Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis


 ,
, 
Abstract
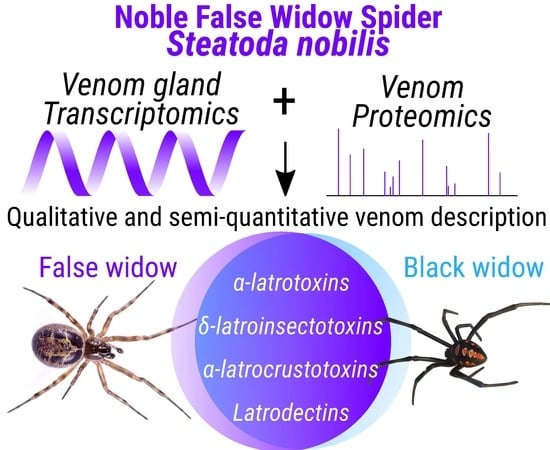
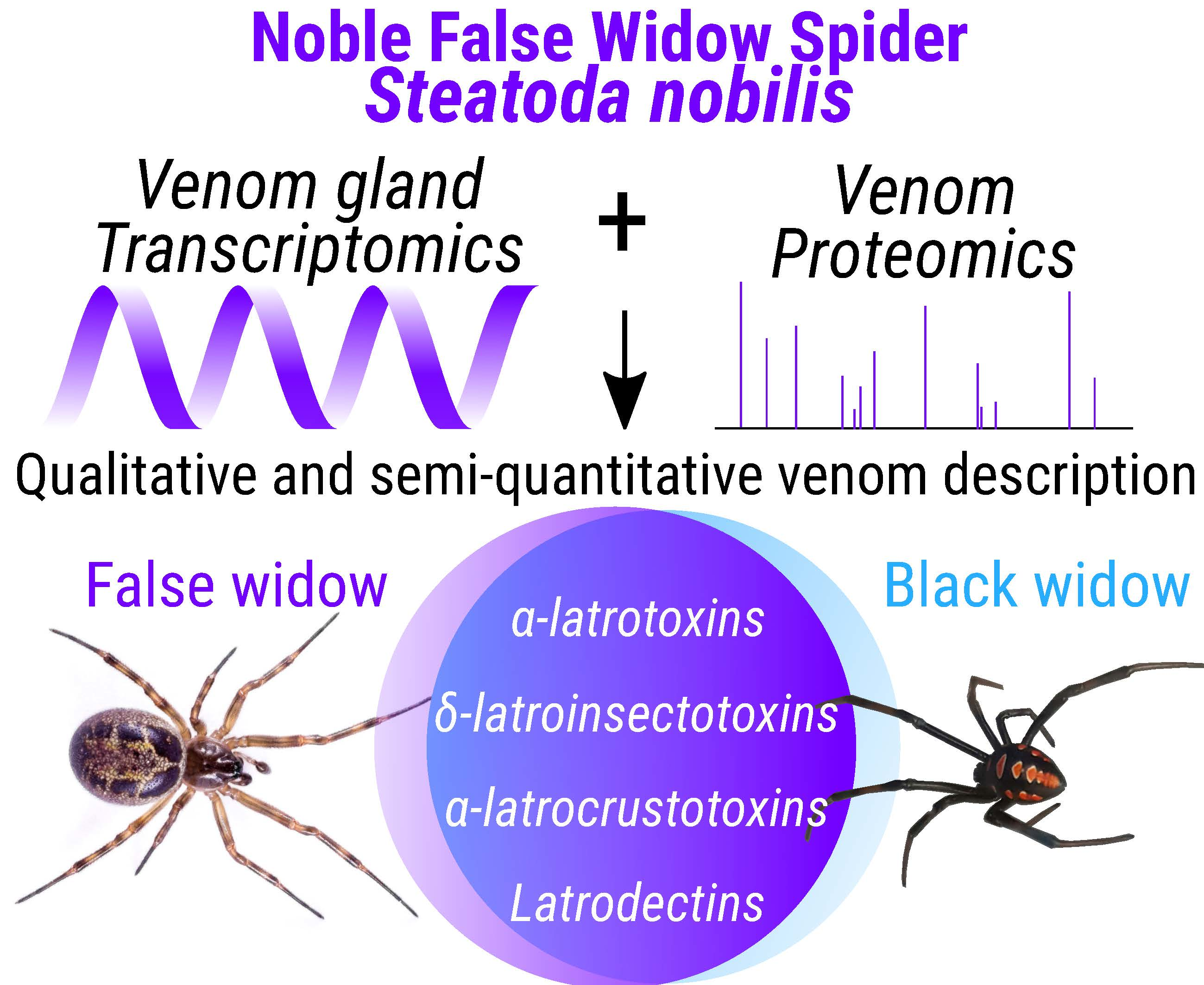
1. Introduction
2. Results
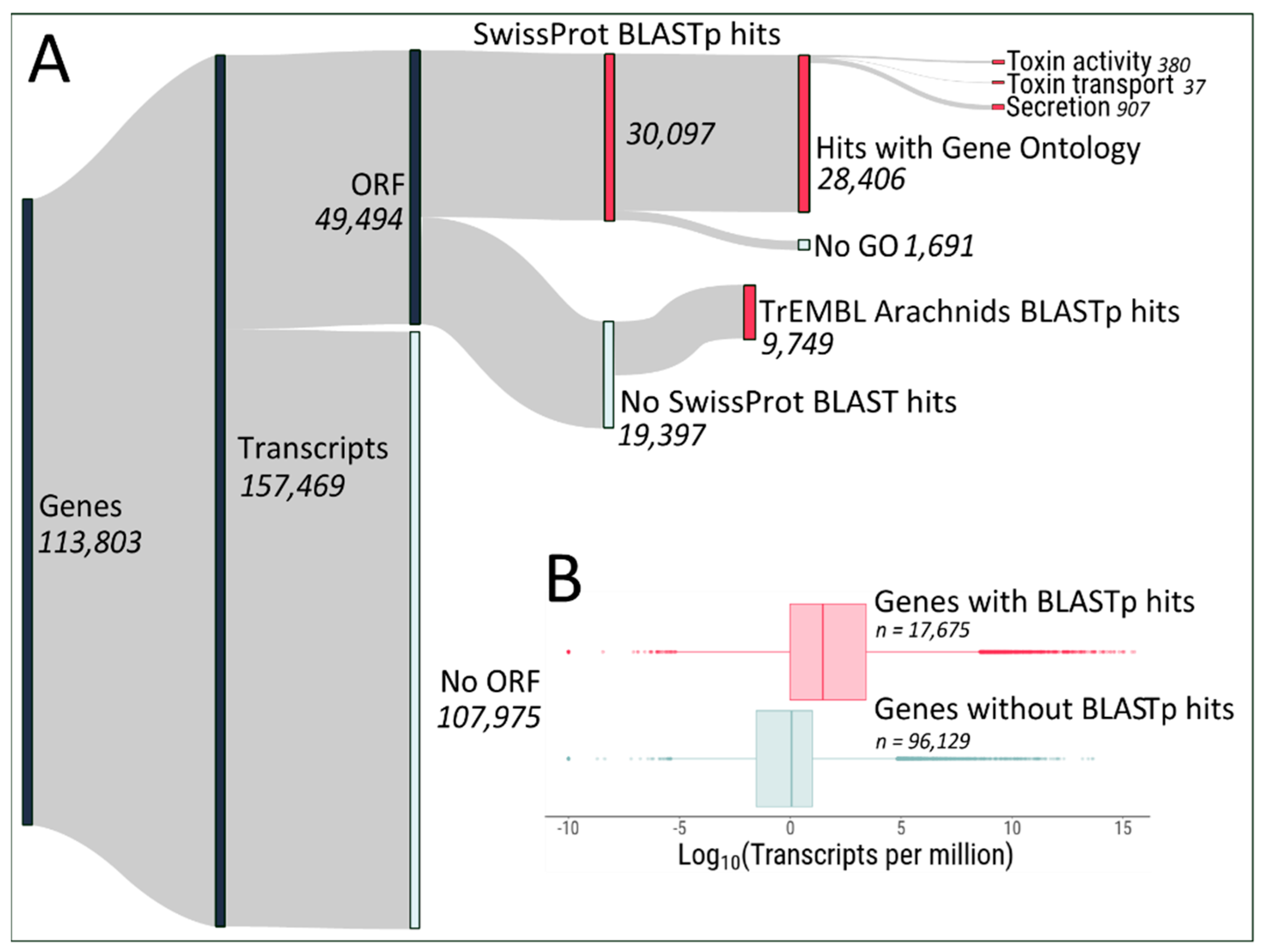
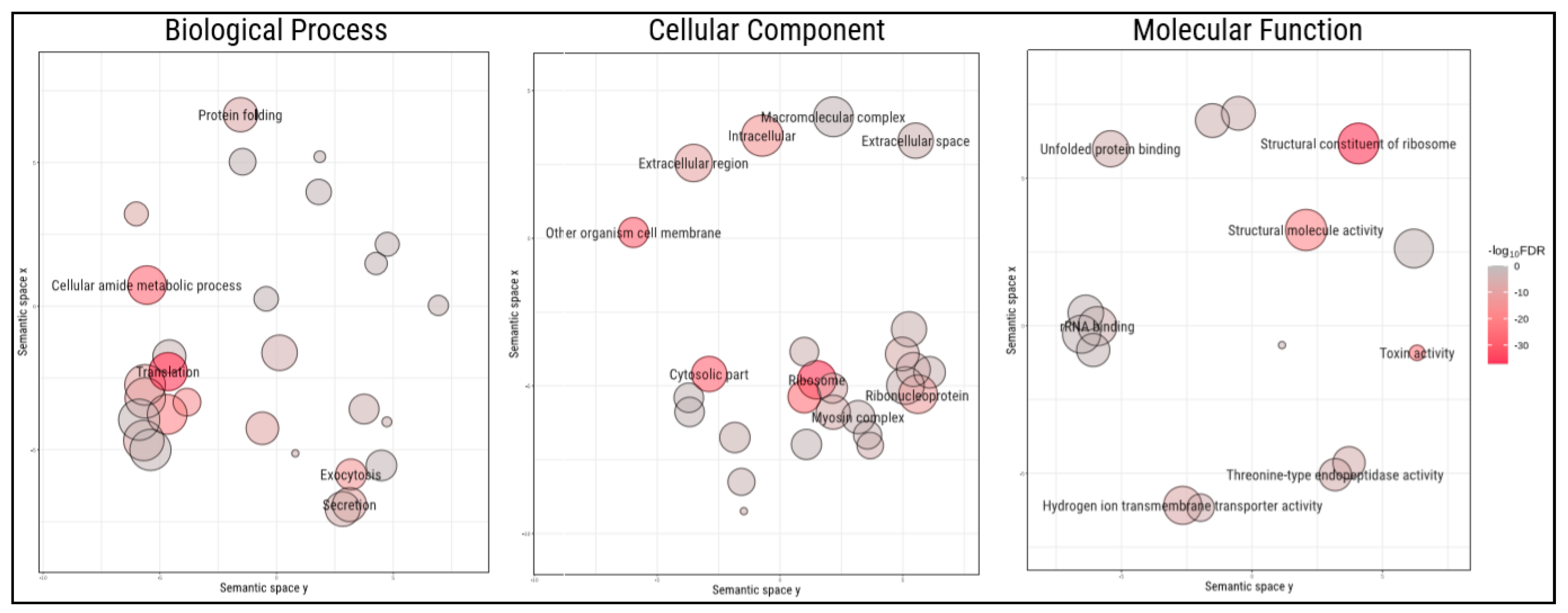
2.1. Protein Identification from Transcripts
2.2. Shotgun Proteomics of Steatoda nobilis
Qualitative Data Analysis
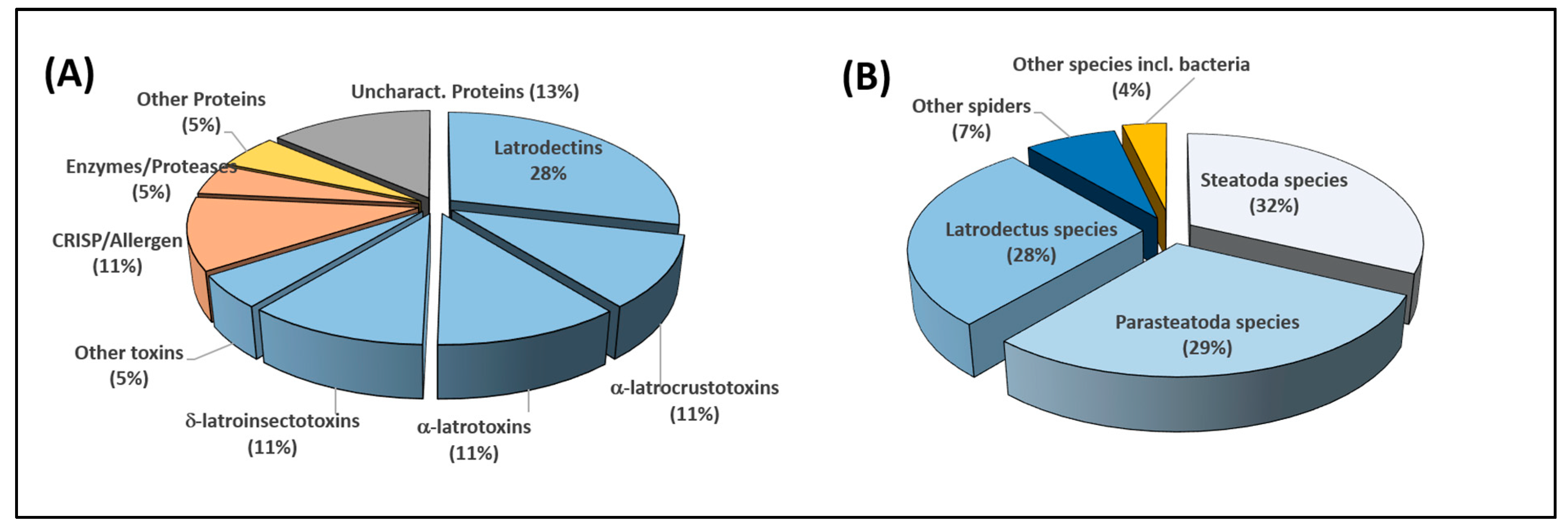
2.3. Integration of Transcriptomics and Proteomics Data
2.3.1. Qualitative Data Analysis
2.3.2. Relative Quantitative Analysis
3. Discussion
3.1. Toxins
3.2. Enzymes
4. Conclusions
5. Materials and Methods
5.1. Spider Collection and Venom Extraction
5.2. Venom Gland Removal and RNA Extraction
5.3. Transcriptomics Assembly and Analysis
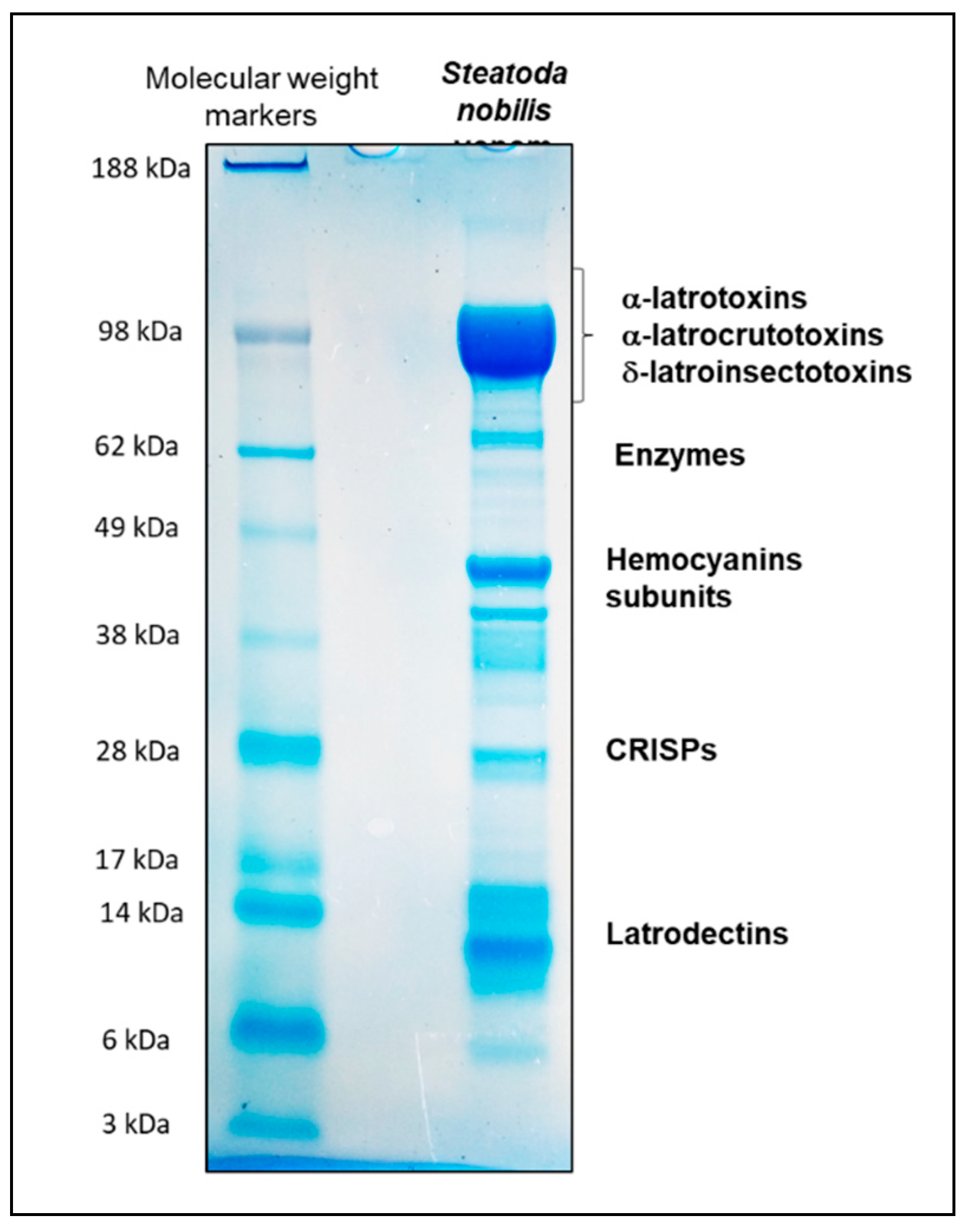
5.4. SDS-PAGE of Steatoda nobilis Female Venom
5.5. Shotgun Proteomics of Steatoda nobilis Female Venom
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lyons, K.; Dugon, M.M.; Healy, K. Diet Breadth Mediates the Prey Specificity of Venom Potency in Snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, G.; Logan, R.A.; Leung, K.Y.; Newberry, F.J.; Rowley, P.D.; Dunbar, J.P.; Wagstaff, S.C.; Casewell, N.R.; Harrison, R.A. Stabilising the Integrity of Snake Venom mRNA Stored under Tropical Field Conditions Expands Research Horizons. PLoS Negl. Trop. Dis. 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Bottrall, J.L.; Madaras, F.; Biven, C.D.; Venning, M.G.; Mirtschin, P.J. Proteolytic Activity of Elapid and Viperid Snake Venoms and Its Implication to Digestion. J. Venom Res. 2010, 1, 18. [Google Scholar] [PubMed]
