


Pesticides in Foods: Towards Bioremediation Biocatalysts?


 ,
,  and
and 
Abstract

1. Introduction
2. Pesticides
- -
- Application: In agriculture, pesticides are used to safeguard crops against pests, in-sects, and weeds; in public health, pesticides are employed to eradicate vectors that cause diseases (e.g., malaria); and domestic pesticides are used to eliminate insects such as cockroaches, bacteria, protozoa, and mice [5].
- -
- Target organisms: Insecticides, which are chemicals designed to eliminate insects; fungicides, which are applied to inhibit or eradicate fungi; herbicides, which are used to control or eliminate weeds; rodenticides, which are pesticides intended to eliminate rodents; fumigants, which are gaseous pesticides used to control or eliminate pests such as bedbugs; and insect repellents, which are applied to the skin or clothing to deter insects from approaching [5].
- -
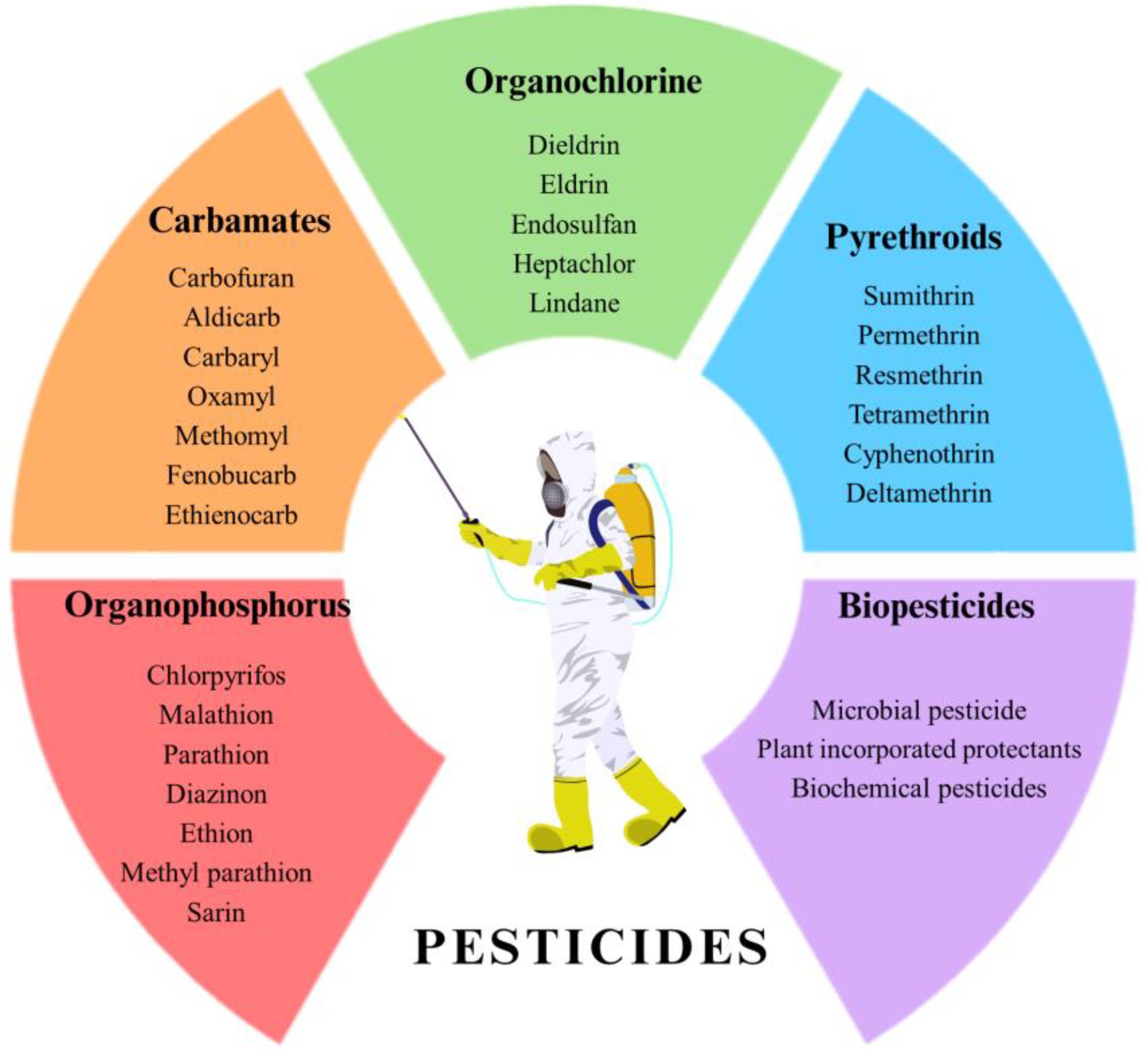
- Chemical composition: Including organochlorines (OC), organophosphates (OP), carbamates, pyrethroids, phenyl amides (carbanilates, acylanalides, toluidines, and acetamides), phenoxyalkonates, trazines, benzoic acid derivatives, benzonitriles, phtalimide derivatives, and dipyrids [5]. The main categories of pesticides are reported in Figure 1.
- -
- Dichlorodiphenylethane or diphenyl aliphatics (DDT (dichlorodiphenyltrichloro-ethane), DDD (dichlorodiphenyldichloroethane), dicofol, ethylan, chlorobenzilate, and methoxychlor);
- -
- Cyclodienes (CHL (chlordane), aldrin, dieldrin, heptachlor, endrin, dodecachloro-pentacyclodecane (mirex), and endosulfan (cyclic ester of sulfuric acid));
- -
- Cylohexanes;
- -
- Chlorinated camphenes (toxaphene and chlordecone).
