
Response of Bread Wheat Cultivars to Terminal Water Stress and Cytokinin Application from a Grain Phenotyping Perspective



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Treatments and Experimental Design
2.3. Imaging
2.4. Image Processing
2.5. Statistical Analyses
3. Results
3.1. TGW Responses under TWS
3.2. Different TGW and Phenotyping Indices Responses across Cultivars
3.3. Relationship between Grain Phenotyping Indices and TWS
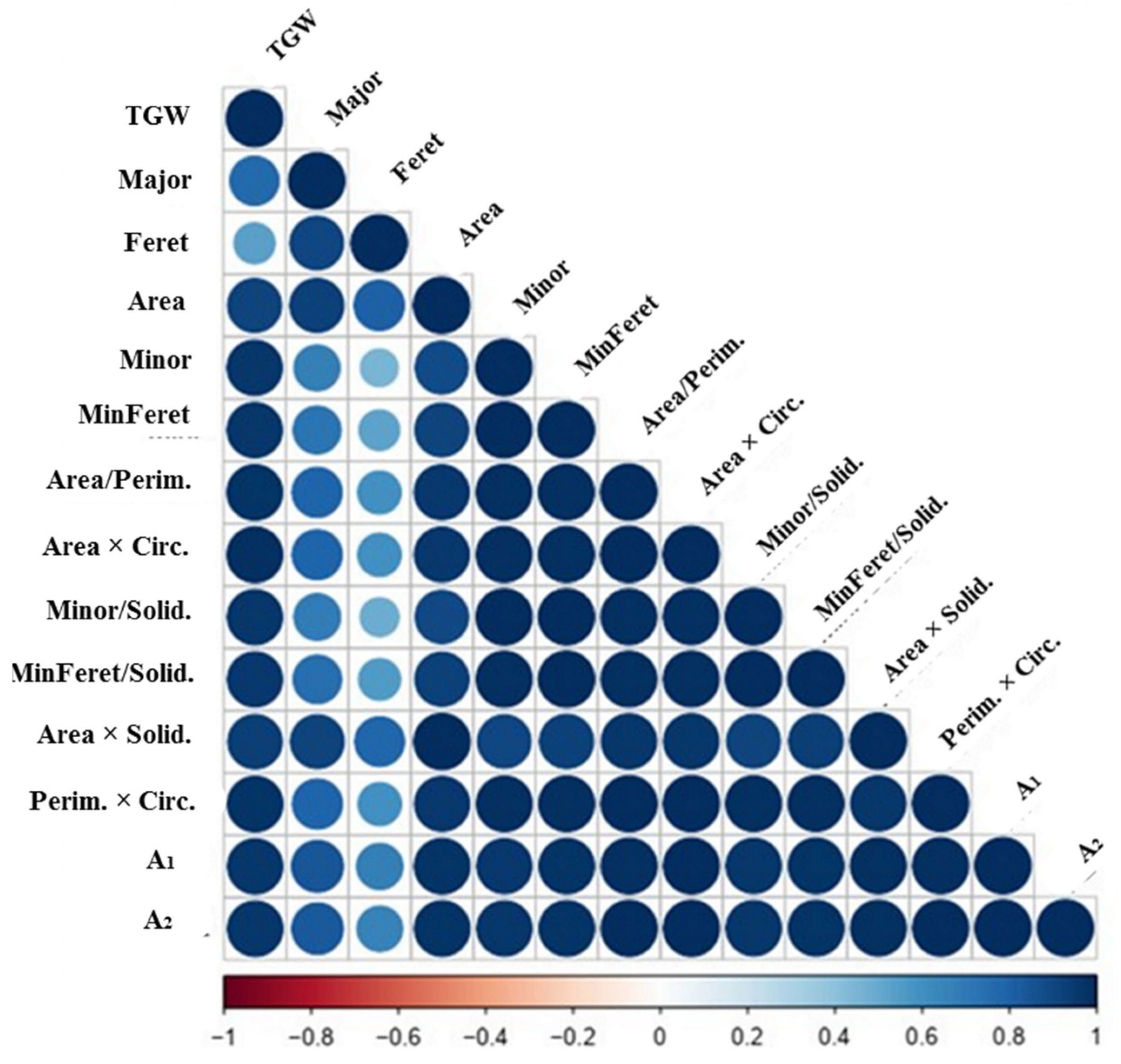
3.4. Dissecting Grain Phenotyping Indices Relationships through Correlations
4. Discussion
4.1. TWS and Changes in TGW as a Function of Water Availability
4.2. Relationship between TGW and Grain Phenotyping Indices
4.3. Responsiveness of Cultivars to Growth Conditions
4.4. Effects of the 6-BA Application
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate Variation Explains a Third of Global Crop Yield Variability. Nat. Commun. 2015, 6, 5989. [Google Scholar] [CrossRef] [PubMed]
- Helman, D.; Bonfil, D.J. Six Decades of Warming and Drought in the World’s Top Wheat-Producing Countries Offset the Benefits of Rising CO2 to Yield. Sci. Rep. 2022, 12, 7921. [Google Scholar] [CrossRef] [PubMed]
- Leng, G.; Hall, J. Crop Yield Sensitivity of Global Major Agricultural Countries to Droughts and the Projected Changes in the Future. Sci. Total Environ. 2019, 654, 811–821. [Google Scholar] [CrossRef]
- Vahamidis, P.; Karamanos, A.J.; Economou, G. Grain Number Determination in Durum Wheat as Affected by Drought Stress: An Analysis at Spike and Spikelet Level. Ann. Appl. Biol. 2019, 174, 190–208. [Google Scholar] [CrossRef]
- Slafer, G.A.; Savin, R.; Pinochet, D.; Calderini, D.F. Wheat. In Crop Physiology Case Histories for Major Crops; Victor, O.S., Daniel, F.C., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 98–163. [Google Scholar]
- Tambussi, E.A.; Bort, J.; Araus, J.L. Water Use Efficiency in C3 Cereals under Mediterranean Conditions: A Review of Physiological Aspects. Ann. Appl. Biol. 2007, 150, 307–321. [Google Scholar] [CrossRef]
- Haghshenas, A.; Emam, Y. Image-Based Tracking of Ripening in Wheat Cultivar Mixtures: A Quantifying Approach Parallel to the Conventional Phenology. Comput. Electron. Agric. 2019, 156, 318–333. [Google Scholar] [CrossRef]
- Brinton, J.; Uauy, C. A Reductionist Approach to Dissecting Grain Weight and Yield in Wheat. J. Integr. Plant Biol. 2019, 61, 337–358. [Google Scholar] [CrossRef]