- Zobel-Thropp, P.A.; Kerins, A.E.; Binford, G.J. Sphingomyelinase D in Sicariid Spider Venom is a Potent Insecticidal Toxin. Toxicon 2012, 60, 265–271. [Google Scholar] [CrossRef]
- Vassilevski, A.; Kozlov, S.; Grishin, E. Molecular Diversity of Spider Venom. Biochemistry (Moscow) 2009, 74, 1505–1534. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L.; Langenegger, N.; Heller, M.; Koua, D.; Nentwig, W. The Dual Prey-Inactivation Strategy of Spiders—In-Depth Venomic Analysis of Cupiennius salei. Toxins 2019, 11, 167. [Google Scholar] [CrossRef]
- Garb, J.E.; Hayashi, C.Y. Molecular Evolution of α-Latrotoxin, the Exceptionally Potent Vertebrate Neurotoxin in Black Widow Spider Venom. Molecular Biol. Evol. 2013, 30, 999–1014. [Google Scholar] [CrossRef]
- Bauer, T.; Feldmeier, S.; Krehenwinkel, H.; Wieczorrek, C.; Reiser, N.; Breitling, R. Steatoda nobilis, a False Widow on the Rise: A Synthesis of Past and Current Distribution Trends. NeoBiota 2019, 42, 19. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Afoullouss, S.; Sulpice, R.; Dugon, M.M. Envenomation by the Noble False Widow Spider Steatoda nobilis (Thorell, 1875)–Five New Cases of Steatodism from Ireland and Great Britain. Clin. Toxicol. 2018, 56, 433–435. [Google Scholar] [CrossRef]
