | dA | 0.00 | e |
| Pera Sweet orange | 0.00 | dA | 0.00 | eA | 0.00 | dA | 0.00 | e |
| Cravo mandarin | 0.00 | dB | 0.26 | eB | 1.50 | dA | 24.30 | e |
| TMxTP 5 | 0.51 | dA | 2.55 | eA | 10.09 | cA | 188.40 | d |
| Ponkan mandarin | 1.43 | cB | 4.26 | dB | 16.18 | cA | 313.50 | d |
| TMxTP 4 | 1.79 | cB | 5.43 | dB | 33.25 | bA | 550.65 | d |
| Murcott tangor | 1.46 | cB | 6.30 | dB | 37.04 | bA | 613.20 | c |
| TMxTP 2 | 1.46 | cB | 9.81 | cB | 40.38 | bA | 737.55 | c |
| TMxTP 9 | 0.70 | cC | 4.06 | dB | 75.00 | aA | 1005.90 | c |
| TMxTP 13 | 2.70 | bC | 13.63 | cB | 78.63 | aA | 1302.90 | b |
| TMxTP 7 | 1.10 | cC | 13.25 | cB | 89.25 | aA | 1402.20 | b |
| TMxTP 12 | 4.06 | aC | 57.75 | bB | 96.13 | aA | 2588.25 | a |
| TMxTP 8 | 4.94 | aB | 86.50 | aA | 94.88 | aA | 3273.75 | a |
| CV (%) | 21.52 | 23.06 | 22.61 | 25.21 | ||||
| SSR | Tandem Repeats | Primer Sequence | Number of Alleles | Fragment Size (bp) | Heterozygosity | |||
|---|---|---|---|---|---|---|---|---|
| Forward (5′-3′) | Reverse (5′-3′) | Observed (Ho) | Expected (Hexp) | PIC | ||||
| CCSM18 | (AG)n | aacagttgatgaagaggaag | Gtgattgctggtgtcgtt | 2 | 200–300 | 0.286 | 0.476 | 0.354 |
| CCSM24 | (GA)26 | attgacagaagtctg | Ccacttaatcactctcct | 2 | 100–200 | 0.214 | 0.389 | 0.305 |
| CCSM29 | (TGA)15(TTA)9 | cgtgattgtgtccga | Cacacttcacaatgttgcac | 3 | 100–200 | 0.714 | 0.624 | 0.523 |
| CCSM46 | (GCA)6(CAA)8 | ataccttatcaagtaacacg | Tcagaatgagtactagctcc | 2 | <150 | 0.714 | 0.519 | 0.375 |
| CCSM-EST-11 | (TC)12 | ttcatggcagcttgagtttc | Agtcatggaagccaaaatgg | 2 | 200–300 | 0.643 | 0.452 | 0.341 |
| CCSM-EST-89 | (ATA)7 | acttatcttgcacccgacga | Gaggtctcgaagtcacggag | 3 | 100–200 | 0.071 | 0.558 | 0.468 |
| CCSM-EST-92 | (GCC)6 | aagcatcgtcaaagtttggg | Ttgatgcatgttctcaaggc | 2 | 300–400 | 0.786 | 0.495 | 0.363 |
| CCSM-EST-159 | (TTCTTG)4 | tgggtcattgatgttgtgct | Cacagatgcagaaggggatt | 2 | 100–200 | 0.857 | 0.508 | 0.370 |
| CCSM-EST-164 | (TC)11 | gagaagcccgtctgcactta | Acgagagcggaaacaagaga | 2 | <150 | 0.714 | 0.476 | 0.354 |
| CCSM-EST-169 | (ATGATC)4 | acgtcgctagatcctgtgct | Catacaccaaacaccgtcca | 2 | 200–300 | 0.071 | 0.071 | 0.067 |
| CCSM-EST-187 | (CTCCAC)4 | cagggactcaaaacacgaca | Tatgatgatggtggtggtcg | 3 | <150 | 0.643 | 0.685 | 0.587 |
| CCSM-EST-191 | (CAG)9 | gagggagtggctatgcaaga | Tcgagattcaattgctgcac | 3 | 100–200 | 0.500 | 0.611 | 0.514 |
| CCSM-EST-234 | (GGC)7 | aatgcgtgggcaataacttc | Ttcaatatcggcccaaactc | 2 | 200–300 | 0.643 | 0.452 | 0.341 |
| TRAPs | Alleles Total | Number of Polymorphic Alleles | P (%) * | PIC | Primers Sequences | |
|---|---|---|---|---|---|---|
| Forward (5′-3′) | Reverse (5′-3′) | |||||
| Trap1 + Arbi1 | 7 | 5 | 71.4 | 0.52 | gcccgtgctgcctgatgatt | gactgcgtacgaattaat |
| Trap9 + Arbi2 | 12 | 8 | 66.6 | 0.33 | gggcggtgatcctgagaa | gactgcgtacgaatttgc |
| Trap2 + Arbi3 | 10 | 5 | 50.0 | 0.32 | acagggccaaaggtaaac | gactgcgtacgaattgac |
| Trap10 + Arbi3 | 5 | 1 | 20.0 | 0.08 | cagtttcttgttgctacg | gactgcgtacgaattgac |
| Trap11 + Arbi4 | 10 | 7 | 63.6 | 0.28 | tccccgaggcacagcatc | gactgcgtacgaatttga |
| Trap5 + Arbi5 | 8 | 5 | 62.5 | 0.46 | acgcgtccgccactctca | gactgcgtacgaattaac |
| Trap2 + Arbi6 | 9 | 5 | 55.5 | 0.26 | acagggccaaaggtaaac | gactgcgtacgaattgca |
| Trap12 + Arbi6 | 7 | 4 | 57.1 | 0.49 | tcggactggtatcatgga | gactgcgtacgaattgca |
| TOTAL | 68 | 40 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristofani-Yaly, M.; Dezotti, A.L.; Cavichioli, T.M.; Roverssi, F.; Novelli, V.M.; Bastianel, M. Genetic Characterization and Fruit Quality Analysis of New Mandarin Hybrids. Agronomy 2024, 14, 2666. https://doi.org/10.3390/agronomy14112666
Cristofani-Yaly M, Dezotti AL, Cavichioli TM, Roverssi F, Novelli VM, Bastianel M. Genetic Characterization and Fruit Quality Analysis of New Mandarin Hybrids. Agronomy. 2024; 14(11):2666. https://doi.org/10.3390/agronomy14112666
Chicago/Turabian StyleCristofani-Yaly, Mariângela, Ana Lúcia Dezotti, Thaís Magni Cavichioli, Fernanda Roverssi, Valdenice Moreira Novelli, and Marinês Bastianel. 2024. "Genetic Characterization and Fruit Quality Analysis of New Mandarin Hybrids" Agronomy 14, no. 11: 2666. https://doi.org/10.3390/agronomy14112666
APA StyleCristofani-Yaly, M., Dezotti, A. L., Cavichioli, T. M., Roverssi, F., Novelli, V. M., & Bastianel, M. (2024). Genetic Characterization and Fruit Quality Analysis of New Mandarin Hybrids. Agronomy, 14(11), 2666. https://doi.org/10.3390/agronomy14112666