Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Drug-Sensitive Cancer Cells and Multidrug-Resistant Cancer Cells Overexpressing ABCB1 or ABCG2 Are Equally Sensitive to Sitravatinib
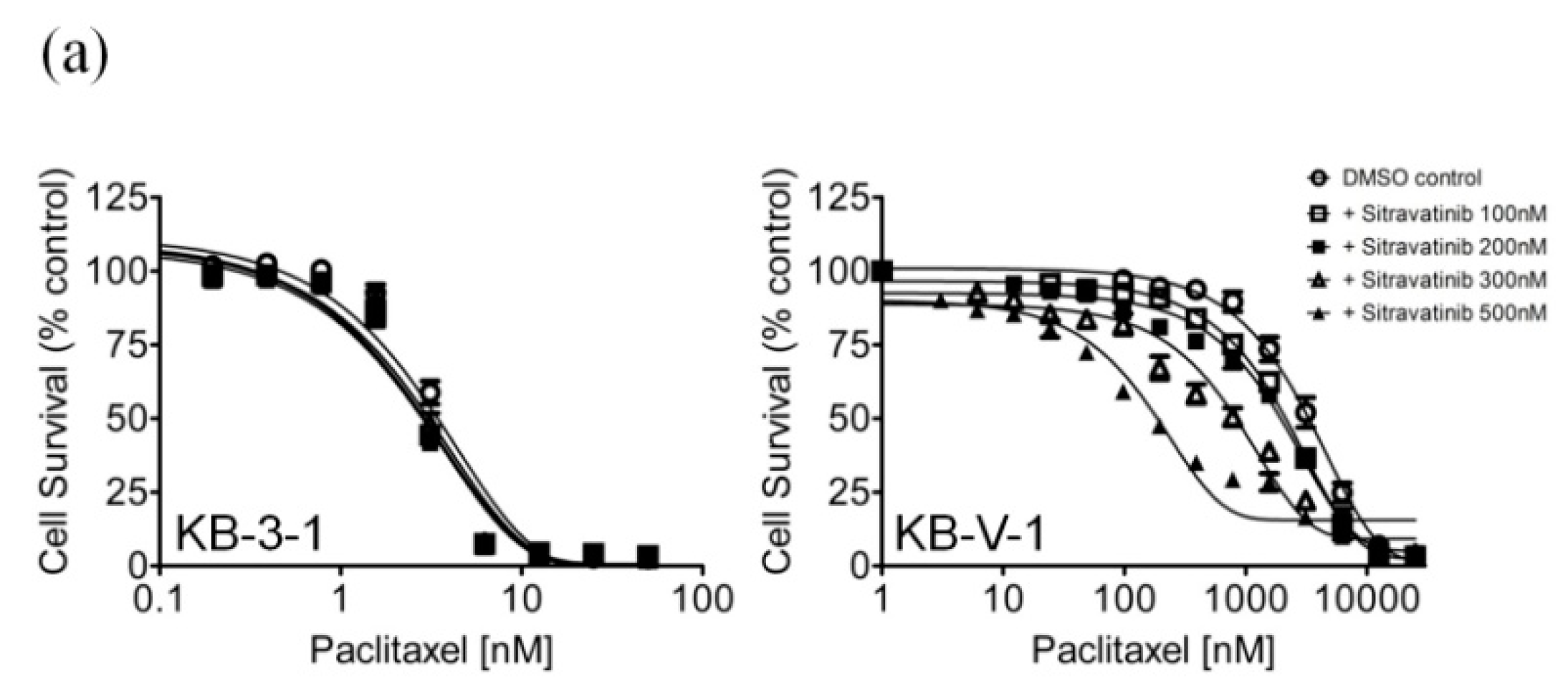
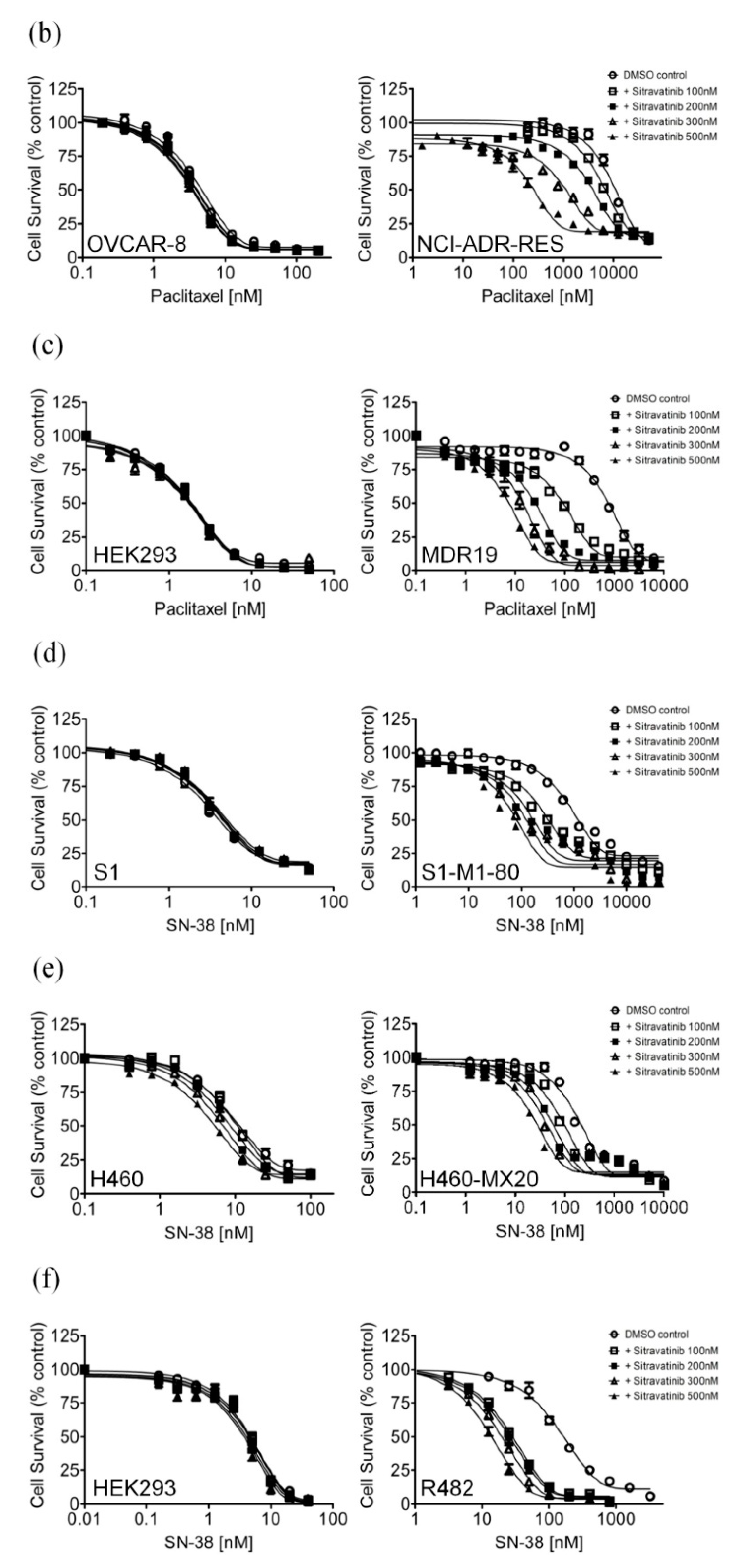
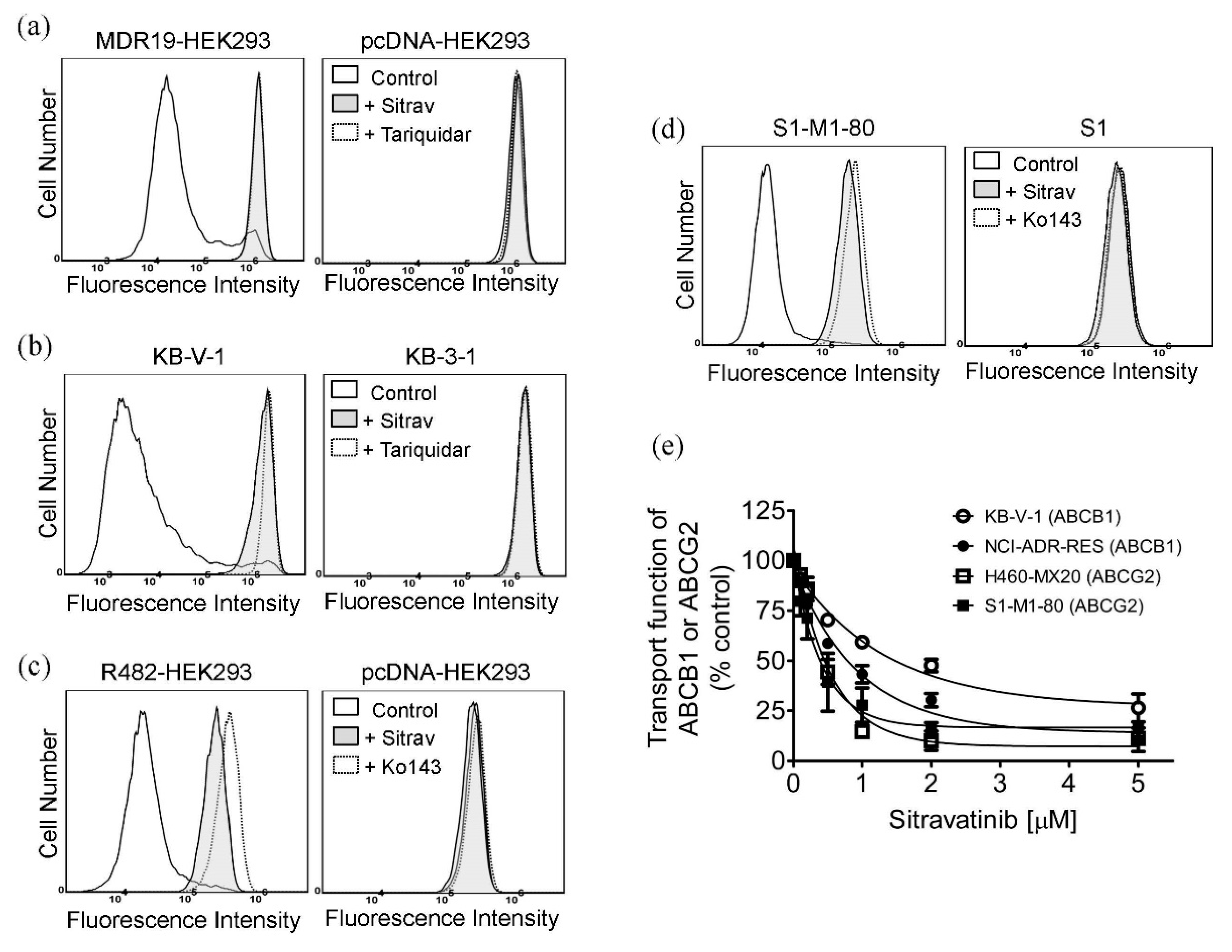
2.2. Sitravatinib Reverses Multidrug Resistance Mediated by ABCB1 and ABCG2
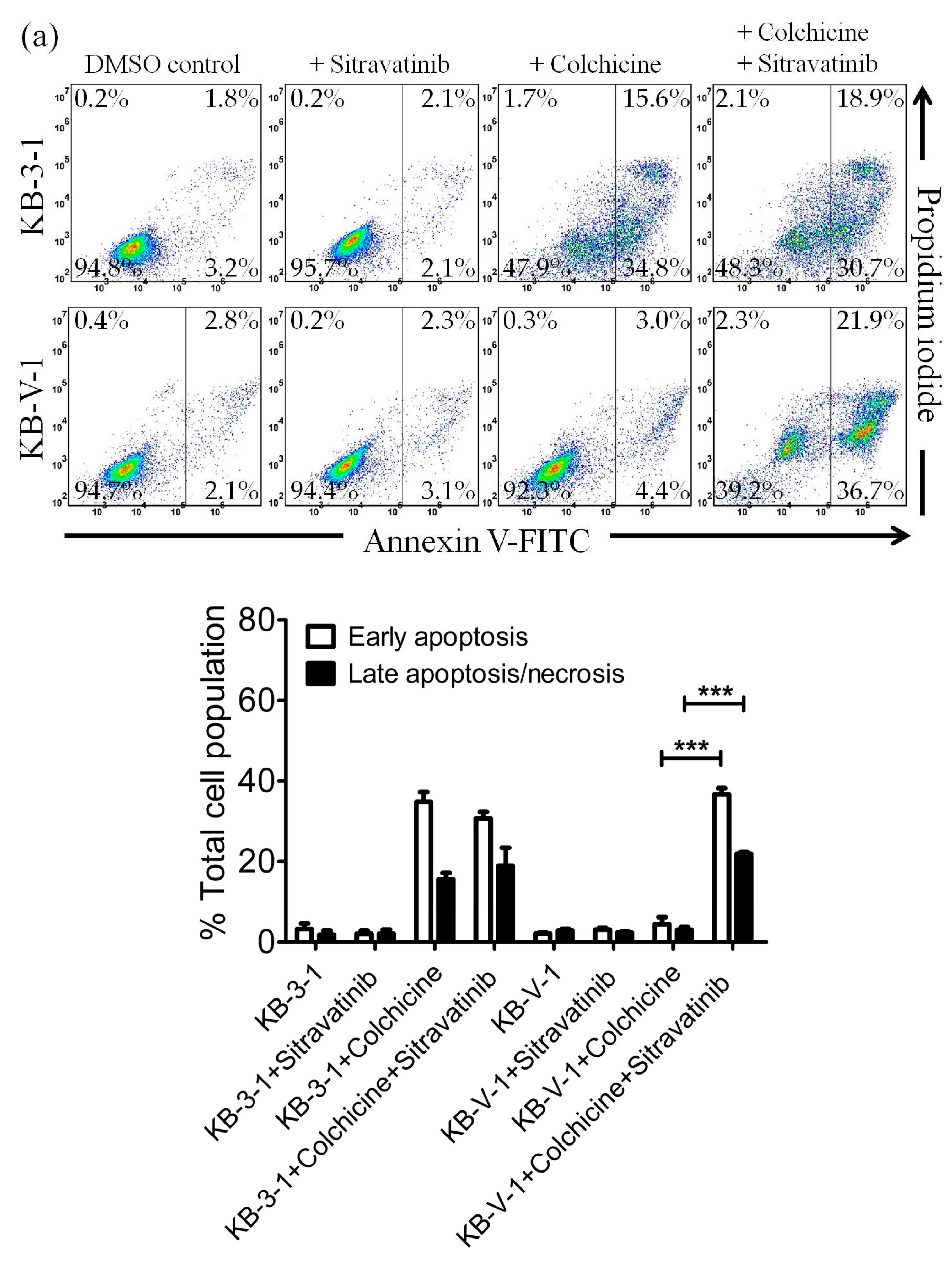
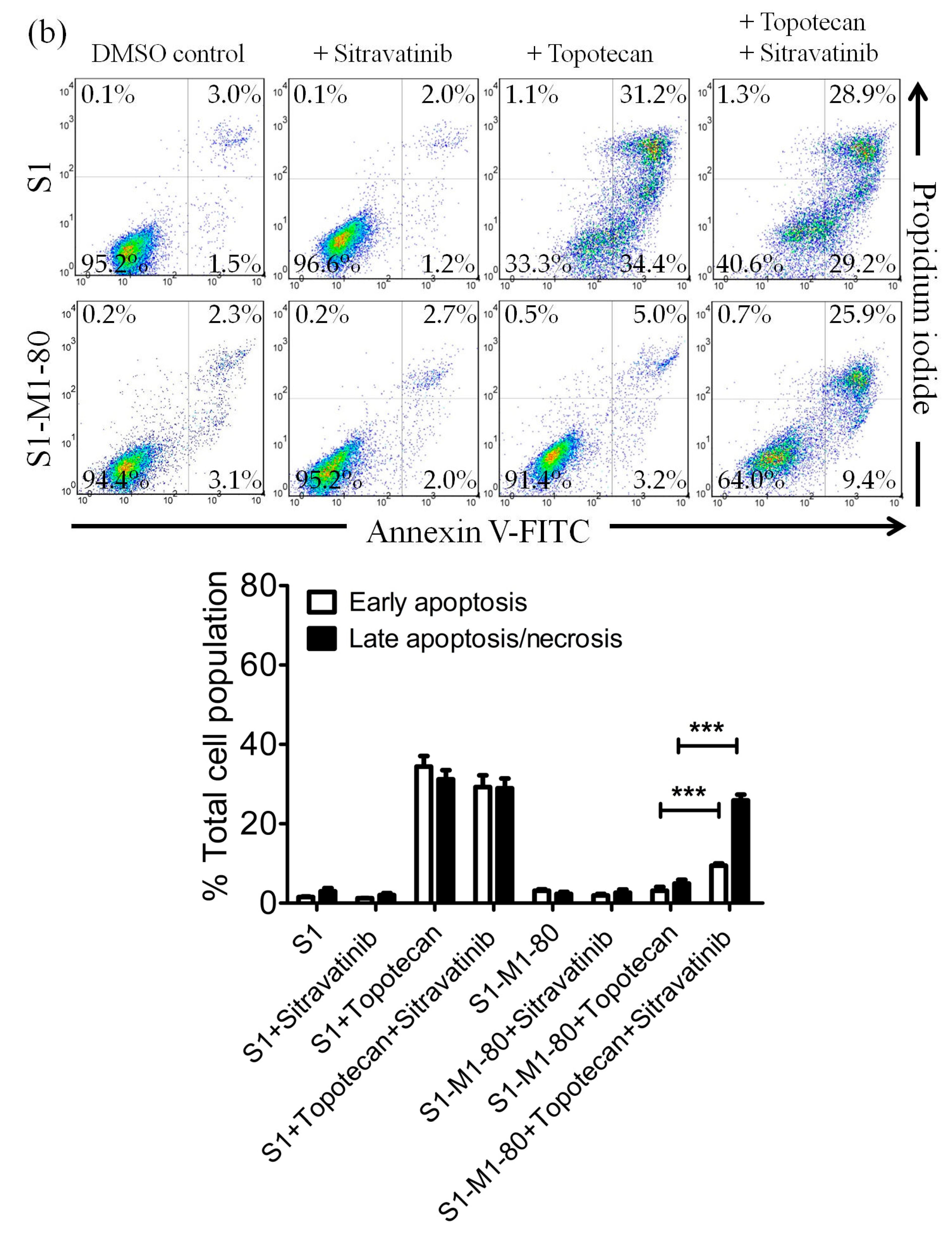
2.3. Sitravatinib Restores Sensitivity for Drug-Induced Apoptosis in ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells
2.4. Sitravatinib Increases Drug Accumulation in Cells Overexpressing ABCB1 or ABCG2
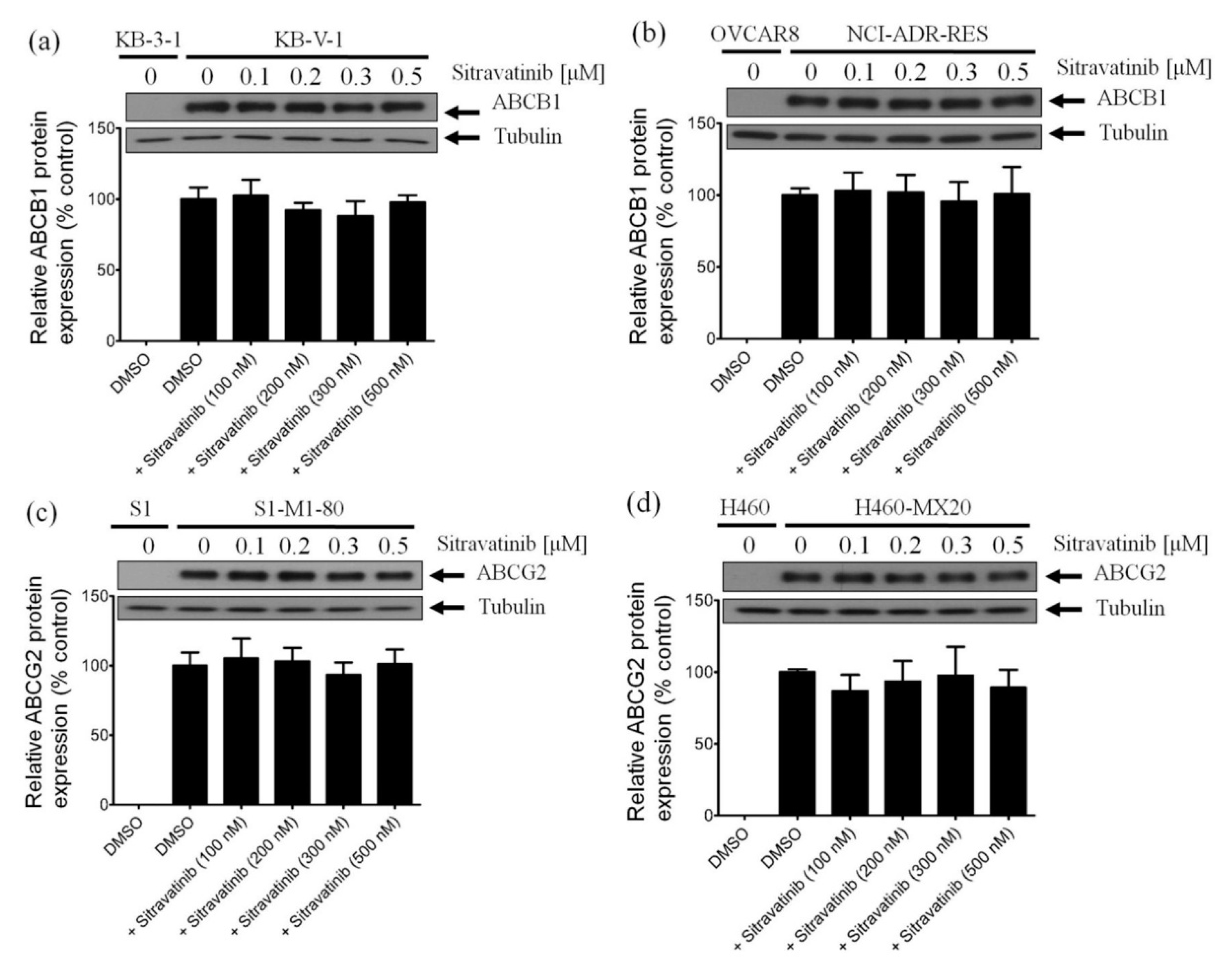
2.5. Sitravatinib Does Not Alter the Protein Expression of ABCB1 or ABCG2 in Multidrug-Resistant Cancer Cells
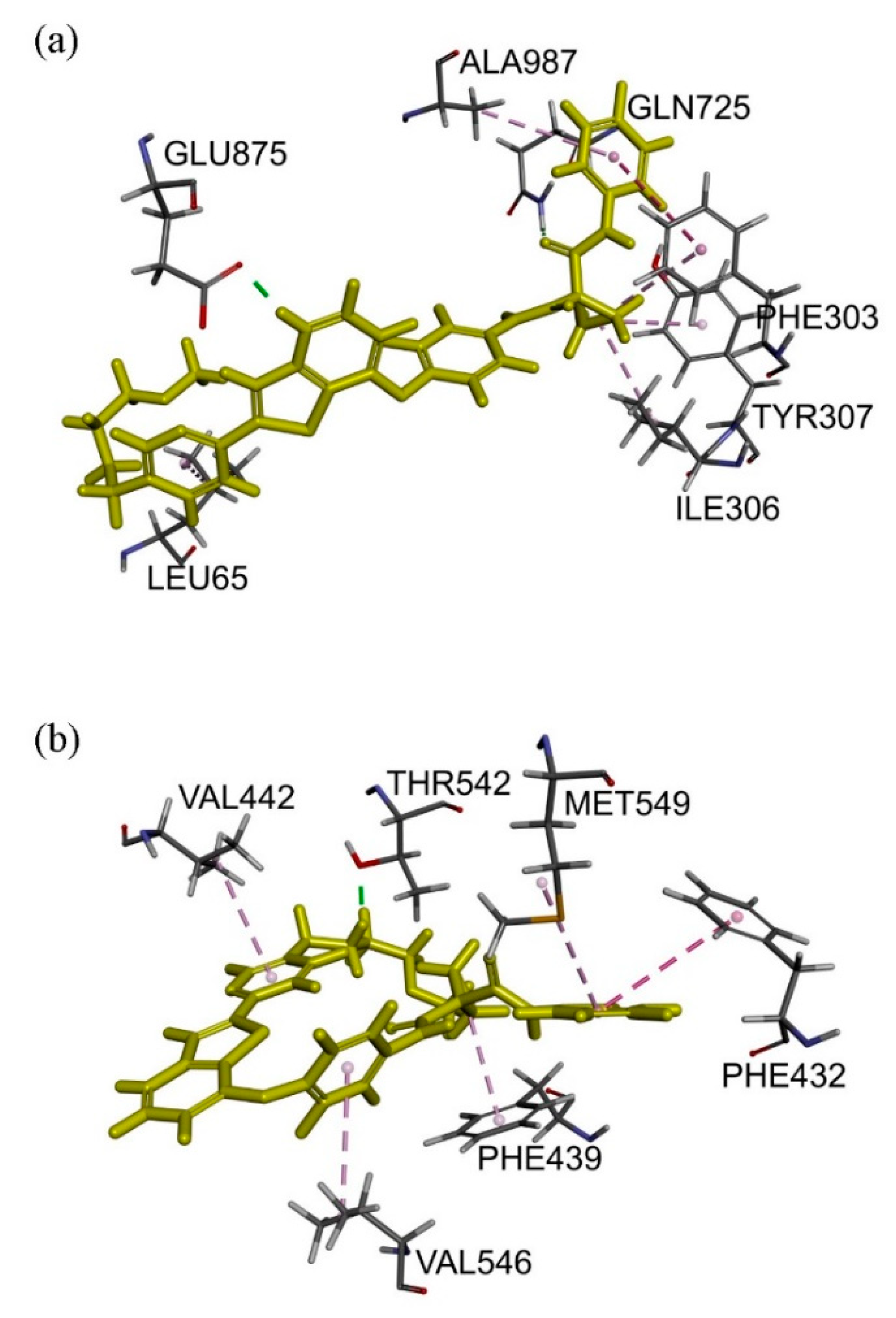
2.6. Docking Analysis of Sitravatinib with Structures of ABCB1 and ABCG2
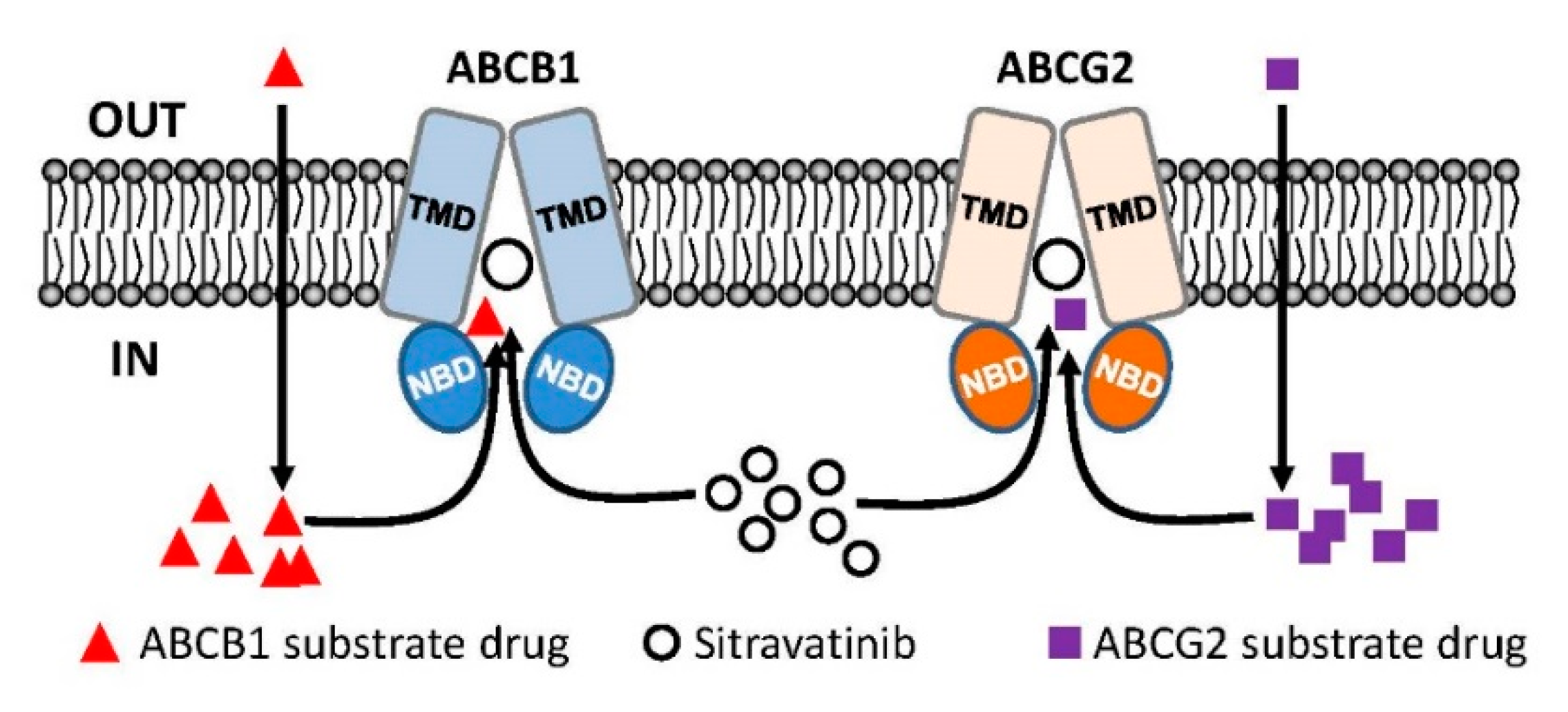
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture Conditions
4.3. Cell Viability Assay
4.4. Annexin V/Propidium Iodide Apoptosis Assay
4.5. Fluorescent Drug Accumulation Assay
4.6. Immunoblotting
4.7. In Silico Docking of Sitravatinib in the Drug-Binding Pockets of ABCB1 and ABCG2
4.8. Quantification and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gottesman, M.; Ambudkar, S.V. Overview: ABC transporters and human disease. J. Bioenergy Biomembr. 2001, 33, 453–458. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Gillet, J.P.; Gottesman, M.M. Mechanisms of multidrug resistance in cancer. Methods Mol. Biol. 2010, 596, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.P.; Hsieh, C.H.; Wu, Y.S. The emergence of drug transporter-mediated multidrug resistance to cancer chemotherapy. Mol. Pharm. 2011, 8, 1996–2011. [Google Scholar] [CrossRef] [PubMed]
- Szakacs, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. 2006, 5, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, K.; Katayama, K.; Sugimoto, Y. Human ABC transporter ABCG2/BCRP expression in chemoresistance: basic and clinical perspectives for molecular cancer therapeutics. Pharmgenomics. Pers. Med. 2014, 7, 53–64. [Google Scholar] [CrossRef]
- Kovalev, A.A.; Tsvetaeva, D.A.; Grudinskaja, T.V. Role of ABC-cassette transporters (MDR1, MRP1, BCRP) in the development of primary and acquired multiple drug resistance in patients with early and metastatic breast cancer. Exp. Oncol. 2013, 35, 287–290. [Google Scholar]
- Schwarzenbach, H. Expression of MDR1/P-glycoprotein, the multidrug resistance protein MRP, and the lung-resistance protein LRP in multiple myeloma. Med. Oncol. 2002, 19, 87–104. [Google Scholar] [CrossRef]
- Tsubaki, M.; Satou, T.; Itoh, T.; Imano, M.; Komai, M.; Nishinobo, M.; Yamashita, M.; Yanae, M.; Yamazoe, Y.; Nishida, S. Overexpression of MDR1 and survivin, and decreased Bim expression mediate multidrug-resistance in multiple myeloma cells. Leuk. Res. 2012, 36, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.G.; Gump, J.L.; Zhang, C.; Cook, J.M.; Marchion, D.; Hazlehurst, L.; Munster, P.; Schell, M.J.; Dalton, W.S.; Sullivan, D.M. ABCG2 expression, function, and promoter methylation in human multiple myeloma. Blood 2006, 108, 3881–3889. [Google Scholar] [CrossRef]
- Matthews, C.; Catherwood, M.A.; Larkin, A.M.; Clynes, M.; Morris, T.C.; Alexander, H.D. MDR-1, but not MDR-3 gene expression, is associated with unmutated IgVH genes and poor prognosis chromosomal aberrations in chronic lymphocytic leukemia. Leuk. Lymphoma 2006, 47, 2308–2313. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.D.; Karp, J.E.; Chen, T.T.; Doyle, L.A. Expression of breast cancer resistance protein in blast cells from patients with acute leukemia. Blood 2000, 96, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, D.; Sell, W.; Voigt, A.; Hermann, J.; Zintl, F.; Sauerbrey, A. BCRP gene expression is associated with a poor response to remission induction therapy in childhood acute myeloid leukemia. Leukemia 2002, 16, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Uggla, B.; Stahl, E.; Wagsater, D.; Paul, C.; Karlsson, M.G.; Sirsjo, A.; Tidefelt, U. BCRP mRNA expression v. clinical outcome in 40 adult AML patients. Leuk. Res. 2005, 29, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Schinkel, A.H. Multidrug resistance and pharmacological protection mediated by the breast cancer resistance protein (BCRP/ABCG2). Mol. Cancer 2002, 1, 427–434. [Google Scholar]