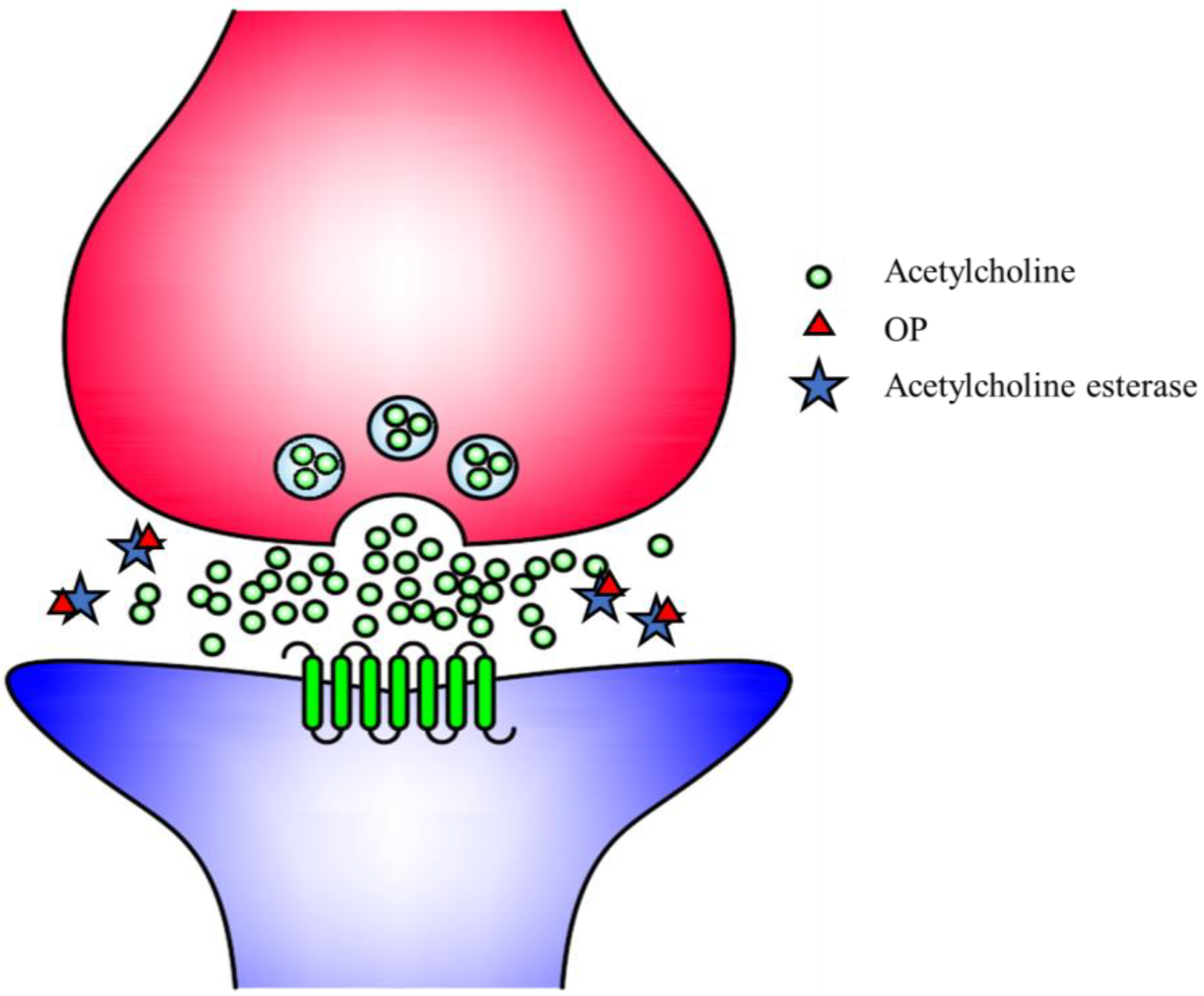
3. Pesticides and Public Health
4. Bioremediation
5. Microbial Degradation of Pesticides in Fermented Foods
6. Lactic Acid Bacteria as Pesticide-Degraders
7. Filamentous Fungi as Pesticide-Degraders
Fungal Enzymes Involved in the Degradation of Pesticides
8. Limits of Pesticide Bioremediation Biocatalysts
8.1. Are Degradation Products Worse Than the Pesticide Itself?
8.2. Applying Bioremediation Biocatalysts in Food
8.2.1. Lactic Acid Bacteria and Their Enzymes
8.2.2. Fungi and Their Enzymes
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lee, S.; Moore, L.; Park, S.; Harris, D.; Blanck, H. Adults Meeting Fruit and Vegetable Intake Recommendations—United States, 2019. Morbid. Mortal. Wkly. Rep. 2022, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Insausti, H.; Chiu, Y.; Wang, Y.; Hart, J.; Bhupathiraju, S.; Mínguez-Alarcón, L.; Ding, M.; Willett, W.; Laden, F.; Chavarro, J. Intake of fruits and vegetables according to pesticide residue status in relation to all-cause and disease-specific mortality: Results from three prospective cohort studies. Environ. Int. 2022, 159, 107024. [Google Scholar] [CrossRef] [PubMed]
- Pesticide Action Network Europe. The dramatic rise in the most toxic pesticides found on fruits and vegetables sold in Europe and evidence that governments are failing their leagal obligations. In Forbidden Fruit; PAN Europe: Brussels, Belgium, 2022. [Google Scholar]
- Sun, S.; Sidhu, V.; Rong, Y.; Zheng, Y. Pesticide pollution in agricultural soils and sustainable remediation methods: A review. Curr. Pollut. Rep. 2018, 4, 240–250. [Google Scholar] [CrossRef]
- Bose, S.; Kumar, P.S.; Vo, D.V.N.; Rajamohan, N.; Saravanan, R. Microbial degradation of recalcitrant pesticides: A review. Environ. Chem. Lett. 2021, 19, 3209–3228. [Google Scholar] [CrossRef]
- Rajmohan, K.S.; Chandrasekaran, R.; Varjani, S. A Review on occurrence of pesticides in environment and current technologies for their remediation and management. Indian J. Microbiol. 2020, 60, 125–138. [Google Scholar] [CrossRef]
- Carriger, J.F.; Rand, G.M.; Gardinali, P.R.; Perry, W.B.; Tompkins, M.S.; Fernandez, A.M. Pesticides of potential ecological concern in sediment from South Florida canals: An ecological risk prioritization for aquatic arthropods. Soil Sediment Contam. 2006, 15, 21–45. [Google Scholar] [CrossRef]
- Mostafalou, S.; Abdollahi, M. Concerns of environmental persistence of pesticides and human chronic diseases. Clin. Exp. Pharmacol. 2012, S5, e002. [Google Scholar] [CrossRef]
- Tudi, M.; Li, H.; Li, H.; Wang, L.; Lyu, J.; Yang, L.; Tong, S.; Yu, Q.J.; Ruan, H.D.; Atabila, A.; et al. Exposure routes and health risks associated with pesticide application. Toxics 2022, 10, 335. [Google Scholar] [CrossRef]
- Wahab, S.; Muzammil, K.; Nasir, N.; Khan, M.S.; Ahmad, M.F.; Khalid, M.; Ahmad, W.; Dawria, A.; Reddy, L.K.V.; Busayli, A.M. Advancement and New Trends in Analysis of Pesticide Residues in Food: A Comprehensive Review. Plants 2022, 11, 1106. [Google Scholar] [CrossRef]
- Mali, H.; Shah, C.; Raghunandan, B.H.; Prajapati, A.S.; Patel, D.H.; Trivedi, U.; Subramanian, R.B. Organophosphate pesticides an emerging environmental contaminant: Pollution, toxicity, bioremediation progress, and remaining challenges. J. Environ. Sci. 2022, 127, 234–250. [Google Scholar] [CrossRef]
- Farnoosh, G.; Latifi, A.M. A review on engineering of organophosphorus hydrolase (OPH) Enzyme. J. Appl. Biotechnol. Rep. 2014, 1, 1–10. [Google Scholar]
- Keswani, C.; Dilnashin, H.; Birla, H.; Roy, P.; Tyagi, R.K.; Singh, D.; Rajput, V.D.; Minkina, T.; Singh, S.P. Global footprints of organochlorine pesticides: A pan-global survey. Environ. Geochem. Health 2022, 44, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Al Antary, T.M.; Alawi, M.A.; Estityah, H.; Haddad, N.; Al-Antary, E.T. Chlorinated pesticide residues in human breast milk collected from southern Jordan in 2012/2013. Toxin Rev. 2015, 34, 190–194. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M.N.; Barceló, D. Persistence of pesticides-based contaminants in the environment and their effective degradation using laccase-assisted biocatalytic systems. Sci. Total Environ. 