
The Influence of Vegan, Vegetarian, and Omnivorous Diets on Protein Metabolism: A Role for the Gut–Muscle Axis?
Abstract
1. Introduction
2. Effects of Vegan, Vegetarian, and Omnivorous Diets on Gut Microbiota Diversity, Composition, and Associated Bacteria-Derived Metabolites
2.1. Modulation of the Microbiome with Diet
2.2. Cross-Sectional Studies Comparing Vegan and Omnivorous Diets
2.3. Intervention Studies Comparing Gut Microbiome Responses to Vegan and Omnivorous Diets
2.4. Limitations of Current Evidence Comparing Gut Microbiome Responses Between Vegan and Omnivorous Diets
3. Effects of Vegan, Vegetarian, and Omnivorous Diets on Muscle Protein Synthesis Responses
3.1. Effects of Supplement Vegan and Omnivorous Protein Sources on Muscle Protein Synthesis Responses
3.2. Effects of Whole-Food Protein Sources on Muscle Protein Synthesis Responses Between Vegan and Omnivorous Diets
3.3. Limitations of Current Evidence Comparing Muscle Protein Synthesis Responses Between Vegan and Omnivorous Diets
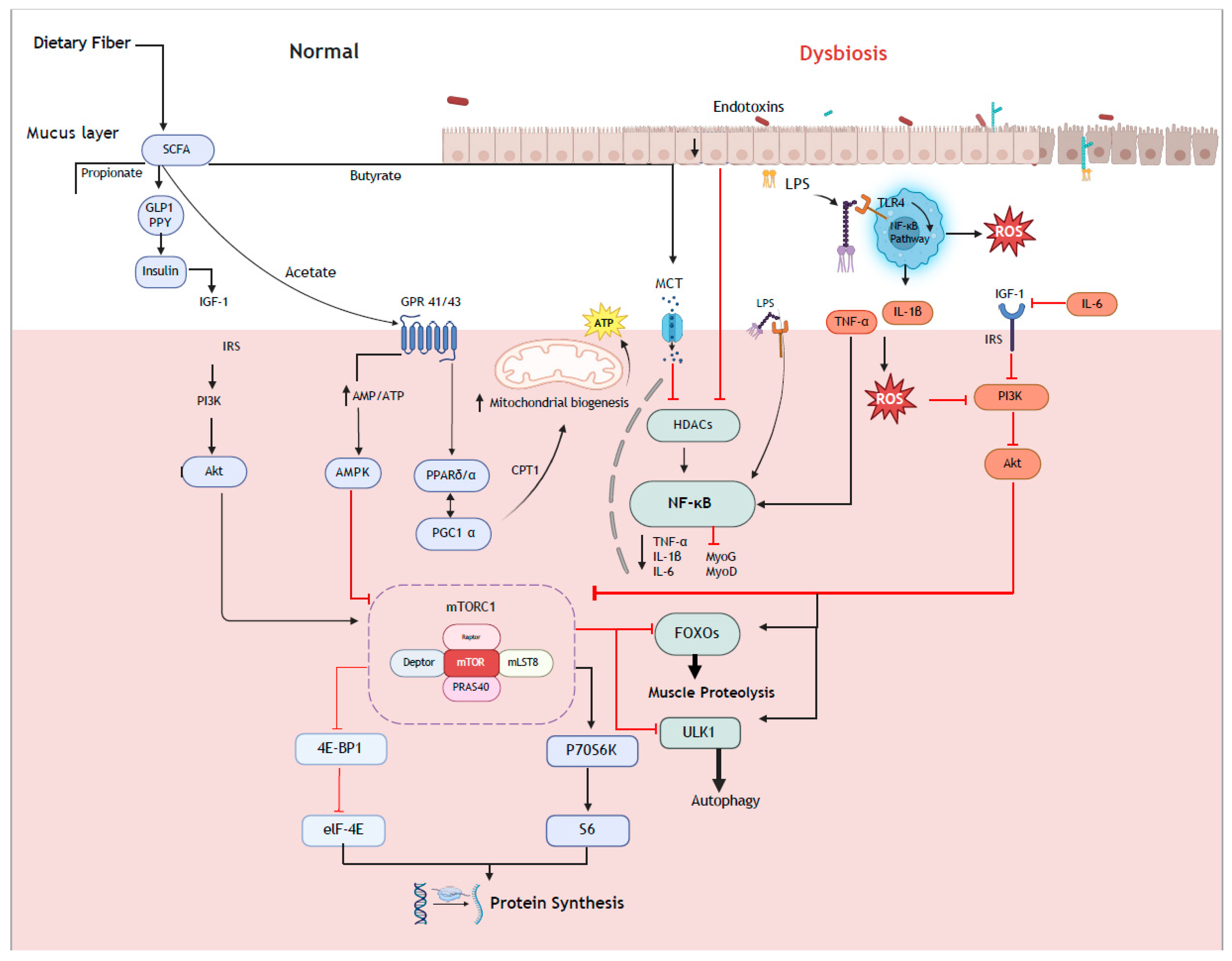
4. The Gut–Muscle Axis: A Mechanism for Altering Muscle Anabolism Between Vegan and Omnivorous Diets?
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Marrone, G.; Guerriero, C.; Palazzetti, D.; Lido, P.; Marolla, A.; Di Daniele, F.; Noce, A. Vegan diet health benefits in metabolic syndrome. Nutrients 2021, 13, 817. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the anthropocene: The EAT-Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Aschemann-Witzel, J.; Gantriis, R.F.; Fraga, P.; Perez-Cueto, F.J.A. Plant-based food and protein trend from a business perspective: Markets, consumers, and the challenges and opportunities in the future. Crit. Rev. Food Sci. Nutr. 2021, 61, 3119–3128. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, S.; Burd, N.A.; van Loon, L.J. The skeletal muscle anabolic response to plant- versus animal-based protein consumption. J. Nutr. 2015, 145, 1981–1991. [Google Scholar] [CrossRef]
- Losno, E.A.; Sieferle, K.; Perez-Cueto, F.J.A.; Ritz, C. Vegan diet and the gut microbiota composition in healthy adults. Nutrients 2021, 13, 2402. [Google Scholar] [CrossRef]
- Dinu, M.; Abbate, R.; Gensini, G.F.; Casini, A.; Sofi, F. Vegetarian, vegan diets and multiple health outcomes: A systematic review with meta-analysis of observational studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3640–3649. [Google Scholar] [CrossRef]
- Satija, A.; Hu, F.B. Plant-based diets and cardiovascular health. Trends Cardiovasc. Med. 2018, 28, 437–441. [Google Scholar] [CrossRef]
- Wilson, P.B. Nutrition behaviors, perceptions, and beliefs of recent marathon finishers. Phys. Sportsmed. 2016, 44, 242–251. [Google Scholar] [CrossRef]
- Turner-McGrievy, G.M.; Moore, W.J.; Barr-Anderson, D. The interconnectedness of diet choice and distance running: Results of the research understanding the nutrition of endurance runners (RUNNER) study. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 205–211. [Google Scholar] [CrossRef]
- Pohl, A.; Schünemann, F.; Bersiner, K.; Gehlert, S. The impact of vegan and vegetarian diets on physical performance and molecular signaling in skeletal muscle. Nutrients 2021, 13, 3884. [Google Scholar] [CrossRef]
- Elorinne, A.L.; Alfthan, G.; Erlund, I.; Kivimäki, H.; Paju, A.; Salminen, I.; Turpeinen, U.; Voutilainen, S.; Laakso, J. Food and nutrient intake and nutritional status of finnish vegans and non-vegetarians. PLoS ONE 2016, 11, e0148235. [Google Scholar] [CrossRef] [PubMed]
- Rogerson, D. Vegan diets: Practical advice for athletes and exercisers. J. Int. Soc. Sports Nutr. 2017, 14, 36. [Google Scholar] [CrossRef]
- Pinckaers, P.J.M.; Kouw, I.W.K.; Hendriks, F.K.; van Kranenburg, J.M.X.; de Groot, L.; Verdijk, L.B.; Snijders, T.; van Loon, L.J.C. No differences in muscle protein synthesis rates following ingestion of wheat protein, milk protein, and their protein blend in healthy, young males. Br. J. Nutr. 2021, 126, 1832–1842. [Google Scholar] [CrossRef] [PubMed]
- Pinckaers, P.J.M.; Smeets, J.S.J.; Kouw, I.W.K.; Goessens, J.P.B.; Gijsen, A.P.B.; de Groot, L.; Verdijk, L.B.; van Loon, L.J.C.; Snijders, T. Post-prandial muscle protein synthesis rates following the ingestion of pea-derived protein do not differ from ingesting an equivalent amount of milk-derived protein in healthy, young males. Eur. J. Nutr. 2024, 63, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009, 107, 987–992. [Google Scholar] [CrossRef]
- Siddharth, J.; Chakrabarti, A.; Pannérec, A.; Karaz, S.; Morin-Rivron, D.; Masoodi, M.; Feige, J.N.; Parkinson, S.J. Aging and sarcopenia associate with specific interactions between gut microbes, serum biomarkers and host physiology in rats. Aging 2017, 9, 1698–1720. [Google Scholar] [CrossRef]
- Zhang, T.; Cheng, J.K.; Hu, Y.M. Gut microbiota as a promising therapeutic target for age-related sarcopenia. Ageing Res. Rev. 2022, 81, 101739. [Google Scholar] [CrossRef]
- Przewłócka, K.; Folwarski, M.; Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Kaczor, J.J. Gut-muscle axis exists and may affect skeletal muscle adaptation to training. Nutrients 2020, 12, 1451. [Google Scholar] [CrossRef]
- Banaszak, M.; Górna, I.; Przysławski, J. Non-pharmacological treatments for insulin resistance: Effective intervention of plant-based diets-a critical review. Nutrients 2022, 14, 1400. [Google Scholar] [CrossRef]
- Craig, W.J.; Mangels, A.R.; Fresán, U.; Marsh, K.; Miles, F.L.; Saunders, A.V.; Haddad, E.H.; Heskey, C.E.; Johnston, P.; Larson-Meyer, E.; et al. The safe and effective use of plant-based diets with guidelines for health professionals. Nutrients 2021, 13, 4144. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of diet on the gut microbiota: Rethinking intervention duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed]
