

Integrative Roles of Functional Foods, Microbiotics, Nutrigenetics, and Nutrigenomics in Managing Type 2 Diabetes and Obesity
 , , , and
, , , and 
Abstract
1. Introduction
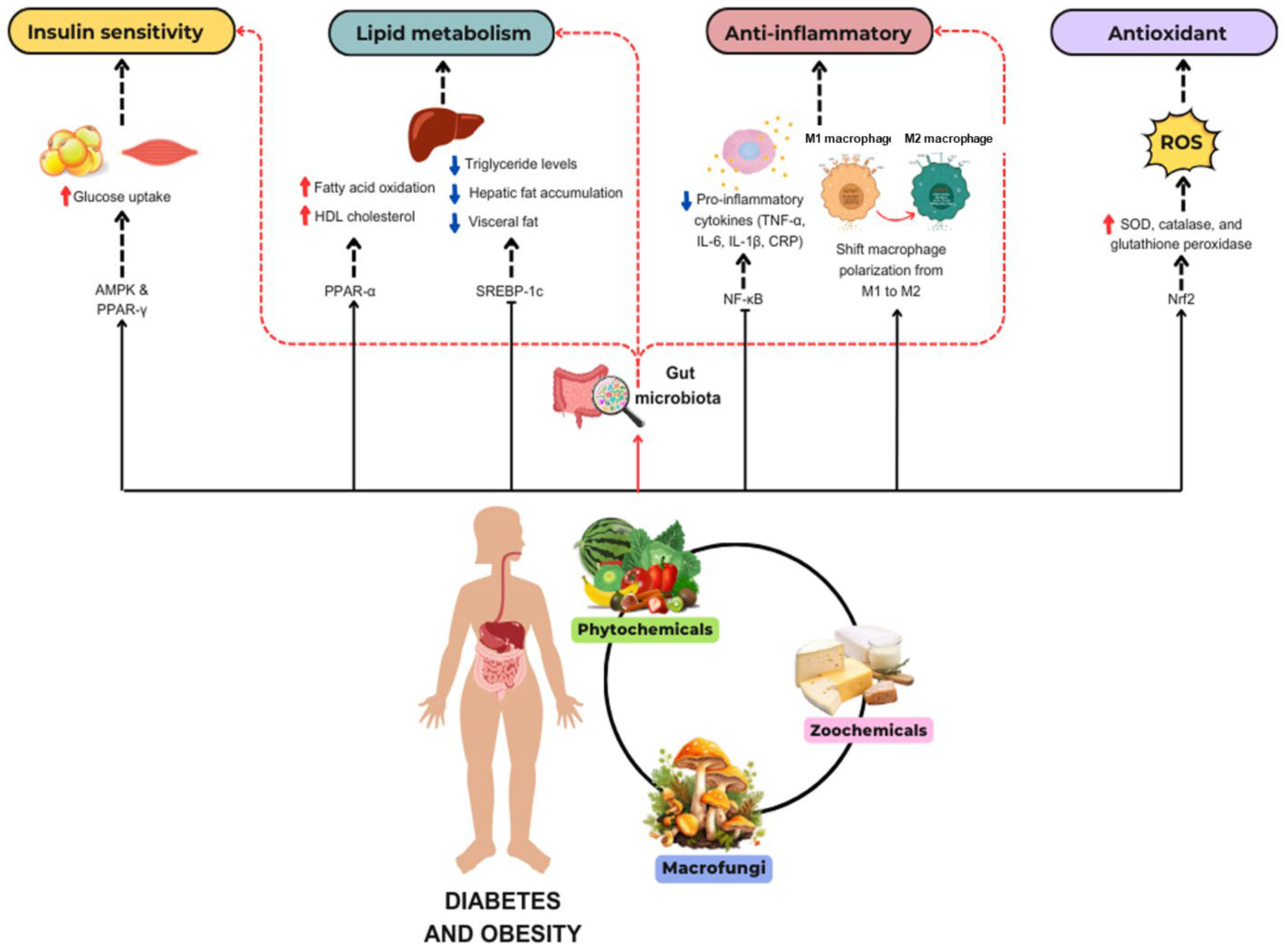
2. Impact of Bioactive Compounds on Metabolic Health in Type 2 Diabetes and Obesity
2.1. Phytochemicals
Mechanism of Action

{kind=link}
{kind=link}
{kind=link}
| Phytochemical | Species | Experiment Model/Dosage | Key Findings | Reference |
|---|---|---|---|---|
Epigallocatechin-3-gallate (EGCG)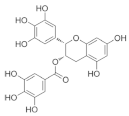 | Green tea (Camellia sinensis) |
|
| [55] ** |
Epigallocatechin gallate (EGCG)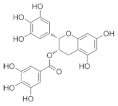 β-Cryptoxanthin 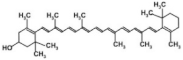 | Catechins from green tea (Camellia sinensis) β-Cryptoxanthin from citrus fruits | Monosodium-glutamate-induced obese male C57BL/6J mice/1.7 mg green tea catechins and 50 µg β-Cryptoxanthin/kg/day. |
| [56] *** |
Epigallocatechin gallate (EGCG) Caffeine  | White tea | Obese human participants (BMI ≥ 30 kg/m2)/consumed 2 cups of white tea daily (brewed from sachets) along with a calorie-restricted diet (1400–1600 kcal/day) and exercise for 12 weeks *. |
| [57] **** |
Curcumin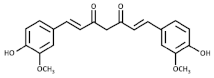 | Turmeric (Curcuma longa) | 3T3-L1 preadipocytes/optimal effects at 10 µM Male C57BL/6J mice with diet induced obesity)/50 mg/kg/day for 8 weeks. |
| [58] ** |
| Male C57BL/6J mice with diet-induced obesity and genetically obese mice/3% dietary curcumin mixed in the diet for 6 weeks *. |
| [59] *** | ||
| Prediabetic human participants (n = 240)/6 capsules per day (1500 mg/day) for 9 months *. |
| [60] **** | ||
| Obese human participants with type 2 diabetes (n = 229)/6 capsules per day (1500 mg/day) for 12 months *. |
| [61] **** | ||
Dihydro-Resveratrol (DR2)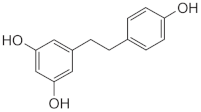 | Dendrobium spp., Dioscorea spp., Bulbophyllum spp. | 3T3-L1 cells and insulin-resistant HepG2 and C2C12 cells/DR2 (10, 20, 40 µM) for 48 h **. High-fat diet (HFD)-induced obese C57BL/6J mice/DR2 (40 or 80 mg/kg/day) for 3 weeks *. |
| [33] ** |
Resveratrol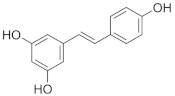 | Grapes (Vitis vinifera), berries, peanuts (Arachis hypogaea). | Patients with type 2 diabetes (n = 110)/200 mg/day (99% pure trans-resveratrol) for 24 weeks. |
| [62] **** |
| Grapes (Vitis vinifera), peanuts (Arachis hypogaea), red wine. | Obese but otherwise healthy male volunteers (n = 11)/150 mg/day (99% pure trans-resveratrol, resVida™) for 30 days. |
| [63] **** | |
Quercetin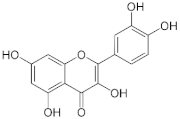 | Apples (Malus domestica), onions (Allium cepa), and berries. | Male C57BL/6J mice were fed a high-fat diet (HF) or HF supplemented with 0.05% quercetin (HFQ) for 6 weeks. |
| [64] *** |
| Patients with type 2 diabetes mellitus (n = 170)/500 mg/day of quercetin dihydrate for 12 weeks. |
| [65,66] **** | ||
β-sitosterol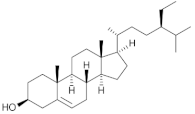 | Chia (Salvia hispanica L.) | Non-polar fractions (light petroleum and dichloromethane) from the aerial parts of Salvia hispanica. |
| [67] * |
n-6 and n-3 PUFA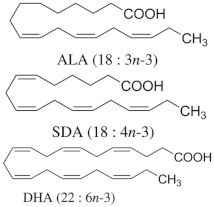 |
| Hypertriglyceridemic adults (n = 59)/ALA group: 20 g/day of linseed oil (7.42 g ALA/day); SDA group: 20 g/day of echium oil (1.57 g SDA/day); DHA group: 12 g/day of microalgae oil (1.64 g DHA/day) for 10 weeks. |
| [68] **** |
2.2. Zoochemicals
Mechanism of Action
| Zoochemical | Experiment Model | Dosage and Duration | Key Findings | Reference |
|---|---|---|---|---|
Cis-9, Trans-11-Conjugated Linoleic Acid (CLA 9,11)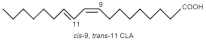 | Bovine Mammary Epithelial Cells (BMECs) | 600 µmol/L H₂O₂ for 8 h |
| [82] * |
| Alzheimer’s disease (AD) mouse model (hAPPSwInd, J20) and C57BL/6 wild-type mice | 0.4% CLA in the diet (~16 mg/day per mouse) from 6 to 14 months of age |
| [83] ** | |
| Cis-9, Trans-11, and Trans-10, Cis-12 Conjugated Linoleic Acid (CLA 9,11 and CLA 10,12)  | EA.hy926 endothelial cells (human umbilical vein endothelial cell lineage) | 1 and 10 µM for CLA9,11 and CLA10,12 for 48 h |
| [84] * |
| Lactating Holstein dairy cows | 120 g/day of CLA supplement providing 12 g/day of each isomer from 21 days pre-calving to 60 days post-calving |
| [85] ** | |
| Overweight and obese adults (n = 68; BMI ≥ 27 and <40 kg/m2) | 60 g/day of PUFA-enriched goat cheese for 12 weeks |
| [72] *** | |
Polyunsaturated Fatty Acids (PUFAs), including n-3 PUFA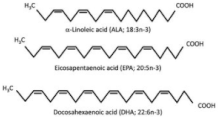 and Conjugated Linoleic Acid (CLA)  | Primary human subcutaneous and visceral adipocytes | 100 μM EPA and/or DHA for 72 h |
| [86] * |
| Omega-3 fatty acids (EPA and DHA) 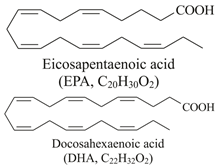 | Fish oil such as: Salmo salar (Atlantic salmon) Clupea harengus (Atlantic herring) Engraulis encrasicolus (European anchovy) Sardinops sagax (Pacific sardine) | Wistar rats on a high-fat diet (HFD)/3.4% fish oil of total dietary energy for 8 weeks. |
| [87] ** |
| Twelve obese women (BMI ≥ 35) and 12 healthy women (BMI < 24)/4.8 g/day (3.2 g EPA + 1.6 g DHA) for 3 months. |
| [88] *** | ||
| 60 diabetic patients with NAFLD/2 g/day (180 mg EPA and 120 mg DHA per capsule, 2 capsules daily) for 12 weeks. |
| [89] *** | ||
