Abstract
Free amino acids in cacao beans are important precursors to the aroma and flavor of chocolate. In this research, we used inferential and explanatory statistical techniques to verify the effect of different edaphic crop conditions on the free amino acid profile of PH-16 dry cacao beans. The decreasing order of free amino acids in PH-16 dry cacao beans is leucine, phenylalanine, glutamic acid, alanine, asparagine, tyrosine, gamma-aminobutyric acid, valine, isoleucine, glutamine, lysine, aspartic acid, serine, tryptophan, threonine, glycine. With the exception of lysine, no other free amino acid showed a significant difference between means of different edaphic conditions under the ANOVA F-test. The hydrophobic free amino acids provided the largest contribution to the explained variance with 58.01% of the first dimension of the principal component analysis. Glutamic acid stands out in the second dimension with 13.09%. Due to the stability of the biochemical profile of free amino acids in this clonal variety, it is recommended that cacao producers consider the genotype as the primary source of variation in the quality of cacao beans and ultimately the chocolate to be produced.
1. Introduction
The amino acid profile of cacao beans is one of the most important biochemical characteristics related to the cacao quality [1,2,3,4], mainly because of its direct influence on the taste and aroma of the by-products of the processed beans, such as chocolate [3,5,6,7]. The amino acid profile in cacao beans is influenced by local or regional crop conditions, in addition to the contribution of fermentation and drying processes [7]. The conditions of growth and fermentation of cacao may interfere with the average weight of the beans, and polyphenols and amino acid concentrations [8]. In Bahia, Brazil, in the same edaphoclimatic conditions and post-harvest processing, the contents and profile of amino acids and bioactive amines differ in relation to the clonal varieties of cacao, resulting in products with different nutritional, functional, safety and sensory attributes [3]. However, the effect of multiple edaphic crop conditions, on the content and nature of amino acids of the cacao beans of the genetic varieties reported, remains unknown.
The most common amino acids found in cacao beans are: aspartic acid, asparagine, glutamine, glutamic acid, alanine, isoleucine, leucine, phenylalanine, tyrosine, valine, gamma-aminobutyric acid (GABA), glycine, lysine, serine, threonine [6,7,9,10] and, in cacao science are classified into acidic, hydrophobic and other [7,10,11,12]. In cacao beans, the GABA non-protein amino acid is also found which is a neurotransmitter that has recently attracted some interest due to its health effects as a hypotensive, diuretic, tranquilizer and diabetes-preventing agent [9]. Alanine, asparagine, aspartate, glutamic acid and serine are non-essential amino acids that are negligible in human diets [13]. The other amino acids are classified as essential, and must be added to the diet through food intake [13].
The profile of free amino acids and their distribution are the result of cacao fermentation practices [10,11,14]. Although the total content of free amino acids in cacao samples is variable, the most representative are the hydrophobic amino acids (leucine, isoleucine, phenylalanine, alanine, valine) and, by and large, leucine is the most abundant amino acid, whereas glycine, histidine and methionine are scarcer [9]. Generally, fermented cacao can be differentiated from unfermented cacao beans by the amount of free amino acids and the ratio of hydrophobic free amino acids to acids [7]. Unfermented cacao beans have few total free amino acids, most of which are categorized as acidic amino acids [7]. Meanwhile, fermented beans have a higher concentration of total free amino acids, with a reduction in acid free amino acids and an increase in hydrophobic free amino acids [7]. Glutamic acid, leucine, phenylalanine and serine are predominant in unfermented cacao beans [14]. Hydrophobic amino acids are predominant at the end of fermentation [12,14]. The hydrophobic amino acids are important precursors of the cacao aroma [9]. Fermentation improves the amino acid profile of the cacao nibs, particularly the essential amino acids lysine, histidine, arginine, threonine, valine, isoleucine and phenylalanine [5]. In the unfermented cacao nibs, glutamic acid was the most abundant amino acid whilst the most concentrated essential amino acid in the same sample was leucine. As for the fermented cacao nibs, a similar trend was observed, with respective values of glutamic acid and leucine [5]. Regarding cacao bean samples from different geographical origins, glutamic acid is present in medium–low concentrations, probably because it partially undergoes conversion to GABA and there is little information about the concentration in cacao of the non-protein amino acid resulting from that conversion in the literature [9]. Cacao beans can be considered an important natural source of the inhibitory neurotransmitter amino acid GABA, even comparable with contents recorded in some enriched foods [9].