2019, 695, 133896. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, Y.; Liu, L.; Yan, H. Pesticide exposure and risk of Alzheimer’s disease: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 32222. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Nicell, J.A. Impact of reaction conditions on the laccase-catalyzedconversion of bisphenol. Bioresour. Technol. 2006, 97, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Mehrpour, O.; Karrari, P.; Zamani, N.; Tsatsakis, A.M.; Abdollahi, M. Occupational exposure to pesticides and consequences on male semen and fertility: A review. Toxicol. Lett. 2014, 230, 146–156. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D. Worldwide pesticide usage and its impacts on ecosystem. Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Verma, J.P.; Jaiswal, D.K.; Sagar, R. Pesticide relevance and their microbial degradation: A-state-of-art. Rev. Environ. Sci. Biotechnol. 2014, 13, 429–466. [Google Scholar] [CrossRef]
- Odukkathil, G.; Vasudevan, N. Enhanced biodegradation of endosulfan and its major metabolite endosulfate by a biosurfactant producing bacterium. J. Environ. Sci. Health Part B 2013, 48, 462–469. [Google Scholar] [CrossRef]
- Bajaj, A.; Pathak, A.; Mudiam, M.R.; Mayilraj, S.; Manickam, N. Isolation and characterization of a Pseudomonas sp. strain IITR01 capable of degrading α-endosulfan and endosulfan sulfate. J. Appl. Microbiol. 2010, 109, 2135–2143. [Google Scholar] [CrossRef]
- Armenova, N.; Tsigoriyna, L.; Arsov, A.; Petrov, K.; Petrova, P. Microbial detoxification of residual pesticides in fermented foods: Current status and prospects. Foods 2023, 12, 1163. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Ghosh, P.; Malyan, S.K.; Sharma, J.; Kumar, V. A comprehensive review on enzymatic degradation of the organophosphate pesticide malathion in the environment. J. Environ. Sci. Health 2019, 37, 288–329. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.U.; Parveen, S. Pesticides pollution and risk assessment of river ganga: A review. Heliyon 2021, 7, e07726. [Google Scholar] [CrossRef] [PubMed]
- Pigłowski, M. Notifications on Pesticide Residues in the Rapid Alert System for Food and Feed (RASFF). Int. J. Environ. Res. Public Health 2022, 19, 8525. [Google Scholar] [CrossRef]
- Adeyeye, A.; Osibanjo, O. Residues of organochlorine pesticides in fruits, vegetables and tubers from Nigerian markets. Sci. Total Environ. 1999, 231, 227–233. [Google Scholar] [CrossRef]
- Ibotom, M.O.; Mohammade, F. Determination of Pesticides Residues in Fruits and Vegetables in Kaduna Metropolis, Nigeria. Int. J. Environ. Sci. Toxicol. Res. 2016, 4, 185–189. [Google Scholar]
- Omwenga, I.; Kanja, L.; Zomer, P.; Louisse, J.; Rietjens, I.M.C.M.; Mol, H. Organophosphate and carbamate pesticide residues and accompanying risks in commonly consumed vegetables in Kenya. Food Addit. Contam. Part B Surv. 2021, 14, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xiao, L.; Li, F.; Xiao, M.; Lin, D.; Long, X.; Wu, Z. Microbial Degradation of Pesticide Residues and an Emphasis on the Degradation of Cypermethrin and 3-phenoxy Benzoic Acid: A Review. Molecules 2018, 23, 2313. [Google Scholar] [CrossRef]
- Abou-Arab, A. Effect of Ras cheese manufacturing on the stability of DDT and its metabolites. Food Chem. 1997, 59, 115–119. [Google Scholar] [CrossRef]
- Navarro, S.; Pérez, G.; Navarro, G.; Vela, N. Decline of pesticide residues from barley to malt. Food Addit. Contam. 2007, 24, 851–859. [Google Scholar] [CrossRef]
- Rajashekar, K.; Kondal, K.; Narasimha, K.; Sudhakar, K. Effect of processing of milk into products on the residue levels of certain pesticides. J. Food. Sci. Technol. 2007, 44, 551–552. [Google Scholar]
- Jung, K.; Park, Y.; Kim, H.; Kang, M.; Yang, Y.; Kang, A.; Chun, K.; Park, Y. Removal effects of Bifenthrin and Metalaxyl pesticides during preparation and fermentation of Baechu Kimchi. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1258–1264. [Google Scholar] [CrossRef]