- Kannan, P.; Telu, S.; Shukla, S.; Ambudkar, S.V.; Pike, V.W.; Halldin, C.; Gottesman, M.M.; Innis, R.B.; Hall, M.D. The “specific” P-glycoprotein inhibitor Tariquidar is also a substrate and an inhibitor for breast cancer resistance protein (BCRP/ABCG2). ACS Chem. Neurosci. 2011, 2, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Weidner, L.D.; Zoghbi, S.S.; Lu, S.; Shukla, S.; Ambudkar, S.V.; Pike, V.W.; Mulder, J.; Gottesman, M.M.; Innis, R.B.; Hall, M.D. The Inhibitor Ko143 Is Not Specific for ABCG2. J. Pharm. Exp. 2015, 354, 384–393. [Google Scholar] [CrossRef]
- Shukla, S.; Wu, C.P.; Ambudkar, S.V. Development of inhibitors of ATP-binding cassette drug transporters: Present status and challenges. Expert Opin. Drug Metab. Toxicol. 2008, 4, 205–223. [Google Scholar] [CrossRef]
- Wu, C.P.; Calcagno, A.M.; Ambudkar, S.V. Reversal of ABC drug transporter-mediated multidrug resistance in cancer cells: Evaluation of current strategies. Curr. Mol. Pharmacol. 2008, 1, 93–105. [Google Scholar] [CrossRef]
- Shi, Z.; Tiwari, A.K.; Shukla, S.; Robey, R.W.; Singh, S.; Kim, I.W.; Bates, S.E.; Peng, X.; Abraham, I.; Ambudkar, S.V.; et al. Sildenafil reverses ABCB1- and ABCG2-mediated chemotherapeutic drug resistance. Cancer Res. 2011, 71, 3029–3041. [Google Scholar] [CrossRef]
- Shukla, S.; Chen, Z.S.; Ambudkar, S.V. Tyrosine kinase inhibitors as modulators of ABC transporter-mediated drug resistance. Drug Resist Updates 2012, 15, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.K.; Sodani, K.; Dai, C.L.; Abuznait, A.H.; Singh, S.; Xiao, Z.J.; Patel, A.; Talele, T.T.; Fu, L.; Kaddoumi, A.; et al. Nilotinib potentiates anticancer drug sensitivity in murine ABCB1-, ABCG2-, and ABCC10-multidrug resistance xenograft models. Cancer Lett. 2013, 328, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Q.; Liu, S.T.; Zhao, B.X.; Yang, F.H.; Wang, Y.T.; Liang, Q.Y.; Sun, Y.B.; Liu, Y.; Song, Z.H.; Cai, Y.; et al. Afatinib reverses multidrug resistance in ovarian cancer via dually inhibiting ATP binding cassette subfamily B member 1. Oncotarget 2015, 6, 26142–26160. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, S.H.; Lu, Y.J.; Li, Y.Q.; Huang, Y.H.; Hsieh, C.H.; Wu, C.P. Osimertinib (AZD9291) Attenuates the Function of Multidrug Resistance-Linked ATP-Binding Cassette Transporter ABCB1 in Vitro. Mol. Pharm. 2016, 13, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, P.P.; Ivy, K.S.; Musi, E.; de Stanchina, E.; Schwartz, G.K. Significant blockade of multiple receptor tyrosine kinases by MGCD516 (Sitravatinib), a novel small molecule inhibitor, shows potent anti-tumor activity in preclinical models of sarcoma. Oncotarget 2016, 7, 4093–4109. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Wang, L.; Clark, R.E.; Pirmohamed, M. Active transport of imatinib into and out of cells: Implications for drug resistance. Blood 2004, 104, 3739–3745. [Google Scholar] [CrossRef]
- Breedveld, P.; Pluim, D.; Cipriani, G.; Wielinga, P.; van Tellingen, O.; Schinkel, A.H.; Schellens, J.H. The effect of Bcrp1 (Abcg2) on the in vivo pharmacokinetics and brain penetration of imatinib mesylate (Gleevec): Implications for the use of breast cancer resistance protein and P-glycoprotein inhibitors to enable the brain penetration of imatinib in patients. Cancer Res. 2005, 65, 2577–2582. [Google Scholar]