Cacao was introduced to the state of Bahia from the Amazon region in 1746, expanding commercially from 1820 and reaching its maximum commercial production in the 1980s [15]. Primarily, the edaphoclimatic conditions made possible the persistence of cacao cultivation in this region [16,17]. In addition to post-harvest management and processing of cacao beans [4], the quality of soils cultivated with cacao reflects different edaphic conditions through different physical and chemical attributes [18]. In cacao beans, the diverse edaphic conditions result in variations in the mineral composition [19] and biochemical attributes [20]. Proteins, total free amino acids, simple carbohydrates (sucrose, fructose and glucose), total acidity, cadmium and barium are the most variable attributes of cacao quality, when edaphic conditions of cultivation are changed [1]. At the regional level, different clonal varieties of cacao, such as PH-16, have been selected, mainly because of their tolerance to the witches’ broom plant (Moniliophthora perniciosa), which one of the major fungal diseases [21]. For this reason, it is necessary to characterize these genotypes in terms of the quality of the chocolate produced [3,22]. The PH-16 cacao clone shows characteristics of medium size, erectness, self-compatible architecture and high precocity of production and productivity [23]. The cacao pods are purplish red and medium in size [23]. The biometric attributes of the cacao pods and beans (purple) are influenced by different crop sites [24]. The PH-16 cacao clone originated from a selection made in a commercial Forastero and Trinitarian group from the “Porto Híbrido” farm in the county of São José da Vitória, Bahia, Brazil [22]. PH-16 cacao has a high nutritional demand similar to other hybrids generated from crossing with Trinitarian plants [25]. PH-16 cacao shows a versatile adaptability to different climatic conditions, being found in commercial crops from humid to semi-arid zones [23,26]. This clonal variety displayed good production responses in semi-mechanized systems with chemical weeding control, sprayed pesticides and pruning, as well as the use of windbreaks and drip irrigation [26]. Several attributes of cacao quality were analysed in samples from the PH-16 clone, including the total amount of free amino acids [1]. The decreasing order of the levels of free amino acids detected in PH-16 dry cacao beans [3] was: leucine, alanine, phenylalanine, arginine, tyrosine, valine, lysine, isoleucine, threonine, proline, aspartic acid, serine, histidine, glycine, glutamine, tryptophan, glutamic acid. Methionine was not detected in PH-16 [3]. Due to their composition of free amino acids, PH-16 dry cacao beans tend to produce bitter and sweeter chocolate [3].
Soil is a good environmental stratifier and the basis for world agricultural production [27]. However, differentiating edaphic conditions from crop sites is a complex task due to the large number of attributes that make up the soil ecosystem. The Soil Quality Index (SQI) for cacao agroecosystems involves the strategy of configuring the different edaphic conditions through the standardization of indicators whose weighted results reflect the needs of the crop [18]. The multivariate solution of the SQI changes in its function with its respective indicators, because each indicator is part of a specific context of plant production, facilitating its interpretation. For example, due to their characteristics, available water capacity, bulk density, total porosity and organic matter are indicators of the “available water” function [18]. In this way, the edaphic condition of available water can be easily identified at different crop sites.
As a result of the relevance of free amino acids to the quality of cacao beans, the main objective of this work was to analyse the effect of different edaphic crop conditions on the free amino acid profile of PH-16 cacao beans. The secondary objectives were the environmental stratification through the different edaphic conditions represented by the SQI and the determination of the content of free amino acids in cacao beans from the samples of the different edaphic conditions of the crop sites.
2. Materials and Methods
2.1. Subsection
The fruit samples of PH-16 cacao were obtained from 12 crop sites in the cacao region of Bahia (Figure 1). According to the Superintendence of Economic and Social Studies of Bahia [28], this region is classified as a wetland according to the climatic types of Thornthwaite: B4r A’, B3r A’, B2r A’, B2r B’, B1r A’, B1r’ A’, B1w A’. The southeastern zone of Bahia is characterized by an average rainfall greater than 2000 mm year−1, with a water deficit from 0 to 345 mm, and a water surplus of 200 to 1409 mm year−1 [28]. The 12 crop sites with their respective geographic coordinates and municipalities, have different types of soils [18], according to the Brazilian Soil Classification System (SiBCS) [29] (Figure 1).
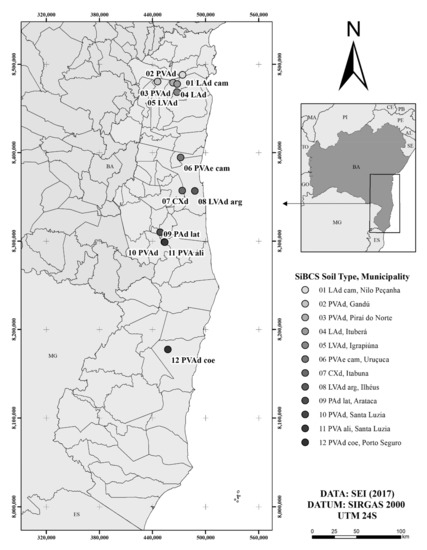
Figure 1.
Cacao crop sites represented by different edaphic conditions in the humid zone of southeast Bahia, Brazil: ambisolic Dystrophic Yellow Latosol (01 LAd cam); typic Dystrophic Red–Yellow Argisol (02 PVAd); abrupt Dystrophic Red–Yellow Argisol (03 PVAd); typic Dystrophic Yellow Latosol (04 LAd); typic Dystrophic Red–Yellow Latosol (05 LVAd); cambisolic Eutrophic Red–Yellow Argisol (06 PVAe cam); typic Dystrophic Haplic Cambisol (07 CXd); argisolic Dystrophic Red–Yellow Latosol (08 LVAd arg); latosolic Dystrophic Yellow Argisol (09 PAd lat); typic Dystrophic Red–Yellow Argisol (10 PVAd); typic Alitic Red–Yellow Argisol (11 PVA ali); abrupt Cohesive Dystrophic Red–Yellow Argisol (12 PVAd coe).
Each crop site, covering approximately one hectare, was subdivided into three collection areas, characterized by the same soil type [1]. Cacao sampling was conducted in November 2008, because it was representative of the second harvest period (August 2008 to January 2009).
2.2. Soil Quality Index and Functions
In this investigation, the standard method [18] was used to calculate the SQI and its functions. The SQI data used in this research came from the original study [18] with the full authorization of the authors. For greater analytical security, the values were recalculated.
2.3. Post-Harvest Processing
Fifty mature cacao pods made up each sample (three per crop site) for the post-harvest processing of fermentation and drying [24,25].
