
Critical Review on Anti-Obesity Effects of Anthocyanins Through PI3K/Akt Signaling Pathways




Abstract

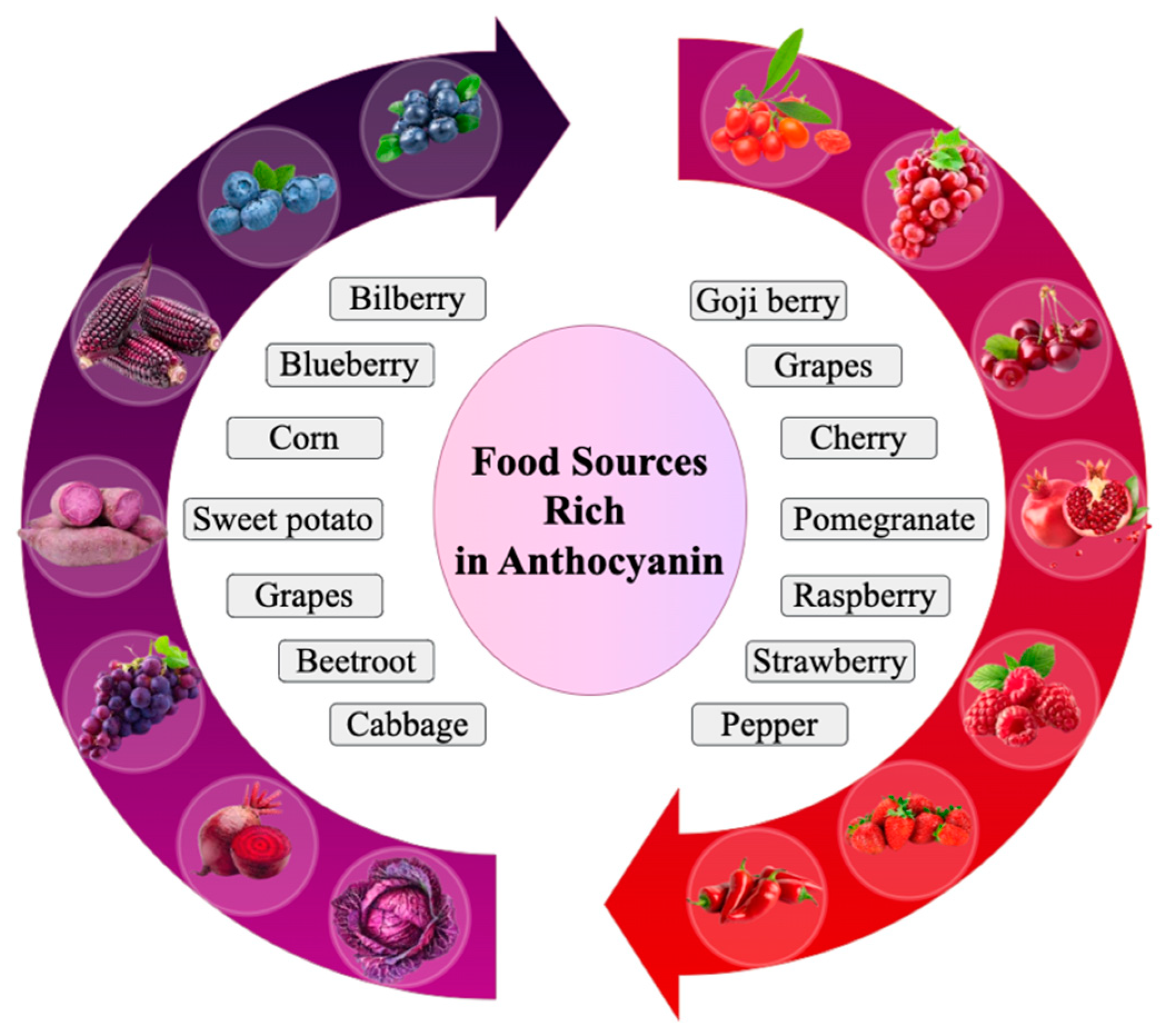
1. Introduction
2. Methodology
3. Anthocyanins and Their Metabolic Impact
4. Molecular Mechanisms of Anthocyanins in Obesity Through PI3K/Akt Signaling Pathway
4.1. Effect of Anthocyanins on PI3K/Akt/GLUT4 Signaling Pathway in Obesity
4.2. Effect of Anthocyanins on PI3K/Akt/FOXO Signaling Pathway in Obesity
4.3. Effect of Anthocyanins on PI3K/Akt/mTOR Signaling Pathway in Obesity
4.4. Effect of Anthocyanins on PI3K/Akt/ GSK3 Signaling Pathway in Obesity
4.5. Cross Link Between PI3K/Akt and AMPK Signaling Pathways
4.6. Role of Anthocyanins on Oxidative Stress and Inflammation in Obesity Through PI3K/Akt Signaling Pathway
5. Clinical Applications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Matrix | Trial Number | Study Design. Length. Doses | Overweight/Obese Patients: n, (Gender), Age | Main Findings | Ref. |
|---|---|---|---|---|---|
| Açaí pulp | RBR-72dvqv | Double-blind RPCT 90 days 200 g Açaí pulp. Anthocyanin content was not measured. | 69 volunteers | ↓Expression of IFN-γ and IL-6 ↓Plasma 8-isoprostane concentration levels ↓Body mass and BMI | [87] |
| Medox capsules; anthocyanins from bilberry and blackcurrant | NCT03415503 | Double-blind RPCT 12 weeks 3 doses of anthocyanins supplement (40, 80, and 320 mg/d) 2 Capsules twice a day | 169 dyslipidemic participants female and male, 35–70 years | ↑Total superoxide dismutase ↓Expression of serum IL-6, TNF-α ↓Expression of urinary 8-iso-PGF2α | [91] |
| Medox capsules; anthocyanins from bilberry and blackcurrant | NCT03415503 | Double-blind RPCT 12 weeks 3 doses of anthocyanins (40, 80, and 320 mg/d) 2 Capsules twice a day | 176 dyslipidemic subjects female and male, 35–70 years | Significant difference in cholesterol efflux capacity, HDL-C, and apoA1 | [93] |
| Juçara berry (Euterpe edulis Mart.) | CEP-UNIFESP No. 0319/2017 | Double-blind RPCT 6 weeks 5 g Juçara freeze-dried pulp Anthocyanin content was not measured. | 27 obese adult female and male, 31–59 years, BMI: 30.0–39.9 kg/m2 | ↓Expression of TLR4, & IL-6 mRNA ↓Expression of IL-6, TNF-α, and MCP-1 | [92] |
| Black rice anthocyanin | CRIS, KCT0005836 | Double-blind RPCT 12-week 500 mg twice daily | 105 participants Female 45–69 years BMI ≥ 25 kg/m2 | ↓Visceral fat region, visceral/total (%), and visceral/subcutaneous (%) ↓Trunk fat and total fat mass ↓Body weight and BMI | [90] |
| Freeze-dried strawberry powders | NCT03441620 | Multicenter, double-blind randomized controlled trial 14 weeks 13 g/day and 32 g/day strawberry powder/day | Female and male, BMI ≥ 30 kg/m2, | ↓Serum LDL-C ↓Postprandial PAI-1 levels | [88] |
| Freeze-dried strawberry powders | NCT02557334 | Double-blinded, RPCT, crossover trial 6 weeks 13 g/d (low-dose), and 40 g/d (high-dose) | Healthy volunteers BMI = 29.4 kg/m2 50 ± 1.0 years | ↓TC levels | [94] |
| Organic bilberry juice | Ethical Committee of the Specialized Hospital of St. Svorad Zobor in Nitra, Slovakia (Study No. 4/071220/202 | Single-arm pre-post intervention study 125 mL/day of 100% bilberry juice or 10 g/day of 100% bilberry fiber | 30 participants, female, BMI > 30 kg/m2, 50–60 years | ↓LDL-C and increased HDLC | [89] |
6. Anthocyanins in Functional Foods and Pharmacological Synergy for Obesity
6.1. Mechanisms of Anthocyanins in Functional Foods
6.2. Types of Functional Foods with Anthocyanins
6.3. Management of Anthocyanins as Adjuncts to Anti-Obesity Therapies
6.4. Challenges in the Development of Anthocyanin-Based Functional Foods for Obesity Management
6.4.1. Anthocyanin Bioavailability and Stability
6.4.2. Variability in Anthocyanin Content
7. Future Directions
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| Akt | Protein kinase B |
| BW | Body weight |
| C3G | Cyanidin-3-glucoside |
| DNL | De novo lipogenesis |
| EDL | Extensor digitorum longus |
| FOXO1 | Forkhead box O1 |
| GLUT4 | Glucose transporter type 4 |
| GPCR | G-protein-coupled receptors |
| GPx | Glutathione peroxidase |
| GSK3 | Glycogen Synthase Kinase 3 |
| IL | Interleukin |
| IR | Insulin resistance |
| IRS | Insulin receptor substrates |
| MAPK | Mitogen-activated protein kinase |
| MAFLD | Metabolic dysfunction-associated fatty liver disease |
| mTOR | Mechanistic target of rapamycin |
| NF-κB | Nuclear factor-kappa B |
| PDK1 | Phosphoinositide-dependent kinase 1 |
| PI3K | Phosphoinositide 3-kinase |
| PI3K/Akt | Phosphoinositide 3-kinase/protein kinase B |
| PIP2 | Phosphatidylinositol-4,5-bisphosphate |
| PIP3 | Phosphatidylinositol-3,4,5-trisphosphate |
| Rheb | Ras homolog enriched in the brain |
| ROS | Reactive oxygen species |
| RTK | Receptor tyrosine kinases |
| SGBS | Human Simpson–Golabi–Behmel syndrome |
| SOD | Superoxide dismutase |
| T2D | Type 2 diabetes |
| TNF | Tumor necrosis factor |
| TSC1/2 | Tuberous sclerosis complex 1/2 |
| ULK1 | Unc-51 Like Autophagy Activating Kinase 1 |
| WHO | World Health Organization |
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 20 February 2025).