- Burger, H.; Van Tol, H.; Brok, M.; Wiemer, E.A.; De Bruijn, E.A.; Guetens, G.; De Boeck, G.; Sparreboom, A.; Verweij, J.; Nooter, K. Chronic imatinib mesylate exposure leads to reduced intracellular drug accumulation by induction of the ABCG2 (BCRP) and ABCB1 (MDR1) drug transport pumps. Cancer Biol. 2005, 4, 747–752. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shiozawa, K.; Hassel, B.A.; Ross, D.D. Complex interaction of BCRP/ABCG2 and imatinib in BCR-ABL-expressing cells: BCRP-mediated resistance to imatinib is attenuated by imatinib-induced reduction of BCRP expression. Blood 2006, 108, 678–684. [Google Scholar] [CrossRef]
- Mizuno, T.; Fukudo, M.; Terada, T.; Kamba, T.; Nakamura, E.; Ogawa, O.; Inui, K.; Katsura, T. Impact of genetic variation in breast cancer resistance protein (BCRP/ABCG2) on sunitinib pharmacokinetics. Drug Metab. Pharm. 2012, 27, 631–639. [Google Scholar] [CrossRef]
- Sato, H.; Siddig, S.; Uzu, M.; Suzuki, S.; Nomura, Y.; Kashiba, T.; Gushimiyagi, K.; Sekine, Y.; Uehara, T.; Arano, Y.; et al. Elacridar enhances the cytotoxic effects of sunitinib and prevents multidrug resistance in renal carcinoma cells. Eur. J. Pharm. 2015, 746, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Mahon, F.X.; Belloc, F.; Lagarde, V.; Chollet, C.; Moreau-Gaudry, F.; Reiffers, J.; Goldman, J.M.; Melo, J.V. MDR1 gene overexpression confers resistance to imatinib mesylate in leukemia cell line models. Blood 2003, 101, 2368–2373. [Google Scholar] [CrossRef] [PubMed]
- Mahon, F.X.; Hayette, S.; Lagarde, V.; Belloc, F.; Turcq, B.; Nicolini, F.; Belanger, C.; Manley, P.W.; Leroy, C.; Etienne, G.; et al. Evidence that resistance to nilotinib may be due to BCR-ABL, Pgp, or Src kinase overexpression. Cancer Res. 2008, 68, 9809–9816. [Google Scholar] [CrossRef] [PubMed]
- Hiwase, D.K.; Saunders, V.; Hewett, D.; Frede, A.; Zrim, S.; Dang, P.; Eadie, L.; To, L.B.; Melo, J.; Kumar, S.; et al. Dasatinib cellular uptake and efflux in chronic myeloid leukemia cells: Therapeutic implications. Clin. Cancer Res. 2008, 14, 3881–3888. [Google Scholar] [CrossRef] [PubMed]
- Beretta, G.L.; Cassinelli, G.; Pennati, M.; Zuco, V.; Gatti, L. Overcoming ABC transporter-mediated multidrug resistance: The dual role of tyrosine kinase inhibitors as multitargeting agents. Eur. J. Med. Chem. 2017, 142, 271–289. [Google Scholar] [CrossRef]
- Wu, S.; Fu, L. Tyrosine kinase inhibitors enhanced the efficacy of conventional chemotherapeutic agent in multidrug resistant cancer cells. Mol. Cancer 2018, 17, 25. [Google Scholar] [CrossRef]
- Kartner, N.; Riordan, J.R.; Ling, V. Cell surface P-glycoprotein associated with multidrug resistance in mammalian cell lines. Science 1983, 221, 1285–1288. [Google Scholar] [CrossRef]
- Riordan, J.R.; Ling, V. Purification of P-glycoprotein from plasma membrane vesicles of Chinese hamster ovary cell mutants with reduced colchicine permeability. J. Biol. Chem. 1979, 254, 12701–12705. [Google Scholar]
- Bates, S.E.; Medina-Perez, W.Y.; Kohlhagen, G.; Antony, S.; Nadjem, T.; Robey, R.W.; Pommier, Y. ABCG2 mediates differential resistance to SN-38 (7-ethyl-10-hydroxycamptothecin) and homocamptothecins. J. Pharm. Exp. 2004, 310, 836–842. [Google Scholar] [CrossRef]
- Miyake, K.; Mickley, L.; Litman, T.; Zhan, Z.; Robey, R.; Cristensen, B.; Brangi, M.; Greenberger, L.; Dean, M.; Fojo, T.; et al. Molecular cloning of cDNAs which are highly overexpressed in mitoxantrone-resistant cells: Demonstration of homology to ABC transport genes. Cancer Res. 1999, 59, 8–13. [Google Scholar]
- Maliepaard, M.; Van Gastelen, M.A.; De Jong, L.A.; Pluim, D.; Van Waardenburg, R.C.; Ruevekamp-Helmers, M.C.; Floot, B.G.; Schellens, J.H. Overexpression of the BCRP/MXR/ABCP gene in a topotecan-selected ovarian tumor cell line. Cancer Res. 1999, 59, 4559–4563. [Google Scholar] [PubMed]
- Dai, C.L.; Tiwari, A.K.; Wu, C.P.; Su, X.D.; Wang, S.R.; Liu, D.G.; Ashby, C.R., Jr.; Huang, Y.; Robey, R.W.; Liang, Y.J.; et al. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res. 2008, 68, 7905–7914. [Google Scholar] [CrossRef]
- Scheffer, G.L.; Maliepaard, M.; Pijnenborg, A.C.; Van Gastelen, M.A.; De Jong, M.C.; Schroeijers, A.B.; Van der Kolk, D.M.; Allen, J.D.; Ross, D.D.; Van der Valk, P.; et al. Breast cancer resistance protein is localized at the plasma membrane in mitoxantrone- and topotecan-resistant cell lines. Cancer Res. 2000, 60, 2589–2593. [Google Scholar]