- Beral, A.; Rincent, R.; Le Gouis, J.; Girousse, C.; Allard, V. Wheat Individual Grain-Size Variance Originates from Crop Development and from Specific Genetic Determinism. PLoS ONE 2020, 15, e0230689. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin Action in Response to Abiotic and Biotic Stresses in Plants. Plant. Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Shi, Y.; Cui, Z.; Luo, Y.; Zheng, M.; Chen, J.; Li, Y.; Yin, Y.; Wang, Z. Exogenous Cytokinins Increase Grain Yield of Winter Wheat Cultivars by Improving Stay-Green Characteristics under Heat Stress. PLoS ONE 2016, 11, e0155437. [Google Scholar] [CrossRef]
- Hai, N.N.; Chuong, N.N.; Tu, N.H.C.; Kisiala, A.; Hoang, X.L.T.; Thao, N.P. Role and Regulation of Cytokinins in Plant Response to Drought Stress. Plants 2020, 9, 422. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Ghorai, M.; Anand, U.; Samanta, D.; Kant, N.; Mishra, T.; Rahman, M.H.; Jha, N.K.; Jha, S.K.; Lal, M.K.; et al. Cytokinin and Abiotic Stress Tolerance -What Has Been Accomplished and the Way Forward? Front. Genet. 2022, 13, 943025. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A.; van Staden, J. The Involvement of Cytokinins in Plant Responses to Environmental Stress. Plant Growth Regul. 1997, 23, 79–103. [Google Scholar] [CrossRef]
- Zarea, M.J.; Karimi, N. Grain Yield and Quality of Wheat Are Improved through Post-Flowering Foliar Application of Zinc and 6-Benzylaminopurine under Water Deficit Condition. Front. Plant Sci. 2022, 13, 1068649. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Peng, S.; Visperas, R.M.; Sanico, A.L.; Zhu, Q.; Gu, S. Grain Filling Pattern and Cytokinin Content in the Grains and Roots of Rice Plants. Plant Growth Regul. 2000, 30, 261–270. [Google Scholar] [CrossRef]
- Ramya, P.; Chaubal, A.; Kulkarni, K.; Gupta, L.; Kadoo, N.; Dhaliwal, H.S.; Chhuneja, P.; Lagu, M.; Gupt, V. QTL Mapping of 1000-Kernel Weight, Kernel Length, and Kernel Width in Bread Wheat (Triticum aestivum L.). J. Appl. Genet. 2010, 51, 421–429. [Google Scholar] [CrossRef]
- García, G.A.; Serrago, R.A.; González, F.G.; Slafer, G.A.; Reynolds, M.P.; Miralles, D.J. Wheat Grain Number: Identification of Favourable Physiological Traits in an Elite Doubled-Haploid Population. Field Crops Res. 2014, 168, 126–134. [Google Scholar] [CrossRef]
- Hughes, A.; Askew, K.; Scotson, C.P.; Williams, K.; Sauze, C.; Corke, F.; Doonan, J.H.; Nibau, C. Non-Destructive, High-Content Analysis of Wheat Grain Traits Using X-Ray Micro Computed Tomography. Plant Methods 2017, 13, 76. [Google Scholar] [CrossRef]