Chitosan | Rats fed with a high-sugar and high-fat diet (HSFD) to induce Type 2 diabetes | Wistar rats on a HSHF/Chitosan microsphere (CMS) supplement providing 40 mg/day for 90 days. |
| [76] ** |
Carboxymethyl chitin | 3T3-L1 preadipocytes as a cell model | Carboxymethyl chitin (CM-chitin) was tested in 3T3-L1 adipocytes at concentrations of 50, 100, and 200 μg/mL |
| [78] * |
Chitin-glucan complex | Randomized, double-blind, placebo-controlled clinical trial | Participants received either 1.5 g/day or 4.5 g/day of chitin–glucan for 6 weeks |
| [79] *** |
2.3. Macrofungi
3. The Potential of Probiotics and Gut Microbiota Modulation in the Treatment of Diabetic Kidney Disease and Chronic Kidney Disease
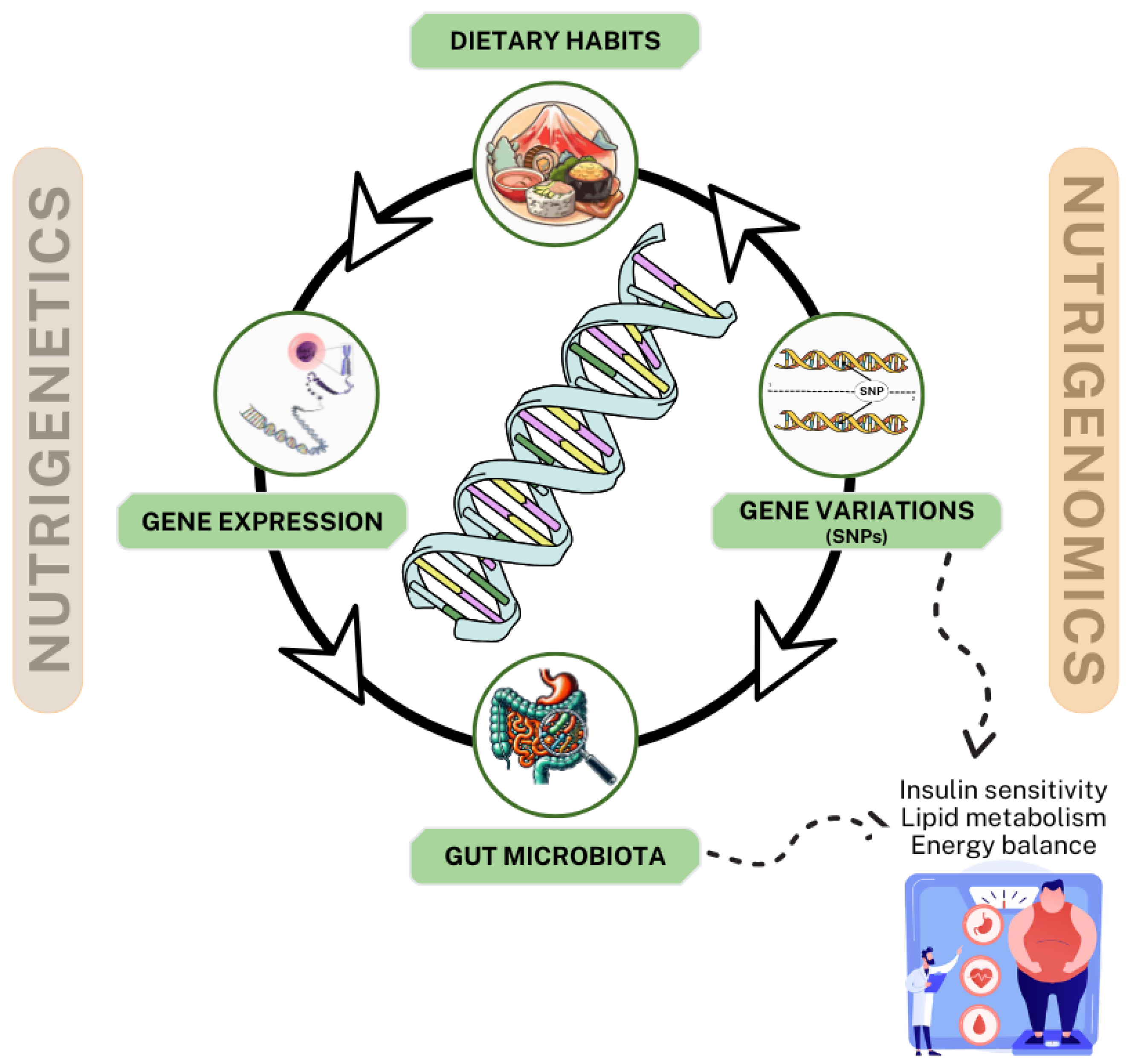
4. Interplay of Nutrigenetics and Nutrigenomics in Personalized Interventions for Obesity and Type 2 Diabetes
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in underweight and obesity from 1990 to 2022: A pooled analysis of 3663 population-representative studies with 222 million children, adolescents, and adults. Lancet 2024, 403, 1027–1050. [Google Scholar] [CrossRef]
- Siqueira, J.S.; Palacio, T.L.N.; Vieira, T.A.; Nakandakare-Maia, E.T.; Grandini, N.A.; Ferron, A.J.T.; Francisqueti-Ferron, F.V.; Correa, C.R. An overview of the complex interaction between obesity and target organ dysfunction: Focus on redox-inflammatory state. Nutrire 2023, 48, 21. [Google Scholar] [CrossRef]
- Magliano, D.J.; Boyko, E.J.; IDF Diabetes Atlas 10th Edition Scientific Committee. IDF Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Romero-Cortes, T.; Lopez-Perez, P.; Toledo, A.; Perez-Espana, V.; Aparicio-Burgos, J.; Cuervo-Parra, J. Nutrigenomics and nutrigenetics in functional foods. Int. J. Bio-Resour. Stress Manag. 2018, 9, 661–672. [Google Scholar] [CrossRef]
- Chen, Y.; Ni, H.; Zhang, H. Exploring the relationship between live microbe intake and obesity prevalence in adults. Sci. Rep. 2024, 14, 21724. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Nutrigenomics and Nutrigenetics in Functional Foods and Personalized Nutrition; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Pan, X.; Olatunji, O.J.; Basit, A.; Sripetthong, S.; Nalinbenjapun, S.; Ovatlarnporn, C. Insights into the phytochemical profiling, antidiabetic and antioxidant potentials of Lepionurus sylvestris Blume extract in fructose/streptozotocin-induced diabetic rats. Front. Pharmacol. 2024, 15, 1424346. [Google Scholar] [CrossRef] [PubMed]
- Bakwo Bassogog, C.B.; Nyobe, C.E.; Sabine, F.Y.; Bruno Dupon, A.A.; Ngui, S.P.; Minka, S.R.; Laure, N.J.; Mune Mune, M.A. Protein hydrolysates of Moringa oleifera seed: Antioxidant and antihyperglycaemic potential as ingredient for the management of type-2 diabetes. Heliyon 2024, 10, e28368. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Gabriel, A.; Tsiani, E. Antidiabetic Properties of Curcumin II: Evidence from In Vivo Studies. Nutrients 2019, 12, 58. [Google Scholar] [CrossRef]
- Nakadate, K.; Kawakami, K.; Yamazaki, N. Synergistic Effect of β-Cryptoxanthin and Epigallocatechin Gallate on Obesity Reduction. Nutrients 2024, 16, 2344. [Google Scholar] [CrossRef]
- Wu, S.; Jia, W.; He, H.; Yin, J.; Xu, H.; He, C.; Zhang, Q.; Peng, Y.; Cheng, R. A New Dietary Fiber Can Enhance Satiety and Reduce Postprandial Blood Glucose in Healthy Adults: A Randomized Cross-Over Trial. Nutrients 2023, 15, 4569. [Google Scholar] [CrossRef] [PubMed]
- Costabile, G.; Della Pepa, G.; Vetrani, C.; Vitaglione, P.; Griffo, E.; Giacco, R.; Vitale, M.; Salamone, D.; Rivellese, A.A.; Annuzzi, G.; et al. An Oily Fish Diet Improves Subclinical Inflammation in People at High Cardiovascular Risk: A Randomized Controlled Study. Molecules 2021, 26, 3369. [Google Scholar] [CrossRef]
- Di Pietro, P.; Lizio, R.; Izzo, C.; Visco, V.; Damato, A.; Venturini, E.; De Lucia, M.; Galasso, G.; Migliarino, S.; Rasile, B.; et al. A Novel Combination of High-Load Omega-3 Lysine Complex (AvailOm®) and Anthocyanins Exerts Beneficial Cardiovascular Effects. Antioxidants 2022, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.C.; Fang, T.J.; Ho, H.H.; Chen, J.F.; Kuo, Y.W.; Huang, Y.Y.; Tsai, S.Y.; Wu, S.F.; Lin, H.C.; Yeh, Y.T. A multi-strain probiotic blend reshaped obesity-related gut dysbiosis and improved lipid metabolism in obese children. Front. Nutr. 2022, 9, 922993. [Google Scholar] [CrossRef] [PubMed]
- Rahayu, E.S.; Mariyatun, M.; Putri Manurung, N.E.; Hasan, P.N.; Therdtatha, P.; Mishima, R.; Komalasari, H.; Mahfuzah, N.A.; Pamungkaningtyas, F.H.; Yoga, W.K.; et al. Effect of probiotic Lactobacillus plantarum Dad-13 powder consumption on the gut microbiota and intestinal health of overweight adults. World J. Gastroenterol. 2021, 27, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Wu, Y.; Gao, D.; Dong, Y.; Pan, Y.; Gu, S. Gut Microbiome and Metabolome Alterations in Overweight or Obese Adult Population after Weight-Loss Bifidobacterium breve BBr60 Intervention: A Randomized Controlled Trial. Int. J. Mol. Sci. 2024, 25, 10871. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Jalali, S.; Mojgani, N.; Sanjabi, M.R.; Saremnezhad, S.; Haghighat, S. Functional properties and safety traits of L. rhamnosus and L. reuteri postbiotic extracts. AMB Express 2024, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Kassaian, N.; Feizi, A.; Aminorroaya, A.; Amini, M. Probiotic and synbiotic supplementation could improve metabolic syndrome in prediabetic adults: A randomized controlled trial. Diabetes Metab. Syndr. 2019, 13, 2991–2996. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Chen, R. Exploring the causal connection: Insights into diabetic nephropathy and gut microbiota from whole-genome sequencing databases. Ren. Fail. 2024, 46, 2385065. [Google Scholar] [CrossRef]