- Bo, L.-Y.; Zhang, Y.-H.; Zhao, X.-H. Degradation kinetics of seven organophosphorus pesticides in milk during yoghurt processing. J. Serb. Chem. Soc. 2011, 76, 353–362. [Google Scholar] [CrossRef]
- Cuš, F.; Cesnik, B.; Bolta, V.; Gregorcic, A. Pesticide residues in grapes and during vinification process. Food Control 2010, 21, 1512–1518. [Google Scholar] [CrossRef]
- Ksiazek-Trela, P.; Szpyrka, E. The effect of natural and biological pesticides. Plant Prot. Sci. 2022, 58, 273–291. [Google Scholar] [CrossRef]
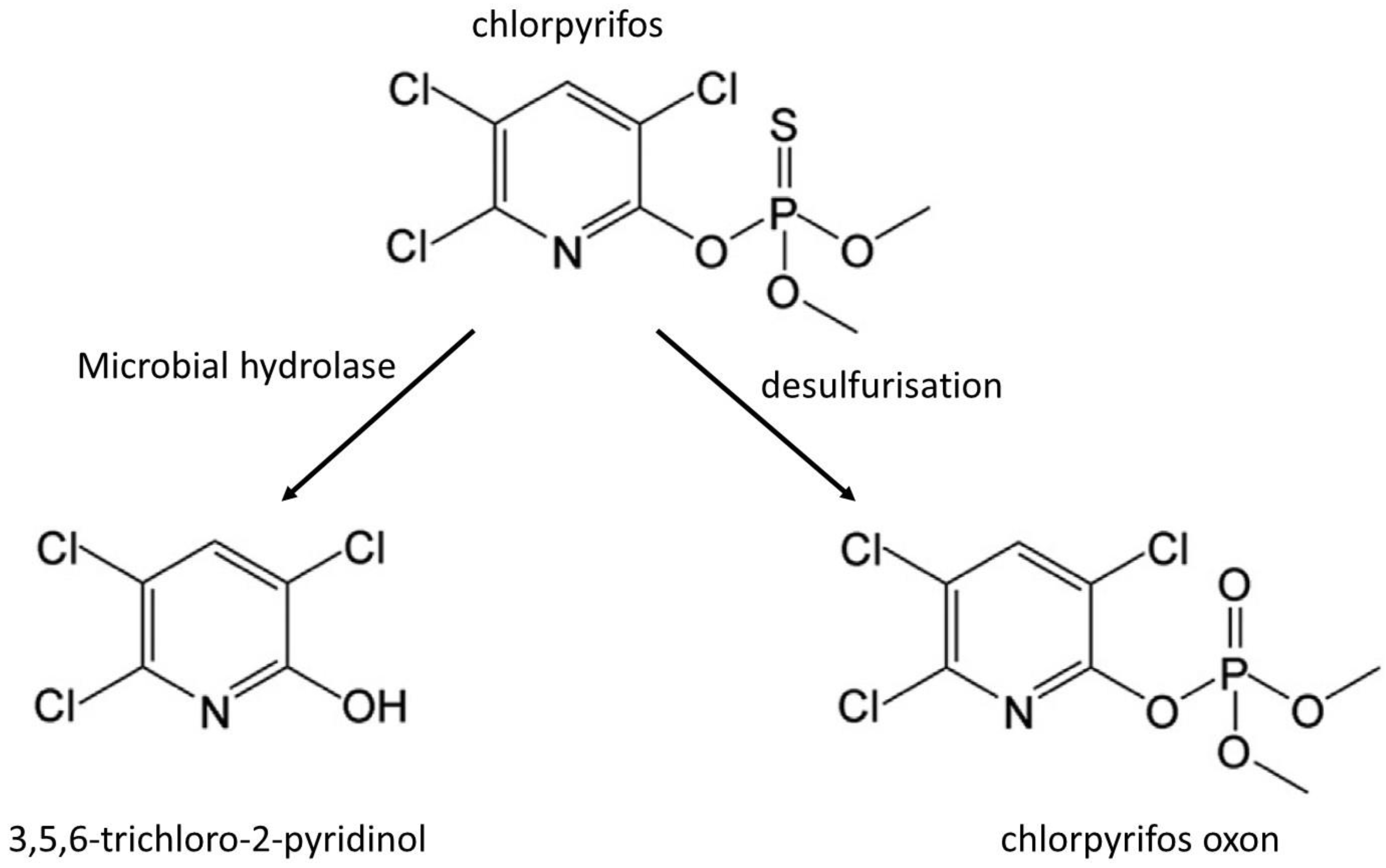
- Nandhini, A.; Harshiny, M.; Gummadi, S. Chlorpyrifos in environment and foods: A critical review of detection methods and degradation pathways. Environ. Sci. Process. Impacts 2021, 23, 1255–1277. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Zaborowska, M.; Kucharski, J. The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities. Int. J. Mol. Sci. 2023, 24, 2892. [Google Scholar]
- Hirozawa, M.; Ono, M.; Suguiura, I.; Bordini, J.; Ono, E. Lactic acid bacteria and Bacillus spp. as fungal biological control agents. J. Appl. Microbiol. 2023, 34, lxac083. [Google Scholar] [CrossRef]
- Jaffar, N.; Jawan, R.; Chong, K. The potential of lactic acid bacteria in mediating the control of plant diseases and plant growth stimulation in crop production—A mini review. Front. Plant. Sci. 2023, 13, 1047945. [Google Scholar] [CrossRef]
- Chu, C.; Yu, L.; Li, Y.; Guo, H.; Zhai, Q.; Chen, W.; Tian, F. Lactobacillus plantarum CCFM405 against Rotenone-Induced Parkinson’s Disease Mice via Regulating Gut Microbiota and Branched-Chain Amino Acids Biosynthesis. Nutrients 2023, 15, 1737. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Cheng, S.; Chang, M.; Lin, Y.; Wu, C.; Tsai, Y. Neuroprotective Effects of Lactobacillus plantarum PS128 in a Mouse Model of Parkinson’s Disease: The Role of Gut Microbiota and MicroRNAs. Int. J. Mol. Sci. 2023, 24, 6794. [Google Scholar] [CrossRef] [PubMed]
- Trinder, M.; Bisanz, J.; Burton, J.; Reid, G. Probiotic lactobacilli: A potential prophylactic treatment for reducing pesticide absorption in humans and wildlife. Benef. Microb. 2015, 6, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Leska, A.; Nowak, A.; Miśkiewicz, K.; Rosicka-Kaczmarek, J. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment-An In Vitro Study. Cells 2022, 11, 3743. [Google Scholar] [CrossRef]
- Guo, X.; Xu, D.; Li, F.; Bai, J.; Su, R. Current approaches on the roles of lactic acid bacteria in crop silage. Microb. Biotechnol. 2023, 16, 67–87. [Google Scholar] [CrossRef]
- Haque, M.A.; Hong, S.Y.; Hwang, C.E.; Kim, S.C.; Cho, K.M. Cloning of an organophosphorus hydrolase (opdD) gene of Lactobacillus sakei WCP904 isolated from chlorpyrifos impregnated kimchi and hydrolysis activities of its gene product for organophosphorus pesticides. Appl. Biol. Chem. 2018, 61, 643–651. [Google Scholar] [CrossRef]
- Zeghal, E.; Vaksmaa, A.; Vielfaure, H.; Boekhout, T.; Niemann, H. The potential role of marine fungi in plastic degradation–A review. Front. Mar. Sci. 2021, 8, 738877. [Google Scholar] [CrossRef]
- Stief, P.; Fuchs-Ocklenburg, S.; Kamp, A.; Manohar, C.-S.; Houbraken, J.; Boekhout, T.; de Beer, D.; Stoeck, T. Dissimilatory nitrate reduction by Aspergillus terreus isolated from the seasonal oxygen minimum zone in the Arabian Sea. BMC Microbiol. 2014, 14, 35. [Google Scholar] [CrossRef]
- Goodell, B.; Winandy, J.E.; Morrell, J.J. Fungal degradation of wood: Emerging data, new insights and changing perceptions. Coatings 2020, 10, 1210. [Google Scholar] [CrossRef]
- Deshmukh, R.; Khardenavis, A.A.; Purohit, H.J. Diverse metabolic capacities of fungi for bioremediation. Indian J. Microbiol. 2016, 56, 247–264. [Google Scholar] [CrossRef]
- Maqbool, Z.; Hussain, S.; Imran, M.; Mahmood, F.; Shahzad, T.; Ahmed, Z.; Azeem, F.; Muzammil, S. Perspectives of using fungi as bioresource for bioremediation of pesticides in the environment: A critical review. Environ. Sci. Pollut. Res. Int. 2016, 23, 16904–16925. [Google Scholar] [CrossRef] [PubMed]