- Le, T.D.Q.; Alvarado, C.; Girousse, C.; Legland, D.; Chateigner-Boutin, A.-L. Use of X-Ray Micro Computed Tomography Imaging to Analyze the Morphology of Wheat Grain through Its Development. Plant Methods 2019, 15, 84. [Google Scholar] [CrossRef]
- Xiong, B.; Wang, B.; Xiong, S.; Lin, C.; Yuan, X. 3D Morphological Processing for Wheat Spike Phenotypes Using Computed Tomography Images. Remote Sens. 2019, 11, 1110. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, C.; Jiang, Y.; Huang, C.; Liu, Q.; Xiong, L.; Yang, W.; Chen, F. Nondestructive 3D Image Analysis Pipeline to Extract Rice Grain Traits Using X-Ray Computed Tomography. Plant Phenomics 2020, 2020, 3414926. [Google Scholar] [CrossRef] [PubMed]
- Haghshenas, A.; Emam, Y.; Jafarizadeh, S. Wheat Grain Width: A Clue for Re-Exploring Visual Indicators of Grain Weight. Plant Methods 2022, 18, 58. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liu, Y.; Zhang, P.; Chen, T.; Tian, T.; Wang, P.; Che, Z.; Shahinnia, F.; Yang, D. Identification of Quantitative Trait Loci (QTL) and Meta-QTL Analysis for Kernel Size-Related Traits in Wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 607. [Google Scholar] [CrossRef] [PubMed]
- Rabieyan, E.; Bihamta, M.R.; Esmaeilzadeh Moghaddam, M.; Mohammadi, V.; Alipour, H. Imaging-Based Screening of Wheat Seed Characteristics towards Distinguishing Drought-Responsive Iranian Landraces and Cultivars. Crop Pasture Sci. 2022, 73, 337–355. [Google Scholar] [CrossRef]
- Halder, J.; Gill, H.S.; Zhang, J.; Altameemi, R.; Olson, E.; Turnipseed, B.; Sehgal, S.K. Genome-Wide Association Analysis of Spike and Kernel Traits in the U.S. Hard Winter Wheat. Plant Genome 2023, 16, e20300. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.S.; Lidon, F.C. Evaluation of Grain Filling Rate and Duration in Bread and Durum Wheat, under Heat Stress after Anthesis. J. Agron. Crop Sci. 2009, 195, 137–147. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought Stress in Wheat during Flowering and Grain-Filling Periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Carpel Size, Grain Filling, and Morphology Determine Individual Grain Weight in Wheat. J. Exp. Bot. 2015, 66, 6715–6730. [Google Scholar] [CrossRef]
- Cohen, I.; Zandalinas, S.I.; Huck, C.; Fritschi, F.B.; Mittler, R. Meta-Analysis of Drought and Heat Stress Combination Impact on Crop Yield and Yield Components. Physiol. Plant. 2021, 171, 66–76. [Google Scholar] [CrossRef]
- Latif, S.; Wang, L.; Khan, J.; Ali, Z.; Sehgal, S.K.; Ali Babar, M.; Wang, J.; Quraishi, U.M. Deciphering the Role of Stay-Green Trait to Mitigate Terminal Heat Stress in Bread Wheat. Agronomy 2020, 10, 1001. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Brocklehurst, P.A. Factors Controlling Grain Weight in Wheat. Nature 1977, 266, 348–349. [Google Scholar] [CrossRef]