- Geetha, V. Drug discovery for diabetes, obesity, and non-alcoholic fatty liver disease (NAFLD). In Multidisciplinary Research in Arts, Science & Commerce; Hill Publication: Elmhurst, NY, USA, 2024; Volume 15, p. 36. [Google Scholar]
- Han, J.C.; Weiss, R. Obesity, metabolic syndrome and disorders of energy balance. In Sperling Pediatric Endocrinology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 939–1003. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Tak, Y.J.; Lee, S.Y. Anti-obesity drugs: Long-term efficacy and safety: An updated review. World J. Mens Health 2021, 39, 208–221. [Google Scholar] [CrossRef]
- Williams, D.M.; Nawaz, A.; Evans, M. Drug therapy in obesity: A review of current and emerging treatments. Diabetes Ther. 2020, 11, 1199–1216. [Google Scholar] [CrossRef]
- Garcia, C.; Blesso, C.N. Antioxidant properties of anthocyanins and their mechanism of action in atherosclerosis. Free Radic. Biol. Med. 2021, 172, 152–166. [Google Scholar] [CrossRef]
- Les, F.; Cásedas, G.; Gómez, C.; Moliner, C.; Valero, M.S.; López, V. The role of anthocyanins as antidiabetic agents: From molecular mechanisms to in vivo and human studies. J. Physiol. Biochem. 2021, 77, 109–131. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Du, B.; Li, J.; Yang, Y.; Zhu, F. An insight into anti-inflammatory activities and inflammation related diseases of anthocyanins: A review of both in vivo and in vitro investigations. Int. J. Mol. Sci. 2021, 22, 11076. [Google Scholar] [CrossRef]
- Escalante-Aburto, A.; Mendoza-Córdova, M.Y.; Mahady, G.B.; Luna-Vital, D.A.; Gutiérrez-Uribe, J.A.; Chuck-Hernández, C. Consumption of dietary anthocyanins and their association with a reduction in obesity biomarkers and the prevention of obesity. Trends. Food Sci. Technol. 2023, 140, 104140. [Google Scholar] [CrossRef]
- Gull, A.; Sheikh, M.A.; Kour, J.; Zehra, B.; Zargar, I.A.; Wani, A.A.; Lone, M.A. Anthocyanins. In Nutraceuticals and Health Care; Academic Press: Cambridge, MA, USA, 2022; pp. 317–329. [Google Scholar] [CrossRef]
- Savova, M.S.; Mihaylova, L.V.; Tews, D.; Wabitsch, M.; Georgiev, M.I. Targeting PI3K/AKT signaling pathway in obesity. Biomed. Pharmacother. 2023, 159, 114244. [Google Scholar] [CrossRef]
- Nistor, M.; Pop, R.; Daescu, A.; Pintea, A.; Socaciu, C.; Rugina, D. Anthocyanins as key phytochemicals acting for the prevention of metabolic diseases: An overview. Molecules 2022, 27, 4254. [Google Scholar] [CrossRef]
- Molonia, M.S.; Quesada-Lopez, T.; Speciale, A.; Muscarà, C.; Saija, A.; Villarroya, F.; Cimino, F. In vitro effects of Cyanidin-3-O-glucoside on inflammatory and insulin-sensitizing genes in human adipocytes exposed to palmitic acid. Chem. Biodivers. 2021, 18, e2100607. [Google Scholar] [CrossRef]
- Shyur, L.F.; Varga, V.; Chen, C.M.; Mu, S.C.; Chang, Y.C.; Li, S.C. Extract of white sweet potato tuber against TNF-α-induced insulin resistance by activating the PI3K/Akt pathway in C2C12 myotubes. Bot. Stud. 2021, 62, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.W.; Lü, H.; Du, L.L.; Li, J.; Chen, H.; Zhao, H.F.; Li, W.L. Five blueberry anthocyanins and their antioxidant, hypoglycemic, and hypolipidemic effects in vitro. Front. Nutri. 2023, 10, 1172982. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Zhou, Y.; Zheng, X. Black rice anthocyanins alleviate hyperlipidemia, liver steatosis and insulin resistance by regulating lipid metabolism and gut microbiota in obese mice. Food Funct. 2021, 12, 10160–10170. [Google Scholar] [CrossRef]
- Kongthitilerd, P.; Suantawee, T.; Cheng, H.; Thilavech, T.; Marnpae, M.; Adisakwattana, S. Anthocyanin-enriched riceberry rice extract inhibits cell proliferation and adipogenesis in 3T3-L1 preadipocytes by downregulating adipogenic transcription factors and their targeting genes. Nutrients 2020, 12, 2480. [Google Scholar] [CrossRef] [PubMed]
- Molonia, M.S.; Occhiuto, C.; Muscarà, C.; Speciale, A.; Bashllari, R.; Villarroya, F.; Saija, A.; Cimino, F.; Cristani, M. Cyanidin-3-O-glucoside restores insulin signaling and reduces inflammation in hypertrophic adipocytes. Arch. Biochem. Biophys. 2020, 691, 108488. [Google Scholar] [CrossRef]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and human health-A focus on oxidative stress, inflammation and disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef]
- Zhang, Y.; Chang, H.; Shao, S.; Zhao, L.; Zhang, R.; Zhang, S. Anthocyanins from Opuntia ficus-indica modulate gut microbiota composition and improve short-chain fatty acid production. Biology 2022, 11, 1505. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, M.; Li, D.; Miao, X.; Hua, M.; Sun, R.; Niu, H. Black rice anthocyanidins regulates gut microbiota and alleviates related symptoms through PI3K/AKT pathway in type 2 diabetic rats. J. Food Biochem. 2023, 2023, 5876706. [Google Scholar] [CrossRef]
- Ngamsamer, C.; Sirivarasai, J.; Sutjarit, N. The benefits of anthocyanins against obesity-induced inflammation. Biomolecules 2022, 12, 852. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Z.; Sun, H.; He, S.; Liu, S.; Zhang, T.; Ma, G. Research progress of anthocyanin prebiotic activity: A review. Phytomedicine 2022, 102, 154145. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Agrizzi Verediano, T.; Agarwal, N.; Stampini Duarte Martino, H.; Kolba, N.; Grancieri, M.; Dias Paes, M.C.; Tako, E. Effect of black corn anthocyanin-rich extract (Zea mays L.) on cecal microbial populations In Vivo (Gallus gallus). Nutrients 2022, 14, 4679. [Google Scholar] [CrossRef]
