First Report of Glyphosate-Resistant Biotype of Eleusine Indica (L.) Gaertn. in Europe

 ,
, 
 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Collection and Plant Growing Conditions
2.2. Dose-Response Experiments
2.3. Statistical Analyses
2.4. EPSPS Sequencing
3. Results
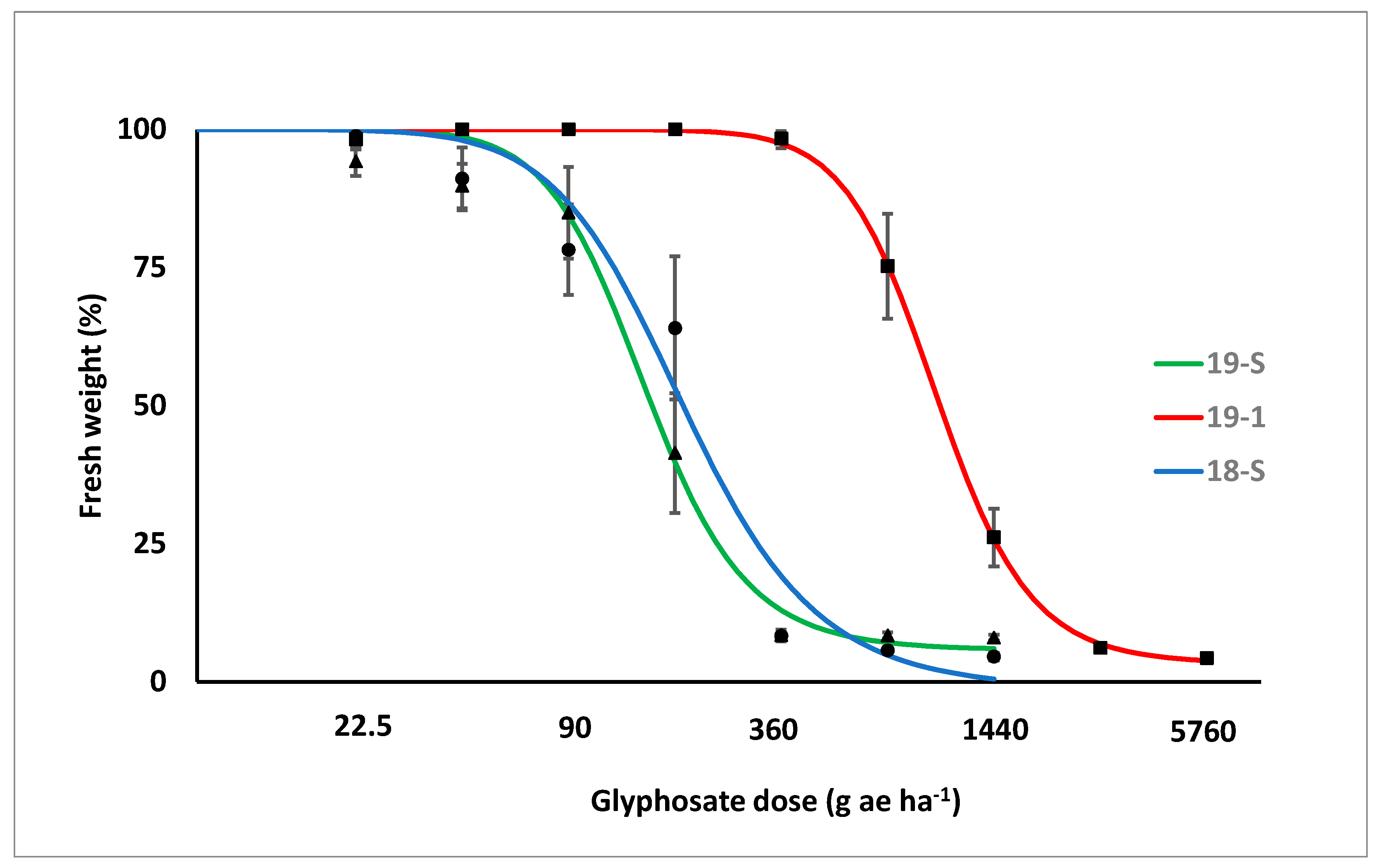
3.1. Dose-Response Experiment
3.2. EPSPS Sequencing
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Lorraine-Colwill, D.F.; Dellow, J.J.; Preston, C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia. Weed Sci. 1998, 46, 604–607. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 30 October 2020).