- Bolte, L.A.; Lee, K.A.; Björk, J.R.; Leeming, E.R.; Campmans-Kuijpers, M.J.E.; de Haan, J.J.; Vila, A.V.; Maltez-Thomas, A.; Segata, N.; Board, R.; et al. Association of a mediterranean diet with outcomes for patients treated with immune checkpoint blockade for advanced melanoma. JAMA Oncol. 2023, 9, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Weisburger, J.H.; Wynder, E.L. Effects of high risk and low risk diets for colon carcinogenesis on fecal microflora and steroids in man. J. Nutr. 1975, 105, 878–884. [Google Scholar] [CrossRef]
- Zimmer, J.; Lange, B.; Frick, J.S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A vegan or vegetarian diet substantially alters the human colonic faecal microbiota. Eur. J. Clin. Nutr. 2012, 66, 53–60. [Google Scholar] [CrossRef]
- Kabeerdoss, J.; Devi, R.S.; Mary, R.R.; Ramakrishna, B.S. Faecal microbiota composition in vegetarians: Comparison with omnivores in a cohort of young women in southern India. Br. J. Nutr. 2012, 108, 953–957. [Google Scholar] [CrossRef]
- Matijašić, B.B.; Obermajer, T.; Lipoglavšek, L.; Grabnar, I.; Avguštin, G.; Rogelj, I. Association of dietary type with fecal microbiota in vegetarians and omnivores in Slovenia. Eur. J. Nutr. 2014, 53, 1051–1064. [Google Scholar] [CrossRef]
- Ferrocino, I.; Di Cagno, R.; De Angelis, M.; Turroni, S.; Vannini, L.; Bancalari, E.; Rantsiou, K.; Cardinali, G.; Neviani, E.; Cocolin, L. Fecal microbiota in healthy subjects following omnivore, vegetarian and vegan diets: Culturable populations and rRNA DGGE profiling. PLoS ONE 2015, 10, e0128669. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Laghi, L.; Gobbetti, M.; Ercolini, D. Unusual sub-genus associations of faecal Prevotella and Bacteroides with specific dietary patterns. Microbiome 2016, 4, 57. [Google Scholar] [CrossRef]
- Wu, G.D.; Compher, C.; Chen, E.Z.; Smith, S.A.; Shah, R.D.; Bittinger, K.; Chehoud, C.; Albenberg, L.G.; Nessel, L.; Gilroy, E.; et al. Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production. Gut 2016, 65, 63–72. [Google Scholar] [CrossRef]
- Losasso, C.; Eckert, E.M.; Mastrorilli, E.; Villiger, J.; Mancin, M.; Patuzzi, I.; Di Cesare, A.; Cibin, V.; Barrucci, F.; Pernthaler, J.; et al. Assessing the influence of vegan, vegetarian and omnivore oriented westernized dietary styles on human gut microbiota: A cross sectional study. Front. Microbiol. 2018, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Trefflich, I.; Dietrich, S.; Braune, A.; Abraham, K.; Weikert, C. Short- and branched-chain fatty acids as fecal markers for microbiota activity in vegans and omnivores. Nutrients 2021, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, M.; Budinska, E.; Kuzma, M.; Pelantova, H.; Hradecky, J.; Heczkova, M.; Daskova, N.; Bratova, M.; Modos, I.; Videnska, P.; et al. Vegan diet is associated with favorable effects on the metabolic performance of intestinal microbiota: A cross-sectional multi-omics study. Front. Nutr. 2021, 8, 783302. [Google Scholar] [CrossRef] [PubMed]
- Stege, P.B.; Hordijk, J.; Shetty, S.A.; Visser, M.; Viveen, M.C.; Rogers, M.R.C.; Gijsbers, E.; Dierikx, C.M.; van der Plaats, R.Q.J.; van Duijkeren, E.; et al. Impact of long-term dietary habits on the human gut resistome in the Dutch population. Sci. Rep. 2022, 12, 1892. [Google Scholar] [CrossRef] [PubMed]
- Tarallo, S.; Ferrero, G.; De Filippis, F.; Francavilla, A.; Pasolli, E.; Panero, V.; Cordero, F.; Segata, N.; Grioni, S.; Pensa, R.G.; et al. Stool microRNA profiles reflect different dietary and gut microbiome patterns in healthy individuals. Gut 2022, 71, 1302–1314. [Google Scholar] [CrossRef]
- Seel, W.; Reiners, S.; Kipp, K.; Simon, M.C.; Dawczynski, C. Role of dietary fiber and energy intake on gut microbiome in vegans, vegetarians, and flexitarians in comparison to omnivores-insights from the nutritional evaluation (NuEva) study. Nutrients 2023, 15, 1914. [Google Scholar] [CrossRef]
- Fackelmann, G.; Manghi, P.; Carlino, N.; Heidrich, V.; Piccinno, G.; Ricci, L.; Piperni, E.; Arrè, A.; Bakker, E.; Creedon, A.C.; et al. Gut microbiome signatures of vegan, vegetarian and omnivore diets and associated health outcomes across 21,561 individuals. Nat. Microbiol. 2025, 10, 41–52. [Google Scholar] [CrossRef]
- van Faassen, A.; Bol, J.; van Dokkum, W.; Pikaar, N.A.; Ockhuizen, T.; Hermus, R.J. Bile acids, neutral steroids, and bacteria in feces as affected by a mixed, a lacto-ovovegetarian, and a vegan diet. Am. J. Clin. Nutr. 1987, 46, 962–967. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Zhang, C.; Björkman, A.; Cai, K.; Liu, G.; Wang, C.; Li, Y.; Xia, H.; Sun, L.; Kristiansen, K.; Wang, J.; et al. Impact of a 3-months vegetarian diet on the gut microbiota and immune repertoire. Front. Immunol. 2018, 9, 908. [Google Scholar] [CrossRef]
- Kohnert, E.; Kreutz, C.; Binder, N.; Hannibal, L.; Gorkiewicz, G.; Müller, A.; Storz, M.A.; Huber, R.; Lederer, A.K. Changes in gut microbiota after a four-week intervention with vegan vs. meat-rich diets in healthy participants: A randomized controlled trial. Microorganisms 2021, 9, 727. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.Q.; Lin, X.; Shen, H.; Liu, H.M.; Qiu, X.; Li, B.Y.; Shen, W.D.; Ge, C.L.; Lv, F.Y.; Shen, J.; et al. Human gut microbiome impacts skeletal muscle mass via gut microbial synthesis of the short-chain fatty acid butyrate among healthy menopausal women. J. Cachexia Sarcopenia Muscle 2021, 12, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Data, O.W.I. Per-Capita-Milk-Consumption. Available online: https://ourworldindata.org/grapher/per-capita-milk-consumption (accessed on 17 July 2023).