- Faúndez, E.I.; Téllez, F. Primer Registro de Una Mordedura De Steatoda nobilis (Thorell, 1875) (Arachnida: Araneae: Theridiidae) en Chile. Arquivos Entomolóxicos 2016, 15, 237–240. [Google Scholar]
- Warrell, D.A.; Shaheen, J.; Hillyard, P.D.; Jones, D. Neurotoxic Envenoming by an Immigrant Spider (Steatoda nobilis) in Southern England. Toxicon 1991, 29, 1263–1265. [Google Scholar] [CrossRef]
- Dugon, M.M.; Dunbar, J.P.; Afoullouss, S.; Schulte, J.; McEvoy, A.; English, M.J.; Hogan, R.; Ennis, C.; Sulpice, R. Occurrence, reproductive rate and identification of the non-native noble false widow spider Steatoda nobilis (Thorell, 1875) in Ireland. Biol. Environ. 2017, 117, 77–89. [Google Scholar]
- Dunbar, J.P.; Ennis, C.; Gandola, R.; Dugon, M.M. Biting off more than one can chew: First record of the non-native noble false widow spider Steatoda nobilis (Thorell, 1875) feeding on the native viviparous lizard Zootoca vivipara. Biol. Environ. 2018, 118B, 45–48. [Google Scholar]
- Dunbar, J.P.; Schulte, J.; Lyons, K.; Fort, A.; Dugon, M.M. New Irish Record for Steatoda triangulosa (Walckenaer, 1802), and New County Records for Steatoda nobilis (Thorell, 1875), Steatoda bipunctata (Linnaeus, 1758) and Steatoda grossa (CL Koch, 1838). Ir. Nat. J. 2018, 36, 39–43. [Google Scholar]
- Nolan, M. Three Spiders (Araneae) New to Ireland: Bolyphantes alticeps, Oonops domesticus and Steatoda nobilis. Ir. Nat. J. 1999, 26, 200–202. [Google Scholar]
- Türkeş, T.; Mergen, O. The Comb-Footed Spider Fauna of the Central Anatolia Region and New Records for the Turkish Fauna (Araneae: Theridiidae). Serket 2007, 10, 112–119. [Google Scholar]