- Touzy, G.; Lafarge, S.; Redondo, E.; Lievin, V.; Decoopman, X.; Le Gouis, J.; Praud, S. Identification of QTLs Affecting Post-Anthesis Heat Stress Responses in European Bread Wheat. Theor. Appl. Genet. 2022, 135, 947–964. [Google Scholar] [CrossRef] [PubMed]
- Samarah, N.H. Effects of Drought Stress on Growth and Yield of Barley. Agron. Sustain. Dev. 2005, 25, 145–149. [Google Scholar] [CrossRef]
- Lizana, X.C.; Riegel, R.; Gomez, L.D.; Herrera, J.; Isla, A.; McQueen-Mason, S.J.; Calderini, D.F. Expansins Expression Is Associated with Grain Size Dynamics in Wheat (Triticum aestivum L.). J. Exp. Bot. 2010, 61, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.K.; Herrera, J.; Lizana, C.; Calderini, D.F. Carpel Weight, Grain Length and Stabilized Grain Water Content Are Physiological Drivers of Grain Weight Determination of Wheat. Field Crops Res. 2011, 123, 241–247. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, L.; Gao, H.; Hu, M.; Mu, L.; Cheng, X.; Wang, J.; Zhao, Y.; Li, Q.; Wang, P.; et al. Genome-Wide Association Study of Yield-Related Traits in Common Wheat (Triticum aestivum L.) under Normal and Drought Treatment Conditions. Front. Plant Sci. 2023, 13, 1098560. [Google Scholar] [CrossRef]
- Gegas, V.C.; Nazari, A.; Griffiths, S.; Simmonds, J.; Fish, L.; Orford, S.; Sayers, L.; Doonan, J.H.; Snape, J.W. A Genetic Framework for Grain Size and Shape Variation in Wheat. Plant Cell 2010, 22, 1046–1056. [Google Scholar] [CrossRef]
- Du, B.; Wang, Q.; Sun, G.; Ren, X.; Cheng, Y.; Wang, Y.; Gao, S.; Li, C.; Sun, D. Mapping Dynamic QTL Dissects the Genetic Architecture of Grain Size and Grain Filling Rate at Different Grain-Filling Stages in Barley. Sci. Rep. 2019, 9, 18823. [Google Scholar] [CrossRef]
- Ji, G.; Xu, Z.; Fan, X.; Zhou, Q.; Chen, L.; Yu, Q.; Liao, S.; Jiang, C.; Feng, B.; Wang, T. Identification and Validation of Major QTL for Grain Size and Weight in Bread Wheat (Triticum aestivum L.). Crop J. 2023, 11, 564–572. [Google Scholar] [CrossRef]
- Gaspar, T.; Franck, T.; Bisbis, B.; Kevers, C.; Jouve, L.; Hausman, J.F.; Dommes, J. Concepts in Plant Stress Physiology. Application to Plant Tissue Cultures. Plant Growth Regul. 2002, 37, 263–285. [Google Scholar] [CrossRef]
- Shah, S.M.; Shabbir, G.; Malik, S.I.; Raja, N.I.; Shah, Z.H.; Rauf, M.; Zahrani, Y.A.; Alghabari, F.; Alsamadany, H.; Shahzad, K.; et al. Delineation of Physiological, Agronomic and Genetic Responses of Different Wheat Genotypes under Drought Condition. Agronomy 2022, 12, 1056. [Google Scholar] [CrossRef]