- Chen, P.Y.; Li, S.; Koh, Y.C.; Wu, J.C.; Yang, M.J.; Ho, C.T.; Pan, M.H. Oolong tea extract and citrus peel polymethoxyflavones reduce transformation of l-carnitine to trimethylamine-n-oxide and decrease vascular inflammation in l-carnitine feeding mice. J. Agric. Food Chem. 2019, 67, 7869–7879. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Bäckhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Toribio-Mateas, M.A.; Bester, A.; Klimenko, N. Impact of plant-based meat alternatives on the gut microbiota of consumers: A real-world study. Foods 2021, 10, 2040. [Google Scholar] [CrossRef]
- Slimani, N.; Fahey, M.; Welch, A.A.; Wirfält, E.; Stripp, C.; Bergström, E.; Linseisen, J.; Schulze, M.B.; Bamia, C.; Chloptsios, Y.; et al. Diversity of dietary patterns observed in the European Prospective Investigation into Cancer and Nutrition (EPIC) project. Public Health Nutr. 2002, 5, 1311–1328. [Google Scholar] [CrossRef]
- Daniel, C.R.; Cross, A.J.; Koebnick, C.; Sinha, R. Trends in meat consumption in the USA. Public Health Nutr. 2011, 14, 575–583. [Google Scholar] [CrossRef]
- Krebs-Smith, S.M.; Guenther, P.M.; Subar, A.F.; Kirkpatrick, S.I.; Dodd, K.W. Americans do not meet federal dietary recommendations. J. Nutr. 2010, 140, 1832–1838. [Google Scholar] [CrossRef]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the gut microbiome using 16S or shotgun metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Quinn, R.A.; Debelius, J.; Xu, Z.Z.; Morton, J.; Garg, N.; Jansson, J.K.; Dorrestein, P.C.; Knight, R. Microbiome-wide association studies link dynamic microbial consortia to disease. Nature 2016, 535, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.B.; Tarnopolsky, M.A.; Macdonald, M.J.; Macdonald, J.R.; Armstrong, D.; Phillips, S.M. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am. J. Clin. Nutr. 2007, 85, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Churchward-Venne, T.A.; Burd, N.A.; Breen, L.; Tarnopolsky, M.A.; Phillips, S.M. Myofibrillar protein synthesis following ingestion of soy protein isolate at rest and after resistance exercise in elderly men. Nutr. Metab. 2012, 9, 57. [Google Scholar] [CrossRef]
- Gorissen, S.H.; Horstman, A.M.; Franssen, R.; Crombag, J.J.; Langer, H.; Bierau, J.; Respondek, F.; van Loon, L.J. Ingestion of wheat protein increases in vivo muscle protein synthesis rates in healthy older men in a randomized trial. J. Nutr. 2016, 146, 1651–1659. [Google Scholar] [CrossRef]
- Oikawa, S.Y.; Bahniwal, R.; Holloway, T.M.; Lim, C.; McLeod, J.C.; McGlory, C.; Phillips, S.M. Potato protein isolate stimulates muscle protein synthesis at rest and with resistance exercise in young women. Nutrients 2020, 12, 1235. [Google Scholar] [CrossRef]
- Pinckaers, P.J.M.; Hendriks, F.K.; Hermans, W.J.H.; Goessens, J.P.B.; Senden, J.M.; JMX, V.A.N.K.; Wodzig, W.; Snijders, T.; LJC, V.A.N.L. Potato protein ingestion increases muscle protein synthesis rates at rest and during recovery from exercise in humans. Med. Sci. Sports Exerc. 2022, 54, 1572–1581. [Google Scholar] [CrossRef]
- Kouw, I.W.; Pinckaers, P.J.; Le Bourgot, C.; van Kranenburg, J.M.; Zorenc, A.H.; de Groot, L.C.; van Loon, L.J. Ingestion of an ample amount of meat substitute based on a lysine-enriched, plant-based protein blend stimulates postprandial muscle protein synthesis to a similar extent as an isonitrogenous amount of chicken in healthy, young men. Br. J. Nutr. 2022, 128, 1955–1965. [Google Scholar] [CrossRef]
- Davies, R.W.; Kozior, M.; Lynch, A.E.; Bass, J.J.; Atherton, P.J.; Smith, K.; Jakeman, P.M. The effect of fava bean (Vicia faba L.) protein ingestion on myofibrillar protein synthesis at rest and after resistance exercise in healthy, young men and women: A randomised control trial. Nutrients 2022, 14, 3688. [Google Scholar] [CrossRef]
- Pinckaers, P.J.M.; Kouw, I.W.K.; Gorissen, S.H.M.; Houben, L.H.P.; Senden, J.M.; Wodzig, W.; de Groot, L.; Verdijk, L.B.; Snijders, T.; van Loon, L.J.C. The muscle protein synthetic response to the ingestion of a plant-derived protein blend does not differ from an equivalent amount of milk protein in healthy young males. J. Nutr. 2023, 152, 2734–2743. [Google Scholar] [CrossRef]
- Pinckaers, P.J.; Domić, J.; Petrick, H.L.; Holwerda, A.M.; Trommelen, J.; Hendriks, F.K.; Houben, L.H.; Goessens, J.P.; van Kranenburg, J.M.; Senden, J.M.; et al. Higher muscle protein synthesis rates following ingestion of an omnivorous meal compared with an isocaloric and isonitrogenous vegan meal in healthy, older adults. J. Nutr. 2024, 154, 2120–2132. [Google Scholar] [CrossRef]
- Pinckaers, P.J.M.; Weijzen, M.E.G.; Houben, L.H.P.; Zorenc, A.H.; Kouw, I.W.K.; de Groot, L.; Verdijk, L.B.; Snijders, T.; van Loon, L.J.C. The muscle protein synthetic response following corn protein ingestion does not differ from milk protein in healthy, young adults. Amino Acids 2024, 56, 8. [Google Scholar] [CrossRef] [PubMed]
- Monteyne, A.J.; Coelho, M.O.C.; Porter, C.; Abdelrahman, D.R.; Jameson, T.S.O.; Jackman, S.R.; Blackwell, J.R.; Finnigan, T.J.A.; Stephens, F.B.; Dirks, M.L.; et al. Mycoprotein ingestion stimulates protein synthesis rates to a greater extent than milk protein in rested and exercised skeletal muscle of healthy young men: A randomized controlled trial. Am. J. Clin. Nutr. 2020, 112, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Monteyne, A.J.; Coelho, M.O.; Porter, C.; Abdelrahman, D.R.; Jameson, T.S.; Finnigan, T.J.; Wall, B.T. Branched-chain amino acid fortification does not restore muscle protein synthesis rates following ingestion of lower-compared with higher-dose mycoprotein. J. Nutr. 2020, 150, 2931–2941. [Google Scholar] [CrossRef] [PubMed]