- Adeoluwa, O.A.; Olayinka, J.N.; Adeoluwa, G.O.; Akinluyi, E.T.; Adeniyi, F.R.; Fafure, A.; Nebo, K.; Edem, E.E.; Eduviere, A.T.; Abubakar, B. Quercetin abrogates lipopolysaccharide-induced depressive-like symptoms by inhibiting neuroinflammation via microglial NLRP3/NFκB/iNOS signaling pathway. Behav. Brain Res. 2023, 450, 114503. [Google Scholar] [CrossRef]
- Wang, X.; Guo, Q.; Liu, Z.; Wang, Y.; Cao, C.; Jin, L.; Li, C.; Xiao, J.; Zhao, W. Alterations in the Gut Microbiota Composition in Obesity with and without Type 2 Diabetes: A Pilot Study. Diabetes Metab. Syndr. Obes. 2024, 17, 3965–3974. [Google Scholar] [CrossRef]
- Nuli, R.; Cai, J.; Kadeer, A.; Zhang, Y.; Mohemaiti, P. Integrative Analysis Toward Different Glucose Tolerance-Related Gut Microbiota and Diet. Front. Endocrinol. 2019, 10, 295. [Google Scholar] [CrossRef]
- Li, Q.; Xie, S.; Liu, Y.; Yue, W.; Wang, L.; Liang, Y.; Chen, Y.; Yuan, H.; Yu, J. Gut microbiota profiling reflects the renal dysfunction and psychological distress in patients with diabetic kidney disease. Front. Endocrinol. 2024, 15, 1410295. [Google Scholar] [CrossRef]
- Wu, Y.; Jha, R.; Li, A.; Liu, H.; Zhang, Z.; Zhang, C.; Zhai, Q.; Zhang, J. Probiotics (Lactobacillus plantarum HNU082) Supplementation Relieves Ulcerative Colitis by Affecting Intestinal Barrier Functions, Immunity-Related Gene Expression, Gut Microbiota, and Metabolic Pathways in Mice. Microbiol. Spectr. 2022, 10, e0165122. [Google Scholar] [CrossRef] [PubMed]
- Vähämiko, S.; Laiho, A.; Lund, R.; Isolauri, E.; Salminen, S.; Laitinen, K. The impact of probiotic supplementation during pregnancy on DNA methylation of obesity-related genes in mothers and their children. Eur. J. Nutr. 2019, 58, 367–377. [Google Scholar] [CrossRef]
- Do, R.; Bailey, S.D.; Desbiens, K.; Belisle, A.; Montpetit, A.; Bouchard, C.; Pérusse, L.; Vohl, M.C.; Engert, J.C. Genetic variants of FTO influence adiposity, insulin sensitivity, leptin levels, and resting metabolic rate in the Quebec Family Study. Diabetes 2008, 57, 1147–1150. [Google Scholar] [CrossRef]
- Muntean, C.; Sasaran, M.O.; Crisan, A.; Banescu, C. Effects of PPARG and PPARGC1A gene polymorphisms on obesity markers. Front. Public Health 2022, 10, 962852. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, M.; Zhuang, Y.; Zhao, L.; Qian, Y.; Xu, J.; Fan, J. Genetic associations between gut microbiota and type 2 diabetes mediated by plasma metabolites: A Mendelian randomization study. Front. Endocrinol. 2024, 15, 1430675. [Google Scholar] [CrossRef]
- Sun, K.; Gao, Y.; Wu, H.; Huang, X. The causal relationship between gut microbiota and type 2 diabetes: A two-sample Mendelian randomized study. Front. Public Health 2023, 11, 1255059. [Google Scholar] [CrossRef]
- Sachs, J. Physiological Notes: II. Contributions to the Theory of the Cell. a) Energids and Cells. Biol. Theory 2022, 17, 231–233. [Google Scholar] [CrossRef]
- Li, W.; Zhu, C.; Liu, T.; Zhang, W.; Liu, X.; Li, P.; Zhu, T. Epigallocatechin-3-gallate ameliorates glucolipid metabolism and oxidative stress in type 2 diabetic rats. Diabetes Vasc. Dis. Res. 2020, 17, 1479164120966998. [Google Scholar] [CrossRef]
- Lam, C.S.; Xia, Y.X.; Chen, B.S.; Du, Y.X.; Liu, K.L.; Zhang, H.J. Dihydro-Resveratrol Attenuates Oxidative Stress, Adipogenesis and Insulin Resistance in In Vitro Models and High-Fat Diet-Induced Mouse Model via AMPK Activation. Nutrients 2023, 15, 3006. [Google Scholar] [CrossRef]
- Kumar, K.S.; Lin, C.; Tseng, Y.-H.; Wang, S.-Y. Fruits of Rosa laevigata and its bio-active principal sitostenone facilitate glucose uptake and insulin sensitivity in hepatic cells via AMPK/PPAR-γ activation. Phytomedicine Plus 2021, 1, 100109. [Google Scholar] [CrossRef]
- Yap, K.H.; Yee, G.S.; Candasamy, M.; Tan, S.C.; Md, S.; Abdul Majeed, A.B.; Bhattamisra, S.K. Catalpol Ameliorates Insulin Sensitivity and Mitochondrial Respiration in Skeletal Muscle of Type-2 Diabetic Mice Through Insulin Signaling Pathway and AMPK/SIRT1/PGC-1α/PPAR-γ Activation. Biomolecules 2020, 10, 1360. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, W.; Yu, B.; Liang, H.; Mao, S.; Hu, X.; Feng, Y.; Xu, J.; Chu, L. Quercetin improves cerebral ischemia/reperfusion injury by promoting microglia/macrophages M2 polarization via regulating PI3K/Akt/NF-κB signaling pathway. Biomed. Pharmacother. 2023, 168, 115653. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.C.; Huang, S.Y.; Chan, S.T.; Liao, J.W.; Yeh, S.L. Combination of β-carotene and quercetin against benzo[a]pyrene-induced pro-inflammatory reaction accompanied by the regulation of antioxidant enzyme activity and NF-κB translocation in Mongolian gerbils. Eur. J. Nutr. 2015, 54, 397–406. [Google Scholar] [CrossRef]