- Wu, C.P.; Hsiao, S.H.; Su, C.Y.; Luo, S.Y.; Li, Y.Q.; Huang, Y.H.; Hsieh, C.H.; Huang, C.W. Human ATP-Binding Cassette transporters ABCB1 and ABCG2 confer resistance to CUDC-101, a multi-acting inhibitor of histone deacetylase, epidermal growth factor receptor and human epidermal growth factor receptor 2. Biochem. Pharm. 2014, 92, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Hollo, Z.; Homolya, L.; Davis, C.W.; Sarkadi, B. Calcein accumulation as a fluorometric functional assay of the multidrug transporter. Biochim. Biophys. Acta 1994, 1191, 384–388. [Google Scholar] [CrossRef]
- Robey, R.W.; Steadman, K.; Polgar, O.; Morisaki, K.; Blayney, M.; Mistry, P.; Bates, S.E. Pheophorbide a is a specific probe for ABCG2 function and inhibition. Cancer Res. 2004, 64, 1242–1246. [Google Scholar] [CrossRef]
- Cuestas, M.L.; Castillo, A.I.; Sosnik, A.; Mathet, V.L. Downregulation of MDR1 and abcg2 genes is a mechanism of inhibition of efflux pumps mediated by polymeric amphiphiles. Bioorg. Med. Chem. Lett. 2012, 22, 6577–6579. [Google Scholar] [CrossRef]
- Natarajan, K.; Bhullar, J.; Shukla, S.; Burcu, M.; Chen, Z.S.; Ambudkar, S.V.; Baer, M.R. The Pim kinase inhibitor SGI-1776 decreases cell surface expression of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) and drug transport by Pim-1-dependent and -independent mechanisms. Biochem. Pharm. 2013, 85, 514–524. [Google Scholar] [CrossRef]
- Wu, C.P.; Shukla, S.; Calcagno, A.M.; Hall, M.D.; Gottesman, M.M.; Ambudkar, S.V. Evidence for dual mode of action of a thiosemicarbazone, NSC73306: A potent substrate of the multidrug resistance linked ABCG2 transporter. Mol. Cancer 2007, 6, 3287–3296. [Google Scholar] [CrossRef]
- Juvale, K.; Wiese, M. Design of inhibitors of BCRP/ABCG2. Future Med. Chem. 2015, 7, 1521–1527. [Google Scholar] [CrossRef]
- Brozik, A.; Hegedus, C.; Erdei, Z.; Hegedus, T.; Ozvegy-Laczka, C.; Szakacs, G.; Sarkadi, B. Tyrosine kinase inhibitors as modulators of ATP binding cassette multidrug transporters: Substrates, chemosensitizers or inducers of acquired multidrug resistance? Expert Opin. Drug Metab. Toxicol. 2011, 7, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Camidge, D.R.; Pao, W.; Sequist, L.V. Acquired resistance to TKIs in solid tumours: Learning from lung cancer. Nat. Rev. Clin. Oncol. 2014, 11, 473–481. [Google Scholar] [CrossRef]
- Du, W.; Huang, H.; Sorrelle, N.; Brekken, R.A. Sitravatinib potentiates immune checkpoint blockade in refractory cancer models. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Dolan, M.; Mastri, M.; Tracz, A.; Christensen, J.G.; Chatta, G.; Ebos, J.M.L. Enhanced efficacy of sitravatinib in metastatic models of antiangiogenic therapy resistance. PLoS ONE 2019, 14, e0220101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Fan, Y.F.; Cai, C.Y.; Wang, J.Q.; Teng, Q.X.; Lei, Z.N.; Zeng, L.; Gupta, P.; Chen, Z.S. Olmutinib (BI1482694/HM61713), a Novel Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor, Reverses ABCG2-Mediated Multidrug Resistance in Cancer Cells. Front. Pharmacol. 2018, 9, 1097. [Google Scholar] [CrossRef]
- Sodani, K.; Tiwari, A.K.; Singh, S.; Patel, A.; Xiao, Z.J.; Chen, J.J.; Sun, Y.L.; Talele, T.T.; Chen, Z.S. GW583340 and GW2974, human EGFR and HER-2 inhibitors, reverse ABCG2- and ABCB1-mediated drug resistance. Biochem. Pharm. 2012, 83, 1613–1622. [Google Scholar] [CrossRef]
- Tsuruo, T.; Iida, H.; Naganuma, K.; Tsukagoshi, S.; Sakurai, Y. Promotion by verapamil of vincristine responsiveness in tumor cell lines inherently resistant to the drug. Cancer Res. 1983, 43, 808–813. [Google Scholar]
- Tsuruo, T.; Iida, H.; Yamashiro, M.; Tsukagoshi, S.; Sakurai, Y. Enhancement of vincristine- and adriamycin-induced cytotoxicity by verapamil in P388 leukemia and its sublines resistant to vincristine and adriamycin. Biochem. Pharm. 1982, 31, 3138–3140. [Google Scholar] [CrossRef]