- Bokade, P.; Purohit, H.J.; Bajaj, A. Myco-remediation of Chlorinated Pesticides: Insights into Fungal Metabolic System. Indian J. Microbiol. 2021, 61, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Vaksmaa, A.; Guerrero-Cruz, S.; Ghosh, P.; Zeghal, E.; Hernando-Morales, V.; Niemann, H. Role of fungi in bioremediation of emerging pollutants. Front. Mar. Sci. 2023, 10, 1070905. [Google Scholar] [CrossRef]
- Nyakundi, W.; Magoma, G.; Ochora, J.; Nyende, A. Biodegradation of diazinon and methomyl pesticides by white rot fungi from selected horticultural farms in rift valley and central Kenya. J. Appl. Technol. Environ. Sanit. 2011, 1, 107–124. [Google Scholar]
- Ellegaard-Jensen, L.; Knudsen, B.E.; Johansen, A.; Albers, C.N.; Aamand, J.; Rosendahl, S. Fungal-bacterial consortia increase diuron degradation in water-unsaturated systems. Sci. Total Environ. 2014, 1, 466–705. [Google Scholar] [CrossRef]
- Knudsen, B.E.; Ellegaard-Jensen, L.; Albers, C.N.; Rosendahl, S.; Aamand, J. Fungal hyphae stimulate bacterial degradation of 2,6-dichlorobenzamide (BAM). Environ. Pollut. 2013, 181, 122–127. [Google Scholar] [CrossRef]
- El-Gendi, H.; Saleh, A.K.; Badierah, R.; Redwan, E.M.; El-Maradny, Y.A.; El-Fakharany, E.M. A comprehensive insight into fungal enzymes: Structure, classification, and their role in mankind’s challenges. J. Fungi 2021, 8, 23. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Hofrichter, M. Review: Lignin conversion by manganese peroxidase (MnP). Enzyme Microb. Technol. 2022, 30, 454–466. [Google Scholar] [CrossRef]
- Magan, N.; Fragoeiro, S.; Bastos, C. Environmental factors and bioremediation of xenobiotics using white rot fungi. Mycobiology 2010, 38, 238–248. [Google Scholar] [CrossRef]
- Zhuo, R.; Fan, F. A comprehensive insight into the application of white rot fungi and their lignocellulolytic enzymes in the removal of organic pollutants. Sci. Total Environ. 2021, 778, 146132. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Peris, A.; Torán, J.; Eljarrat, E.; Sarrà, M.; Blánquez, P.; Caminal, G. Exploring the degradation capability of trametes versicolor on selected hydrophobic pesticides through setting sights simultaneously on culture broth and biological matrix. Chemosphere 2020, 250, 126293. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Moreira, J.; Brugnari, T.; Sá-Nakanishi, A.; Castoldi, R.; Souza, C.; Bracht, A.; Peralta, R.M. Evaluation of diuron tolerance and biotransformation by the white-rot fungus Ganoderma lucidum. Fungal Biol. 2017, 122, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Wolfand, J.M.; Lefevre, G.H.; Luthy, R.G. Metabolization and degradation kinetics of the urban-use pesticide fipronil by white rot fungus Trametes versicolor. Environ. Sci. Process. Impacts 2016, 18, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Barbieri, M.V.; López-García, E.; Postigo, C.; López De Alda, M.; Caminal, G.; Sarrà, M. Fungal degradation of selected medium to highly polar pesticides by Trametes versicolor: Kinetics, biodegradation pathways, and ecotoxicity of treated waters. Anal. Bioanal. Chem. 2022, 414, 439–449. [Google Scholar] [CrossRef]
- Doddapaneni, H.; Chakraborty, R.; Yadav, J.S. Genome-wide structural and evolutionary analysis of the P450 monooxygenase genes (P450ome) in the white rot fungus Phanerochaete chrysosporium: Evidence for gene duplications and extensive gene clustering. BMC Genom. 2005, 6, 92. [Google Scholar] [CrossRef]
- Mori, T.; Ohno, H.; Ichinose, H.; Kawagishi, H.; Hirai, H. White-rot fungus Phanerochaete chrysosporium metabolizes chloropyridinyl-type neonicotinoid insecticides by an n-dealkylation reaction catalyzed by two cytochrome P450s. J. Hazard. Mater. 2021, 402, 123831. [Google Scholar] [CrossRef]
- Kullman, S.W.; Matsumura, F. Metabolic pathways utilized by Phanerochaete chrysosporium for degradation of the cyclodiene pesticide endosulfan. Appl. Environ. Microbiol. 1996, 62, 593–600. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Losito, I.; Facchini, L.; Katina, K.; Palmisano, F.; Gobbetti, M.; Coda, R. Degradation of vicine, convicine and their aglycones during fermentation of faba bean flour. Sci. Rep. 2016, 6, 32452. [Google Scholar] [CrossRef]