- Monteyne, A.J.; Dunlop, M.V.; Machin, D.J.; Coelho, M.O.C.; Pavis, G.F.; Porter, C.; Murton, A.J.; Abdelrahman, D.R.; Dirks, M.L.; Stephens, F.B.; et al. A mycoprotein-based high-protein vegan diet supports equivalent daily myofibrillar protein synthesis rates compared with an isonitrogenous omnivorous diet in older adults: A randomised controlled trial. Br. J. Nutr. 2021, 126, 674–684. [Google Scholar] [CrossRef]
- Monteyne, A.J.; Coelho, M.O.C.; Murton, A.J.; Abdelrahman, D.R.; Blackwell, J.R.; Koscien, C.P.; Knapp, K.M.; Fulford, J.; Finnigan, T.J.A.; Dirks, M.L.; et al. Vegan and omnivorous high protein diets support comparable daily myofibrillar protein synthesis rates and skeletal muscle hypertrophy in young adults. J. Nutr. 2023, 153, 1680–1695. [Google Scholar] [CrossRef]
- Churchward-Venne, T.A.; Pinckaers, P.J.M.; Smeets, J.S.J.; Peeters, W.M.; Zorenc, A.H.; Schierbeek, H.; Rollo, I.; Verdijk, L.B.; van Loon, L.J.C. Myofibrillar and mitochondrial protein synthesis rates do not differ in young men following the ingestion of carbohydrate with whey, soy, or leucine-enriched soy protein after concurrent resistance- and endurance-type exercise. J. Nutr. 2019, 149, 210–220. [Google Scholar] [CrossRef]
- West, S.; Monteyne, A.J.; Whelehan, G.; van der Heijden, I.; Abdelrahman, D.R.; Murton, A.J.; Finnigan, T.J.A.; Stephens, F.B.; Wall, B.T. Ingestion of mycoprotein, pea protein, and their blend support comparable postexercise myofibrillar protein synthesis rates in resistance-trained individuals. Am. J. Physiol. Endocrinol. Metab. 2023, 325, E267–E279. [Google Scholar] [CrossRef]
- West, S.; Monteyne, A.J.; Whelehan, G.; Abdelrahman, D.R.; Murton, A.J.; Finnigan, T.J.A.; Blackwell, J.R.; Stephens, F.B.; Wall, B.T. Mycoprotein ingestion within or without its wholefood matrix results in equivalent stimulation of myofibrillar protein synthesis rates in resting and exercised muscle of young men. Br. J. Nutr. 2023, 130, 20–32. [Google Scholar] [CrossRef]
- van der Heijden, I.; West, S.; Monteyne, A.J.; Finnigan, T.J.; Abdelrahman, D.R.; Murton, A.J.; Wall, B.T. Algae ingestion increases resting and exercised myofibrillar protein synthesis rates to a similar extent as mycoprotein in young adults. J. Nutr. 2023, 153, 3406–3417. [Google Scholar] [CrossRef]
- Lim, C.; Janssen, T.A.; Currier, B.S.; Paramanantharajah, N.; McKendry, J.; Abou Sawan, S.; Phillips, S.M. Muscle protein synthesis in response to plant-based protein isolates with and without added leucine versus whey protein in young men and women. Curr. Dev. Nutr. 2024, 8, 103769. [Google Scholar] [CrossRef]
- van der Heijden, I.; Monteyne, A.J.; West, S.; Morton, J.P.; Langan-Evans, C.; Hearris, M.A.; Wall, B.T. Plant protein blend ingestion stimulates post-exercise myofibrillar protein synthesis rates equivalently to whey in resistance-trained adults. Med. Sci. Sports Exerc. 2024, 56, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Domić, J.; Pinckaers, P.J.; Grootswagers, P.; Siebelink, E.; Gerdessen, J.C.; van Loon, L.J.; de Groot, L.C. A well-balanced vegan diet does not compromise daily mixed muscle protein synthesis rates when compared with an omnivorous diet in active older adults: A randomized controlled cross-over trial. J. Nutr. 2024. [Google Scholar] [CrossRef]
- Wall, B.T.; Gorissen, S.H.; Pennings, B.; Koopman, R.; Groen, B.B.; Verdijk, L.B.; van Loon, L.J. Aging is accompanied by a blunted muscle protein synthetic response to protein ingestion. PLoS ONE 2015, 10, e0140903. [Google Scholar] [CrossRef]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005, 19, 422–424. [Google Scholar] [CrossRef]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. Aging is associated with diminished accretion of muscle proteins after the ingestion of a small bolus of essential amino acids. Am. J. Clin. Nutr. 2005, 82, 1065–1073. [Google Scholar] [CrossRef]
- Shad, B.J.; Thompson, J.L.; Breen, L. Does the muscle protein synthetic response to exercise and amino acid-based nutrition diminish with advancing age? A systematic review. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E803–E817. [Google Scholar] [CrossRef]
- Wilkinson, K.; Koscien, C.P.; Monteyne, A.J.; Wall, B.T.; Stephens, F.B. Association of postprandial postexercise muscle protein synthesis rates with dietary leucine: A systematic review. Physiol. Rep. 2023, 11, e15775. [Google Scholar] [CrossRef]
- Finnigan, T.; Needham, L.; Abbott, C. Chapter 19—Mycoprotein: A healthy new protein with a low environmental impact. In Sustainable Protein Sources; Nadathur, S.R., Wanasundara, J.P.D., Scanlin, L., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 305–325. [Google Scholar] [CrossRef]
- Borack, M.S.; Reidy, P.T.; Husaini, S.H.; Markofski, M.M.; Deer, R.R.; Richison, A.B.; Lambert, B.S.; Cope, M.B.; Mukherjea, R.; Jennings, K.; et al. Soy-dairy protein blend or whey protein isolate ingestion induces similar postexercise muscle mechanistic target of rapamycin complex 1 signaling and protein synthesis responses in older men. J. Nutr. 2016, 146, 2468–2475. [Google Scholar] [CrossRef]
- Reidy, P.T.; Walker, D.K.; Dickinson, J.M.; Gundermann, D.M.; Drummond, M.J.; Timmerman, K.L.; Cope, M.B.; Mukherjea, R.; Jennings, K.; Volpi, E.; et al. Soy-dairy protein blend and whey protein ingestion after resistance exercise increases amino acid transport and transporter expression in human skeletal muscle. J. Appl. Physiol. 2014, 116, 1353–1364. [Google Scholar] [CrossRef]