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef]
- Amaro-Blanco, I.; Fernández-Moreno, P.T.; Osuna-Ruiz, M.D.; Bastida, F.; De Prado, R. Mechanisms of glyphosate resistance and response to alternative herbicide-based management in populations of the three Conyza species introduced in southern Spain. Pest Manag. Sci. 2018, 74, 1925–1937. [Google Scholar] [CrossRef]
- Tahmasebi, B.K.; Alebrahim, M.T.; Roldán-Gómez, R.A.; Silveira, H.M.D.; Carvalho, L.B.D.; Alcántara-de la Cruz, R.; De Prado, R. Effectiveness of alternative herbicides on three Conyza species from Europe with and without glyphosate resistance. Crop Prot. 2018, 112, 350–355. [Google Scholar] [CrossRef]
- Travlos, I.S.; Chachalis, D. Assessment of glyphosate-resistant horseweed (Conyza canadensis L. Cronq.) and fleabane (Conyza albida Willd. ex Spreng) populations from perennial crops in Greece. Int. J. Plant Prod. 2013, 7, 665–676. [Google Scholar]
- Fernández-Moreno, P.T.; Travlos, I.; Brants, I.; De Prado, R. Different levels of glyphosate-resistant Lolium rigidum L. among major crops in southern Spain and France. Sci. Rep. 2017, 7, 13116. [Google Scholar] [CrossRef]
- Collavo, A.; Sattin, M. Resistance to glyphosate in Lolium rigidum selected in Italian perennial crops: Bioevaluation, management and molecular bases of target-site resistance. Weed Res. 2012, 52, 16–24. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Palma-Bautista, C.; Rojano-Delgado, A.M.; De Prado, R.; Menendez, J. The first case of glyphosate resistance in johnsongrass (Sorghum halepense (L.) Pers.) in Europe. Plants 2020, 9, 313. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Castro, P.; Torra, J.; Alcántara-de la Cruz, R.; De Prado, R. Resistance evolution to EPSPS inhibiting herbicides in false barley (Hordeum murinum) harvested in Southern Spain. Agronomy 2020, 10, 992. [Google Scholar] [CrossRef]
- USDA-ARS. Germplasm Resources Information Network (GRIN). Online Database. Beltsville, Maryland, USA: National Germplasm Resources Laboratory. Available online: https://npgsweb.ars-grin.gov/gringlobal/taxon/taxonomydetail?id=14997 (accessed on 30 October 2020).
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds: Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Pignatti, S. Flora d’Italia, 1st ed.; Edagricole: Bologna, Italy, 1982; Volume 3, p. 600. [Google Scholar]
- Zanin, G.; Cantele, A.; Della Pietà, S.; Lorenzoni, G.G.; Tei, F.; Vazzana, C. Le Erbe Infestanti Graminacee Nelle a Moderna Agricoltura: Dinamica, Problemi e Possibili Soluzioni; Atti Società Italiana per lo Studio della Lotta alle Malerbe (S.I.L.M): Verona, Italy, 14 November 1985; Tipografia Guerra: Perugia, Italy, 1985; pp. 13–248. [Google Scholar]
- Motti, R.; Espodito, A.; Stinca, A. New additions s to the exotic vascular flora of Campania (Southern Italy). Ann. Bot. Italy 2018, 8, 75–85. [Google Scholar] [CrossRef]
- Ampong-Nyarko, K.; de Datta, S.K.; Dingkuhn, M. Physiological response of rice and weeds to low light intensity at different growth stages. Weed Res. 1992, 32, 465–472. [Google Scholar] [CrossRef]
- Lee, L.J.; Ngim, J. A first report of glyphosate-resistant goosegrass (Eleusine indica (L) Gaertn) in Malaysia. Pest Manag. Sci. 2000, 56, 336–339. [Google Scholar] [CrossRef]
- Chen, J.-C.; Huang, H.-J.; Wei, S.-H.; Zhang, C.-X.; Huang, Z.-F. Characterization of glyphosate-resistant goosegrass (Eleusine indica) populations in China. J. Integr. Agric. 2015, 14, 919–925. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Zelaya, I.A.; Dale, R.P.; Lycett, A.J.; Carter, P.; Sharples, K.R.; McIndoe, E. Importance of the P106S target-site mutation in conferring resistance to glyphosate in a goosegrass (Eleusine indica) population from the Philippines. Weed Sci. 2008, 56, 637–646. [Google Scholar] [CrossRef]