- Zamani, A.; Mirshamsi, O.; Jannesar, B.; Marusik, Y.M.; Esyunin, S.L. New Data on Spider Fauna of Iran (Arachnida: Araneae), Part II. Zool. Ecol. 2015, 25, 339–346. [Google Scholar] [CrossRef]
- Faúndez, E.I.; Carvajal, M.A.; Darquea-Schettini, D.; González-Cano, E. Nuevos Registros de Steatoda nobilis (Thorell, 1875)(Araneae: Theridiidae) de Sudamérica. Revista Ibérica de Aracnología 2018, 33, 52–54. [Google Scholar]
- Taucare-Ríos, A.; Mardones, D.; Zúñiga-Reinoso, Á. Steatoda nobilis (Araneae: Theridiidae) in South America: A New Alien Species for Chile. Can. Entomol. 2016, 148, 479–481. [Google Scholar] [CrossRef][Green Version]
- Vetter, R.S.; Adams, R.J.; Berrian, J.E.; Vincent, L.S. The European Spider Steatoda nobilis (Thorell, 1875)(Araneae: Theridiidae) becoming Widespread in California. Pan-Pac. Entomol. 2015, 91, 98–101. [Google Scholar] [CrossRef]
- Snazell, R.; Jones, D. The Theridiid Spider Steatoda nobilis (Thorell, 1875) in Britain. Bull. Br. Arachnol. Soc. 1993, 9, 164–167. [Google Scholar]
- Kulczycki, A.; Simeon, E.; Legittimo, C.M.; Di Pompeo, P. New Records of Steatoda nobilis (Thorell, 1875)(Araneae, Theridiidae), an Introduced Species on the Italian Mainland and in Sardinia. Arachnology 2012, 15, 269–272. [Google Scholar] [CrossRef]
- Gendreau, K.L.; Haney, R.A.; Schwager, E.E.; Wierschin, T.; Stanke, M.; Richards, S.; Garb, J.E. House Spider Genome uncovers Evolutionary Shifts in the Diversity and Expression of Black Widow Venom Proteins Associated with Extreme Toxicity. BMC Genom. 2017, 18, 178. [Google Scholar] [CrossRef]