- Maddah Hosseini, S.; Poustini, K.; Ahmadi, A. Effects of Foliar Application of BAP on Source and Sink Strength in Four Six-Rowed Barley (Hordeum vulgare L.) Cultivars. Plant Growth Regul. 2008, 54, 231–239. [Google Scholar] [CrossRef]
- Ren, B.; Zhu, Y.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Effects of Spraying Exogenous Hormone 6-Benzyladenine (6-BA) after Waterlogging on Grain Yield and Growth of Summer Maize. Field Crops Res. 2016, 188, 96–104. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Regulations of 6-Benzyladenine (6-BA) on Leaf Ultrastructure and Photosynthetic Characteristics of Waterlogged Summer Maize. J. Plant Growth Regul. 2017, 36, 743–754. [Google Scholar] [CrossRef]
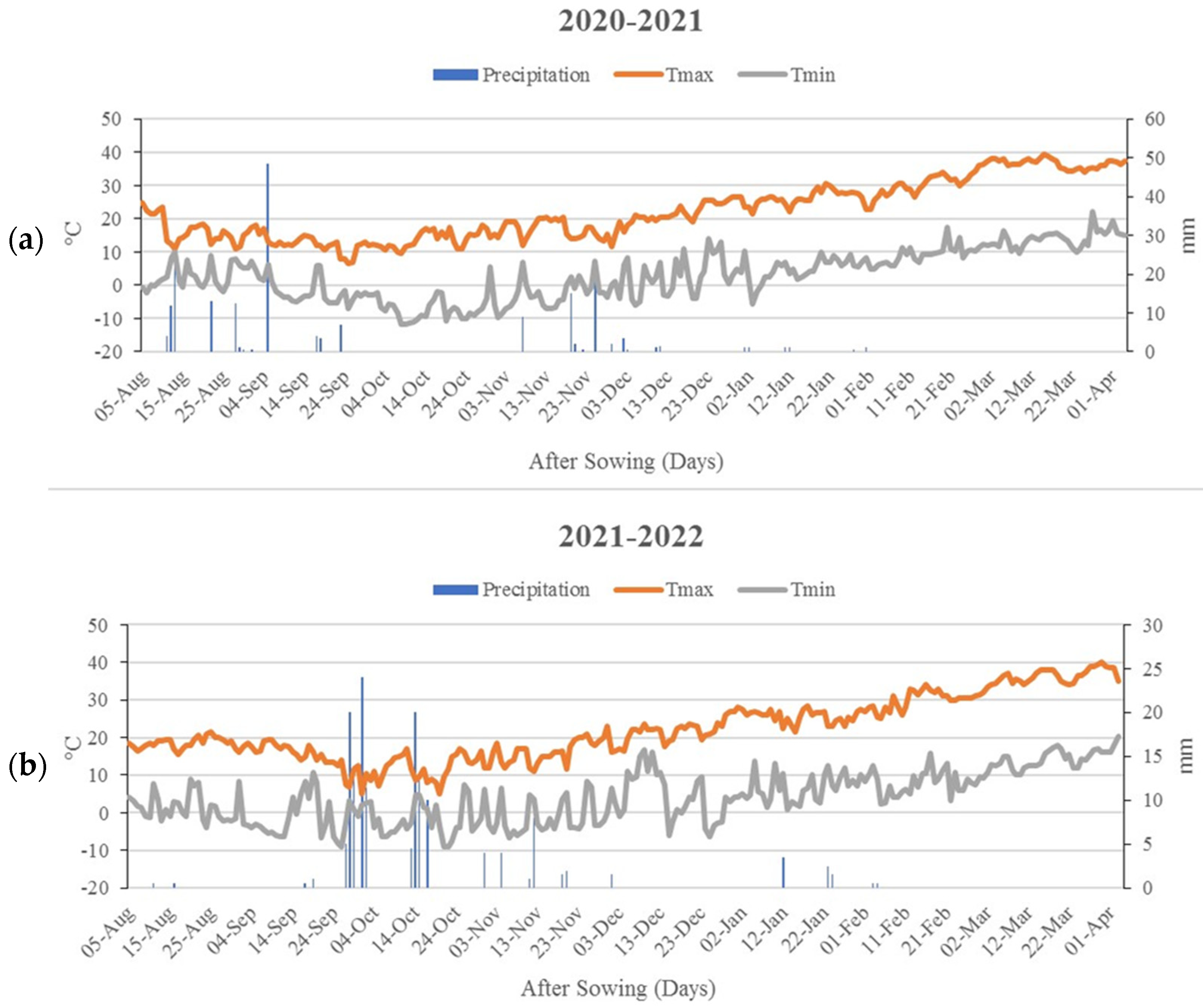


{kind=link}
{kind=link}
{kind=link}
| Basic Indices | Synthesized Indices |
|---|---|
| Area | Perim. × Circ. |
| Minor (the minor axes of the best fitted ellipses to the grains) | Area/Perim. |
| Major (the major axes of the best fitted ellipses to the grains) | Area × Circ. |
| MinFeret (minimum caliper diameter) | Minor/Solid. |
| Feret (maximum caliper diameter) | MinFeret/Solid. |
| Area × Solid. | |
| A1 (Area × Perim. × Circ. × Solidity × MinFeret) | |
| A2 (Area × Perim. × Circ. × Solidity × Minor) |
| Source | DF | TGW | Major | Feret | Area | Minor | MinFeret | Area/Perim | Area × Circ. | Minor/Solid. | MinFeret/Solid. | Area × Solid | Perim. × Circ. | A1 | A2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rep | 2 | 0.314 | 0.043 | 0.0482 | 0.404 | 0.002 | 0.002 | 0.0002 | 0.125 | 0.002 | 0.002 | 0.365 | 0.037 | 1513.28 | 1363.63 |
| Irrigation | 1 | 71.844 *** | 0.171 | 0.261 | 27.814 *** | 0.8217 *** | 0.845 *** | 0.052 *** | 26.179 *** | 0.798 *** | 0.813 *** | 27.830 *** | 8.396 *** | 183,973.5 *** | 173,902.7 *** |
| Irrigation × rep | 2 | 3.862 | 0.027 | 0.023 | 1.204 | 0.022 | 0.022 | 0.002 | 0.864 | 0.023 | 0.022 | 1.145 | 0.266 | 6791.90 | 6434.85 |
| Cytokinin | 1 | 1.5477 | 0.0001 | 0.001 | 0.344 | 0.018 | 0.019 | 0.001 | 0.477 | 0.018 | 0.019 | 0.339 | 0.170 | 2453.31 | 2266.16 |
| Irrigation × Cytokinin | 1 | 0.787 | 0.0007 | 0.0003 | 0.124 | 0.003 | 0.002 | 0.0001 | 0.107 | 0.002 | 0.002 | 0.132 | 0.031 | 940.96 | 890.60 |
| Cytokin × rep (Irrigat) | 4 | 0.821 | 0.010 | 0.006 | 0.389 | 0.009 | 0.01 | 0.0006 | 0.313 | 0.009 | 0.01 | 0.376 | 0.107 | 1920.38 | 1791.81 |