- Thadhani, V.M. Resveratrol in management of diabetes and obesity: Clinical applications, bioavailability, and nanotherapy. In Resveratrol—Adding Life to Years, Not Adding Years to Life; IntechOpen: London, UK, 2019; Volume 10, pp. 1–166. [Google Scholar]
- Tsai, C.-F.; Chen, G.-W.; Chen, Y.-C.; Shen, C.-K.; Lu, D.-Y.; Yang, L.-Y.; Chen, J.-H.; Yeh, W.-L. Regulatory Effects of Quercetin on M1/M2 Macrophage Polarization and Oxidative/Antioxidative Balance. Nutrients 2022, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, H.; Moustafa, N.; Ahmed, R.R.; El-Shahawy, A.A.G.; Eldin, Z.E.; Al-Jameel, S.S.; Amin, K.A.; Ahmed, O.M.; Abdul-Hamid, M. Therapeutic effect of oral insulin-chitosan nanobeads pectin-dextrin shell on streptozotocin-diabetic male albino rats. Heliyon 2024, 10, e35636. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xue, Y.; Zhang, Y.; Wang, Q.; Qiu, J.; Zhang, J.; Yang, C.; Zhao, Y.; Zhang, Y. Association between dietary antioxidant capacity and type 2 diabetes mellitus in Chinese adults: A population-based cross-sectional study. Nutr. Metab. 2024, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of NF-κB and Nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Orhan, C.; Sahin, N.; Padigaru, M.; Morde, A.; Lal, M.; Dhavan, N.; Erten, F.; Bilgic, A.A.; Ozercan, I.H.; et al. Lutein/Zeaxanthin Isomers and Quercetagetin Combination Safeguards the Retina from Photo-Oxidative Damage by Modulating Neuroplasticity Markers and the Nrf2 Pathway. Pharmaceuticals 2023, 16, 1543. [Google Scholar] [CrossRef]
- Zhang, X.; Cui, K.; Wang, X.; Tong, Y.; Liu, C.; Zhu, Y.; You, Q.; Jiang, Z.; Guo, X. Novel Hydrogen Sulfide Hybrid Derivatives of Keap1-Nrf2 Protein–Protein Interaction Inhibitor Alleviate Inflammation and Oxidative Stress in Acute Experimental Colitis. Antioxidants 2023, 12, 1062. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.O.M.; Evangelista-Silva, P.H.; Neves, N.N.; Moreno, L.G.; Santos, C.S.; Rocha, K.L.S.; Ottone, V.O.; Batista-da-Silva, B.; Dias-Peixoto, M.F.; Magalhães, F.C.; et al. Caloric restriction-induced weight loss with a high-fat diet does not fully recover visceral adipose tissue inflammation in previously obese C57BL/6 mice. Appl. Physiol. Nutr. Metab. 2020, 45, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Baek, S.Y.; Jang, E.J.; Ku, S.K.; Kim, K.M.; Ki, S.H.; Kim, C.E.; Park, K.I.; Kim, S.C.; Kim, Y.W. Oxyresveratrol ameliorates nonalcoholic fatty liver disease by regulating hepatic lipogenesis and fatty acid oxidation through liver kinase B1 and AMP-activated protein kinase. Chem. Biol. Interact. 2018, 289, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Ortega, M.P.; Almanza-Pérez, J.C.; Sánchez-Muñoz, F.; Hong, E.; Velázquez-Reyes, E.; Romero-Nava, R.; Villafaña-Rauda, S.; Pérez-Ontiveros, A.; Blancas-Flores, G.; Huang, F. Effect of Supplementation with Omega-3 Polyunsaturated Fatty Acids on Metabolic Modulators in Skeletal Muscle of Rats with an Obesogenic High-Fat Diet. Pharmaceuticals 2024, 17, 222. [Google Scholar] [CrossRef]
- Kabirifar, R.; Rezaifar, A.; Binesh, F.; Bamdad, K.; Moradi, A. Curcumin, quercetin and atorvastatin protected against the hepatic fibrosis by activating AMP-activated protein kinase. J. Funct. Foods 2018, 40, 341–348. [Google Scholar] [CrossRef]
- Naiini, M.R.; Shahouzehi, B.; Azizi, S.; Shafiei, B.; Nazari-Robati, M. Trehalose-induced SIRT1/AMPK activation regulates SREBP-1c/PPAR-α to alleviate lipid accumulation in aged liver. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Azuma, N.; Mawatari, T.; Saito, Y.; Tsukamoto, M.; Sampei, M.; Iwama, Y. Effect of Continuous Ingestion of Bifidobacteria and Dietary Fiber on Improvement in Cognitive Function: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2023, 15, 4175. [Google Scholar] [CrossRef]
- Baba, Y.; Saito, Y.; Kadowaki, M.; Azuma, N.; Tsuge, D. Effect of Continuous Ingestion of Bifidobacteria and Inulin on Reducing Body Fat: A Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Comparison Study. Nutrients 2023, 15, 5025. [Google Scholar] [CrossRef]
- Corrêa, R.O.; Castro, P.R.; Fachi, J.L.; Nirello, V.D.; El-Sahhar, S.; Imada, S.; Pereira, G.V.; Pral, L.P.; Araújo, N.V.P.; Fernandes, M.F.; et al. Inulin diet uncovers complex diet-microbiota-immune cell interactions remodeling the gut epithelium. Microbiome 2023, 11, 90. [Google Scholar] [CrossRef]
- Kaewarsar, E.; Chaiyasut, C.; Lailerd, N.; Makhamrueang, N.; Peerajan, S.; Sirilun, S. Optimization of Mixed Inulin, Fructooligosaccharides, and Galactooligosaccharides as Prebiotics for Stimulation of Probiotics Growth and Function. Foods 2023, 12, 1591. [Google Scholar] [CrossRef] [PubMed]
- Ezzatpour, S.; Mondragon Portocarrero, A.D.C.; Cardelle-Cobas, A.; Lamas, A.; López-Santamarina, A.; Miranda, J.M.; Aguilar, H.C. The Human Gut Virome and Its Relationship with Nontransmissible Chronic Diseases. Nutrients 2023, 15, 977. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.W.; Cai, S.; Zhao, T.S.; Li, M.; Tian, Y. Green tea derivative (-)-epigallocatechin-3-gallate (EGCG) confers protection against ionizing radiation-induced intestinal epithelial cell death both in vitro and in vivo. Free Radic. Biol. Med. 2020, 161, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Nakadate, K.; Kawakami, K.; Yamazaki, N. Anti-Obesity and Anti-Inflammatory Synergistic Effects of Green Tea Catechins and Citrus β-Cryptoxanthin Ingestion in Obese Mice. Int. J. Mol. Sci. 2023, 24, 7054. [Google Scholar] [CrossRef]