- Camera, D.M.; Smiles, W.J.; Hawley, J.A. Exercise-induced skeletal muscle signaling pathways and human athletic performance. Free Radic Biol. Med. 2016, 98, 131–143. [Google Scholar] [CrossRef]
- Trommelen, J.; van Loon, L.J.C. Quantification and interpretation of postprandial whole-body protein metabolism using stable isotope methodology: A narrative review. Front. Nutr. 2024, 11, 1391750. [Google Scholar] [CrossRef] [PubMed]
- Camera, D.M.; Burniston, J.G.; Pogson, M.A.; Smiles, W.J.; Hawley, J.A. Dynamic proteome profiling of individual proteins in human skeletal muscle after a high-fat diet and resistance exercise. FASEB J. 2017, 31, 5478–5494. [Google Scholar] [CrossRef] [PubMed]
- Medawar, E.; Huhn, S.; Villringer, A.; Veronica Witte, A. The effects of plant-based diets on the body and the brain: A systematic review. Transl. Psychiatry 2019, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef]
- Camilleri, M.; Lyle, B.J.; Madsen, K.L.; Sonnenburg, J.; Verbeke, K.; Wu, G.D. Role for diet in normal gut barrier function: Developing guidance within the framework of food-labeling regulations. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G17–G39. [Google Scholar] [CrossRef]
- Myles, I.A. Fast food fever: Reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef]
- Rodrigues, P.B.; Dátilo, M.N.; Sant’Ana, M.R.; Nogueira, G.; Marin, R.M.; Nakandakari, S.; de Moura, L.P.; da Silva, A.S.R.; Ropelle, E.R.; Pauli, J.R.; et al. The early impact of diets enriched with saturated and unsaturated fatty acids on intestinal inflammation and tight junctions. J. Nutr. Biochem. 2023, 119, 109410. [Google Scholar] [CrossRef]
- Seiwert, N.; Heylmann, D.; Hasselwander, S.; Fahrer, J. Mechanism of colorectal carcinogenesis triggered by heme iron from red meat. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188334. [Google Scholar] [CrossRef]
- Page, M.J.; Kell, D.B.; Pretorius, E. The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress 2022, 6, 24705470221076390. [Google Scholar] [CrossRef]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review article: Insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharmacol. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Wang, H.; Liang, F. Mechanisms linking inflammation to insulin resistance. Int. J. Endocrinol. 2015, 2015, 508409. [Google Scholar] [CrossRef] [PubMed]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct. Target Ther. 2023, 8, 375. [Google Scholar] [CrossRef] [PubMed]
- Camera, D.M.; Edge, J.; Short, M.J.; Hawley, J.A.; Coffey, V.G. Early time course of Akt phosphorylation after endurance and resistance exercise. Med. Sci. Sports Exerc. 2010, 42, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Fritz, P.; Fritz, R.; Bóday, P.; Bóday, Á.; Bató, E.; Kesserű, P.; Oláh, C. Gut microbiome composition: Link between sports performance and protein absorption? J. Int. Soc. Sports Nutr. 2024, 21, 2297992. [Google Scholar] [CrossRef]
- Jäger, R.; Zaragoza, J.; Purpura, M.; Iametti, S.; Marengo, M.; Tinsley, G.M.; Anzalone, A.J.; Oliver, J.M.; Fiore, W.; Biffi, A.; et al. Probiotic administration increases amino acid absorption from plant protein: A placebo-controlled, randomized, double-blind, multicenter, crossover study. Probiotics Antimicrob. Proteins 2020, 12, 1330–1339. [Google Scholar] [CrossRef]
- Keller, D.; Van Dinter, R.; Cash, H.; Farmer, S.; Venema, K. Bacillus coagulans GBI-30, 6086 increases plant protein digestion in a dynamic, computer-controlled in vitro model of the small intestine (TIM-1). Benef. Microbes 2017, 8, 491–496. [Google Scholar] [CrossRef]

{kind=link}
| Author, Country (PMID) | Participant Demographic (N, Male% and Age M ± SD) | Dietary Groups | Minimum Duration of Diet | Dietary Analysis | Method of Gut Microbiota and Metabolomic Assessment | Composition of Gut Microbiota—Main Findings |
|---|---|---|---|---|---|---|
| Reddy et al., 1975 ([24]) USA (1138032) | N = 8 Sex: Both but not specified Age: 30–50 yrs | Habitual mixed Western diet (n = 8) Non-meat diet (n = 8) | No details for Western diet. Four-week intervention after consuming habitual Western diet. | NA | One stool sample per person for 4 consecutive days. Only one sample used for microbial analysis. Traditional plate count culture. |
|
| Zimmer et al., 2012 ([25]) Germany (21811294) | N = 498 Sex: Vegan 43%, Vegetarian 34%, Omnivore 38% Age: Vegan 49.2 ± 14.6 yrs, Vegetarian 56.7 ± 15.1 yrs, Omnivore 53.7 ± 14.8 yrs | Vegan (n = 105) Vegetarian (n = 144) Omnivore (n = 249) | Not specified for each group (≥one month inclusion criteria). | Short questionnaire that included specific food intake. | One stool sample per person. Viable bacterial cell counts via agar plates. |
|
| Kabeerdoss et al., 2012 ([26]) India (22182464) | N = 56 Sex: 0% Age *: Vegetarian: 19 yrs (18–19) Omnivore: 19 yrs (18–20) | Lacto-vegetarian (n = 32) Omnivore (n = 24) | Not specified. | 24 h diet recall and FFQ (3 months). | One stool sample per person. 16srRNA sequencing (variable region not specified). |
|
| Matijasic et al., 2014 ([27]) Slovenia (24173964) | N = 60 Sex: Vegan 60%, Lacto-vegetarian 58.3%, Vegetarian 59.4%, Omnivore 43.3% Age **: Vegan 35 (2–63) Lacto-vegetarian 34 (30–67) Vegetarian 35 (2–67) Omnivore 30 (1.5–61) | Vegan (n = 20) Lacto-vegetarian (n = 11) Vegetarian (n = 31) Omnivore (n = 29) | Not specified for each group (≥12 months inclusion criteria). | In-house questionnaire including consumption of particular foods. | One stool sample per person. PCR-DGGE finger printing of 16S rRNA V3 region. |