- Cai, C.Y.; Zhai, H.; Lei, Z.N.; Tan, C.P.; Chen, B.L.; Du, Z.Y.; Wang, J.Q.; Zhang, Y.K.; Wang, Y.J.; Gupta, P.; et al. Benzoyl indoles with metabolic stability as reversal agents for ABCG2-mediated multidrug resistance. Eur. J. Med. Chem. 2019, 179, 849–862. [Google Scholar] [CrossRef]
- Zhang, G.N.; Zhang, Y.K.; Wang, Y.J.; Gupta, P.; Ashby, C.R., Jr.; Alqahtani, S.; Deng, T.; Bates, S.E.; Kaddoumi, A.; Wurpel, J.N.D.; et al. Epidermal growth factor receptor (EGFR) inhibitor PD153035 reverses ABCG2-mediated multidrug resistance in non-small cell lung cancer: In vitro and in vivo. Cancer Lett. 2018, 424, 19–29. [Google Scholar] [CrossRef]
- Moore, M.J.; Goldstein, D.; Hamm, J.; Figer, A.; Hecht, J.R.; Gallinger, S.; Au, H.J.; Murawa, P.; Walde, D.; Wolff, R.A.; et al. Erlotinib plus gemcitabine compared with gemcitabine alone in patients with advanced pancreatic cancer: A phase III trial of the National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol. 2007, 25, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Yuan, J.Q.; Di, M.Y.; Zheng, D.Y.; Chen, J.Z.; Ding, H.; Wu, X.Y.; Huang, Y.F.; Mao, C.; Tang, J.L. Gemcitabine plus erlotinib for advanced pancreatic cancer: A systematic review with meta-analysis. PLoS ONE 2013, 8, e57528. [Google Scholar] [CrossRef] [PubMed]
- Geyer, C.E.; Forster, J.; Lindquist, D.; Chan, S.; Romieu, C.G.; Pienkowski, T.; Jagiello-Gruszfeld, A.; Crown, J.; Chan, A.; Kaufman, B.; et al. Lapatinib plus capecitabine for HER2-positive advanced breast cancer. N. Engl. J. Med. 2006, 355, 2733–2743. [Google Scholar] [CrossRef] [PubMed]
- Cetin, B.; Benekli, M.; Turker, I.; Koral, L.; Ulas, A.; Dane, F.; Oksuzoglu, B.; Kaplan, M.A.; Koca, D.; Boruban, C.; et al. Lapatinib plus capecitabine for HER2-positive advanced breast cancer: A multicentre study of Anatolian Society of Medical Oncology (ASMO). J. Chemother. 2014, 26, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Alemany, R.; Moura, D.S.; Redondo, A.; Martinez-Trufero, J.; Calabuig, S.; Saus, C.; Obrador-Hevia, A.; Ramos, R.F.; Villar, V.H.; Valverde, C.; et al. Nilotinib as co-adjuvant treatment with doxorubicin in patients with sarcomas: A phase I trial of the Spanish Group for Research on Sarcoma. Clin. Cancer Res. 2018, 24, 5239–5249. [Google Scholar] [CrossRef]
- Brendel, C.; Scharenberg, C.; Dohse, M.; Robey, R.W.; Bates, S.E.; Shukla, S.; Ambudkar, S.V.; Wang, Y.; Wennemuth, G.; Burchert, A.; et al. Imatinib mesylate and nilotinib (AMN107) exhibit high-affinity interaction with ABCG2 on primitive hematopoietic stem cells. Leukemia 2007, 21, 1267–1275. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Sodani, K.; Wang, S.R.; Kuang, Y.H.; Ashby, C.R., Jr.; Chen, X.; Chen, Z.S. Nilotinib (AMN107, Tasigna) reverses multidrug resistance by inhibiting the activity of the ABCB1/Pgp and ABCG2/BCRP/MXR transporters. Biochem. Pharm. 2009, 78, 153–161. [Google Scholar] [CrossRef]
- Dohse, M.; Scharenberg, C.; Shukla, S.; Robey, R.W.; Volkmann, T.; Deeken, J.F.; Brendel, C.; Ambudkar, S.V.; Neubauer, A.; Bates, S.E. Comparison of ATP-binding cassette transporter interactions with the tyrosine kinase inhibitors imatinib, nilotinib, and dasatinib. Drug Metab. Dispos. Biol. Fate Chem. 2010, 38, 1371–1380. [Google Scholar] [CrossRef]
- Shen, D.W.; Fojo, A.; Chin, J.E.; Roninson, I.B.; Richert, N.; Pastan, I.; Gottesman, M.M. Human multidrug-resistant cell lines: Increased MDR1 expression can precede gene amplification. Science 1986, 232, 643–645. [Google Scholar] [CrossRef]
- Honjo, Y.; Hrycyna, C.A.; Yan, Q.W.; Medina-Perez, W.Y.; Robey, R.W.; van de Laar, A.; Litman, T.; Dean, M.; Bates, S.E. Acquired mutations in the MXR/BCRP/ABCP gene alter substrate specificity in MXR/BCRP/ABCP-overexpressing cells. Cancer Res. 2001, 61, 6635–6639. [Google Scholar]