- Akyildiz, K.; Yilmaz, A.; Avci, U.; Toraman, M.N.; Yazici, Z.A. White Tea Consumption Alleviates Anthropometric and Metabolic Parameters in Obese Patients. Medicina 2024, 60, 1568. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Pan, Y.; Yu, N.; Bai, Y.; Ma, R.; Mo, F.; Zuo, J.; Chen, B.; Jia, Q.; Zhang, D.; et al. Curcumin improves adipocytes browning and mitochondrial function in 3T3-L1 cells and obese rodent model. R. Soc. Open Sci. 2021, 8, 200974. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Leibel, R.; Tortoriello, D.V. Dietary curcumin significantly improves obesity-associated inflammation and diabetes in mouse models of diabesity. Endocrinology 2008, 149, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Luechapudiporn, R.; Phisalaphong, C.; Jirawatnotai, S. Curcumin extract for prevention of type 2 diabetes. Diabetes Care 2012, 35, 2121–2127. [Google Scholar] [CrossRef]
- Yaikwawong, M.; Jansarikit, L.; Jirawatnotai, S.; Chuengsamarn, S. Curcumin extract improves beta cell functions in obese patients with type 2 diabetes: A randomized controlled trial. Nutr. J. 2024, 23, 119. [Google Scholar] [CrossRef] [PubMed]
- Mahjabeen, W.; Khan, D.A.; Mirza, S.A. Role of resveratrol supplementation in regulation of glucose hemostasis, inflammation and oxidative stress in patients with diabetes mellitus type 2: A randomized, placebo-controlled trial. Complement. Ther. Med. 2022, 66, 102819. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Tam, C.C.; Rolston, M.; Alves, P.; Chen, L.; Meng, S.; Hong, H.; Chang, S.K.C.; Yokoyama, W. Quercetin Ameliorates Insulin Resistance and Restores Gut Microbiome in Mice on High-Fat Diets. Antioxidants 2021, 10, 1251. [Google Scholar] [CrossRef]
- Mantadaki, A.E.; Linardakis, M.; Tsakiri, M.; Baliou, S.; Fragkiadaki, P.; Vakonaki, E.; Tzatzarakis, M.N.; Tsatsakis, A.; Symvoulakis, E.K. Benefits of Quercetin on Glycated Hemoglobin, Blood Pressure, PiKo-6 Readings, Night-Time Sleep, Anxiety, and Quality of Life in Patients with Type 2 Diabetes Mellitus: A Randomized Controlled Trial. J. Clin. Med. 2024, 13, 3504. [Google Scholar] [CrossRef]
- Mantadaki, A.E.; Baliou, S.; Linardakis, M.; Vakonaki, E.; Tzatzarakis, M.N.; Tsatsakis, A.; Symvoulakis, E.K. Quercetin Intake and Absolute Telomere Length in Patients with Type 2 Diabetes Mellitus: Novel Findings from a Randomized Controlled Before-and-After Study. Pharmaceuticals 2024, 17, 1136. [Google Scholar] [CrossRef] [PubMed]
- Abdel Ghani, A.E.; Al-Saleem, M.S.M.; Abdel-Mageed, W.M.; AbouZeid, E.M.; Mahmoud, M.Y.; Abdallah, R.H. UPLC-ESI-MS/MS Profiling and Cytotoxic, Antioxidant, Anti-Inflammatory, Antidiabetic, and Antiobesity Activities of the Non-Polar Fractions of Salvia hispanica L. Aerial Parts. Plants 2023, 12, 1062. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, M.; Jahreis, G.; Bothor, K.; Drechsel, C.; Kiehntopf, M.; Blüher, M.; Dawczynski, C. Benefits of foods supplemented with vegetable oils rich in α-linolenic, stearidonic or docosahexaenoic acid in hypertriglyceridemic subjects: A double-blind, randomized, controlled trail. Eur. J. Nutr. 2015, 54, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Vass, N.; Czegledi, L.; Javor, A. Significance of functional foods of animal origin in human health. Sci. Pap. Anim. Sci. Biotechnol. 2008, 41, 263. [Google Scholar]
- Gonzalez-Becerra, K.; Barron-Cabrera, E.; Muñoz-Valle, J.F.; Torres-Castillo, N.; Rivera-Valdes, J.J.; Rodriguez-Echevarria, R.; Martinez-Lopez, E. A Balanced Dietary Ratio of n-6:n-3 Polyunsaturated Fatty Acids Exerts an Effect on Total Fatty Acid Profile in RBCs and Inflammatory Markers in Subjects with Obesity. Healthcare 2023, 11, 2333. [Google Scholar] [CrossRef]
- Spencer, M.; Finlin, B.S.; Unal, R.; Zhu, B.; Morris, A.J.; Shipp, L.R.; Lee, J.; Walton, R.G.; Adu, A.; Erfani, R.; et al. Omega-3 fatty acids reduce adipose tissue macrophages in human subjects with insulin resistance. Diabetes 2013, 62, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Santurino, C.; López-Plaza, B.; Fontecha, J.; Calvo, M.V.; Bermejo, L.M.; Gómez-Andrés, D.; Gómez-Candela, C. Consumption of Goat Cheese Naturally Rich in Omega-3 and Conjugated Linoleic Acid Improves the Cardiovascular and Inflammatory Biomarkers of Overweight and Obese Subjects: A Randomized Controlled Trial. Nutrients 2020, 12, 1315. [Google Scholar] [CrossRef]
- Ma, N.; Chang, G.; Huang, J.; Wang, Y.; Gao, Q.; Cheng, X.; Liu, J.; Shen, X. cis-9, trans-11-Conjugated Linoleic Acid Exerts an Anti-inflammatory Effect in Bovine Mammary Epithelial Cells after Escherichia coli Stimulation through NF-κB Signaling Pathway. J. Agric. Food Chem. 2019, 67, 193–200. [Google Scholar] [CrossRef]
- Hwang, S.; Ha, A.W. Intakes of Dairy and Soy Products and 10-Year Coronary Heart Disease Risk in Korean Adults. Nutrients 2024, 16, 2959. [Google Scholar] [CrossRef] [PubMed]