| Cultivar | 2 | 6.965 * | 0.493 *** | 0.5823 *** | 5.69 *** | 0.046 * | 0.071 * | 0.004 * | 2.166 * | 0.047 * | 0.076 ** | 5.148 *** | 0.659 * | 24,257.2 * | 20,392.73 *** |
| Irrigation × Cultivar | 2 | 4.921 | 0.0015 | 0.0002 | 0.535 | 0.023 | 0.022 | 0.0015 | 0.747 | 0.021 | 0.020 | 0.582 | 0.236 | 4692.9 | 4603.90 |
| Cytokinin × Cultivar | 2 | 0.031 | 0.012 | 0.014 | 0.076 | 0.0002 | 0.0002 | 0.00006 | 0.026 | 0.0003 | 0.0002 | 0.067 | 0.009 | 159.72 | 166.68 |
| Irriga × Cytoki × Cultiv | 2 | 1.336 | 0.005 | 0.012 | 0.490 | 0.015 | 0.017 | 0.0008 | 0.466 | 0.016 | 0.017 | 0.471 | 0.143 | 3615.4 | 3367.9 |
| Error | 15 | 1.620 | 0.018 | 0.016 | 0.618 | 0.012 | 0.012 | 0.0009 | 0.451 | 0.012 | 0.012 | 0.597 | 0.146 | 3189.3 | 3020.1 |
| CV% | 11.611 | 2.224 | 2.032 | 5.840 | 3.891 | 3.768 | 3.497 | 6.900 | 3.668 | 3.558 | 6.044 | 3.489 | 12.840 | 12.975 |
| Source | DF | TGW | Major | Feret | Area | Minor | MinFeret | Area/Perim. | Area × Circ. | Minor/Solid. | MinFeret/Solid. | Area × Solid | Perim. × Circ. | A1 | A2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rep | 2 | 0.093 | 0.008 | 0.009 | 0.112 | 0.006 | 0.006 | 0.0003 | 0.156 | 0.006 | 0.006 | 0.112 | 0.039 | 1702.34 | 1594.910 |
| Irrigation | 1 | 43.670 *** | 0.054 | 0.073 | 12.360 *** | 0.393 *** | 0.395 *** | 0.024 *** | 12.745 *** | 0.387 *** | 0.387 *** | 12.296 *** | 3.848 *** | 96,923.93 *** | 92,400.13 *** |
| Irrigation × rep | 2 | 0.487 | 0.003 | 0.007 | 0.049 | 0.004 | 0.003 | 0.0002 | 0.107 | 0.003 | 0.003 | 0.056 | 0.034 | 596.44 | 602.82 |
| Cytokinin | 1 | 2.805 | 0.015 | 0.021 | 0.402 | 0.004 | 0.006 | 0.0004 | 0.234 | 0.004 | 0.005 | 0.402 | 0.066 | 2493.64 | 2165.76 |
| Irrigation × Cytokinin | 1 | 0.899 | 0.017 | 0.025 | 0.260 | 0.002 | 0.002 | 0.0002 | 0.102 | 0.002 | 0.003 | 0.243 | 0.030 | 1146.26 | 988.48 |
| Cytokin × rep (Irrigat) | 4 | 1.843 | 0.027 | 0.029 | 0.656 | 0.010 | 0.011 | 0.0008 | 0.438 | 0.010 | 0.010 | 0.629 | 0.127 | 3917.28 | 3617.09 |
| Cultivar | 2 | 13.276 *** | 0.851 *** | 0.680 *** | 8.955 *** | 0.036 * | 0.037 * | 0.007 *** | 3.51 8*** | 0.036 * | 0.038 * | 8.294 *** | 1.101 *** | 29,504.88 *** | 27,244.93 *** |
| Irrigation × Cultivar | 2 | 0.037 | 0.007 | 0.006 | 0.205 | 0.003 | 0.003 | 0.0003 | 0.165 | 0.003 | 0.002 | 0.210 | 0.054 | 1116.59 | 1101.68 |
| Cytokinin × Cultivar | 2 | 1.015 | 0.008 | 0.009 | 0.333 | 0.006 | 0.007 | 0.0005 | 0.245 | 0.006 | 0.007 | 0.322 | 0.078 | 1932.01 | 1760.01 |
| Irriga × Cytoki × Cultiv | 2 | 0.004 | 0.011 | 0.012 | 0.144 | 0.0008 | 0.0008 | 0.0001 | 0.055 | 0.0009 | 0.001 | 0.126 | 0.017 | 573.25 | 535.41 |