- Đorđević, T.; Đurović-Pejčev, R. The potency of Saccharomyces cerevisiae and Lactobacillus plantarum to dissipate organophosphorus pesticides in wheat during fermentation. J. Food Sci. Technol. 2016, 53, 4205–4215. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, B.; Yang, Z.; Kai, L.; Liu, Z. Alternative splicing and expression reduction of p450 genes mediating the oxidation of chlorpyrifos revealed a novel resistance mechanism in Nilaparvata lugens. J. Agric. Food Chem. 2023, 71, 4036–4042. [Google Scholar] [CrossRef]
- Daisley, B.; Trinder, M.; McDowell, T.; Collins, S.; Sumarah, M.; Reid, G. Microbiota-Mediated Modulation of Organophosphate Insecticide Toxicity by Species-Dependent Interactions with Lactobacilli in a Drosophila melanogaster Insect Model. Appl. Environ. Microbiol. 2018, 84, e02820-17. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Chuenchomrat, P.; Pereyron, V.; Haure, M.; Lorn, D.; Quan, L.-H.; Ho, P.-H.; Nguyen, T.-T.; Do, T.-Y.; Phi, Q.-T.; et al. Microorganisms, the Ultimate Tool for Clean Label Foods? Inventions 2021, 6, 31. [Google Scholar] [CrossRef]
- Laulund, S.; Wind, A.; Derkx, P.M.F.; Zuliani, V. Regulatory and safety requirements for food cultures. Microorganisms 2017, 5, 28. [Google Scholar] [CrossRef]
- Lechardeur, D.; Cesselin, B.; Fernandez, A.; Lamberet, G.; Garrigues, C.; Pedersen, M.; Gaudu, P.; Gruss, A. Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 2011, 22, 143–149. [Google Scholar] [CrossRef]
- Ly, M.H.; Vo, N.H.; Le, T.M.; Belin, J.-M.; Waché, Y. Diversity of the surface properties of Lactococci and consequences on adhesion to food components. Colloids Surf. B 2006, 52, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Adunphatcharaphon, S.; Petchkongkaew, A.; Visessanguan, W. In Vitro Mechanism Assessment of Zearalenone Removal by Plant-Derived Lactobacillus plantarum BCC 47723. Toxins 2021, 13, 286. [Google Scholar] [CrossRef]
- Trinder, M.; McDowell, T.; Daisley, B.; Ali, S.; Leong, H.; Sumarah, M.; Reid, G. Probiotic Lactobacillus rhamnosus reduces organophosphate pesticide absorption and toxicity to Drosophila melanogaster. Appl. Environ. Microbiol. 2016, 82, 6204–6213. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Szwajgier, D.; Waśko, A. Biosorption of Al+3 and Cd+2 by an exopolysaccharide from Lactobacillus rhamnosus. J. Food Sci. 2014, 79, T2404–T2408. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, A.; Bertilsson, S.; Goedkoop, W.E. Effects of extracellular polymeric and humic substances on chlorpyrifos bioavailability to Chironomus riparius. Ecotoxicology 2010, 19, 614–622. [Google Scholar] [CrossRef]
- López-Cabeza, R.; Francioso, A. Chiral pesticides with asymmetric sulfur: Extraction, separation, and determination in different environmental matrices. Separations 2022, 9, 29. [Google Scholar] [CrossRef]
- Boopathy, R. Factors limiting bioremediation technologies. Bioresour. Technol. 2000, 74, 63–67. [Google Scholar] [CrossRef]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase thatexhibit significant differences in redox potential substrate specificity, and stability. Biochim. Biophys. Acta 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Qin, X.; Xia, L. Degradation of the herbicide isoproturon by laccase mediator systems. Biochem. Eng. J. 2017, 119, 92–100. [Google Scholar] [CrossRef]
- Kupski, L.; Salcedo, G.M.; Caldas, S.S.; de Souza, T.D.; Furlong, E.B.; Primel, E.G. Optimization of a laccase-mediator system with natural redox-mediating compounds for pesticide removal. Environ. Sci. Pollut. Res. 2019, 26, 5131–5139. [Google Scholar] [CrossRef]
- Vidal-Limon, A.; García Suárez, P.C.; Arellano-García, E.; Contreras, O.E.; Aguila, S.A. Enhanced degradation of pesticide dichlorophen by laccase immobilized on nanoporous materials: A cytotoxic and molecular simulation investigation. Bioconjug. Chem. 2018, 29, 1073–1080. [Google Scholar] [CrossRef]
- Das, A.; Singh, J.; Yogalakshmi, K.N. Laccase immobilized magnetic iron nanoparticles: Fabrication and its performance evaluation in chlorpyrifos degradation. Int. Biodeter. Biodegr. 2017, 117, 183–189. [Google Scholar] [CrossRef]