2.4. High-Performance Liquid Chromatography for the Determination of Free Amino Acids
The following steps of the method for determining free amino acids in cacao beans were adapted from the reference study with high-performance liquid chromatography (HPLC) [7].
2.4.1. Material Analyzed
For the chromatographic analysis, 300 g of dry cacao cotyledons (vegetal material) were used.
2.4.2. Pre-Analytical Physical Treatments
The dry cacao cotyledon samples were frozen for at least 24 h at −20 °C and ground slowly by an ultracentrifugal mill (ZU 100, Comp. Retsch, without sieve, Crown 12er) at 15,000 rpm. The dry cacao cotyledons were sieved through a 3 mm diameter sieve. The residue of material was crushed and sieved again until 99% was less than 3 mm in diameter. A stainless-steel ball homogenizer was used to grind this vegetal material.
2.4.3. Pre-Analytical Chemical Treatments
In a centrifuge tube with petroleum ether, the ground dry cacao cotyledons were defatted by multiple cold extraction. Then, the water content of the defatted ground cacao powder was measured according to Methodenbuch Band III [30].
The contents of free amino acids were analysed according to the method described by Kirchhoff et al. [11] with the following modifications: 0.5 g of defatted ground cacao powder was stirred at temperature less than 4 °C for 1 h with 1.4 g of polyvinylpolypyrrolidone (PVPP) and 50 mL of double distilled water. The pH value was adjusted to pH 2.5 with 50% aqueous trifluoroacetic acid solution, and the homogenate solution was centrifuged for 10 min at 5000 rpm [7]. Clear supernatant solution was filtered through a 0.45 µm filter membrane (Multoclear, CS-Chromatographie). Thirty microliters of each sample was lyophilized (1 h, −20 °C, 0.05 mbar) directly in the flask, where it was kept at −20 °C until analysis.
2.4.4. Separation, Identification and Quantitative Determination of Free Amino Acids
Free amino acids of dry cacao cotyledons were separated by reversed-phase HPLC equipment, then converted into ophthaldehyde derivatives (OPA).
The HPLC analytical conditions of the OPA derivatization were as follows [7]: separating column: LicChroCART 250-4 (Merck, Darmstadt, Germany). Pre-column: LiChrospher® 100 RP-18 (5 µm) (Merck, VWR international). Column temperature: 30 °C. Eluent A: 1.6 l sodium acetate solution/glacial acetic acid (50 mmol L−1; pH 6.2), 50 mL of MeOH [LiChrosolv® (Merck, Darmstadt, Germany), gradient grade], 20 mL of tetrahydrofuran [LiChrosolv® (Merck, Darmstadt, Germany), gradient grade]. Eluent B: 200 mL of sodium acetate/glacial acetic acid solution (50 mmol L−1; pH 6.2), 800 mL of MeOH [LiChrosolv® (Merck, Darmstadt, Germany), gradient grade], flow rate of 1.3 mL min−1. A column was balanced as Eluent A. Gradient (A + B = 100% v/v): (1) 2 min 100–95% A, (2) 10 min 95–80% A, (3) 8 min 80–70% A, (4) 5 min 62–50% A, (5) 15 min 50–0% A, (6) 10 min constant 0% A, (7) 5 min 0–100% A, (8) 20 min constant 100% A. Detector: Hitachi F-1050 fluorescence spectrophotometer detector (λex = 334 nm, λem = 425 nm). Autosampler: Merck-Hitachi AS-4000 automatic display. Separation HPLC measurement: 800 μL borate buffer (200 mmol L−1 boric acid; adjusted to pH 9.5 with concentrated KOH, boiled for 5 min) were added to the lyophilized samples. Standard solutions: 1–10 pmol μL−1 of each amino acid, dissolved in borate buffer. Subsequently, 400 μL OPA-reagent was added to these solutions prior to each injection. Preparation of OPA reagent (24 h before being used): 100 mg ophthaldialdehyde (Merck, No. 11452) previously solubilized in 2.5 mL MeOH [LiChrosolv® (Merck, Darmstadt, Germany), gradient grade] plus 22.4 mL borate buffer (pH 9.5) plus 100 μL 2-mercaptoethanol (Merck, No. 15433). After injection of 20 μL of this mixture, the derivatization was stopped after exactly after 2 min by passing the eluent into the column. Solution degasser: Degassex DG-4400, Phenomenex. Quantification: calculated via peak area of chromatograms from standard mixtures containing 1–10 pmol μL−1 of each amino acid.
2.4.5. Statistical Analysis
The experimental design was completely randomized, with 12 crop sites characterized by different soil types, with three replications in each site. The statistical procedures used in this study were performed using the R software [31]. The missing values were imputed into the dataset with the PCA model using the imputed PCA function of the package “missMDA” [32]. The PCA was carried out with the “FactoMineR” package [33] and graphical analysis with the “ggplot2” package [34].
3. Results
The cacao crop sites represented different edaphic conditions in the southeast of Bahia, Brazil (Figure 1, Table 1). In order to detect significant statistical differences between averages at 5% error, the SQI values and their functions were subjected to an ANOVA F-test (Table 1). Through the Scott–Knott test, four SQI average groups were formed (Table 1). The environmental safety function did not correlate with the SQI (r = 0.05) but, on the other hand, the available water (r = 0.68), root growth (r = 0.88) and mineral nutrition of plant (r = 0.77) functions are strongly associated with the SQI scores (Table 1). Available water, root growth and mineral nutrition of plants (Table 1) were the functions with the greatest contribution to the group with the highest scores of the SQI: typic Alitic Red–Yellow Argisol (0.72 ± 0.08), typic Dystrophic Red–Yellow Argisol (0.67 ± 0.09), argisolic Dystrophic Red–Yellow Latosol (0.62 ± 0.08). On the other hand, the abrupt Cohesive Dystrophic Red–Yellow Argisol (Table 1) had the lowest score (0.29). Table 2 shows soil agricultural management in the crop sites.