- Wu, Y.; Han, Y.; Tao, Y.; Li, D.; Xie, G.; Show, P.L.; Lee, S.Y. In vitro gastrointestinal digestion and fecal fermentation reveal the effect of different encapsulation materials on the release, degradation and modulation of gut microbiota of blueberry anthocyanin extract. Food Res. Int. 2020, 132, 109098. [Google Scholar] [CrossRef]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef]
- Wen, X.; Zhang, B.; Wu, B.; Xiao, H.; Li, Z.; Li, R.; Xu, X.; Li, T. Signaling pathways in obesity: Mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 298. [Google Scholar] [CrossRef]
- Naomi, R.; Teoh, S.H.; Embong, H.; Balan, S.S.; Othman, F.; Bahari, H.; Yazid, M.D. The role of oxidative stress and inflammation in obesity and its impact on cognitive impairments—A narrative review. Antioxidants 2023, 12, 1071. [Google Scholar] [CrossRef]
- Tong, Y.; Xu, S.; Huang, L.; Chen, C. Obesity and insulin resistance: Pathophysiology and treatment. Drug Discov. Today. 2022, 27, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.; Fang, X.; Wei, J.; Wu, H.; Wang, X.; Tian, J. Akt: A potential drug target for metabolic syndrome. Front. Physiol. 2022, 13, 822333. [Google Scholar] [CrossRef]
- Kim, J.N.; Han, S.N.; Kim, H.K. Anti-inflammatory and anti-diabetic effect of black soybean anthocyanins: Data from a dual cooperative cellular system. Molecules 2021, 26, 3363. [Google Scholar] [CrossRef]
- Ye, X.; Chen, W.; Huang, X.F.; Yan, F.J.; Deng, S.G.; Zheng, X.D.; Shan, P.F. Anti-diabetic effect of anthocyanin cyanidin-3-O-glucoside: Data from insulin resistant hepatocyte and diabetic mouse. Nutri. Diabetes 2024, 14, 7. [Google Scholar] [CrossRef]
- Jia, Y.; Wu, C.; Rivera-Piza, A.; Kim, Y.J.; Lee, J.H.; Lee, S.J. Mechanism of action of cyanidin 3-O-glucoside in gluconeogenesis and oxidative stress-induced cancer cell senescence. Antioxidants 2022, 11, 749. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.Y.; Wu, S.J.; Chang, Y.C.; Ng, L.T.; Chang, S.J. Stimulation of GLUT4 glucose uptake by anthocyanin-rich extract from black rice (Oryza sativa L.) via PI3K/Akt and AMPK/p38 MAPK signaling in C2C12 cells. Metabolites 2022, 12, 856. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Balandrano, D.D.; Chai, Z.; Hutabarat, R.P.; Beta, T.; Feng, J.; Ma, K.; Li, D.; Huang, W. Hypoglycemic and hypolipidemic effects of blueberry anthocyanins by AMPK activation: In vitro and in vivo studies. Redox Biol. 2021, 46, 102100. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, L.; Fang, Z.; Nisar, T.; Zou, L.; Li, D.; Guo, Y. Lycium ruthenicum Murray anthocyanins effectively inhibit α-glucosidase activity and alleviate insulin resistance. Food Biosci. 2021, 41, 100949. [Google Scholar] [CrossRef]
- Li, R.; Ye, Z.; Xu, Y.J.; Liu, Y. Cyanidin-3-O-glucoside from blueberry anthocyanin extracts protects ARPE-19 cells against high glucose damage via REDD1/GSK3β pathway. Food Biosci. 2023, 56, 103322. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, W.; Xu, X. Cyanidin-3-glucoside suppresses the progression of lung adenocarcinoma by downregulating TP53I3 and inhibiting PI3K/AKT/mTOR pathway. World J. Surg. Oncol. 2021, 19, 232. [Google Scholar] [CrossRef]
- Silveira Rabelo, A.C.; Mertens-Talcott, S.U.; Chew, B.P.; Noratto, G. Dark sweet cherry (Prunus avium) anthocyanins suppressed ERK1/2-Akt/mTOR cell signaling and oxidative stress: Implications for TNBC growth and invasion. Molecules 2022, 27, 7245. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, J.; Meng, X. Aronia melanocarpa anthocyanin extracts are an effective regulator of suppressor of cytokine signaling 3-dependent insulin resistance in HepG2 and C2C12 cells. J. Funct. Foods 2020, 75, 104258. [Google Scholar] [CrossRef]
- Shi, M.; Mathai, M.L.; Xu, G.; Su, X.Q.; McAinch, A.J. The effect of dietary supplementation with blueberry, cyanidin-3-O-β-glucoside, yoghurt and its peptides on gene expression associated with glucose metabolism in skeletal muscle obtained from a high-fat-high-carbohydrate diet induced obesity model. PLoS ONE 2022, 17, e0270306. [Google Scholar] [CrossRef]
- Teixeira, L.D.L.; Pilon, G.; Coutinho, C.P.; Dudonné, S.; Dube, P.; Houde, V.; Hassimotto, N.M. Purple grumixama anthocyanins (Eugenia brasiliensis Lam.) attenuate obesity and insulin resistance in high-fat diet mice. Food Funct. 2021, 12, 3680–3691. [Google Scholar] [CrossRef]