- Cakir, B.; Tunali-Akbay, T. Potential anticarcinogenic effect of goat milk-derived bioactive peptides on HCT-116 human colorectal carcinoma cell line. Anal. Biochem. 2021, 622, 114166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bhatia, Z.; Seshadri, S. Formulated chitosan microspheres remodelled the altered gut microbiota and liver miRNA in diet-induced Type-2 diabetic rats. Carbohydr. Res. 2025, 547, 109301. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Mondragon, A.d.C.; Lamas, A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Animal-Origin Prebiotics Based on Chitin: An Alternative for the Future? A Critical Review. Foods 2020, 9, 782. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.-S.; Kim, J.-A.; Bak, S.-S.; Byun, H.-G.; Kim, S.-K. Anti-obesity effect of carboxymethyl chitin by AMPK and aquaporin-7 pathways in 3T3-L1 adipocytes. J. Nutr. Biochem. 2011, 22, 276–281. [Google Scholar] [CrossRef]
- Bays, H.E.; Evans, J.L.; Maki, K.C.; Evans, M.; Maquet, V.; Cooper, R.; Anderson, J.W. Chitin-glucan fiber effects on oxidized low-density lipoprotein: A randomized controlled trial. Eur. J. Clin. Nutr. 2013, 67, 2–7. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, T.; Wang, L. Chitosan oligosaccharide improves the therapeutic efficacy of sitagliptin for the therapy of Chinese elderly patients with type 2 diabetes mellitus. Ther. Clin. Risk Manag. 2017, 13, 739–750. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Possemiers, S.; Verstraete, W.; De Backer, F.; Cani, P.D.; Delzenne, N.M. Dietary modulation of clostridial cluster XIVa gut bacteria (Roseburia spp.) by chitin–glucan fiber improves host metabolic alterations induced by high-fat diet in mice. J. Nutr. Biochem. 2012, 23, 51–59. [Google Scholar] [CrossRef]
- Zhang, H.; Dan, N.; Wang, Y.Q.; Gou, C.L. Protection effect of cis 9, trans 11-conjugated linoleic acid on oxidative stress and inflammatory damage in bovine mammary epithelial cells. Sci. Rep. 2024, 14, 26295. [Google Scholar] [CrossRef]
- Fujita, Y.; Kano, K.; Kishino, S.; Nagao, T.; Shen, X.; Sato, C.; Hatakeyama, H.; Ota, Y.; Niibori, S.; Nomura, A.; et al. Dietary cis-9, trans-11-conjugated linoleic acid reduces amyloid β-protein accumulation and upregulates anti-inflammatory cytokines in an Alzheimer’s disease mouse model. Sci. Rep. 2021, 11, 9749. [Google Scholar] [CrossRef]
- Valenzuela, C.A.; Baker, E.J.; Miles, E.A.; Calder, P.C. Conjugated Linoleic Acids Have Anti-Inflammatory Effects in Cultured Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 874. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, B.; Taghizadeh, A.; Paya, H.; Daghigh Kia, H. Conjugated linoleic acid (CLA) supplementation effects on performance, metabolic parameters and reproductive traits in lactating Holstein dairy cows. Vet. Res. Forum 2021, 12, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.F.; Roberts-Lee, K.; Borcea, C.; Smith, H.M.; Midgette, Y.; Shah, R. Omega-3 polyunsaturated fatty acids attenuate inflammatory activation and alter differentiation in human adipocytes. J. Nutr. Biochem. 2019, 64, 45–49. [Google Scholar] [CrossRef]
- Chacińska, M.; Zabielski, P.; Książek, M.; Szałaj, P.; Jarząbek, K.; Kojta, I.; Chabowski, A.; Błachnio-Zabielska, A.U. The Impact of OMEGA-3 Fatty Acids Supplementation on Insulin Resistance and Content of Adipocytokines and Biologically Active Lipids in Adipose Tissue of High-Fat Diet Fed Rats. Nutrients 2019, 11, 835. [Google Scholar] [CrossRef] [PubMed]
- Borja-Magno, A.; Guevara-Cruz, M.; Flores-López, A.; Carrillo-Domínguez, S.; Granados, J.; Arias, C.; Perry, M.; Sears, B.; Bourges, H.; Gómez, F.E. Differential effects of high dose omega-3 fatty acids on metabolism and inflammation in patients with obesity: Eicosapentaenoic and docosahexaenoic acid supplementation. Front. Nutr. 2023, 10, 1156995. [Google Scholar] [CrossRef]
- Sangouni, A.A.; Orang, Z.; Mozaffari-Khosravi, H. Effect of omega-3 supplementation on fatty liver and visceral adiposity indices in diabetic patients with non-alcoholic fatty liver disease: A randomized controlled trial. Clin. Nutr. ESPEN 2021, 44, 130–135. [Google Scholar] [CrossRef]
- Das, A.; Chen, C.-M.; Mu, S.-C.; Yang, S.-H.; Ju, Y.-M.; Li, S.-C. Medicinal Components in Edible Mushrooms on Diabetes Mellitus Treatment. Pharmaceutics 2022, 14, 436. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Ho, K.-J.; Hsieh, Y.-J.; Wang, L.-T.; Mau, J.-L. Contents of lovastatin, γ-aminobutyric acid and ergothioneine in mushroom fruiting bodies and mycelia. Lwt 2012, 47, 274–278. [Google Scholar] [CrossRef]
- Balakrishnan, P.; Loganayagi, C. Antihyperglycemic activity of Agaricus bisporus mushroom extracts on alloxan induced diabetic rats. Int. J. Pharma Res. Health Sci. 2018, 6, 2475–2479. [Google Scholar]