| Error | 16 | 1.236 | 0.015 | 0.017 | 0.489 | 0.009 | 0.009 | 0.0006 | 0.351 | 0.009 | 0.009 | 0.477 | 0.109 | 2839.39 | 2673.09 |
| CV% | 7.921 | 2.025 | 2.114 | 4.740 | 3.123 | 3.033 | 2.814 | 5.336 | 2.949 | 2.860 | 4.896 | 2.814 | 9.597 | 9.642 |
| First Growing Season | Second Growing Season | |||||
|---|---|---|---|---|---|---|
| Well-Watered | Terminal Water Stress | Relative Change % | Well-Watered | Terminal Water Stress | Relative Change % | |
| TGW (g) | 31.127 a | 23.462 b | −24.62 | 37.854 a | 32.340 b | −14.55 |
| Major (mm) | 6.115 a | 5.955 a | −2.62 | 6.172 a | 6.094 a | −1.26 |
| Feret (mm) | 6.310 a | 6.115 a | −3.08 | 6.298 a | 6.207 a | −1.43 |
| Area (mm2) | 14.396 a | 12.481 b | −13.30 | 15.335 a | 14.164 b | −7.64 |
| Minor (mm) | 2.974 a | 2.649 b | −10.91 | 3.139 a | 2.930 b | −6.65 |
| MinFeret (mm) | 3.088 a | 2.757 b | −10.70 | 3.250 a | 3.041 b | −6.44 |
| Area/Perim (mm) | 0.913 a | 0.831 b | −9.01 | 0.959 a | 0.907 b | −5.42 |
| Area × Circ. (mm) | 10.621 a | 8.783 b | −17.30 | 11.705 a | 10.514 b | −10.17 |
| Minor/Solid. (mm) | 3.124 a | 2.807 b | −10.26 | 3.278 a | 3.071 b | −6.32 |
| MinFeret/Solid. (mm) | 3.243 a | 2.918 b | −10.01 | 3.394 a | 3.187 b | −6.11 |
| Area × Solid (mm) | 13.708 a | 11.797 b | −13.94 | 14.690 a | 13.521 b | −7.96 |
| Perim. × Circ. (mm) | 11.478 a | 10.436 b | −9.07 | 12.057 a | 11.403 b | −5.42 |
| A1 (mm) | 514.78 a | 360.41 b | −29.99 | 607.133 a | 503.357 b | −17.09 |
| A2 (mm) | 496.32 a | 346.44 b | −30.20 | 586.868 a | 485.543 b | −17.27 |
| First Growing Season | Second Growing Season | |||||
|---|---|---|---|---|---|---|
| Sirvan | Pishgam | Torabi | Sirvan | Pishgam | Torabi | |
| TGW (g) | 25.700 b | 27.110 ab | 29.600 a | 37.500 a | 32.275 b | 35.475 a |
| Major (mm) | 5.940 b | 5.901 b | 6.292 a | 6.291 a | 5.825 b | 6.282 a |
| Feret (mm) | 6.066 b | 6.110 b | 6.494 a | 6.346 a | 5.982 b | 6.429 a |
| Area (mm2) | 12.912 b | 13.249 b | 14.306 a | 15.371 a | 13.763 b | 15.114 a |
| Minor (mm) | 2.748 b | 2.834 ab | 2.871 a | 3.088 a | 2.979 b | 3.038 ab |
| MinFeret (mm) | 2.845 b | 2.938 ab | 3.004 a | 3.187 a | 3.083 b | 3.167 ab |
| Area/Perim (mm) | 0.855 b | 0.872 ab | 0.893 a | 0.954 a | 0.907 b | 0.938 a |
| Area × Circ. (mm) | 9.305 b | 9.728 ab | 10.191 a | 11.585 a | 10.520 b | 11.223 a |
| Minor/Solid. (mm) | 2.900 b | 2.982 ab | 3.028 a | 3.226 a | 3.117 b | 3.181 ab |
| MinFeret/Solid. (mm) | 3.004 b | 3.092 ab | 3.168 a | 3.330 a | 3.226 b | 3.316 ab |
| Area × Solid (mm) | 12.597 b | 12.597 b | 13.573 a | 14.716 a | 13.160 b | 14.443 a |