Table 1.
Edaphic crop conditions of the PH-16 cacao clon in the humid zone of southeast Bahia, Brazil represented by the Soil Quality Index (SQI) and its functions.
Table 2.
Soil management of the 12 crop sites for the PH-16 cacao clone in the humid region of southeast Bahia: correction of soil acidity, mineral fertilization and irrigation.
Table 3 and Table 4 show the contents of the free amino acids in dry cacao beans from cacao trees grown in different soil edaphic conditions (Table 1 and Table 2). By the ANOVA F-test, no significant difference was detected at 5% error between the averages of the asparagine, aspartic acid, glutamine or glutamic (acidic free amino acid) contents in PH-16 dry cacao beans in the different edaphic crop conditions (Table 3). There was also no significant effect of different edaphic crop conditions on the average contents of alanine, isoleucine, leucine, phenylalanine, tyrosine or valine (hydrophobic amino acids) in dry cacao beans (Table 3). There was no effect of edaphic crop conditions on average GABA contents in dry cacao beans (Table 4). The average contents of glycine, serine, threonine and tryptophan in the dry cacao beans did not vary in relation to the different edaphic crop conditions (Table 4). Only the average content of lysine in the dry beans varied in relation to the different edaphic conditions of the cacao crop (Table 4). Two groups of average lysine contents were formed by the Scott–Knott test at 5% error (Table 4), and the highest group corresponded to the cambisolic Eutrophic Red–Yellow Argisol (PVAe cam, 1204.92 ± 618.90) and typic Dystrophic Haplic Cambisol soils (CXd, 922.80 ± 153.29). According to the SQI interpretation scale proposed by Araujo et al. (2018), the average score of the CXd is classified as “bad” and PVAe as “regular”.
Table 3.
Effect of different edaphic crop conditions in the humid zone of southeast Bahia, Brazil on acidic and hydrophobic free amino acids of PH-16 dry cacao beans.
Table 4.
Effect of different edaphic crop conditions in the humid zone of southeast Bahia, Brazil on non-protein and other free amino acids of PH-16 dry cacao beans.
Table 3 and Table 4 show the contents of the free amino acids in dry cacao beans found in different soil conditions (Table 1 and Table 2). By the ANOVA F-test, no significant difference was detected at 5% error between the averages of the asparagine, aspartic acid, glutamine or glutamic (acidic free amino acid) contents in PH-16 dry cacao beans in the different edaphic crop conditions (Table 3). There was also no significant effect of different edaphic crop conditions in the average contents of alanine, isoleucine, leucine, phenylalanine, tyrosine or valine (hydrophobic amino acids) in dry cacao beans (Table 3). There was no effect of edaphic crop conditions on average GABA contents in dry cacao beans (Table 4). The average contents of glycine, serine, threonine and tryptophan in the dry cacao beans did not vary in relation to the different edaphic crop conditions (Table 4).
Only the average content of lysine in the dry beans varied in relation to the different edaphic conditions of the cacao crop (Table 4). Two groups of average lysine contents were formed by the Scott–Knott test at 5% error (Table 4), and the highest group corresponded to the cambisolic Eutrophic Red–Yellow Argisol (PVAe cam, 1204.92 ± 618.90) and typic Dystrophic Haplic Cambisol soils (CXd, 922.80 ± 153.29). According to the SQI interpretation scale proposed by Araujo et al. (2018), the average score of the CXd is classified as “bad” and PVAe as “regular”.
The specific information of the edaphic conditions provided by the SQI and its functions (Table 1) are not linearly associated with the contents of free amino acids in dry cacao beans (Table 5). The weak correlations of lysine and tyrosine with the available water function could not be confirmed by graphical analysis (Figure 2).
Table 5.
Pearson (r) linear correlations between free amino acids in PH-16 dry cacao beans and Soil Quality Index (SQI) and its functions (0–15 cm) in 12 crop sites in the humid zone of southeast Bahia, Brazil.
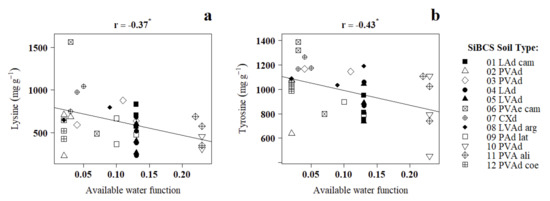
Figure 2.
Pearson’s linear correlations (r) between the free amino acids lysine and glutamine, in PH-16 dry cacao beans, and the functions of the Soil Quality Index available water (a) and environmental safety (b) calculated for 12 crop sites in the southeast of Bahia. Significant at 5% error (*).
According to the general average content of the samples (Table 3 and Table 4), the free amino acids of PH-16 dry cacao beans were classified in descending order: leucine, phenylalanine, glutamic acid, alanine, asparagine, tyrosine, gamma-aminobutyric acid, valine, isoleucine, glutamine, lysine, aspartic acid, serine, tryptophan, threonine, glycine. As the contents of the free amino acids were determined in the same subsamples of the same cacao genotype, high collinearity between them is expected (Table 6). However, the variation between samples (Table 3 and Table 4) affects the order of classification from highest to lowest content for each free amino acid in each subsample, reflecting non-significant correlations between some of the free amino acids in dry cacao beans (Table 6).