- Elhusseiny, S.M.; El-Mahdy, T.S.; Awad, M.F.; Elleboudy, N.S.; Farag, M.M.S.; Yassein, M.A.; Aboshanab, K.M. Proteome Analysis and In Vitro Antiviral, Anticancer and Antioxidant Capacities of the Aqueous Extracts of Lentinula edodes and Pleurotus ostreatus Edible Mushrooms. Molecules 2021, 26, 4623. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Bolívar, V.; García-Fontana, B.; García-Fontana, C.; Muñoz-Torres, M. Mechanisms Involved in the Relationship between Vitamin D and Insulin Resistance: Impact on Clinical Practice. Nutrients 2021, 13, 3491. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Chen, C.-M.; Ju, Y.-M.; Wu, Y.-C.; Hsieh, H.-M.; Yang, S.-H.; Su, C.-T.; Fang, T.-C.; Setyaningsih, W.; Li, S.-C. Effects of Consuming Pulsed UV Light-Treated Pleurotus citrinopileatus on Vitamin D Nutritional Status in Healthy Adults. Foods 2025, 14, 259. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Q.; Liu, H.; Guo, L.; Ma, C.; Kang, W. Regulating role of Pleurotus ostreatus insoluble dietary fiber in high fat diet induced obesity in rats based on proteomics and metabolomics analyses. Int. J. Biol. Macromol. 2024, 282, 136857. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, C.; Liu, Z.; Cui, X.; Guo, X.; Wang, H. Chemical Composition, Antioxidant and Anti-Inflammatory Activity of Shiitake Mushrooms (Lentinus edodes). J. Fungi 2024, 10, 552. [Google Scholar] [CrossRef]
- Srivastava, M.; Kumari, M.; Karn, S.K.; Bhambri, A.; Mahale, V.G.; Mahale, S. Submerged cultivation and phytochemical analysis of medicinal mushrooms (Trametes sp.). Front. Fungal Biol. 2024, 5, 1414349. [Google Scholar] [CrossRef] [PubMed]
- Harada, E.; Morizono, T.; Kanno, T.; Saito, M.; Kawagishi, H. Medicinal Mushroom, Grifola gargal (Agaricomycetes), Lowers Triglyceride in Animal Models of Obesity and Diabetes and in Adults with Prediabetes. Int. J. Med. Mushrooms 2020, 22, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Gunawardena, D.; Bennett, L.; Shanmugam, K.; King, K.; Williams, R.; Zabaras, D.; Head, R.; Ooi, L.; Gyengesi, E.; Münch, G. Anti-inflammatory effects of five commercially available mushroom species determined in lipopolysaccharide and interferon-γ activated murine macrophages. Food Chem. 2014, 148, 92–96. [Google Scholar] [CrossRef]
- Kim, Y.M.; Woo, H.W.; Shin, M.H.; Koh, S.B.; Kim, H.C.; Kim, M.K. A prospective association between dietary mushroom intake and the risk of type 2 diabetes: The Korean Genome and Epidemiology Study-Cardiovascular Disease Association Study. Epidemiol. Health 2024, 46, e2024017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, L.; Lu, Y.; Zhang, J.; Yang, M.; Tian, Y.; Dong, J.; Liao, L. Protective effect of Cordyceps sinensis against diabetic kidney disease through promoting proliferation and inhibiting apoptosis of renal proximal tubular cells. BMC Complement. Med. Ther. 2023, 23, 109. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Song, J.; Teng, M.; Zheng, X.; Li, X.; Tian, Y.; Pan, M.; Li, Y.; Lee, R.J.; Wang, D. Antidiabetic and Antinephritic Activities of Aqueous Extract of Cordyceps militaris Fruit Body in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats. Oxid. Med. Cell. Longev. 2016, 2016, 9685257. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.-Y.; Wu, H.; Cheng, Y.-L.; Kang, J.; Xu, Z.-G. The Intestinal Microbiota Composition in Early and Late Stages of Diabetic Kidney Disease. Microbiol. Spectr. 2023, 11, e00382-23. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Gnanasambandan, R. Intestinal microbiome diversity of diabetic and non-diabetic kidney disease: Current status and future perspective. Life Sci. 2023, 316, 121414. [Google Scholar] [CrossRef] [PubMed]
- Lohia, S.; Vlahou, A.; Zoidakis, J. Microbiome in Chronic Kidney Disease (CKD): An Omics Perspective. Toxins 2022, 14, 176. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xi, Y.; Yan, M.; Sun, C.; Tang, J.; Dong, X.; Yang, Z.; Wu, L. Lactiplantibacillus plantarum NKK20 Increases Intestinal Butyrate Production and Inhibits Type 2 Diabetic Kidney Injury through PI3K/Akt Pathway. J. Diabetes Res. 2023, 2023, 8810106. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, K.; Lee, Y.; Chen, M. Preventive Effects of Lactobacillus Mixture against Chronic Kidney Disease Progression through Enhancement of Beneficial Bacteria and Downregulation of Gut-Derived Uremic Toxins. J. Agric. Food Chem. 2021, 69, 7353–7366. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; DeSantis, T.Z.; Ni, Z.; Nguyen, T.H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.W.; Huang, Y.Y.; Tsai, S.Y.; Wang, J.Y.; Lin, J.H.; Syu, Z.J.; Wang, H.S.; Hsu, Y.C.; Chen, J.F.; Hsia, K.C.; et al. Probiotic Formula Ameliorates Renal Dysfunction Indicators, Glycemic Levels, and Blood Pressure in a Diabetic Nephropathy Mouse Model. Nutrients 2023, 15, 2803. [Google Scholar] [CrossRef]
- Ferguson, L.R.; De Caterina, R.; Görman, U.; Allayee, H.; Kohlmeier, M.; Prasad, C.; Choi, M.S.; Curi, R.; de Luis, D.A.; Gil, Á.; et al. Guide and Position of the International Society of Nutrigenetics/Nutrigenomics on Personalised Nutrition: Part 1—Fields of Precision Nutrition. J. Nutr. Nutr. 2016, 9, 12–27. [Google Scholar] [CrossRef]
- Kohlmeier, M.; De Caterina, R.; Ferguson, L.R.; Görman, U.; Allayee, H.; Prasad, C.; Kang, J.X.; Nicoletti, C.F.; Martinez, J.A. Guide and Position of the International Society of Nutrigenetics/Nutrigenomics on Personalized Nutrition: Part 2—Ethics, Challenges and Endeavors of Precision Nutrition. J. Nutr. Nutr. 2016, 9, 28–46. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Boba, A.; Czemplik, M.; Kulma, A.; Wojtasik, W. Influence of the Bioactive Diet Components on the Gene Expression Regulation. Nutrients 2021, 13, 3673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kutateladze, T.G. Diet and the epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef]