- Jiang, T.; Shuai, X.; Li, J.; Yang, N.; Deng, L.; Li, S.; He, Y.; Guo, H.; Li, Y.; He, J. Protein-bound anthocyanin compounds of purple sweet potato ameliorate hyperglycemia by regulating hepatic glucose metabolism in high-fat diet/streptozotocin-induced diabetic mice. J. Agric. Food Chem. 2020, 68, 1596–1608. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Bao, T.; Xie, L.; Xu, Y.; Chen, W. Transcriptome profiling reveals the antihyperglycemic mechanism of pelargonidin-3-O-glucoside extracted from wild raspberry. J. Funct. Foods 2020, 64, 103657. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Chu, Q.; Zheng, X. Red raspberry (poly) phenolic extract improves diet-induced obesity, hepatic steatosis and insulin resistance in obese mice. J. Berry Res. 2021, 11, 349–362. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Maleki, M.; Sathyapalan, T.; Jamialahmadi, T.; Sahebkar, A. Obesity and insulin resistance: A review of molecular interactions. Curr. Mol. Med. 2021, 21, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Flores-Opazo, M.; McGee, S.L.; Hargreaves, M. Exercise and GLUT4. Exerc. Sport Sci. Rev. 2020, 48, 110–118. [Google Scholar] [CrossRef]
- Chen, D.; Chen, X.; He, C.; Chen, Z.; Chen, Q.; Chen, J.; Bo, H. Sanhuang xiexin decoction synergizes insulin/PI3K-Akt/FoxO signaling pathway to inhibit hepatic glucose production and alleviate T2DM. J. Ethnopharmacol. 2023, 306, 116162. [Google Scholar] [CrossRef]
- Cho, C.H.; Patel, S.; Rajbhandari, P. Adipose tissue lipid metabolism: Lipolysis. Curr. Opin. Genet. Dev. 2023, 83, 102114. [Google Scholar] [CrossRef]
- Deng, H.; Chen, Y.; Li, P.; Hang, Q.; Zhang, P.; Jin, Y.; Chen, M. PI3K/AKT/mTOR pathway, hypoxia, and glucose metabolism: Potential targets to overcome radioresistance in small cell lung cancer. Cancer Pathog. Ther. 2023, 1, 56–66. [Google Scholar] [CrossRef]
- Cao, Y.; Han, S.; Lu, H.; Luo, Y.; Guo, T.; Wu, Q.; Luo, F. Targeting mTOR signaling by dietary polyphenols in obesity prevention. Nutrients 2022, 14, 5171. [Google Scholar] [CrossRef]
- Grabner, G.F.; Xie, H.; Schweiger, M.; Zechner, R. Lipolysis: Cellular mechanisms for lipid mobilization from fat stores. Nat. Metab. 2021, 3, 1445–1465. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C. The role of mTORC1 in the regulation of skeletal muscle mass. Fac. Rev. 2022, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Marcondes-de-Castro, I.A.; Reis-Barbosa, P.H.; Marinho, T.S.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. AMPK/mTOR pathway significance in healthy liver and non-alcoholic fatty liver disease and its progression. J. Gastroenterol. Hepatol. 2023, 38, 1868–1876. [Google Scholar] [CrossRef] [PubMed]
- Asahara, S.I.; Inoue, H.; Watanabe, H.; Kido, Y. Roles of mTOR in the regulation of pancreatic β-cell mass and insulin secretion. Biomolecules 2022, 12, 614. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhou, X.; Li, D.L.; Ye, J.M. Role of the mTOR-autophagy-ER stress pathway in high fructose-induced metabolic-associated fatty liver disease. Acta Pharmacol. Sin. 2022, 43, 10–14. [Google Scholar] [CrossRef]
- Sukumaran, A.; Choi, K.; Dasgupta, B. Insight on transcriptional regulation of the energy sensing AMPK and biosynthetic mTOR pathway genes. Front. Cell Dev. Biol. 2020, 8, 671. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, L.; Yan, Y.; Zhao, L.; Han, S.; Wu, D.; Ji, X. cPKCγ-modulated autophagy contributes to ischemic preconditioning–induced neuroprotection in mice with ischemic stroke via mTOR-ULK1 pathway. Transl. Stroke Res. 2023, 14, 790–801. [Google Scholar] [CrossRef]
- Hussain, I.; Sureshkumar, H.K.; Bauer, M.; Rubio, I. Starvation protects hepatocytes from inflammatory damage through Paradoxical mTORC1 signaling. Cells 2023, 12, 1668. [Google Scholar] [CrossRef]
- Lee, M.G.; Hong, H.J.; Nam, K.S. Anthocyanin oligomers induce apoptosis and autophagy by inhibiting the mTOR signaling pathway in human breast cancer cells. Pharmaceuticals 2023, 17, 24. [Google Scholar] [CrossRef]
- Liu, K.; Yang, Y.; Zhou, F.; Xiao, Y.; Shi, L. Inhibition of PI3K/AKT/mTOR signaling pathway promotes autophagy and relieves hyperalgesia in diabetic rats. Neuroreport 2020, 31, 644–649. [Google Scholar] [CrossRef]
- El-Ashmawy, N.E.; Khedr, E.G.; Alfeky, N.H.; Ibrahim, A.O. Upregulation of GLUT4 and PI3K, and downregulation of GSK3 mediate the anti-hyperglycemic effects of proanthocyanidins. Med. Int. 2022, 2, 14. [Google Scholar] [CrossRef]
- Yan, Z.; Cao, X.; Sun, S.; Sun, B.; Gao, J. Inhibition of GSK3B phosphorylation improves glucose and lipid metabolism disorder. Biochim. Biophys. Acta 2023, 1869, 166726. [Google Scholar] [CrossRef]
- Delangre, E.; Liu, J.; Tolu, S.; Maouche, K.; Armanet, M.; Cattan, P.; Movassat, J. Underlying mechanisms of glucocorticoid-induced β-cell death and dysfunction: A new role for glycogen synthase kinase 3. Cell Death Dis. 2021, 12, 1136. [Google Scholar] [CrossRef]
- Fontana, F.; Giannitti, G.; Marchesi, S.; Limonta, P. The PI3K/Akt pathway and glucose metabolism: A dangerous liaison in cancer. Int. J. Biol. Sci. 2024, 20, 3113. [Google Scholar] [CrossRef]
- Canbolat, E.; Cakıroglu, F.P. The importance of AMPK in obesity and chronic diseases and the relationship of AMPK with nutrition: A literature review. Crit. Rev. Food Sci. Nutr. 2023, 63, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Yaman, M.; Kismiroglu, C.; Ugur, H.; Belli, I.; Ozgur, B. Biochemical development of insulin resistance due to excess nutrition and the function of AMP-activated protein kinase (AMPK). Eur. J. Sci. Technol. 2020, 20, 67–76. [Google Scholar] [CrossRef]