- Henrich, C.J.; Bokesch, H.R.; Dean, M.; Bates, S.E.; Robey, R.W.; Goncharova, E.I.; Wilson, J.A.; McMahon, J.B. A high-throughput cell-based assay for inhibitors of ABCG2 activity. J. Biomol. Screen. 2006, 11, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, M.; Tominaga, H.; Shiga, M.; Sasamoto, K.; Ohkura, Y.; Ueno, K. A combined assay of cell viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt, neutral red and crystal violet. Biol. Pharm. Bull. 1996, 19, 1518–1520. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.A.; Maylock, C.A.; Williams, J.A.; Paweletz, C.P.; Shu, H.; Shacter, E. Serum-derived protein S binds to phosphatidylserine and stimulates the phagocytosis of apoptotic cells. Nat. Immunol. 2003, 4, 87–91. [Google Scholar] [CrossRef]
- Wu, C.P.; Hsiao, S.H.; Sim, H.M.; Luo, S.Y.; Tuo, W.C.; Cheng, H.W.; Li, Y.Q.; Huang, Y.H.; Ambudkar, S.V. Human ABCB1 (P-glycoprotein) and ABCG2 mediate resistance to BI 2536, a potent and selective inhibitor of Polo-like kinase 1. Biochem. Pharm. 2013, 86, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Gribar, J.J.; Ramachandra, M.; Hrycyna, C.A.; Dey, S.; Ambudkar, S.V. Functional characterization of glycosylation-deficient human P-glycoprotein using a vaccinia virus expression system. J. Membr. Biol. 2000, 173, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Kowal, J.; Broude, E.; Roninson, I.; Locher, K.P. Structural insight into substrate and inhibitor discrimination by human P-glycoprotein. Science 2019, 363, 753–756. [Google Scholar] [CrossRef]
- Taylor, N.M.I.; Manolaridis, I.; Jackson, S.M.; Kowal, J.; Stahlberg, H.; Locher, K.P. Structure of the human multidrug transporter ABCG2. Nature 2017, 546, 504–509. [Google Scholar] [CrossRef]
- Wu, C.P.; Hsieh, Y.J.; Murakami, M.; Vahedi, S.; Hsiao, S.H.; Yeh, N.; Chou, A.W.; Li, Y.Q.; Wu, Y.S.; Yu, J.S.; et al. Human ATP-binding cassette transporters ABCB1 and ABCG2 confer resistance to histone deacetylase 6 inhibitor ricolinostat (ACY-1215) in cancer cell lines. Biochem. Pharm. 2018, 155, 316–325. [Google Scholar] [CrossRef]
- Stewart, C.F.; Leggas, M.; Schuetz, J.D.; Panetta, J.C.; Cheshire, P.J.; Peterson, J.; Daw, N.; Jenkins, J.J., 3rd; Gilbertson, R.; Germain, G.S.; et al. Gefitinib enhances the antitumor activity and oral bioavailability of irinotecan in mice. Cancer Res. 2004, 64, 7491–7499. [Google Scholar] [CrossRef]
- Leggas, M.; Panetta, J.C.; Zhuang, Y.; Schuetz, J.D.; Johnston, B.; Bai, F.; Sorrentino, B.; Zhou, S.; Houghton, P.J.; Stewart, C.F. Gefitinib modulates the function of multiple ATP-binding cassette transporters in vivo. Cancer Res. 2006, 66, 4802–4807. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Type | Transporter Expressed | IC50 (μM) 1 | R.F 2 |
|---|---|---|---|---|
| KB-3-1 | epidermal | - | 2.34 ± 0.70 | 1.0 |
| KB-V-1 | epidermal | ABCB1 | 2.29 ± 0.73 | 1.0 |
| OVCAR-8 | ovarian | - | 3.61 ± 1.64 | 1.0 |
| NCI-ADR-RES | ovarian | ABCB1 | 5.51 ± 1.26 | 1.5 |
| H460 | lung | - | 2.69 ± 1.07 | 1.0 |
| H460-MX20 | lung | ABCG2 | 2.24 ± 0.70 | 0.8 |
| S1 | colon | - | 1.69 ± 0.42 | 1.0 |
| S1-M1-80 | colon | ABCG2 | 2.13 ± 0.51 | 1.3 |
| pcDNA-HEK293 | - | - | 1.96 ± 0.50 | 1.0 |
| MDR19-HEK293 | - | ABCB1 | 2.72 ± 0.51 | 1.4 |
| R482-HEK293 | - | ABCG2 | 1.98 ± 0.45 | 1.0 |
| Treatment | Concentration (μM) | Mean IC50 † ± SD and (FR ‡) | |
|---|---|---|---|
| OVCAR-8 (Parental) [nM] | NCI-ADR-RES (Resistant) [μM] | ||
| Paclitaxel | - | 3.59 ± 0.61 (1.0) | 10.04 ± 1.89 (1.0) |
| +Sitravatinib | 0.1 | 3.00 ± 0.50 (1.2) | 6.84 ± 0.78 (1.5) |
| +Sitravatinib | 0.2 | 3.13 ± 0.65 (1.1) | 4.60 ± 0.47 ** (2.2) |
| +Sitravatinib | 0.3 | 2.73 ± 0.44 (1.3) | 1.87 ± 0.43 ** (5.4) |
| +Sitravatinib | 0.5 | 2.63 ± 0.39 (1.4) | 0.44 ± 0.10 *** (22.8) |
| +Verapamil | 5.0 | 2.67 ± 0.57 (1.3) | 0.34 ± 0.05 *** (29.5) |
| [nM] | [μM] | ||
| Colchicine | - | 30.91 ± 8.92 (1.0) | 3.32 ± 0.71 (1.0) |
| +Sitravatinib | 0.1 | 32.00 ± 9.09 (1.0) | 1.84 ± 0.51 * (1.8) |
| +Sitravatinib | 0.2 | 32.46 ± 9.51 (1.0) | 1.03 ± 0.33 ** (3.2) |
| +Sitravatinib | 0.3 | 32.66 ± 10.21 (1.0) | 0.69 ± 0.09 ** (4.8) |
| +Sitravatinib | 0.5 | 32.14 ± 10.01 (1.0) | 0.36 ± 0.07 ** (9.2) |
| +Verapamil | 5.0 | 22.86 ± 6.94 (1.4) | 0.75 ± 0.14 ** (4.4) |
| [nM] | [μM] | ||
| Vincristine | - | 7.68 ± 1.16 (1.0) | 4.29 ± 1.04 (1.0) |