|
| Ferrocino et al., 2015 ([28]) Italy (26035837) | N = 153 Sex: 35% in Ovo-lacto-vegetarian, 45% in both Vegan and Omnivore Age: Ovo-lacto-vegetarian (39.0 ± 9.0) Vegan (37.0 ± 10.0) Omnivore (37.0 ± 9.0) | Ovo-lacto-vegetarian (n = 51) Vegan (n = 51) Omnivore (n = 51) | Not specified for each group (≥12 months inclusion criteria). | Not stated. | One stool sample per person. Ribosomal RNA Denaturing Gradient Gel Electrophoresis (rRNA-DGGE). |
|
| De Filippis et al., 2016 ([29]) Italy (27769291) | N = 153 Sex: 35% Ovo-lacto-vegetarian, 45% in both Vegan and Omnivore Age: Ovo-lacto-vegetarian (39.0 ± 9.0) Vegan (37.0 ± 10.0) Omnivore (37.0 ± 9.0) | Ovo-lacto-vegetarian (n = 51) Vegan (n = 51) Omnivore (n = 51) | Not specified for each group (≥ 12 months inclusion criteria). | 7-day weighed food diary. | One stool sample per person. 16S rRNA gene sequencing (V1-V3 region). |
|
| Wu et al., 2016 ([30]) USA (25431456) | N = 31 Sex: Not specified Age: Not specified (18–40 yrs) | Vegan (n = 15) Omnivore (n = 16) | Not specified for each group (≥ six months inclusion criteria). | 3 × 24 h dietary recalls. | One stool sample per person. 16S rRNA sequencing (V1 and V2 regions). Metabolite profiling (LC-MS and GC-MS). |
|
| Losasso et al., 2018 ([31]) Italy (29556222) | N = 101 Sex: Vegetarian (30%), Vegan (34.7%), Omnivore (26.7%) Age: Vegetarian: 42.3 ± 13.2 yrs Vegan: 39.4 ± 11.1 yrs Omnivore: 45.0 ± 13.9 yrs | Vegetarian (n = 32) Vegan (n = 26) Omnivorous (n = 43) | Not specified for each group (≥ 12 months inclusion criteria). | 14-day semi-quantitative FFQ. 24 h recall. | One stool sample per person. 16S rRNA sequencing (V3 and V4 regions). |
|
| Trefflich et al., 2021 ([32]) Germany (34073495) | N = 72 Sex: 50% for both Vegan and Omnivore Age: Vegan: 37.5 yrs Omnivore: 38.5 yrs | Vegan (n = 36) Omnivorous (n = 36) | Vegan: 4.8 yrs. Omnivore: No detail. | 3-day weighed food diary. | One stool sample per person. 16S rRNA sequencing (V3 and V4 regions). Metabolite profiling (GC-MS). |
|
| Prochazkova et al., 2022 ([33]) Czech Republic (35071294) | N = 95 Sex: Vegan (60%), Omnivore (48%) Age: Vegan: 30.9 ± 10.5 yrs Omnivore: 31.3 ± 11.2 yrs | Vegan (n = 62) Omnivore (n = 33) | Not specified for each group (≥ 36 months inclusion criteria). | 3-day prospective dietary record supervised by trained dietitian. | One stool sample per person. 16S rRNA sequencing (V4 region). Metabolite profiling (NMR and LC/GC-MS). |
|
| Stege et al., 2022 ([34]) Netherlands (35115599) | N = 149 Sex: Vegetarian (32%), Vegan (34%), Pescatarian (36%), Omnivore (32%) Age ***: Vegetarian 45 yrs Vegan 37 yrs Pescatarian 51 yrs Omnivore 47 yrs | Vegetarian (n = 34) Vegan (n = 32) Pescatarian (n = 33) Omnivore (n = 50) | Not specified for each group (≥ six months inclusion criteria). | None. | One stool sample per person. Metagenomic shotgun sequencing. ResCap sequencing (resistome analysis). |
|
| Tarallo et al., 2022 ([35]) Italy (34315772) | N = 120 Sex: 40% in each group Age: Vegetarian (40.6 ± 11.7) Vegan (39.1 ± 11.6) Omnivore (40.5 ± 13.2) | Vegetarian (n = 40) Vegan (n = 40) Omnivore (n = 40) | Vegetarian: 9.7 ± 9.0 yrs Vegan: 8 ± 2.5 yrs | FFQ. | One stool sample per person. Metagenomic shotgun sequencing. |
|
| Seel et al., 2023 ([36]) Germany (37111133) | N = 258 Sex: 38% Omnivore 20% Flexitarian 28% Ovo-lacto-vegetarian 29% Vegan Age *: Omnivore 33.0 (17.0) Flexitarian 29.5 (16.8) Ovo-lacto-vegetarian 28.0 (14.0) Vegan 25.0 (9.8) | Ovo-lacto-vegetarian (n = 65, n = 51 final), Vegan (n = 58, final 52), Flexitarian (n = 70, n = 52 final), Omnivore (n = 65 screening, n = 51 final) | Ovo-lacto-vegetarian 6.0 ± 10.0 yrs, Vegan: 3.0 ± 3.0 yrs, Flexitarian: 8.0 ± 17.8 yrs, Omnivore 32.0 ± 20.0 yrs. | 5-day self-reported dietary record. | One stool sample for each participant. 16S rRNA gene sequencing (V3-V4 region). |
|
| Fackelmann et al., 2025 ([37]) Italy, UK, and USA (39762435) | N = 21,561 Sex: Not included Age: 52 ± 12.5 yrs | Vegetarian (n = 1088), Vegan (n = 656), Omnivorous (n = 19,817) | Not specified for each group (≥ 12 months inclusion criteria). | Food Frequency Questionnaire. | One stool sample per person. Metagenomic shotgun sequencing. |
|
| Author, Country (PMID) | Participant Demographic (N, Male% and Age M ± SD) | Dietary Groups | Study Design and Intervention Duration | Dietary Analysis | Method of Gut Microbiota and Metabolomic Assessment | Composition of Gut Microbiota—Main Findings |
|---|---|---|---|---|---|---|
| van Faassen et al., 1987 ([38]) Netherlands (3120571) | N = 12 Sex: 100% Age: Not specified, age range 20–27 yrs | Mixed Western (n = 12), Lacto-ovo-vegetarian (n = 12), Vegan (n = 12) in cross-over design Mixed Western: 38% fat, 17% protein. Lacto-ovo-vegetarian: 32% fat, 16% protein (0% meat. Vegan: 33% fat, 17% protein. All meals were provided in a controlled metabolic ward setting in randomized order. | Cross-over study design. 20 days for each diet period, with an 8-day adaptation period on mixed diet first. No details regarding washout length provided. | NA | Two stool samples collected per person. Traditional plate count (Agar- based culturing). |
|
| David et al., 2014 ([39]) USA (24336217) | N = 11 (n = 10 per diet; 9 individuals completed both diet interventions) Sex:55% Age: 28.1 ± 3.72 yrs | Plant-based diet (n = 9). Animal-based diet (n = 9). Animal-based diet: dietary fat 69.5 ± 0.4% kcal, dietary protein 30.1 ± 0.5% kcal, and dietary fiber ~0%. Plant-based diet: dietary fiber 25.6 ± 1.1 g/1000 kcal, dietary fat 22.1 ± 1.7% kcal, and dietary protein 10.0 ± 0.3% kcal. | Cross-over study design. Five-day controlled diet intervention with meals provided. Six-day wash out period. | NA | Daily stool samples from each participant. 16S rRNA gene sequencing (V4 region). |