- Haney, R.A.; Ayoub, N.A.; Clarke, T.H.; Hayashi, C.Y.; Garb, J.E. Dramatic Expansion of the Black Widow Toxin Arsenal uncovered by Multi-Tissue Transcriptomics and Venom Proteomics. BMC Genom. 2014, 15, 366. [Google Scholar] [CrossRef]
- Orlova, E.V.; Rahman, M.A.; Gowen, B.; Volynski, K.E.; Ashton, A.C.; Manser, C.; van Heel, M.; Ushkaryov, Y.A. Structure of α-Latrotoxin Oligomers Reveals that Divalent Cation-Dependent Tetramers form Membrane Pores. Nat. Struct. Mol. Biol. 2000, 7, 48. [Google Scholar]
- Ushkaryov, Y.A.; Rohou, A.; Sugita, S. receptors. In Pharmacology of Neurotransmitter Release; Springer: Berlin/Heidelberg, Germany, 2008; pp. 171–206. [Google Scholar]
- O’Shea, M.; Kelly, K. Predation on a Weasel Skink (Saproscincus mustelinus)(Squamata: Scincidae: Lygosominae) by a Redback Spider (Latrodectus hasselti)(Araneae: Araneomorpha: Theridiidae), with a Review of other Latrodectus Predation Events Involving Squamates. Herpetofauna 2017, 44, 49–55. [Google Scholar]
- Degueldre, M.; Verdenaud, M.; Legarda, G.; Minambres, R.; Zuniga, S.; Leblanc, M.; Gilles, N.; Ducancel, F.; De Pauw, E.; Quinton, L. Diversity in Sequences, Post-Translational Modifications and Expected Pharmacological Activities of Toxins from Four Conus Species Revealed by the Combination of Cutting-Edge Proteomics, Transcriptomics and Bioinformatics. Toxicon 2017, 130, 116–125. [Google Scholar] [CrossRef]