| Perim. × Circ. (mm) | 10.744 b | 10.963 ab | 11.231 a | 11.996 a | 11.400 b | 11.793 a |
| A1 (mm) | 396.140 b | 436.980 b | 490.490 a | 592.800 a | 499.040 b | 573.890 a |
| A2 (mm) | 382.820 b | 422.170 ab | 469.390 a | 574.720 a | 482.920 b | 550.970 a |
| Indices | Overall | 1st Year | 2nd Year | WW (2 Years) | TWS (2 Years) | 1st Year WW | 1st Year TWS | 2nd Year WW | 2nd Year TWS |
|---|---|---|---|---|---|---|---|---|---|
| Major | 0.654 | 0.710 | 0.710 | 0.626 | 0.652 | 0.671 | 0.734 | 0.846 | 0.769 |
| Feret | 0.590 | 0.704 | 0.704 | 0.531 | 0.540 | 0.708 | 0.616 | 0.830 | 0.714 |
| Area | 0.955 | 0.948 | 0.948 | 0.917 | 0.951 | 0.953 | 0.923 | 0.927 | 0.943 |
| Minor | 0.965 | 0.943 | 0.943 | 0.964 | 0.969 | 0.948 | 0.925 | 0.946 | 0.901 |
| MinFeret | 0.964 | 0.944 | 0.944 | 0.962 | 0.966 | 0.965 | 0.909 | 0.927 | 0.908 |
| Area/Perim | 0.981 | 0.973 | 0.973 | 0.979 | 0.982 | 0.985 | 0.954 | 0.961 | 0.959 |
| Area × Circ. | 0.983 | 0.973 | 0.973 | 0.980 | 0.982 | 0.988 | 0.954 | 0.959 | 0.956 |
| Minor/Solid. | 0.962 | 0.943 | 0.943 | 0.961 | 0.967 | 0.951 | 0.924 | 0.944 | 0.902 |
| MinFeret/Solid | 0.959 | 0.942 | 0.942 | 0.957 | 0.961 | 0.960 | 0.904 | 0.922 | 0.904 |
| Area × Solid | 0.962 | 0.953 | 0.953 | 0.931 | 0.959 | 0.963 | 0.929 | 0.931 | 0.949 |
| Perim. × Circ. | 0.981 | 0.973 | 0.973 | 0.979 | 0.982 | 0.985 | 0.955 | 0.961 | 0.959 |
| A1 | 0.978 | 0.966 | 0.966 | 0.965 | 0.976 | 0.984 | 0.933 | 0.939 | 0.947 |
| A2 | 0.979 | 0.969 | 0.969 | 0.969 | 0.978 | 0.987 | 0.940 | 0.945 | 0.948 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamani, A.; Emam, Y.; Edalat, M. Response of Bread Wheat Cultivars to Terminal Water Stress and Cytokinin Application from a Grain Phenotyping Perspective. Agronomy 2024, 14, 182. https://doi.org/10.3390/agronomy14010182
Zamani A, Emam Y, Edalat M. Response of Bread Wheat Cultivars to Terminal Water Stress and Cytokinin Application from a Grain Phenotyping Perspective. Agronomy. 2024; 14(1):182. https://doi.org/10.3390/agronomy14010182
Chicago/Turabian StyleZamani, Afshin, Yahya Emam, and Mohsen Edalat. 2024. "Response of Bread Wheat Cultivars to Terminal Water Stress and Cytokinin Application from a Grain Phenotyping Perspective" Agronomy 14, no. 1: 182. https://doi.org/10.3390/agronomy14010182
APA StyleZamani, A., Emam, Y., & Edalat, M. (2024). Response of Bread Wheat Cultivars to Terminal Water Stress and Cytokinin Application from a Grain Phenotyping Perspective. Agronomy, 14(1), 182. https://doi.org/10.3390/agronomy14010182