Table 6.
Pearson’s linear correlations (r) between free amino acids in PH-16 dry cacao beans in 12 crop sites in the humid zone of southeast Bahia, Brazil.
The exploratory analysis of the data was performed with the PCA technique (Table 7; Figure 3). The data were reduced to two dimensions that represent 71.09% (Table 7, Figure 3) of the cumulative variance and were: hydrophobic free amino acids (PC1) and glutamic acid (PC2). The hydrophobic free amino acids in dry cacao beans were strongly correlated with PC1 (Table 7). On the other hand, the most correlated free amino acid with PC2 was glutamic acid (Table 7), which is the third most abundant amino acid in samples of PH-16 dry cacao beans (Table 3).
Table 7.
Summary of principal component analysis applied to the free amino acids of PH-16 dry cacao beans correlated with samples represented by Scott–Knott average groups for the Soil Quality Index.
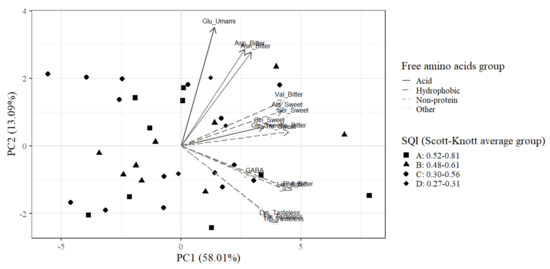
Figure 3.
Biplot of principal component analysis (PCA) applied to the free amino acids of PH-16 dry cacao beans correlated with samples represented by Scott-Knott average groups (decreasing order A, B, C and D) for the Soil Quality Index (SQI): asparagine (Asn), aspartic acid (Asp), glutamine (Gln), glutamic acid (Glu), alanine (Ala), isoleucine (Ile), leucine (Leu), phenylalanine (Phe), tyrosine (Tyr), valine (Val), gamma-aminobutyric acid (GABA), glycine (Gly), lysine (Lys), serine (Ser), threonine (Thr), tryptophan (Trp). Taste classification: bitter, sweet, tasteless, umami. Thirty-six sample observations.
The absence of significant differences between the average contents of free amino acids in the ANOVA (Table 3 and Table 4) is reflected in the multivariate analysis of the PCA biplot (Figure 3). There is no distribution pattern of observations in the sample (individuals) represented by different sets of Scott–Knott averages for the SQI. The internal variability of the SQI values for each sample observation does not show any discriminating relationship with the free amino acid contents (Figure 3). In the biplot of the PCA (Figure 3), asparagine and aspartic acid are strongly correlated with each other, and more correlated with glutamic acid compared to the other free amino acids. The hydrophobic free amino acids leucine and phenylalanine and the non-protein GABA are strongly correlated (Figure 3). Leucine and phenylalanine are the most abundant free amino acids in PH-16 dry cacao beans (Table 3). Lysine, tyrosine and tryptophan were strongly correlated with each other, and are closer to leucine, phenylalanine and GABA compared to the other free amino acids (Figure 3). Very weak correlations were verified between free amino acids found at the ends of the PCA biplot (Figure 3). GABA, glycine and glutamine have the lowest contributions to the explained variation in the PCA biplot (Figure 3).
Valine, alanine, serine, glycine, glutamine, threonine and isoleucine form a group of variables highly correlated with each other and with weak or very weak correlation with the other free amino acids (Figure 3). Valine, alanine and serine show similar contributions to the explained variance and are strongly correlated (Figure 3). Glycine and glutamine are strongly associated with each other (Figure 3). Glycine is the least concentrated free amino acid in the PH-16 dry cacao beans (Table 3). Threonine and isoleucine are strongly correlated (Figure 3).
4. Discussion
The average groups formed by Scott-Knott test after the ANOVA F-test (Table 1) indicate at least four different edaphic conditions that may or may not favourable for a cacao crop. These conditions (Table 1) do not reflect only the intrinsic characteristics of each type of soil, such as natural fertility, because the edaphic environment is also influenced by soil agricultural management (Table 2).
In general, soil management in cultivated areas with the PH-16 clone was quite irregular (Table 2). The lack of more accurate information (Table 2) is due to the fact that not all rural properties had adequate administrative control over the agricultural inputs applied in the cacao crop fields. Since the soils in the humid region of southeastern Bahia are predominantly acidic [16], the practice of liming to stabilize the acidity of the soils is a common practice found in seven out of 12 cacao crop sites, in addition to N–P–K fertilization (Table 2). Plaster application and fertigation are the least common soil management practices in the cacao humid region, because of climatic conditions such as high rainfall and rugged topography (Table 2). Albeit the information on soil management is incomplete, the favourable edaphic condition of the crop sites that showed the highest SQI scores is undoubtedly related to the correction of soil acidity and mineral fertilization with macronutrients (Table 2). The PH-16 cacao genotype [23,25] requires semi-intensive or intensive soil management to achieve high productivity, based on previous information.
Arginine, histidine and proline were identified in chocolate samples made with PH-16 dry cacao beans [14], however, they were not detected in the dry bean samples of the present research. When comparing the content of free amino acids between chocolate samples and PH-16 dry cacao beans, the difference is from two to 15 times lower (Table 3 and Table 4). Despite its content reduction of approximately four times in chocolate samples [14], leucine is the most abundant free amino acid. Incomplete sample observations (missing values) of glutamine and tyrosine (Table 3) and glycine and lysine (Table 5) were not deleted from the statistical analyses.