- Giribaldi, J.; Kazandjian, T.; Amorim, F.G.; Whiteley, G.; Wagstaff, S.C.; Cazals, G.; Enjalbal, C.; Quinton, L.; Casewell, N.R.; Dutertre, S. Venomics of the asp viper Vipera aspis aspis from France. J. Proteom. 2020, 218, 103707. [Google Scholar] [CrossRef]
- Silva, J.C.; Gorenstein, M.V.; Li, G.Z.; Vissers, J.P.; Geromanos, S.J. Absolute Quantification of Proteins by LCMSE: A Virtue of Parallel MS Acquisition. Mol. Cell. Proteom. 2006, 5, 144–156. [Google Scholar] [CrossRef]
- Pineda, S.S.; Chaumeil, P.A.; Kunert, A.; Kaas, Q.; Thang, M.W.; Le, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B.; et al. ArachnoServer 3.0: An Online Resource for Automated Discovery, Analysis and Annotation of Spider Toxins. Bioinformatics 2018, 34, 1074–1076. [Google Scholar] [CrossRef]
- Casas, J. Spider Physiology and Behaviour: Behaviour; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- McMahon, H.T.; Rosenthal, L.; Meldolesi, J.; Nicholls, D.G. α-Latrotoxin Releases both Vesicular and Cytoplasmic Glutamate from Isolated Nerve Terminals. J. Neurochem. 1990, 55, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Mackessy, S.P. Handbook of Venoms and Toxins of Reptiles; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Langenegger, N.; Koua, D.; Schürch, S.; Heller, M.; Nentwig, W.; Kuhn-Nentwig, L. Identification of a Precursor Processing Protease from the Spider Cupiennius salei Essential for Venom Neurotoxin Maturation. J. Biol. Chem. 2018, 293, 2079–2090. [Google Scholar] [CrossRef] [PubMed]