- Tampubolon, K.; Purba, E.; Basyuni, M.; Hanafiah, D.S. Glyphosate resistance of Eleusine indica populations from North Sumatra, Indonesia. Biodiversitas 2019, 20, 1910–1916. [Google Scholar] [CrossRef]
- Takano, H.K.; Oliveira, R.S.; Constantin, J.; Braz, G.B.P.; Gheno, E.A. Goosegrass resistant to glyphosate in Brazil [Capim-pé-de-galinha resistente ao glyphosate no Brasil]. Planta Daninha 2017, 35, e017163071. [Google Scholar] [CrossRef]
- Gherekhloo, J.; Fernández-Moreno, P.T.; Alcántara-De La Cruz, R.; Sánchez-González, E.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Pro-106-Ser mutation and EPSPS overexpression acting together simultaneously in glyphosate-resistant goosegrass (Eleusine indica). Sci. Rep. 2017, 7, 6702. [Google Scholar] [CrossRef]
- Mueller, T.C.; Barnett, K.A.; Brosnan, J.T.; Steckel, L.E. Glyphosate-resistant goosegrass (Eleusine indica) confirmed in Tennessee. Weed Sci. 2011, 59, 562–566. [Google Scholar] [CrossRef]
- Jalaludin, A.; Yu, Q.; Powles, S.B. Multiple resistance across glufosinate, glyphosate, paraquat and ACCase-inhibiting herbicides in an Eleusine indica population. Weed Res. 2015, 55, 82–89. [Google Scholar] [CrossRef]
- Deng, W.; Yang, Q.; Chen, Y.; Yang, M.; Xia, Z.; Zhu, J.; Chen, Y.; Cai, J.; Yuan, S. Cyhalofop-butyl and glyphosate multiple-herbicide resistance evolved in an Eleusine indica population collected in Chinese direct-seeding rice. J. Agric. Food Chem. 2020, 68, 2623–2630. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, C.; Huang, H.; Wei, S.; Huang, Z.; Wang, H.; Zhao, D.; Zhang, C. Characterization of Eleusine indica with gene mutation or amplification in EPSPS to glyphosate. Pestic. Biochem. Physiol. 2017, 143, 201–206. [Google Scholar] [CrossRef]
- Chen, J.; Huang, H.; Wei, S.; Cui, H.; Li, X.; Zhang, C. Glyphosate resistance in Eleusine indica: EPSPS overexpression and P106A mutation evolved in the same individuals. Pestic. Biochem. Physiol. 2020, 164, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.L.; Riggins, C.W.; Steckel, L.E.; Tranel, P.J. The EPSPS Pro106Ser substitution solely accounts for glyphosate resistance in a goosegrass (Eleusine indica) population from Tennessee, United States. J. Integr. Agric. 2016, 15, 1304–1312. [Google Scholar] [CrossRef]
- Chen, J.; Huang, H.; Zhang, C.; Wei, S.; Huang, Z.; Chen, J.; Wang, X. Mutations and amplification of EPSPS gene confer resistance to glyphosate in goosegrass (Eleusine indica). Planta 2015, 242, 859–868. [Google Scholar] [CrossRef]
- Ng, C.H.; Wickneswari, R.; Salmijah, S.; Teng, Y.T.; Ismail, B.S. Gene polymorphisms in glyphosate-resistant and -susceptible biotypes of Eleusine indica from Malaysia. Weed Res. 2003, 43, 108–115. [Google Scholar] [CrossRef]
- Takano, H.K.; Mendes, R.R.; Scoz, L.B.; Lopez Ovejero, R.F.; Constantin, J.; Gaines, T.A.; Westra, P.; Dayan, F.E.; Oliveira, R.S. Proline-106 EPSPS mutation imparting glyphosate resistance in goosegrass (Eleusine indica) Emerges in South America. Weed Sci. 2019, 67, 48–56. [Google Scholar] [CrossRef]
- Yu, Q.; Jalaludin, A.; Han, H.; Chen, M.; Douglas Sammons, R.; Powles, S.B. Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant Physiol. 2015, 167, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Vila-Aiub, M.M.; Jalaludin, A.; Yu, Q.; Powles, S.B. A double EPSPS gene mutation endowing glyphosate resistance shows a remarkably high resistance cost. Plant Cell. Environ. 2017, 40, 3031–3042. [Google Scholar] [CrossRef]
- Zhang, C.; Feng, L.; He, T.T.; Yang, C.H.; Chen, G.Q.; Tian, X.S. Investigating the mechanisms of glyphosate resistance in goosegrass (Eleusine indica) population from South China. J. Integr. Agric. 2015, 14, 909–918. [Google Scholar] [CrossRef]