|
| Zhang et al., 2018 ([40]) China, Sweden, and USA (29755475) | N = 29 Sex: 47% Omnivore to lacto-ovo-vegetarian 71% Long-term omnivore 43% Long-term vegetarian Age: Omnivore to lacto-ovo-vegetarian 35.4 ± 9.8 yrs Long-term omnivore 35.7 ± 10.3 yrs Long-term vegetarian 33.7 ± 5.8 yrs | Study groups: Omnivore to lacto-ovo-vegetarian (n = 15); Control 1: long-term omnivore (n = 7); Control 2: long-term vegetarian (n = 7). Study group changed from omnivorous to lacto-ovo-vegetarian diet for 3 months (self-selected foods, no meals provided). Control Group 1: Maintained regular omnivorous diet (no dietary changes required). Control Group 2: Long-term vegetarians maintained their lacto-ovo-vegetarian diet (no dietary changes required). | Single arm, unblinded study design. 3-month diet change from omnivore to lacto-ovo-vegetarian. The other 2 groups were monitored during that time and continued their usual diet (long-term omnivore and vegetarian). | 3-day weighted food diary and dietary questionnaire. | One stool sample per person at days 0 and 91. Metagenomic shotgun sequencing. |
|
| Kohnert et al., 2021 ([41]) Germany (33807447) | N = 53 Sex: 31% Vegan, 44% Meat-rich Age: Vegan: 33.2 ± 11.2 Meat-rich: 29.9 ± 9.5 | Meat-rich diet (>150 g of meat per day, n = 27). Strict vegan (n = 26). Every participant received extensive training on his/her assigned diet and detailed written information, including a recipe book. No meals were provided, and participants were free to choose their food within their assigned diet. | Monocentric, controlled, randomized trial with a parallel group design. Four-week intervention. | NA | Two stool samples were collected at the beginning and at the end of the trial. 16S rRNA gene sequencing (V4 region). |
|
| Study (PMID) | Cohort Details | Vegan Protein Source | Comparison Protein Source | Exercise | Key Findings |
|---|---|---|---|---|---|
| Wilkinson et al., 2007 ([53]) (17413102) | 8 healthy men (21.6 ± 0.3 y; mean ± SEM) | Isonitrogenous, isoenergetic, and macronutrient-matched soy beverage (18 g protein, 750 kJ) | Isonitrogenous, isoenergetic, and macronutrient-matched milk beverage (18 g protein, 750 kJ) | Unilateral leg press, hamstring curl, and knee extension: 3 × 10 reps @ 80% of 1 RM + 1 × AMRAP of each exercise |
|
| Tang et al., 2009 ([15]) (19589961) | 3 × 6 groups of resistance-trained young males (22.8 ± 3.9 y; mean ± SD) | Soy (22.2 g) protein dissolved in 250 mL water with sucralose (1 g Splenda) | Whey (21.4 g) OR Casein (21.9 g) protein dissolved in 250 mL water with sucralose (1 g, Splenda) | Unilateral leg press and knee extension (4 × AMRAP @ 10- to 12-RM) |
|
| Yang et al., 2012 ([54]) (22698458) | 30 older males (age 71 ± 5 y; mean ± SD) | 20 g or 40 g soy protein isolate (S20 and S40) | 0 g, 20 g, or 40 g of whey protein isolate (W20 and W40) | Unilateral knee-extensor exercise (3 × AMRAP @ 10 RM) |
|
| Gorissen et al., 2016 ([55]) (27440260) | 60 healthy older men (71 ± 1 y; mean ± SEM) | 35 g wheat protein (n = 12), 35 g wheat protein hydrolysate (WPH-35; n = 12), or 60 g wheat protein hydrolysate (WPH-60; n = 12) | 35 g micellar casein (MCas-35; n = 12), 35 g whey protein (Whey-35; n = 12), hydrolysate (WPH-60; n = 12) | No exercise |
|
| Oikawa et al., 2020 ([56]) (32349353) | 24 healthy females (20 ± 1 y; mean ± SD) | Potato protein (25 g providing 1.6 g/kg body mass per d total protein) | Control (0.8 g/kg body mass per d total protein) | 3 d/wk unilateral leg press and extension: 6 sets @30% 1-RM |
|
| Pinckaers et al., 2021 ([13]) (33597056) | 36 healthy young males (23 ± 3 y; mean ± SD) | 30 g wheat protein (WHEAT) | 30 g milk protein (MILK) OR 30 g blend combining 15 g wheat plus 15 g milk protein (WHEAT + MILK) | No exercise |
|
| Pinckaers et al., 2022 ([57]) (35438672) | 24 healthy young males (24 ± 4 y; mean ± SD) | 30 g potato-derived protein | 30 g milk protein | Unilateral leg press (3 × 8 reps @ ~80% 1-RM + AMRAP) + unilateral seated knee extension machine (3 × of 8 reps @ ~80% 1-RM + AMRAP) |
|
| Kouw et al., 2022 ([58]) (34881688) | 24 men (age 24 ± 5 y; mean ± SD) | 40 g of protein as a lysine-enriched wheat and chickpea protein product (plant, n = 12) | 40 g chicken breast fillet (chicken, n = 12) | No exercise |
|
| Davies et al., 2022 ([59]) (36145064) | 16 healthy adults (age = 25 ± 5; mean ± SD) | 0.33 g·kg fava bean protein (FBP) | 0.33 g·kg control (CON) beverage (EAA-free mixture) | 6 × 10 maximal effort unilateral (dominant limb) isokinetic knee extensor contractions (concentric and eccentric) at a velocity of 90°·s−1 |
|
| Pinckaers et al., 2023 ([60]) (36170964) | 24 young males (aged 24 ± 4 y; mean ± SD) | 30 g plant-derived protein blend combining 15 g wheat, 7.5 g corn, and 7.5 g pea protein (PLANT-BLEND) | 30 g milk protein (MILK) | No exercise |
|
| Pinckaers et al., 2024 ([14])(38228945) | 24 young males (24 ± 3 y; mean ± SD) | 30 g pea (PEA) | 30 g milk-derived protein (MILK) | No exercise |
|
| Pinckaers et al., 2024 ([61]) (37972895) | 16 older adults (8 males, 8 females) (65–85 y; mean ± SD) | Isonitrogenous and isocaloric whole-food vegan meal (PLANT) | Whole-food omnivorous meal containing beef as the primary source of protein (0.45 g protein/kg body mass; MEAT) | No exercise |
|