- Tanyıldız, S.N.; Yıldırım, H.; Ugur, H.; Yaman, M. AMPK’s natural activators and relationships with diseases. Eur. J. Sci. Technol. 2021, 21, 389–401. [Google Scholar] [CrossRef]
- Kismiroğlu, C.; Cengiz, S.; Yaman, M. Biochemistry of AMPK: Mechanisms of action and importance in the treatment of diabetes. Eur. J. Sci. Technol. 2020, 18, 162–170. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Maroyi, A. The therapeutic potential of anthocyanins: Current approaches based on their molecular mechanism of action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef]
- Palabiyik, A.A.; Palabiyik, E. Pharmacological approaches to enhance mitochondrial biogenesis: Focus on PGC-1A, AMPK, and SIRT1 in cellular health. Mol. Biol. Rep. 2025, 52, 270. [Google Scholar] [CrossRef]
- Fang, C.; Pan, J.; Qu, N.; Lei, Y.; Han, J.; Zhang, J.; Han, D. The AMPK pathway in fatty liver disease. Front. Physiol. 2022, 13, 970292. [Google Scholar] [CrossRef]
- Xu, H.; Liu, M.; Liu, H.; Zhao, B.; Zheng, M.; Liu, J. Anthocyanins from purple corn ameliorated obesity in high fat diet-induced obese mice through activating hepatic AMPK. J. Funct. Foods 2021, 84, 104582. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Luzardo-Ocampo, I.; Cuellar-Nuñez, M.L.; Loarca-Piña, G.; de Mejia, E.G. Maize extract rich in ferulic acid and anthocyanins prevents high-fat-induced obesity in mice by modulating SIRT1, AMPK and IL-6 associated metabolic and inflammatory pathways. J. Nutr. Biochem. 2020, 79, 108343. [Google Scholar] [CrossRef]
- Panic, A.; Stanimirovic, J.; Sudar-Milovanovic, E.; Isenovic, E.R. Oxidative stress in obesity and insulin resistance. Explorat. Med. 2022, 3, 58–70. [Google Scholar] [CrossRef]
- Taşkın, S.; Çelik, H.; Taşkın, A. The effect of anthocyanin administration on redox balance in acute exercise: An experimental study. Cukurova Med. J. 2024, 49, 471–478. [Google Scholar] [CrossRef]
- Li, Z.; Tian, J.; Cheng, Z.; Teng, W.; Zhang, W.; Bao, Y.; Li, B. Hypoglycemic bioactivity of anthocyanins: A review on proposed targets and potential signaling pathways. Crit. Rev. Food Sci. Nutr. 2023, 63, 7878–7895. [Google Scholar] [CrossRef]
- Fang, Z.; Luo, Y.; Ma, C.; Dong, L.; Chen, F. Blueberry anthocyanins extract attenuates acrylamide-induced oxidative stress and neuroinflammation in rats. Oxid. Med. Cell. Longev. 2022, 2022, 7340881. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, A.; Dzierżanowski, T. Targeting inflammation by anthocyanins as the novel therapeutic potential for chronic diseases: An update. Molecules 2021, 26, 4380. [Google Scholar] [CrossRef]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A chronic low-grade inflammation and its markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef]
- Vugic, L.; Colson, N.; Nikbakht, E.; Gaiz, A.; Holland, O.J.; Kundur, A.R.; Singh, I. Anthocyanin supplementation inhibits secretion of pro-inflammatory cytokines in overweight and obese individuals. J. Funct. Foods 2020, 64, 103596. [Google Scholar] [CrossRef]
- He, X.; Li, Y.; Deng, B.; Lin, A.; Zhang, G.; Ma, M.; Kang, X. The PI3K/AKT signalling pathway in inflammation, cell death and glial scar formation after traumatic spinal cord injury: Mechanisms and therapeutic opportunities. Cell Prolif. 2022, 55, e13275. [Google Scholar] [CrossRef] [PubMed]
- Santamarina, A.B.; Calder, P.C.; Estadella, D.; Pisani, L.P. Anthocyanins ameliorate obesity-associated metainflammation: Preclinical and clinical evidence. Nutr. Res. 2023, 114, 50–70. [Google Scholar] [CrossRef] [PubMed]
- Aranha, L.N.; Silva, M.G.; Uehara, S.K.; Luiz, R.R.; Nogueira Neto, J.F.; Rosa, G.; Moraes de Oliveira, G.M. Effects of a hypoenergetic diet associated with açaí (Euterpe oleracea Mart.) pulp consumption on antioxidant status, oxidative stress and inflammatory biomarkers in overweight, dyslipidemic individuals. Clin. Nutr. 2020, 39, 1464–1469. [Google Scholar] [CrossRef]
- Basu, A.; Izuora, K.; Betts, N.M.; Kinney, J.W.; Salazar, A.M.; Ebersole, J.L.; Scofield, R.H. Dietary strawberries improve cardiometabolic risks in adults with obesity and elevated serum LDL cholesterol in a randomized controlled crossover trial. Nutrients 2021, 13, 1421. [Google Scholar] [CrossRef]
- Habanova, M.; Bihari, M.; Latal, R.; Gažarová, M.; Lenártová, P.; Pastrnakova, J.; Hamulka, J. Short-term supplementation with 100% bilberry products and its effects on body composition and lipid profile in overweight/obese women. Preprints 2025, 15, 218. [Google Scholar] [CrossRef]
- Jung, A.J.; Sharma, A.; Lee, S.H.; Lee, S.J.; Kim, J.H.; Lee, H.J. Efficacy of black rice extract on obesity in obese postmenopausal women: A 12-week randomized, double-blind, placebo-controlled preliminary clinical trial. Menopause 2021, 28, 1391–1399. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, Z.; Zhao, H.; Wang, X.; Pang, J.; Li, Q.; Yang, Y.; Ling, W. Anthocyanin supplementation improves anti-oxidative and anti-inflammatory capacity in a dose-response manner in subjects with dyslipidemia. Redox Biol. 2020, 32, 101474. [Google Scholar] [CrossRef] [PubMed]
- Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; Cesar, H.D.C.; Vasconcelos, J.R.; Oyama, L.M.; Pisani, L.P. Obesity-related inflammatory modulation by juçara berry (Euterpe edulis Mart.) supplementation in Brazilian adults: A double-blind randomized controlled trial. Eur. J. Nutr. 2020, 59, 1693–1705. [Google Scholar] [CrossRef]
- Xu, Z.; Xie, J.; Zhang, H.; Pang, J.; Li, Q.; Wang, X.; Ling, W. Anthocyanin supplementation at different doses improves cholesterol efflux capacity in subjects with dyslipidemia—A randomized controlled trial. Eur. J. Clin. Nutr. 2021, 75, 345–354. [Google Scholar] [CrossRef]
- Richter, C.K.; Skulas-Ray, A.C.; Gaugler, T.L.; Meily, S.; Petersen, K.S.; Kris-Etherton, P.M. Randomized double-blind controlled trial of freeze-dried strawberry powder supplementation in adults with overweight or obesity and elevated cholesterol. J. Am. Nutr. Assoc. 2023, 42, 148–158. [Google Scholar] [CrossRef]
- Vlaicu, P.A.; Untea, A.E.; Varzaru, I.; Saracila, M.; Oancea, A.G. Designing nutrition for health—Incorporating dietary by-products into poultry feeds to create functional foods with insights into health benefits, risks, bioactive compounds, food component functionality and safety regulations. Foods 2023, 12, 4001. [Google Scholar] [CrossRef]
- Danielewski, M.; Matuszewska, A.; Szeląg, A.; Sozański, T. The impact of anthocyanins and iridoids on transcription factors crucial for lipid and cholesterol homeostasis. Int. J. Mol. Sci. 2021, 22, 6074. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Cianciosi, D.; Alvarez-Suarez, J.M.; Quiles, J.L.; Forbes-Hernández, T.Y.; Navarro-Hortal, M.D.; Diaz, Y.A. Anthocyanins: What do we know until now? J. Berry Res. 2023, 13, 1–6. [Google Scholar] [CrossRef]
- Nikbakht, E.; Singh, I.; Vider, J.; Williams, L.T.; Vugic, L.; Gaiz, A.; Colson, N. Potential of anthocyanin as an anti-inflammatory agent: A human clinical trial on type 2 diabetic, diabetic at-risk and healthy adults. Inflamm. Res. 2021, 70, 275–284. [Google Scholar] [CrossRef]
- Ma, Y.; Feng, Y.; Diao, T.; Zeng, W.; Zuo, Y. Experimental and theoretical study on antioxidant activity of the four anthocyanins. J. Mol. Struct. 2020, 1204, 127509. [Google Scholar] [CrossRef]
- Echegaray, N.; Munekata, P.E.; Gullón, P.; Dzuvor, C.K.; Gullón, B.; Kubi, F.; Lorenzo, J.M. Recent advances in food products fortification with anthocyanins. Crit. Rev. Food Sci. Nutr. 2022, 62, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Palencia-Argel, M.; Rodríguez-Villamil, H.; Bernal-Castro, C.; Díaz-Moreno, C.; Fuenmayor, C.A. Probiotics in anthocyanin-rich fruit beverages: Research and development for novel synbiotic products. Crit. Rev. Food Sci. Nutr. 2024, 64, 110–126. [Google Scholar] [CrossRef]
- Boyle, K.A.; Verghese, M. Development of a functional food snack product utilizing underexplored and underutilized ingredients: Spirulina microalgae and bilberry. Food Nutr. Sci. 2024, 15, 974–990. [Google Scholar] [CrossRef]
- Çetin-Babaoğlu, H.; Yalım, N.; Kale, E.; Tontul, S.A. Pigmented whole maize grains for functional value added and low glycemic index snack production. Food Biosci. 2021, 44, 101349. [Google Scholar] [CrossRef]
- Neder-Suárez, D.; Lardizabal-Gutiérrez, D.; Zazueta-Morales, J.D.J.; Meléndez-Pizarro, C.O.; Delgado-Nieblas, C.I.; Ramírez Wong, B.; Quintero-Ramos, A. Anthocyanins and functional compounds change in a third-generation snacks prepared using extruded blue maize, black bean, and chard: An optimization. Antioxidants 2021, 10, 1368. [Google Scholar] [CrossRef]
- Neacsu, M.; Christie, J.S.; Duncan, G.J.; Vaughan, N.J.; Russell, W.R. Buckwheat, fava bean and hemp flours fortified with anthocyanins and other bioactive phytochemicals as sustainable ingredients for functional food development. Nutraceuticals 2022, 2, 150–161. [Google Scholar] [CrossRef]
- Singh, S.; Nwabor, O.F.; Syukri, D.M.; Voravuthikunchai, S.P. Chitosan-poly (vinyl alcohol) intelligent films fortified with anthocyanins isolated from Clitoria ternatea and Carissa carandas for monitoring beverage freshness. Int. J. Biol. Macromol. 2021, 182, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Di Stefano, V.; Lauria, A.; Pitonzo, R.; Gentile, C. Vaccinium macrocarpon (Cranberry)-based dietary supplements: Variation in mass uniformity, proanthocyanidin dosage and anthocyanin profile demonstrates quality control standard needed. Nutrients 2020, 12, 992. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. The influence of supplementation of anthocyanins on obesity-associated comorbidities: A concise review. Foods 2020, 9, 687. [Google Scholar] [CrossRef]
- Yang, B. Microencapsulation of Polyphenols Extracted from Pomegranate Peels and Their Application in Salad Dressing. Master’s Thesis, University of Georgia, Athens, GA, USA, 2021. [Google Scholar]
- Zhao, R.; Chen, J.; Yu, S.; Niu, R.; Yang, Z.; Wang, H.; Wang, W. Active chitosan/gum Arabic-based emulsion films reinforced with thyme oil encapsulating blood orange anthocyanins: Improving multi-functionality. Food Hydrocoll. 2023, 134, 108094. [Google Scholar] [CrossRef]
- Tian, J.L.; Si, X.; Shu, C.; Wang, Y.H.; Tan, H.; Zang, Z.H.; Li, B. Synergistic effects of combined anthocyanin and metformin treatment for hyperglycemia in vitro and in vivo. J. Agric. Food Chem. 2022, 70, 1182–1195. [Google Scholar] [CrossRef]