Cacao bean productivity (Table 2) is reflected in the SQI classifications. In addition to the cambisolic character that appears in the two types of soil where samples of dry cacao beans with higher lysin contents were found (Table 1 and Table 4), the averages of the functions of available water, root growth, mineral nutrition of plants and environmental safety are in the same groups (Table 2), indicating similar edaphic conditions.
Hydrophobic free amino acids are the most representative group in cacao samples [9]. Leucine, valine, alanine, isoleucine and phenylalanine contribute to the formation of the typical cacao aroma components in the presence of reducing sugar upon roasting [36]. Chocolate’s sweet taste is related to alanine, a free amino acid whose content is eight times higher in chocolate samples [14] compared to dry cacao bean samples (Table 3).
Besides being the third most abundant free amino acid in PH-16 dry cacao beans (Table 3), glutamic acid belongs to a different multivariate dimension (Table 7). It is possible that this is a characteristic of genotypes associated with the Forastero group. Glutamic acid was the most abundant free amino acid in the unfermented and fermented cacao nibs from Nigeria [5]. The glutamic acid content remains somewhat stable even after fermentation and drying of the cacao beans [6]. Both aspartic and glutamic acids are related to a sour and astringent taste of dry cacao beans [35]. Umami is the fifth basic taste, after sweet, sour, salty and bitter, which is associated with glutamate, the sodium salt of glutamic acid, one of the most essential non-essential amino acids in nature [37]. Umami taste is only possible with protein hydrolysis favoured by fermentation, aging, ripening and heating processes that release glutamate [37,38].
The SQI values could not discriminate the samples in relation to the free amino acid contents (Figure 3), which implies that the effect of different edaphic conditions on the free amino acid profile of dry PH-16 cacao beans was not detected. However, the different relationships between free amino acids shown in PCA biplot (Figure 3) are an important biochemical characterization of the profile of PH-16 dry cacao beans.
GABA is a neurotransmitter that has been extensively studied for its effects on human health [9], and represents a relatively large contribution to the total content of free amino acids in dry cacao beans, of about 7% (Table 4), making them a rich source of GABA.
In addition, lysine is one of the essential free amino acids that shows an increase in content after fermentation [5], and it was also the only one that had variation in its content in dry cacao beans in relation to crop site (Figure 3). The fermentation process of this study was completely standardized [25], however, lysine also presented missing values in the analytical determination (Table 4). It is possible that the pH and acidity of the cacao mucilage [25] influenced the differences observed for lysine in dry cacao beans and are not a direct result of the edaphic systems.
Furthermore, valine, alanine, serine, glutamine, glycine and threonine are free amino acids associated with the sweet taste of chocolate [3]. Conversely, in terms of the relative content in PH-16 dry cacao beans, leucine and phenylalanine, which are linked with bitter taste, add up to 29.39% (16.22 + 13.17) and glutamic acid, with its umami taste, represents 9.09%, whilst only the free amino acid alanine represents 8.65% (Table 3). Chocolate made from PH-16 cacao can vary between bitter and sweet tastes [3]. The intensification of the two tastes is probably related to the biochemical transformations and consequent increase in the content of these free amino acids in the chocolate.
Based on the results of this research in relation to PH-16 cacao (Table 3 and Table 4, Figure 3), it is possible to affirm that the free amino acid profile is a very stable biochemical characteristic of the cacao genotype. However, the environmental effect on the biochemical composition of cacao beans is extremely important for the quality of products such as chocolate. The combination of genetic and environmental factors enables the appellation of the origin of cacao that brings benefits for producers, traders and consumers. One of the achievements of this research was the environmental stratification by different crop edaphic conditions of the PH-16 cacao, a fact that made it possible to isolate the soil’s effect on the free amino acid profile. Likewise, the total number of observations of the sample (n = 36) guaranteed a robust exploitative analysis to find the relationships between free amino acids of PH-16 dry cacao beans.
In terms of the taste of the final chocolate product, cacao producers must, above all, consider the clonal variety or the “blend” of clonal varieties for manufacturing. Moreover, other production factors such as adequate environmental intervention through soil management and cacao crop are essential, mainly because of their direct relationship with production.
5. Conclusions
Leucine, phenylalanine, glutamic acid, alanine, asparagine, tyrosine, gamma-aminobutyric acid, valine, isoleucine, glutamine, lysine, aspartic acid, serine, tryptophan, threonine and glycine are the free amino acids found in the PH-16 dry cacao beans.
With the exception of lysine, under the ANOVA F-test, no other free amino acid showed a significant difference between means of crop sites represented by different edaphic conditions.
The hydrophobic free amino acids showed a great contribution to the explained variance of 58.01% of the first dimension of the principal component analysis. Glutamic acid stands out in the second dimension with 13.09% of the variation explained.