- Du, Y.; Jia, X.; Zhong, L.; Jiao, Y.; Zhang, Z.; Wang, Z.; Feng, Y.; Bila, M.; Cui, J.; Jia, S. Metal-organic frameworks with different dimensionalities: An ideal host platform for enzyme@MOF composites. Coord. Chem. Rev. 2022, 454, 214327. [Google Scholar] [CrossRef]
- Ladole, M.R.; Pokale, P.B.; Patil, S.S.; Belokar, P.G.; Pandit, A.B. Laccase immobilized peroxidase mimicking magnetic metal organic frameworks for industrial dye degradation. Bioresour Technol. 2020, 317, 124035. [Google Scholar] [CrossRef]
- Shen, X.; Du, Y.; Du, Z.; Tang, X.; Li, P.; Cheng, J.; Yan, R.; Cui, J. Construction of enzyme@glutathione hybrid metal-organicframeworks: Glutathione-boosted microenvironment fine-tuning of biomimetic immobilization for improving catalytic performance. Mater. Today Chem. 2023, 27, 101326. [Google Scholar] [CrossRef]
- Jain, M.; Yadav, P.; Joshi, A.; Kodgire, P. Advances in detection of hazardous organophosphorus compounds using organophosphorus hydrolase based biosensors. Crit. Rev. Toxicol. 2019, 49, 387–410. [Google Scholar] [CrossRef] [PubMed]
- Vyas, T.; Singh, V.; Kodgire, P.; Joshi, A. Insights in detection and analysis of organophosphates using organophosphorus acid anhydrolases (OPAA) enzyme-based biosensors. Crit. Rev. Biotechnol. 2023, 43, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, P.; Wang, Z.; Du, Y.; Kuang, G.; Feng, Y.; Jia, S.; Cui, J. Glutamate oxidase-integrated biomimetic metal–organic framework hybrids as cascade nanozymes for ultrasensitive glutamate detection. J. Agric. Food Chem. 2022, 70, 3785–3794. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Ganai, B. Deciphering the recent trends in pesticide bioremediation using genome editing and multi-omics approaches: A review. World J. Microbiol. Biotechnol. 2023, 39, 151. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | Pesticide | Microbes Involved |
|---|---|---|
| Dairy products | Dimethoate, fenthion, malathion, methyl parathion, monocrotophos, phorate, trichlorphon | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, yeasts |
| α-hexachlorocyclohexane (HCH), hexachlorobenzene (HCB), ϒ-HCH, β-chlordane, α-chlordane | Lactobacillus acidophilus, L. delbrueckii subsp. bulgaricus, Lactiplantibacillus plantarum, Lacticaseibacillus rhamnosus, Lacticaseibacillus casei, S. thermophilus, Bifidobacterium bifidum | |
| α-HCH, β-HCH, ϒ-HCH, 1,1-bis-(4-chlorophenyl)-2,2-dichloroethene, 1-chloro-4-[2,2-dichloro-1-(4-chlorophenyl) ethyl] benzene, 1,10-(2,2,2-trichloroethane-1,1-diyl) bis (4-chlorobenzene) | L. acidophilus, Bif. animalis subsp. lactis | |
| Dimethoate | Lp. plantarum | |
| Chlorpyrifos, chlorpyrifos-methyl, diazinon, dichlorvos, fenthion, malathion, phorate, pirimiphos-methyl, trichlorphon | L. delbrueckii subsp. bulgaricus, L. acidophilus, Lc. casei, Lc. rhamnosus, S. thermophilus | |
| Chlorpyrifos, diazinon, fenitrothion, malathion, methyl parathion | Lp. plantarum, Levilactobacillus brevis, Lactobacillus helveticus, Lactobacillus lactis, L. delbrueckii subsp. bulgaricus, S. thermophilus | |
| Dimethoate, fenthion, malathion, methyl parathion, monocrotophos, phorate, trichlorphon | L. bulgaricus, Lacticaseibacillus paracasei, Lp. plantarum | |
| Vegetables | Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | Leuconostoc mesenteroides, Lev. brevis, Lp. plantarum, L. sakei |
| Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | Lev. brevis | |
| Chlorpyrifos, dichlorvos, phorate, trichlorphon, deltamethrin, cyhalothrin, malathion, chlorpyrifos-methyl, deltamethrin, dimethoate, Imidacloprid | Lp. plantarum | |
| Cereals and bakery products | Chlorpyrifos, phorate, pirimiphos, pirimiphos-methyl, bifenthrin | Lp. plantarum |
| Glyphosate, endosulfan, deltamethrin, malathion, propiaconazole, chlorpyriphos, hexaconazole, pirimiphos methyl | S. cerevisiae | |
| Beverages | β-cypermethrin, 3-phenoxybenzoic acid | Aspergillus niger |
| Chlorpyrifos, dicofol, chlorothalonil, procymidone | Oenococcus oeni | |
| Tebuconazole | Saccharomyces cerevisiae, O. oeni | |
| Diazinon | L. acidophilus | |
| Meat products | 1,1,1-trichloro-2,2-bis (4-chlorophenyl) ethane (DDT), lindane | Lp. plantarum, Micrococcus varians |
| Fungi | Pesticide Degraded | Category | Degradation Rate (%) |
|---|---|---|---|