- Núñez, S.; Millán-Laleona, A.; Cano-Lou, J.; Corella, A.; Moliner, C.; Cásedas, G.; Gómez-Rincón, C. Black mulberry (Morus nigra L.) prevents deleterious effects of excess glucose in obese C. elegans decreasing lipofuscin accumulation and ROS production. Heliyon 2025, 11, e41898. [Google Scholar] [CrossRef]
- Koh, J.; Xu, Z.; Wicker, L. Binding kinetics of blueberry pectin-anthocyanins and stabilization by non-covalent interactions. Food Hydrocoll. 2020, 99, 105354. [Google Scholar] [CrossRef]
- Liao, M.; Ma, L.; Miao, S.; Hu, X.; Liao, X.; Chen, F.; Ji, J. The in-vitro digestion behaviors of milk proteins acting as wall materials in spray-dried microparticles: Effects on the release of loaded blueberry anthocyanins. Food Hydrocoll. 2021, 115, 106620. [Google Scholar] [CrossRef]
- Yang, W.; Guo, Y.; Liu, M.; Chen, X.; Xiao, X.; Wang, S.; Chen, F. Structure and function of blueberry anthocyanins: A review of recent advances. J. Funct. Foods 2022, 88, 104864. [Google Scholar] [CrossRef]
- Enaru, B.; Drețcanu, G.; Pop, T.D.; Stǎnilǎ, A.; Diaconeasa, Z. Anthocyanins: Factors affecting their stability and degradation. Antioxidants 2021, 10, 1967. [Google Scholar] [CrossRef] [PubMed]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the encapsulation in bioavailability of phenolic compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wang, L.; Liao, J.; Sun, P. Progress on anthocyanidin nano-carriers. Sci. Technol. Food Ind. 2021, 42, 330–336. [Google Scholar] [CrossRef]
- Fu, Z.; Ju, H.; Xu, G.S.; Wu, Y.C.; Chen, X.; Li, H.J. Recent development of carrier materials in anthocyanins encapsulation applications: A comprehensive literature review. Food Chem. 2024, 439, 138104. [Google Scholar] [CrossRef]
- Fang, J.; Luo, Y.; Yuan, K.; Guo, Y.; Jin, S. Preparation and evaluation of an encapsulated anthocyanin complex for enhancing the stability of anthocyanins. Food Sci. Technol. 2020, 117, 108543. [Google Scholar] [CrossRef]
- Ma, Y.; Li, S.; Ji, T.; Wu, W.; Sameen, D.E.; Ahmed, S.; Qin, W.; Dai, J.; Liu, Y. Development and optimization of dynamic gelatin/chitosan nanoparticles incorporated with blueberry anthocyanins for milk freshness monitoring. Carbohydr. Polym. 2020, 247, 116738. [Google Scholar] [CrossRef]
- Xie, C.; Wang, Q.; Ying, R.; Wang, Y.; Wang, Z.; Huang, M. Binding a chondroitin sulfate-based nanocomplex with kappa-carrageenan to enhance the stability of anthocyanins. Food Hydrocoll. 2020, 100, 105448. [Google Scholar] [CrossRef]
- Lang, Y.; Li, B.; Gong, E.; Shu, C.; Si, X.; Gao, N.; Meng, X. Effects of α-casein and β-casein on the stability, antioxidant activity and bioaccessibility of blueberry anthocyanins with an in vitro simulated digestion. Food Chem. 2021, 334, 127526. [Google Scholar] [CrossRef]
- Lang, Y.; Tian, J.; Meng, X.; Si, X.; Tan, H.; Wang, Y.; Shu, C.; Chen, Y.; Zang, Z.; Zhang, Y.; et al. Effects of α-Casein on the Absorption of Blueberry Anthocyanins and Metabolites in Rat Plasma Based on Pharmacokinetic Analysis. J. Agric. Food Chem. 2021, 69, 6200–6213. [Google Scholar] [CrossRef]
- Zang, Z.; Chou, S.; Tian, J.; Lang, Y.; Shen, Y.; Ran, X.; Gao, N.; Li, B. Effect of whey protein isolate on the stability and antioxidant capacity of blueberry anthocyanins: A mechanistic and in vitro simulation study. Food Chem. 2021, 336, 127700. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, metabolism and bioavailability of flavonoids: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef] [PubMed]
- Gui, H.; Sun, L.; Liu, R.; Si, X.; Li, D.; Wang, Y.; Tian, J. Current knowledge of anthocyanin metabolism in the digestive tract: Absorption, distribution, degradation, and interconversion. Crit. Rev. Food Sci. Nutr. 2023, 63, 5953–5966. [Google Scholar] [CrossRef]
- Liang, A.; Leonard, W.; Beasley, J.T.; Fang, Z.; Zhang, P.; Ranadheera, C.S. Anthocyanins-gut microbiota-health axis: A review. Crit. Rev. Food Sci. Nutr. 2024, 64, 7563–7588. [Google Scholar] [CrossRef]
- Chen, K.; Kortesniemi, M.K.; Linderborg, K.M.; Yang, B. Anthocyanins as promising molecules affecting energy homeostasis, inflammation, and gut microbiota in type 2 diabetes with special reference to impact of acylation. J. Agric. Food Chem. 2023, 71, 1002–1017. [Google Scholar] [CrossRef] [PubMed]
- Ponder, A.; Hallmann, E.; Kwolek, M.; Średnicka-Tober, D.; Kazimierczak, R. Genetic differentiation in anthocyanin content among berry fruits. Curr. Issues Mol. Biol. 2021, 43, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Simkova, K.; Veberic, R.; Hudina, M.; Grohar, M.C.; Ivancic, T.; Smrke, T.; Jakopic, J. Variability in ‘Capri’everbearing strawberry quality during a harvest season. Foods 2023, 12, 1349. [Google Scholar] [CrossRef]
- Chen, S.; Jia, Y.; Wu, Y.; Ren, F. Anthocyanin and its bioavailability, health benefits, and applications: A comprehensive review. Food Rev. Int. 2024, 40, 3666–3689. [Google Scholar]