- La Scala, S.; Naselli, F.; Quatrini, P.; Gallo, G.; Caradonna, F. Drought-Adapted Mediterranean Diet Plants: A Source of Bioactive Molecules Able to Give Nutrigenomic Effects per sè or to Obtain Functional Foods. Int. J. Mol. Sci. 2024, 25, 2235. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, A.C.P.; Assis, I.S.S.; Salum, K.C.R.; Palhinha, L.; Abreu, G.M.; Zembrzuski, V.M.; Campos, M., Jr.; Nogueira Neto, J.F.; Mattos, F.C.C.; Cambraia, A.; et al. SH2B1 variants as potential causes of non-syndromic monogenic obesity in a Brazilian cohort. Eat. Weight. Disord. 2022, 27, 3665–3674. [Google Scholar] [CrossRef]
- Mohammed, I.; Haris, B.; Al-Barazenji, T.; Vasudeva, D.; Tomei, S.; Al Azwani, I.; Dauleh, H.; Shehzad, S.; Chirayath, S.; Mohamadsalih, G.; et al. Understanding the Genetics of Early-Onset Obesity in a Cohort of Children from Qatar. J. Clin. Endocrinol. Metab. 2023, 108, 3201–3213. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Ha, N.; Ji, Y.; Do, A.; Seo, J.H.; Oh, B.; Choi, S.; Choe, E.K.; Lee, W.; Son, J.W.; et al. Genetic determinants of obesity in Korean populations: Exploring genome-wide associations and polygenic risk scores. Brief. Bioinform. 2024, 25, bbae389. [Google Scholar] [CrossRef]
- Dashti, H.S.; Scheer, F.; Saxena, R.; Garaulet, M. Impact of polygenic score for BMI on weight loss effectiveness and genome-wide association analysis. Int. J. Obes. 2024, 48, 694–701. [Google Scholar] [CrossRef]
- Garaulet, M.; Smith, C.E.; Hernández-González, T.; Lee, Y.-C.; Ordovás, J.M. PPARγ Pro12Ala interacts with fat intake for obesity and weight loss in a behavioural treatment based on the Mediterranean diet. Mol. Nutr. Food Res. 2011, 55, 1771–1779. [Google Scholar] [CrossRef]
- Saber-Ayad, M.; Manzoor, S.; Radwan, H.; Hammoudeh, S.; Wardeh, R.; Ashraf, A.; Jabbar, H.; Hamoudi, R. The FTO genetic variants are associated with dietary intake and body mass index amongst Emirati population. PLoS ONE 2019, 14, e0223808. [Google Scholar] [CrossRef] [PubMed]
- Franzago, M.; Di Nicola, M.; Fraticelli, F.; Marchioni, M.; Stuppia, L.; Vitacolonna, E. Nutrigenetic variants and response to diet/lifestyle intervention in obese subjects: A pilot study. Acta Diabetol. 2022, 59, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Ramya, K.; Ayyappa, K.A.; Ghosh, S.; Mohan, V.; Radha, V. Genetic association of ADIPOQ gene variants with type 2 diabetes, obesity and serum adiponectin levels in south Indian population. Gene 2013, 532, 253–262. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Ping, Z.; Li, Y.; Wang, C.; Shi, Y.; Zhou, W.; Zhang, L. Interaction analysis of gene variants of TCF7L2 and body mass index and waist circumference on type 2 diabetes. Clin. Nutr. 2020, 39, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Ku, E.J.; Won, G.W.; Lee, Y.H.; Lee, D.-H.; Jeon, H.J.; Oh, T.K. Genetic variation in TCF7L2 rs7903146 correlating with peripheral arterial disease in long-standing type 2 diabetes. Diabetes Vasc. Dis. Res. 2019, 17, 1479164119888475. [Google Scholar] [CrossRef]
- Zúñiga-Hernández, S.R.; García-Iglesias, T.; Macías-Carballo, M.; Pérez-Larios, A.; Gutiérrez-Mercado, Y.K.; Camargo-Hernández, G.; Rodríguez-Razón, C.M. Targets and Effects of Common Biocompounds of Hibiscus sabdariffa (Delphinidin-3-Sambubiosid, Quercetin, and Hibiscus Acid) in Different Pathways of Human Cells According to a Bioinformatic Assay. Nutrients 2024, 16, 566. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Lahuerta, A. The Parathyroid Hormone-Related Protein/Parathyroid Hormone 1 Receptor Axis in Adipose Tissue. Biomolecules 2021, 11, 1570. [Google Scholar] [CrossRef]
- Hernando Boigues, J.F.; Mach, N. The effect of polyunsaturated fatty acids on obesity through epigenetic modifications. Endocrinol. Nutr. 2015, 62, 338–349. [Google Scholar] [CrossRef]
- Saad, B.; Ghareeb, B.; Kmail, A. Metabolic and Epigenetics Action Mechanisms of Antiobesity Medicinal Plants and Phytochemicals. Evid.-Based Complement. Altern. Med. 2021, 2021, 9995903. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, H.N.; Lin, S.-P.; Nguyen, D.H.N.; Chen, C.-M.; Su, C.-T.; Fang, T.-C.; Li, S.-C. Integrative Roles of Functional Foods, Microbiotics, Nutrigenetics, and Nutrigenomics in Managing Type 2 Diabetes and Obesity. Nutrients 2025, 17, 608. https://doi.org/10.3390/nu17040608
Lam HN, Lin S-P, Nguyen DHN, Chen C-M, Su C-T, Fang T-C, Li S-C. Integrative Roles of Functional Foods, Microbiotics, Nutrigenetics, and Nutrigenomics in Managing Type 2 Diabetes and Obesity. Nutrients. 2025; 17(4):608. https://doi.org/10.3390/nu17040608
Chicago/Turabian StyleLam, Hong Nhung, Shih-Ping Lin, Dang Hien Ngan Nguyen, Chiao-Ming Chen, Chien-Tien Su, Te-Chao Fang, and Sing-Chung Li. 2025. "Integrative Roles of Functional Foods, Microbiotics, Nutrigenetics, and Nutrigenomics in Managing Type 2 Diabetes and Obesity" Nutrients 17, no. 4: 608. https://doi.org/10.3390/nu17040608
APA StyleLam, H. N., Lin, S.-P., Nguyen, D. H. N., Chen, C.-M., Su, C.-T., Fang, T.-C., & Li, S.-C. (2025). Integrative Roles of Functional Foods, Microbiotics, Nutrigenetics, and Nutrigenomics in Managing Type 2 Diabetes and Obesity. Nutrients, 17(4), 608. https://doi.org/10.3390/nu17040608