| +Sitravatinib | 0.1 | 7.10 ± 1.06 (1.1) | 2.31 ± 0.42 * (1.9) |
| +Sitravatinib | 0.2 | 6.52 ± 0.93 (1.2) | 1.49 ± 0.19 * (2.9) |
| +Sitravatinib | 0.3 | 6.29 ± 0.95 (1.2) | 0.60 ± 0.09 ** (7.2) |
| +Sitravatinib | 0.5 | 6.44 ± 1.05 (1.2) | 0.15 ± 0.02 ** (28.6) |
| +Verapamil | 5.0 | 1.48 ± 0.22 *** (5.2) | 0.12 ± 0.02 ** (35.8) |
| Treatment | Concentration (μM) | KB-3-1 (Parental) [nM] | KB-V-1 (Resistant) [μM] |
| Paclitaxel | - | 2.37 ± 0.86 (1.0) | 2.76 ± 0.51 (1.0) |
| +Sitravatinib | 0.1 | 2.28 ± 0.77 (1.0) | 1.90 ± 0.26 (1.5) |
| +Sitravatinib | 0.2 | 2.01 ± 0.69 (1.2) | 1.85 ± 0.21 * (1.5) |
| +Sitravatinib | 0.3 | 2.20 ± 0.76 (1.1) | 0.88 ± 0.10 ** (3.1) |
| +Sitravatinib | 0.5 | 2.31 ± 0.80 (1.0) | 0.28 ± 0.05 ** (9.9) |
| +Verapamil | 5.0 | 1.85 ± 0.62 (1.3) | 66.96 ± 5.99 *** [nM] (41.2) |
| [nM] | [μM] | ||
| Colchicine | - | 12.50 ± 4.88 (1.0) | 1.66 ± 0.12 (1.0) |
| +Sitravatinib | 0.1 | 10.04 ± 3.41 (1.2) | 0.43 ± 0.05 *** (3.9) |
| +Sitravatinib | 0.2 | 11.85 ± 4.33 (1.1) | 0.80 ± 0.02 *** (2.1) |
| +Sitravatinib | 0.3 | 11.57 ± 4.38 (1.1) | 0.35 ± 0.05 *** (4.7) |
| +Sitravatinib | 0.5 | 11.68 ± 4.27 (1.1) | 0.37 ± 0.04 *** (4.5) |
| +Verapamil | 5.0 | 7.29 ± 2.19 (1.7) | 0.24 ± 0.03 *** (6.9) |
| [nM] | [nM] | ||
| Vincristine | - | 1.23 ± 0.46 (1.0) | 1479.50 ± 275.52 (1.0) |
| +Sitravatinib | 0.1 | 1.19 ± 0.40 (1.0) | 1207.91 ± 183.53 (1.2) |
| +Sitravatinib | 0.2 | 1.31 ± 0.45 (0.9) | 916.44 ± 164.61 * (1.6) |
| +Sitravatinib | 0.3 | 1.38 ± 0.50 (0.9) | 555.14 ± 99.66 ** (2.7) |
| +Sitravatinib | 0.5 | 1.38 ± 0.45 (0.9) | 118.37 ± 44.58 ** (12.5) |
| +Verapamil | 5.0 | 0.24 ± 0.09 * (5.1) | 25.49 ± 4.05 *** (58.0) |
| Treatment | Concentration (μM) | pcDNA-HEK293 (Parental) [nM] | MDR19-HEK293 (Resistant) [nM] |
| Paclitaxel | - | 1.45 ± 0.21 (1.0) | 817.35 ± 93.18 (1.0) |
| +Sitravatinib | 0.1 | 1.44 ± 0.25 (1.0) | 126.61 ± 18.40 *** (6.5) |
| +Sitravatinib | 0.2 | 1.33 ± 0.24 (1.1) | 30.55 ± 4.40 *** (26.8) |
| +Sitravatinib | 0.3 | 1.44 ± 0.27 (1.0) | 13.94 ± 1.62 *** (58.6) |
| +Sitravatinib | 0.5 | 1.41 ± 0.27 (1.0) | 7.81 ± 0.97 *** (104.65) |
| +Verapamil | 5.0 | 1.02 ± 0.18 (1.4) | 9.60 ± 1.52 *** (85.1) |
| [nM] | [nM] | ||
| Colchicine | - | 13.26 ± 3.73 (1.0) | 169.27 ± 34.71 (1.0) |
| +Sitravatinib | 0.1 | 11.67 ± 3.05 (1.1) | 74.28 ± 10.60 * (2.3) |
| +Sitravatinib | 0.2 | 11.70 ± 3.06 (1.1) | 45.81 ± 7.81 ** (3.7) |
| +Sitravatinib | 0.3 | 11.64 ± 3.25 (1.1) | 40.74 ± 9.43 ** (4.2) |
| +Sitravatinib | 0.5 | 9.38 ± 2.11 (1.4) | 25.18 ± 5.72 ** (6.7) |
| +Verapamil | 5.0 | 11.80 ± 3.05 (1.1) | 57.09 ± 9.89 ** (3.0) |
| [nM] | [nM] | ||
| Vincristine | - | 1.32 ± 0.23 (1.0) | 193.67 ± 31.50 (1.0) |
| +Sitravatinib | 0.1 | 1.31 ± 0.30 (1.0) | 54.89 ± 6.11 ** (3.5) |
| +Sitravatinib | 0.2 | 1.15 ± 0.29 (1.1) | 8.81 ± 1.52 *** (22.0) |
| +Sitravatinib | 0.3 | 1.15 ± 0.27 (1.1) | 9.53 ± 1.20 *** (20.3) |
| +Sitravatinib | 0.5 | 1.04 ± 0.24 (1.3) | 2.40 ± 0.43 *** (80.7) |
| +Verapamil | 5.0 | 0.54 ± 0.14 ** (2.4) | 1.79 ± 0.32 *** (108.2) |
| Treatment | Concentration (μM) | Mean IC50 † ± SD and (FR ‡) | |
|---|---|---|---|
| H460 (Parental) [nM] | H460-MX20 (Resistant) [nM] | ||
| Mitoxantrone | - | 57.25 ± 9.90 (1.0) | 820.80 ± 118.43 (1.0) |
| +Sitravatinib | 0.1 | 39.65 ± 6.73 (1.4) | 592.76 ± 102.60 (1.4) |
| +Sitravatinib | 0.2 | 48.17 ± 9.95 (1.2) | 447.77 ± 89.18 * (1.8) |
| +Sitravatinib | 0.3 | 38.98 ± 6.05 (1.5) | 309.55 ± 70.08 ** (2.7) |
| +Sitravatinib | 0.5 | 50.58 ± 9.62 (1.1) | 238.35 ± 64.00 ** (3.4) |
| +Ko143 | 1.0 | 50.83 ± 10.60 (1.1) | 120.15 ± 29.28 *** (6.8) |
| [nM] | [nM] | ||
| Topotecan | - | 67.27 ± 9.29 (1.0) | 2005.50 ± 588.05 (1.0) |
| +Sitravatinib | 0.1 | 53.29 ± 7.46 (1.3) | 993.81 ± 312.10 (2.0) |
| +Sitravatinib | 0.2 | 45.94 ± 7.52 * (1.5) | 590.88 ± 206.01 * (3.4) |
| +Sitravatinib | 0.3 | 49.11 ± 9.28 (1.4) | 316.30 ± 114.34 ** (6.3) |
| +Sitravatinib | 0.5 | 37.07 ± 6.41 ** (1.8) | 333.40 ± 131.66 ** (6.0) |
| +Ko143 | 1.0 | 35.89 ± 6.98 ** (1.9) | 75.50 ± 25.41 ** (26.6) |
| [nM] | [nM] | ||