- Hess, M.; Barralis, G.; Bleiholder, H.; Buhr, L.; Eggers, T.; Hack, H.; Stauss, R. Use of the extended BBCH scale—General for the descriptions of the growth stages of mono- and dicotyledonous weed species. Weed Res. 1997, 37, 433–441. [Google Scholar] [CrossRef]
- Onofri, A. Bioassay97: A new Excel VBA macro to perform statistical analyses on herbicide dose-response data. Riv. Ital. Agrometeorol. 2005, 3, 40–45. [Google Scholar]
- Aras, S.; Duran, A.; Yenilmez, G. Isolation of DNA for RAPD analysis from dry leaf material of some Hesperis L. specimens. Plant Mol. Biol. Rep. 2003, 21, 461–462. [Google Scholar] [CrossRef]
- Milani, A.; Panozzo, S.; Farinati, S.; Loddo, D. Molecular data for: First report of glyphosate-resistant biotype of Eleusine indica in Europe. Mendeley Data 2020, V1. [Google Scholar] [CrossRef]
- Molin, W.; Wright, A.; Nandula, V. Glyphosate-resistant goosegrass from Mississippi. Agronomy 2013, 3, 474–487. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S. Glyphosate, paraquat and ACCase multiple herbicide resistance evolved in a Lolium rigidum biotype. Planta 2007, 225, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.B.; McCarty, L.B.; Tharayil, N.; McElroy, J.S.; Chen, S.; McCullough, P.E.; Powell, B.A.; Bridges, W.C. A Pro 106 to Ala Substitution is associated with resistance to glyphosate in annual bluegrass (Poa annua). Weed Sci. 2015, 63, 613–622. [Google Scholar] [CrossRef]
- Bi, B.; Wang, Q.; Coleman, J.J.; Porri, A.; Peppers, J.M.; Patel, J.D.; Betz, M.; Lerchl, J.; McElroy, J.S. A novel mutation A212T in chloroplast Protoporphyrinogen oxidase (PPO1) confers resistance to PPO inhibitor Oxadiazon in Eleusine indica. Pest Manag. Sci. 2020, 76, 1786–1794. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Nishimoto, R.K.; DeFrank, J. Metribuzin-resistant goosegrass (Eleusine indica) in bermudagrass turf. Weed Technol. 2008, 22, 675–678. [Google Scholar] [CrossRef]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Permingeat, H.R. Herbicide resistant weeds: A call to integrate conventional agricultural practices, molecular biology knowledge and new technologies. Plant Sci. 2020, 290, 110255. [Google Scholar] [CrossRef] [PubMed]
- Masin, R.; Zuin, M.C.; Otto, S.; Zanin, G. Seed longevity and dormancy of four summer annual grass weeds in turf. Weed Res. 2006, 46, 362–370. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biotype | ED50 | ED90 | RI |
|---|---|---|---|
| g ae ha−1 | |||
| 19-1 | 1402 ± 20.9 | 2030 ± 172.1 | 7.3 |
| 18-S | 199 ± 30.7 | 264 ± 146.1 | |
| 19-5 | 183 ± 7.6 | 295 ± 44.9 | |
| GR50 | GR90 | RI | |
| g ae ha−1 | |||
| 19-1 | 999 ± 49.5 | 1931 ± 206.4 | 5.8 |
| 18-S | 192 ± 20.5 | 480 ± 124.0 | |
| 19-5 | 50 ± 12.0 | 311 ± 56.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loddo, D.; Imperatore, G.; Milani, A.; Panozzo, S.; Farinati, S.; Sattin, M.; Zanin, G. First Report of Glyphosate-Resistant Biotype of Eleusine Indica (L.) Gaertn. in Europe. Agronomy 2020, 10, 1692. https://doi.org/10.3390/agronomy10111692
Loddo D, Imperatore G, Milani A, Panozzo S, Farinati S, Sattin M, Zanin G. First Report of Glyphosate-Resistant Biotype of Eleusine Indica (L.) Gaertn. in Europe. Agronomy. 2020; 10(11):1692. https://doi.org/10.3390/agronomy10111692
Chicago/Turabian StyleLoddo, Donato, Gaetano Imperatore, Andrea Milani, Silvia Panozzo, Silvia Farinati, Maurizio Sattin, and Giuseppe Zanin. 2020. "First Report of Glyphosate-Resistant Biotype of Eleusine Indica (L.) Gaertn. in Europe" Agronomy 10, no. 11: 1692. https://doi.org/10.3390/agronomy10111692
APA StyleLoddo, D., Imperatore, G., Milani, A., Panozzo, S., Farinati, S., Sattin, M., & Zanin, G. (2020). First Report of Glyphosate-Resistant Biotype of Eleusine Indica (L.) Gaertn. in Europe. Agronomy, 10(11), 1692. https://doi.org/10.3390/agronomy10111692