| Trametes versicolor | Chlorpyrifos | OP | 94.7 |
| Dicofol | OC | 87.9 | |
| Cypermethrin | Pyrethroid | 93.1 | |
| Diuron | Herbicide | 93 | |
| Bentazon | Herbicide | 93 | |
| Phlebia brevispora | Heptachlor | OC | 74 |
| Heptachlor epoxide | OC | 16 | |
| Aldrin | OC | 97.6 | |
| Aspergillus niger | Heptachlor | OC | - |
| A-endosulfan | OC | 100 | |
| Phlebia aurea | Heptachlor epoxide | OC | 25 |
| Aldrin | OC | 96.4 | |
| Dieldrin | OC | 54 | |
| Fusarium ventricosum | α-endosulfan | OC | 91 |
| β-endosulfan | OC | 89.9 | |
| Phlebia acanthocystis | Heptachlor | OC | 90 |
| Heptachlor epoxide | OC | 16 | |
| Aldrin | OC | 96 | |
| Dieldrin | OC | 56 | |
| Ganoderma lucidum | Diuron | Herbicide | >50 |
| Lindane | OC | 75.5 | |
| Penicillium miczynskii | Dieldrin | OC | 90 |
| Trichoderma hamatum | Dichlorodiphenyltrichloroethane (DDT) | OC | - |
| Phanerochete chrysosporium | Chlordane | OC | - |
| DDT | OC | - | |
| Aldrin | OC | - | |
| Dieldrin | OC | - | |
| Endosulfan | OC | - | |
| Heptachlor | OC | - | |
| Parathion | OP | >90% | |
| Pentachlorophenol | OC | - | |
| Hexachlorocyclohexane | OC | - | |
| Lindane | OC | >10 | |
| Aspergillus sydowii | Methyl parathion | OP | 80 |
| Chlorpyrifos | OP | 32 | |
| Profenofos | OP | 52 | |
| Penicillium citrinum | Methyl parathion | OP | 100 |
| Enzyme | Reaction Type | Site of Reaction | Pesticides Degraded |
|---|---|---|---|
| Laccases | Oxidation of phenolic substrates, requiring only atmospheric molecular oxygen as their co-substrate | Extra-cellular | Pesticides having a phenolic ring structure. Laccases can also catalyze the breakdown of non-phenolic molecules in the presence of mediator systems |
| Manganese peroxidase | Oxidation, with H2O2 as terminal electron acceptor | Chlorophenol, lindane, tribromophenol, glyphosate | |
| P450 Oxidases | Epoxidations, hydroxylation | Intra-cellular | Lindane, DDT, Endrin, Fibronil, Alachlor, Acetamiprid, Clothianidin, Fenazaquin, Diuron, Chlorothalonil, Cyhalothrin |
| Transferases | Removal of hydroxyl groups to produce conjugates | Chlorpyrifos, methyl parathion, and profenofos | |
| Aromatic nitro reductases | Reductions of nitro groups for further extra-cellular degradation | Picloram, bentazon, and carbofuran |
| Parameter | Mechanism |
|---|---|
| Temperature | High temperatures can denature or modify the characteristics of microbial enzymes, which are sensitive to heat. For instance, the ideal temperature for laccases is between 20 and 40 °C, while the ideal temperature for lipases and peroxidases is between 15 and 30 °C. |
| pH | The ideal pH varies depending on the enzyme. Lower enzyme activity occurs when the pH value is outside the ideal range. The majority of enzymes operate in the neutral pH range. Manganese peroxidase, on the other hand, performs best in a pH range of 4–5, whereas some enzymes, such as alkaline proteases, function at pH levels above 8.0. |
| Enzyme concentration | Bioremediation depends heavily on the enzymatic dose used for the degradation process. Typically, when enzyme concentrations increase, the breakdown efficiency increases up to a certain degree. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perpetuini, G.; Nzobouh Fossi, P.A.; Kwak, S.; Namessi, O.; Petchkongkaew, A.; Tofalo, R.; Waché, Y. Pesticides in Foods: Towards Bioremediation Biocatalysts? Catalysts 2023, 13, 1055. https://doi.org/10.3390/catal13071055
Perpetuini G, Nzobouh Fossi PA, Kwak S, Namessi O, Petchkongkaew A, Tofalo R, Waché Y. Pesticides in Foods: Towards Bioremediation Biocatalysts? Catalysts. 2023; 13(7):1055. https://doi.org/10.3390/catal13071055
Chicago/Turabian StylePerpetuini, Giorgia, Pilar Anais Nzobouh Fossi, Seyi Kwak, Oscar Namessi, Awanwee Petchkongkaew, Rosanna Tofalo, and Yves Waché. 2023. "Pesticides in Foods: Towards Bioremediation Biocatalysts?" Catalysts 13, no. 7: 1055. https://doi.org/10.3390/catal13071055
APA StylePerpetuini, G., Nzobouh Fossi, P. A., Kwak, S., Namessi, O., Petchkongkaew, A., Tofalo, R., & Waché, Y. (2023). Pesticides in Foods: Towards Bioremediation Biocatalysts? Catalysts, 13(7), 1055. https://doi.org/10.3390/catal13071055