| SN-38 | - | 11.29 ± 1.01 (1.0) | 246.22 ± 38.74 (1.0) |
| +Sitravatinib | 0.1 | 9.67 ± 1.59 (1.2) | 123.55 ± 24.08 ** (2.0) |
| +Sitravatinib | 0.2 | 8.32 ± 1.32 * (1.4) | 77.41 ± 18.10 ** (3.2) |
| +Sitravatinib | 0.3 | 6.35 ± 1.04 ** (1.8) | 62.12 ± 16.50 ** (4.0) |
| +Sitravatinib | 0.5 | 5.43 ± 0.78 ** (2.1) | 45.26 ± 14.37 ** (5.4) |
| +Ko143 | 1.0 | 3.27 ± 0.73 *** (3.5) | 9.86 ± 3.51 *** (25.0) |
| Treatment | Concentration (μM) | S1 (Parental) [nM] | S1-M1-80 (Resistant) [μM] |
| Mitoxantrone | - | 6.77 ± 1.18 (1.0) | 55.46 ± 5.66 (1.0) |
| +Sitravatinib | 0.1 | 7.28 ± 1.53 (0.9) | 14.25 ± 1.99 *** (3.9) |
| +Sitravatinib | 0.2 | 6.87 ± 1.43 (1.0) | 7.13 ± 0.87 *** (7.8) |
| +Sitravatinib | 0.3 | 6.01 ± 1.29 (1.1) | 4.46 ± 0.53 *** (12.4) |
| +Sitravatinib | 0.5 | 5.59 ± 1.18 (1.2) | 1.74 ± 0.23 *** (31.9) |
| +Ko143 | 1.0 | 7.10 ± 1.55 (1.0) | 0.53 ± 0.07 *** (104.6) |
| [nM] | [nM] | ||
| Topotecan | - | 35.38 ± 5.38 (1.0) | 2995.95 ± 504.50 (1.0) |
| +Sitravatinib | 0.1 | 34.75 ± 4.48 (1.0) | 1250.80 ± 253.93 ** (2.4) |
| +Sitravatinib | 0.2 | 35.25 ± 5.06 (1.0) | 609.29 ± 127.64 ** (4.9) |
| +Sitravatinib | 0.3 | 35.50 ± 4.72 (1.0) | 422.24 ± 91.32 *** (7.1) |
| +Sitravatinib | 0.5 | 40.24 ± 6.04 (0.9) | 279.53 ± 63.52 *** (11.2) |
| +Ko143 | 1.0 | 36.39 ± 5.57 (1.0) | 91.76 ± 19.15 *** (32.6) |
| [nM] | [nM] | ||
| SN-38 | - | 4.07 ± 0.43 (1.0) | 1642.50 ± 236.20 (1.0) |
| +Sitravatinib | 0.1 | 4.28 ± 0.50 (1.0) | 597.00 ± 128.03 ** (2.8) |
| +Sitravatinib | 0.2 | 4.57 ± 0.55 (0.9) | 316.89 ± 74.75 *** (5.2) |
| +Sitravatinib | 0.3 | 4.57 ± 0.57 (0.9) | 205.26 ± 45.33 *** (8.0) |
| +Sitravatinib | 0.5 | 5.09 ± 0.68 (0.9) | 121.59 ± 26.89 *** (13.5) |
| +Ko143 | 1.0 | 4.11 ± 0.49 (1.0) | 12.39 ± 2.47 *** (132.6) |
| Treatment | Concentration (μM) | pcDNA-HEK293 (Parental) [nM] | R482-HEK293 (Resistant) [nM] |
| Mitoxantrone | - | 3.86 ± 0.47 (1.0) | 116.36 ± 10.39 (1.0) |
| +Sitravatinib | 0.1 | 2.60 ± 0.46 * (1.5) | 41.18 ± 4.02 *** (2.8) |
| +Sitravatinib | 0.2 | 2.56 ± 0.44 * (1.5) | 27.06 ± 3.07 *** (4.3) |
| +Sitravatinib | 0.3 | 3.16 ± 0.66 (1.2) | 11.44 ± 1.58 *** (10.2) |
| +Sitravatinib | 0.5 | 2.66 ± 0.42 * (1.5) | 12.54 ± 1.53 *** (9.3) |
| +Ko143 | 1.0 | 3.90 ± 0.56 (1.0) | 9.71 ± 1.55 *** (12.0) |
| [nM] | [nM] | ||
| Topotecan | - | 26.09 ± 6.11 (1.0) | 169.46 ± 22.27 (1.0) |
| +Sitravatinib | 0.1 | 29.26 ± 8.03 (1.0) | 71.52 ± 10.98 (1.0) |
| +Sitravatinib | 0.2 | 33.39 ± 9.19 (1.0) | 59.49 ± 10.19 (1.0) |
| +Sitravatinib | 0.3 | 27.42 ± 7.35 (1.0) | 51.96 ± 9.75 (1.0) |
| +Sitravatinib | 0.5 | 29.80 ± 7.99 (1.0) | 39.22 ± 8.51 (1.0) |
| +Ko143 | 1.0 | 28.67 ± 7.40 (1.0) | 30.83 ± 8.26 (1.0) |
| [nM] | [nM] | ||
| SN-38 | - | 4.05 ± 0.75 (1.0) | 155.78 ± 13.95 (1.0) |
| +Sitravatinib | 0.1 | 3.92 ± 0.83 (1.0) | 23.74 ± 2.99 *** (6.6) |
| +Sitravatinib | 0.2 | 4.06 ± 1.01 (1.0) | 21.58 ± 2.64 *** (7.2) |
| +Sitravatinib | 0.3 | 3.65 ± 0.94 (1.1) | 16.17 ± 2.57 *** (9.6) |
| +Sitravatinib | 0.5 | 3.39 ± 1.01 (1.2) | 11.29 ± 1.79 *** (13.8) |
| +Ko143 | 1.0 | 4.27 ± 0.84 (0.9) | 2.88 ± 0.81 *** (54.1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-P.; Hsiao, S.-H.; Huang, Y.-H.; Hung, L.-C.; Yu, Y.-J.; Chang, Y.-T.; Hung, T.-H.; Wu, Y.-S. Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. Cancers 2020, 12, 195. https://doi.org/10.3390/cancers12010195
Wu C-P, Hsiao S-H, Huang Y-H, Hung L-C, Yu Y-J, Chang Y-T, Hung T-H, Wu Y-S. Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. Cancers. 2020; 12(1):195. https://doi.org/10.3390/cancers12010195
Chicago/Turabian StyleWu, Chung-Pu, Sung-Han Hsiao, Yang-Hui Huang, Lang-Cheng Hung, Yi-Jou Yu, Yu-Tzu Chang, Tai-Ho Hung, and Yu-Shan Wu. 2020. "Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs" Cancers 12, no. 1: 195. https://doi.org/10.3390/cancers12010195
APA StyleWu, C.-P., Hsiao, S.-H., Huang, Y.-H., Hung, L.-C., Yu, Y.-J., Chang, Y.-T., Hung, T.-H., & Wu, Y.-S. (2020). Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. Cancers, 12(1), 195. https://doi.org/10.3390/cancers12010195