- Forster, L. The behavioural ecology of Latrodectus hasselti (Thorell), the Australian Redback Spider (Araneae: Theridiidae): A Review. Rec. West. Aust. Mus. 1995, 52, 13–24. [Google Scholar]
- Vetter, R.; Swanson, D.; Weinstein, S.; White, J. Do Spiders Vector Bacteria during Bites? The Evidence Indicates Otherwise. Toxicon 2015, 93, 171–174. [Google Scholar] [CrossRef]
- Südhof, T.C. α-Latrotoxin and Its Receptors: Neurexins and CIRL/Latrophilins. Annu. Rev. Neurosci. 2001, 24, 933–962. [Google Scholar] [CrossRef]
- Morgon, A.M.; Belisario-Ferrari, M.R.; Trevisan-Silva, D.; Meissner, G.O.; Vuitika, L.; Marin, B.; Tashima, A.K.; Gremski, L.H.; Gremski, W.; Senff-Ribeiro, A.; et al. Expression and immunological cross-reactivity of LALP3, a novel astacin-like metalloprotease from brown spider (Loxosceles intermedia) venom. Biochimie 2016, 128, 8–19. [Google Scholar] [CrossRef]
- Song, L.; Florea, L. Rcorrector: Efficient and Accurate Error Correction for Illumina RNA-seq Reads. GigaScience 2015, 4, s13742-015-0089-y. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a Full-Length Transcriptome without a Genome from RNA-Seq Data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494. [Google Scholar] [CrossRef]
- Haas, B.; Papanicolaou, A. TransDecoder (Find Coding Regions within Transcripts). Github, nd. Available online: https://github.com/TransDecoder/TransDecoder (accessed on 17 May 2018).