Author Contributions
Q.R.d.A. conceived the study and was in charge of overall direction and planning; Q.R.d.A. carried out the experiment and collected data; Q.R.d.A. and D.S. performed laboratory analysis; G.A.H.d.A.L. and C.E.M.P. performed statistical analysis; D.S., G.A.H.d.A.L. and S.S.d.A. reviewed the methodology; D.A. and V.C.B. supervised the investigation; Q.R.d.A. and G.A.H.d.A.L. wrote the original draft preparation. All authors contributed to review and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This paper is part of the project “Linking soil quality and cacao quality in Bahia, Brazil”. To run fundamental steps of this research, the first author was supported by the Brazilian National Council for Scientific and Technological Development (CNPq) with a postdoctoral fellowship. The project SCA 58-1245-3-237F of USDA-ARS-FUNPAB partially supported lab activities. We thank Marshall Elson for an excellent review of this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Araujo, Q.R.d.; Loureiro, G.A.H.d.A.; Baligar, V.C.; Ahnert, D.; Faria, J.C.; Valle, R.R.; Lieberei, R. Cacao quality index for cacao agroecosystems in Bahia, Brazil. Int. J. Food Prop. 2019, 22, 1799–1814. [Google Scholar] [CrossRef]
- Araujo, Q.R.; Fernandes, C.A.F.C.A.F.; Ribeiro, D.O.; Efraim, P.; Steinmacher, D.; Lieberei, R.; Bastide, P.; Araujo, T.G. Cocoa Quality Index—A proposal. Food Control 2014, 46, 49–54. [Google Scholar] [CrossRef]
- Deus, V.L.; Bispo, E.S.; Franca, A.S.; Gloria, M.B.A. Influence of cocoa clones on the quality and functional properties of chocolate—Nitrogenous compounds. LWT 2020, 134, 110202. [Google Scholar] [CrossRef]
- Loureiro, G.A.H.A.; Araujo, Q.R.; Sodré, G.A.; Valle, R.R.; Souza, J.O.; Ramos, E.M.L.S.; Comerford, N.B.; Grierson, P.F. Cacao quality: Highlighting selected attributes. Food Rev. Int. 2017, 33, 382–405. [Google Scholar] [CrossRef]
- Adeyeye, E.I.; Akinyeye, R.O.; Ogunlade, I.; Olaofe, O.; Boluwade, J.O. Effect of farm and industrial processing on the amino acid profile of cocoa beans. Food Chem. 2010, 118, 357–363. [Google Scholar] [CrossRef]
- De Brito, E.S.; Pezoa García, N.H.; Gallão, M.I.; Cortelazzo, A.L.; Fevereiro, P.S.; Braga, M.R. Structural and chemical changes in cocoa (Theobroma cacao L.) during fermentation, drying and roasting. J. Sci. Food Agric. 2001, 81, 281–288. [Google Scholar] [CrossRef]
- Rohsius, C.; Matissek, R.; Lieberei, R. Free amino acid amounts in raw cocoas from different origins. Eur. Food Res. Technol. 2006, 222, 432–438. [Google Scholar] [CrossRef]
- Gu, F.; Tan, L.; Wu, H.; Fang, Y.; Xu, F.; Chu, Z.; Wang, Q. Comparison of cocoa beans from China, Indonesia and Papua New Guinea. Foods 2013, 2, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, A.; Palla, G.; Caligiani, A. Presence and variation of γ-aminobutyric acid and other free amino acids in cocoa beans from different geographical origins. Food Res. Int. 2014, 63, 360–366. [Google Scholar] [CrossRef]
- Kirchhoff, P.M.; Biehl, B.; Crone, G. Peculiarity of the accumulation of free amino acids during cocoa fermentation. Food Chem. 1989, 31, 295–311. [Google Scholar] [CrossRef]
- Kirchhoff, P.-M.; Biehl, B.; Ziegeler-Berghausen, H.; Hammoor, M.; Lieberei, R. Kinetics of the formation of free amino acids in cocoa seeds during fermentation. Food Chem. 1989, 34, 161–179. [Google Scholar] [CrossRef]
- Brunetto, M.d.R.; Gallignani de Bernardi, M.A.; Orozco Contreras, W.J.; Clavijo Roa, S.d.S.; Delgado Cayama, Y.J.; Ayala Montilla, C.D.; Zambrano García, A. RP-HPLC-DAD determination of free amino acids in cocoa samples during fermentation and roasting. Rev. Colomb. Química 2020, 49, 11–19. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; W. H. Freeman: New York, NY, USA, 2017. [Google Scholar]
- Deus, V.L.; Bispo, E.S.; Franca, A.S.; Gloria, M.B.A. Understanding amino acids and bioactive amines changes during on-farm cocoa fermentation. J. Food Compos. Anal. 2021, 97, 103776. [Google Scholar] [CrossRef]
- Piasentin, F.B.; Saito, C.H. The different methods of cocoa farming in southeastern Bahia, Brazil: Historical aspects and perceptions. Bol. Do Mus. Para. Emílio Goeldi. Ciências Hum. 2014, 9, 61–78. [Google Scholar] [CrossRef]
- Chepote, R.E.; Santana, S.O.; Araujo, Q.R.; Sodré, G.A.; Reis, E.L.; Pacheco, R.G.; Marrocos, P.C.L.; Serôdio, M.H.d.C.F.; Valle, R.R. Aptidão agrícola e fertilidade de solos para a cultura do cacaueiro. In Ciência, Tecnologia e Manejo do Cacaueiro; Valle, R.R., Ed.; MAPA/CEPLAC/CEPEC: Brasília, DF, Brazil, 2012; pp. 67–114. [Google Scholar]