| Pinckaers et al., 2024 ([62]) (38315260) | 36 healthy young males (26 ± 4 y; mean ± SD) | 30 g corn protein (CORN) | 30 g milk protein (MILK) or a 30 g protein blend with 15 g corn plus 15 g milk protein (CORN + MILK) | No exercise |
|
| Monteyne et al., 2020 ([63]) (32438401) | 20 resistance-trained healthy young males (age: 22 ± 1 y; mean ± SEM) | 70 g (31.5 g protein/2.5 g leucine) mycoprotein (MYCO) | 31 g (26.2 g protein/2.5 g leucine) milk protein (MILK) | 5 × 30 maximal concentric isokinetic leg-extension and leg-flexion contractions on a Biodex System 3 isokinetic dynamometer |
|
| Monteyne et al., 2020 ([64]) (32886108) | 19 healthy males (22 ± 1 y; mean ± SEM) | 70 g mycoprotein (31.5 g protein; MYCO; n = 10) OR 35 g BCAA-enriched mycoprotein (18.7 g protein: matched on BCAA content; ENR; n = 9) | N/A | 5 × 30 maximal concentric isokinetic leg-extension and leg-flexion contractions on a Biodex System 3 isokinetic dynamometer |
|
| Monteyne et al., 2021 ([65]) (33172506) | 19 healthy older adults (66 ± 1 y; mean ± SEM) | 3 d isoenergetic high-protein (1·8 g/kg body mass per d) diet, where the protein was provided from exclusively vegan (VEG; n 10; six males, four females; mycoprotein providing 57% of daily protein intake) sources | 3 d isoenergetic high-protein (1·8 g/kg body mass per d) diet, where the protein was provided from predominantly (71%) animal (OMNI; n 9; six males, three females) sources | Unilateral leg extension exercise (5 × 30 reps of maximal concentric isokinetic knee extension contractions of their dominant leg) |
|
| Monteyne et al., 2023 ([66]) (36822394) | 16 healthy young adults (8 males and 8 females) (23 ± 1 y; mean ± SD) | 3 d dietary intervention (high protein, 1.8 g/kg body mass per d) where protein was derived from exclusively non-animal (VEG1; n = 8) sources | 3 d dietary intervention (high protein, 1.8 g/kg body mass per d) where protein was derived from omnivorous (OMNI1; n = 8) sources | Unilateral leg extension exercise (5 × 30 reps of maximal concentric isokinetic knee extension contractions of their dominant leg) |
|
| Churchward-Venne et al., 2019 ([67]) (30698812) | 36 healthy young males (23 ± 0.4 y; mean ± SEM) | 45 g carbohydrate + 20 g soy protein OR leucine-enriched soy (same level of leucine as in whey) | 45 g carbohydrate + 20 g whey | Concurrent exercise: 4 × 8 reps @ ∼80% 1 RM + 30 min of continuous cycling at ∼60% of their previously determined maximal workload (Wmax) |
|
| West et al., 2023 ([68]) (37529834) | 33 healthy, young females and males (age: 21 ± 1 y; mean ± SEM) | 25 g of protein from mycoprotein (MYC, n = 11), pea protein (PEA, n = 11), or a blend (39% MYC and 61% PEA) of the two (BLEND, n = 11) | N/A | Whole-body REX program (1 × 10 reps @ 75% 10-RM + 3 × AMRAP @ 10 RM) |
|
| West et al., 2023 ([69]) (36172885) | 24 healthy young (age, 21 ± 2 y; mean ± SEM) | 70 g mycoprotein (MYC; 31·4 g protein and 2·5 g leucine; n 12) or 38·2 g of a protein concentrate obtained from mycoprotein (PCM; 28·0 g protein and 2·5 g leucine; n 12) | N/A | 5 × 30 maximal concentric isokinetic leg-extension and leg-flexion contractions on a Biodex System 3 isokinetic dynamometer |
|
| van der Heijden et al., 2023 ([70]) (37716611) | 36 healthy young adults females and males (age: 22 ± 3 y; mean ± SD) | 25 g protein from (fungal-derived mycoprotein [MYCO]) (n = 12), Spirulina [SPIR] (cyanobacterium) (n = 12), or chlorella [CHLO] (microalgae) (n = 12; m/f, 6/6) | N/A | 4 unilateral sets of leg press, followed by 4 sets of leg extension at the predetermined 10 RM until volitional failure (8–12 repetitions) |
|
| Lim et al., 2024 ([71]) (38846451) | 8 healthy and recreationally active (men: n = 4; women: n = 4; 18–30 y; mean ± SD) | 25 g WHEY plant-based blend protein (88% pea protein and 12% canola protein or plant-based blend protein + leucine matched with WHEY; PBP) | 25 g WHEY (SureProtein WPI 895) | No exercise |
|
| van der Heijden et al., 2024 ([72]) (38537270) | 10 resistance-trained, young adults (male/female: 8/2; age: 26 ± 6 y; mean ± SD) | 32 g plant protein blend (BLEND; 39.5% pea, 39.5% brown rice, and 21.0% canola) | 32 g protein from whey (WHEY) | Bilateral leg resistance exercise |
|
| Domic et al., 2024 ([73] (39732437) | 34 community-dwelling older adults (18 males and 16 females) (72 ± 4 y; mean ± SD) | 1.3 g/kg body mass per d protein from all vegan sources: soy-based dairy alternatives, legumes, nuts, cereals, and plant-based meat analogues based on (isolated) pea protein and/or rice protein | 1.3 g/kg body mass per d protein from omnivorous sources: dairy products, cheese, chicken, beef, and pork sausage | No exercise |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Refai, W.; Keenan, S.; Camera, D.M.; Cooke, M.B. The Influence of Vegan, Vegetarian, and Omnivorous Diets on Protein Metabolism: A Role for the Gut–Muscle Axis? Nutrients 2025, 17, 1142. https://doi.org/10.3390/nu17071142
Al-Refai W, Keenan S, Camera DM, Cooke MB. The Influence of Vegan, Vegetarian, and Omnivorous Diets on Protein Metabolism: A Role for the Gut–Muscle Axis? Nutrients. 2025; 17(7):1142. https://doi.org/10.3390/nu17071142
Chicago/Turabian StyleAl-Refai, Waed, Stephen Keenan, Donny M. Camera, and Matthew B. Cooke. 2025. "The Influence of Vegan, Vegetarian, and Omnivorous Diets on Protein Metabolism: A Role for the Gut–Muscle Axis?" Nutrients 17, no. 7: 1142. https://doi.org/10.3390/nu17071142
APA StyleAl-Refai, W., Keenan, S., Camera, D. M., & Cooke, M. B. (2025). The Influence of Vegan, Vegetarian, and Omnivorous Diets on Protein Metabolism: A Role for the Gut–Muscle Axis? Nutrients, 17(7), 1142. https://doi.org/10.3390/nu17071142