- Rokyta, D.R.; Ward, M.J. Venom-Gland Transcriptomics and Venom Proteomics of the Black-Back Scorpion (Hadrurus spadix) Reveal Detectability Challenges and an Unexplored Realm of Animal Toxin Diversity. Toxicon 2017, 128, 23–37. [Google Scholar] [CrossRef]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl de Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.U. UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam Protein Families Database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Goseq: Gene Ontology Testing for RNA-seq Datasets. R Bioconduct. 2012, 8, 1–25. [Google Scholar]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Development Core Team, R. R: A Language and Environment for Statistical Computing; R foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Bederson, B.B.; Shneiderman, B.; Wattenberg, M. Ordered and Quantum Treemaps: Making Effective Use of 2D Space to Display Hierarchies. AcM Trans. Graph. (TOG) 2002, 21, 833–854. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latrodectus Toxins | Abbreviation | Function/Activity |
|---|---|---|
| α-latrotoxin | α-LTX | Toxic to vertebrates, forms calcium channels on pre-synaptic neurons, triggers neurotransmitter release. |
| α- latrocrustotoxin | α-LCT | Toxic to crustaceans, forms calcium channels on pre-synaptic neurons, triggers neurotransmitter release. |
| α,d-latroinsectotoxin | α-LIT / d-LIT | Toxic to insects, forms calcium channels on pre-synaptic neurons, triggers neurotransmitter release. |
| Latrodectin | α-LTX LMWPs | Enhances potency of latrotoxins |
| Cystein Rich Secretory Protein | CRISPs | Block Calcium channels |
| Metalloprotease | MPs | Tissue lysis, facilitates spread of latrotoxins |
| Serine protease | SPs | Tissue lysis, facilitates spread of latrotoxins |
| Hyaluronidase | -- | Tissue lysis, facilitates spread of latrotoxins |
| Chitinase | -- | Degrades arthropod exoskeletons |
| Inhibitor cystine knot | ICK | Alters ion channel function |
| Steatoda nobilis Proteins | Best Match (BLAST) | Accession Number | Species | Quantification in % | Sequence Coverage % | −10lgP |
|---|---|---|---|---|---|---|
| Latrodectin-Sno1a | Latrodectin; Alpha-latrotoxin associated LMWP-2 | V9QFH8 | Steatoda grossa | 23.5 | 63 | 210.63 |
| Alpha-latrocrustotoxin-Sno1a | Alpha-latrocrustotoxin-Lt1a | Q9XZC0 | Latrodectus tredecimguttatus | 10.6 | 70 | 447.19 |
| CRISP-Sno1a | CRISP/Allergen/PR-1-like | XP_015912134 | Parasteatoda tepidariorum | 9.4 | 81 | 274.42 |
| Uncharacterized Protein-Sno1a | Uncharacterized protein LOC107442339 | XP_015911366 | Parasteatoda tepidariorum | 6.1 | 66 | 242.45 |
| Alpha-latrotoxin-Sno1a | Alpha-latrotoxin-Lhe1a | P0DJE3 | Latrodectus hesperus | 4.7 | 60 | 295.77 |
| Putative neurotoxin-Sno1a | Putative neurotoxin LTDF 06-01 | A0A0K1D8C3 | Dolomedes fimbriatus | 3.7 | 48 | 167.43 |
| Delta-latroinsectotoxin-Sno1a | Delta-latroinsectotoxin-Lt1a | Q25338 | Latrodectus tredecimguttatus | 2.8 | 55 | 369.06 |
| Latrodectin-Sno1b | Latrodectin; Alpha-latrotoxin associated LMWP-2 | V9QFH8 | Steatoda grossa | 2.5 | 60 | 222.56 |
| Uncharacterized Protein-Sno1b | Uncharacterized protein LOC107437515 | XP_015905073 | Parasteatoda tepidariorum | 2.4 | 56 | 175.17 |
| Latrodectin-Sno1c | Latrodectin; Alpha-latrotoxin-associated LMWP | AHC13266.1 | Steatoda grossa | 2.1 | 63 | 178.46 |
| Delta-latroinsectotoxin-Sno1b | Delta-latroinsectotoxin-Lt1a | Q25338 | Latrodectus tredecimguttatus | 1.2 | 47 | 352.89 |
| CRISP-Sno1b | CRISP/Allergen/PR-1-like | XP_015912134 | Parasteatoda tepidariorum | 1.1 | 69 | 209.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunbar, J.P.; Fort, A.; Redureau, D.; Sulpice, R.; Dugon, M.M.; Quinton, L. Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis. Toxins 2020, 12, 402. https://doi.org/10.3390/toxins12060402
Dunbar JP, Fort A, Redureau D, Sulpice R, Dugon MM, Quinton L. Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis. Toxins. 2020; 12(6):402. https://doi.org/10.3390/toxins12060402
Chicago/Turabian StyleDunbar, John P., Antoine Fort, Damien Redureau, Ronan Sulpice, Michel M. Dugon, and Loïc Quinton. 2020. "Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis" Toxins 12, no. 6: 402. https://doi.org/10.3390/toxins12060402
APA StyleDunbar, J. P., Fort, A., Redureau, D., Sulpice, R., Dugon, M. M., & Quinton, L. (2020). Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis. Toxins, 12(6), 402. https://doi.org/10.3390/toxins12060402