- Loureiro, G.A.H.d.A.; Dantas, P.A.d.S.; de Araujo, Q.R. Manejo e conservação do solo para cacaueiro. In Cacau: Cultivo, Pesquisa e Inovação; De Souza Júnior, J.O., Ed.; Editus: Ilhéus, BA, Brazil, 2018; pp. 399–436. [Google Scholar]
- Araujo, Q.; Ahnert, D.; Loureiro, G.; Faria, J.; Fernandes, C.; Baligar, V. Soil quality index for cacao cropping systems. Arch. Agron. Soil Sci. 2018, 64, 1892–1909. [Google Scholar] [CrossRef]
- De Araujo, Q.R.; Baligar, V.C.; Loureiro, G.A.H.d.A.; de Souza Júnior, J.O.; Comerford, N.B. Impact of soils and cropping systems on mineral composition of dry cacao beans. J. Soil Sci. Plant Nutr. 2017, 17, 410–428. [Google Scholar] [CrossRef][Green Version]
- Araujo, Q.R.; Loureiro, G.A.H.A.; Rohsius, C.; Baligar, V.C. Impact of soils and cropping systems on biochemical attributes of dry cacao beans. Agrotrópica 2018, 30, 175–194. [Google Scholar] [CrossRef]
- Mandarino, E.P.; Sena Gomes, A.R. Produtividade do Cacaueiro (Theobroma cacao L.) Cultivado em Blocos Monoclonais, no sul da Bahia, Brazil; Boletim Técnico, No. 197; CEPLAC/CEPEC: Ilhéus, BA, Brazil, 2009. [Google Scholar]
- Cruz, J.F.M.; Leite, P.B.; Soares, S.E.; Bispo, E.d.S.; Leite, P.B.; Soares, S.E.; Bispo, E.d.S. Assessment of the fermentative process from different cocoa cultivars produced in Southern Bahia, Brazil. Afr. J. Biotechnol. 2013, 12, 5218–5225. [Google Scholar] [CrossRef]
- Sodré, G.A.; Marrocos, P.C.L.; Sarmento, D.A. Cultivo do Cacaueiro no Estado do Ceará; Boletim Técnico, No. 209; CEPLAC/CEPEC: Ilheus, BA, Brazil, 2017. [Google Scholar]
- Loureiro, G.A.H.d.A.; de Araujo, Q.R.; Valle, R.R.; Sodré, G.A.; Santos Filho, L.P.; Oliveira, S.J.R.; Dantas, P.A.S.; Couto, L.R.; Lopes, M.R. Influence of environmental factors on cacao biometric attributes. Agrotrópica 2016, 28, 123–140. [Google Scholar] [CrossRef]
- Loureiro, G.A.H.d.A.; de Araujo, Q.R.; Valle, R.R.; Sodré, G.A.; de Souza, S.M.M. Influencia de factores agroambientales sobre la calidad del clon de cacao (Theobroma cacao L.) PH-16 en la región cacaotera de Bahia, Brazil. Ecosistemas y Recur. Agropecu. 2017, 4, 579. [Google Scholar] [CrossRef]
- Sodré, G.A.; Gomes, A.R.S. Cocoa propagation, technologies for production of seedlings. Rev. Bras. Frutic. 2019, 41. [Google Scholar] [CrossRef]
- Resende, M.; Curi, N.; de Rezende, S.B.; Corrêa, G.F. Pedologia: Base Para Distinção de Ambientes, 6th ed.; Editora UFLA: Lavras, MG, Brazil, 2014. [Google Scholar]
- Superintendência de Estudos Econômicos e Sociais da Bahia (SEI). Tipologia Climática—Segundo Thornthwaite—Pluviometria 1943–1983/Temperatura 1961–1990. Estado da Bahia. Available online: http://www.sei.ba.gov.br/site/geoambientais/cartogramas/pdf/carto_tip_clim.pdf (accessed on 2 February 2014).
- Dos Santos, H.G.; Jacomine, P.K.T.; dos Anjos, L.H.C.; de Oliveira, V.Á.; Lumbreras, J.F.; Coelho, M.R.; de Almeida, J.A.; Cunha, T.J.F.; de Oliveira, J.B. Sistema Brasileiro de Classificação de Solos, 3rd ed.; Embrapa: Brasilia, DF, Brazil, 2013. [Google Scholar]
- Methodenbuch Band III Methode, Nr. 3.6. In Ergänzungslieferung; Naumann, C., Bassler, R., Seibold, R., Barth, C., Eds.; VDLUFA-Verlag: Darmstadt, Germany, 1976. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Husson, F.; Josse, J. Package ‘missMDA’. 2020, p. 41. Available online: https://cran.rediris.es/web/packages/FactoMineR/FactoMineR.pdf (accessed on 1 June 2021).
- Husson, F.; Josse, J.; Le, S.; Mazet, J. Package ‘FactoMineR’. 2020, p. 100. Available online: http://203.178.137.175/pub/lang/R/CRAN/web/packages/FactoMineR/FactoMineR.pdf (accessed on 1 June 2021).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. RStudio Package ‘ggplot2′. 2020, p. 284. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 1 June 2021).
- Stark, T.; Hofmann, T. Isolation, Structure Determination, Synthesis, and Sensory Activity of N -Phenylpropenoyl- l -amino Acids from Cocoa (Theobroma cacao). J. Agric. Food Chem. 2005, 53, 5419–5428. [Google Scholar] [CrossRef] [PubMed]
- Voigt, J.; Heinrichs, H.; Voigt, G.; Biehl, B. Cocoa-specific aroma precursors are generated by proteolytic digestion of the vicilin-like globulin of cocoa seeds. Food Chem. 1994, 50, 177–184. [Google Scholar] [CrossRef]
- Jinap, S.; Hajeb, P. Glutamate. Its applications in food and contribution to health. Appetite 2010, 55, 1–10. [Google Scholar] [CrossRef]
- Yoshida, Y. Umami taste and traditional seasonings. Food Rev. Int. 1998, 14, 213–246. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).