- Rosales, T.K.O.; Fabi, J.P. Nanoencapsulated anthocyanin as a functional ingredient: Technological application and future perspectives. Colloids Surf. B. 2022, 218, 112707. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Wang, X.; Lu, B.; Zhang, J. Preparation of blueberry anthocyanin liposomes and changes of vesicle properties, physicochemical properties, in vitro release, and antioxidant activity before and after chitosan modification. Food Sci. Nutr. 2022, 10, 75–87. [Google Scholar] [CrossRef]
- Tarone, A.G.; Cazarin, C.B.B.; Junior, M.R.M. Anthocyanins: New techniques and challenges in microencapsulation. Food Res. Int. 2020, 133, 109092. [Google Scholar] [CrossRef]
- Biondi, F.; Balducci, F.; Capocasa, F.; Visciglio, M.; Mei, E.; Vagnoni, M.; Mazzoni, L. Environmental conditions and agronomical factors influencing the levels of phytochemicals in Brassica vegetables responsible for nutritional and sensorial properties. Appl. Sci. 2021, 11, 1927. [Google Scholar] [CrossRef]






| Compound | Source | Study Model | Dose | Effects | Reference |
|---|---|---|---|---|---|
| C3G | SGBS cells 3T3-L1 cells | 10–20 μm | ↑Expression of GLUT-1 and GLUT-4 protein levels ↑IRS-1 phosphorylation Glucose uptake ↓Lipid accumulation, | [15] | |
| Petunidin-3-O-glucoside, delphinidin-3-O-glucoside Malvidin-3-O-galactoside | Blueberry | HepG2 cells | 100 μm | ↑Glucose uptake ↓Lipid accumulation | [17] |
| C3G | Black soybean | 3T3-L1 cells | 12.5 μg/mL | ↑Expression of pparγ protein ↓ROS levels | [34] |
| White sweet potato | C2C12 cells | 12.5–200 µg/mL | ↑Glucose uptake ↑PI3K/Akt signaling pathway activation | [16] | |
| C3G | Red bayberry | HepG2 cells L02 cells | 20, 40, and 80 μg/mL | ↑Glucose consumption ↑Glucose uptake | [35] |
| C3G | Red bayberry | HepG2 cells HEK293 cells | 10 and 50 μm | ↓Transactivation of FOXO1 ↓Phosphorylation of S6K1 (mtorc1 substrate) | [36] |
| Hualien black rice | C2C12 cells | 10, 50, and 100 μg/mL | ↑Expression of GLUT4 level ↑Phosphorylation of IRS-1 ↑Expression levels of P13K, p-Akt/Akt, and p-p38/p38 | [37] | |
| Malvidin | Brightwell rabbiteye blueberries | HepG2 cells | 5 μg/mL | ↓ROS levels ↓Expression levels of FOXO1 ↑Phosphorylation GSK3β at Ser9 ↓Expression levels of SREBP-1c | [38] |
| Petunidin-3-O-glucoside-5-O-galactoside | Lycium ruthenicum Murray | Caco-2 cells HepG2 cells | 25, 50, 100 and 200 μg/mL | ↑Glucose consumption ↑Glucose uptake ↑Protein levels of p-Akt/Akt ↓Activity of the downstream GSK3β and FOXO1 ↑Protein levels of p-IRS2/IRS2 | [39] |
| C3G | Blueberry | ARPE-19 cells | 10 μm | ↓ROS generation ↓Expression level of GSK3β | [40] |
| C3G | Fruits and vegetables | Human H1299 and A549 cells Lung epithelial cell line BEAS-2B | 40 μm | ↓Expression level of p-mtor/ mtor ↓Expression level of p-Akt/Akt | [41] |
| Dark sweet cherry | 4T1 murine BC cells | - | ↓Expression level of p-mtor/mtor ↓ROS levels | [42] | |
| C3G Cyanidin-3-O-galactoside Cyanidin-3-O-arabinoside | Aronia melanocarpa | HepG2 cells C2C12 cells | 40 μg/mL | ↑mRNA and protein levels of GLUT-4 ↑Glucose uptake ↓Expression levels of p-GSK-3β (Ser9) | [43] |
| Compound | Source | Model | Dose | Effects | Reference |
|---|---|---|---|---|---|
| C3G | Blueberry | Male C57BL/6 mice, 6-week-old, n = 10 | Blueberries—6.4 g/kg BW/d C3G—0.02 g/kg BW/d | ↓FoxO1 expression in the EDL ↑GLUT4 expression ↑AMPK and PI3K expression in the EDL skeletal | [44] |
| C3G | Purple grumixama | Male C57BL/6J mice, 7-week old, n = 12 | 200 mg/kg/BW/d | ↑Insulin sensitivity ↓Insulin resistance ↓Triglyceride accumulation in the liver ↓Akt1 and Slc1a2 gene expression | [45] |
| C3G | Red bayberry | Male C57BL/6J mice, 4-week-old, n = 6 | 150 mg/kg/BW/d | ↓Blood glucose levels | [35] |
| cyanidin or peonidin | Purple sweet potatoes | Male Institute of Cancer Research (ICR) mice, 4-week, old, n = 6 | 200 mg/kg/BW/d | ↑Glucose tolerance ↑Hypoglycemic activity ↑Glycolysis ↓Gluconeogenesis in the liver | [46] |
| Pelargonidin-3-O-glucoside | Wild raspberry | The (db/db) mice with C57BL/6J background, 6-week-old, n = 12 | 50 mg/kg/BW and 150 mg/kg/BW | ↑Glucose tolerance ↑Insulin sensitivity ↓Hepatic genes sterol regulatory element-binding transcription factor 1 (Srebp1c) | [47] |
| malvidin | Brightwell rabbit eye blueberries | Male C57BL/6J mice | 100 mg/kg/BW, and 400 mg/kg/BW | ↓Blood glucose levels | [38] |
| - | Red raspberry | Male C57BL/6J mice, n = 12 | 150 mg/kg/BW | ↑Lipolysis ↓Levels of total cholesterol, LDL cholesterol | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randeni, N.; Luo, J.; Xu, B. Critical Review on Anti-Obesity Effects of Anthocyanins Through PI3K/Akt Signaling Pathways. Nutrients 2025, 17, 1126. https://doi.org/10.3390/nu17071126
Randeni N, Luo J, Xu B. Critical Review on Anti-Obesity Effects of Anthocyanins Through PI3K/Akt Signaling Pathways. Nutrients. 2025; 17(7):1126. https://doi.org/10.3390/nu17071126
Chicago/Turabian StyleRandeni, Nidesha, Jinhai Luo, and Baojun Xu. 2025. "Critical Review on Anti-Obesity Effects of Anthocyanins Through PI3K/Akt Signaling Pathways" Nutrients 17, no. 7: 1126. https://doi.org/10.3390/nu17071126
APA StyleRandeni, N., Luo, J., & Xu, B. (2025). Critical Review on Anti-Obesity Effects of Anthocyanins Through PI3K/Akt Signaling Pathways. Nutrients, 17(7), 1126. https://doi.org/10.3390/nu17071126