Hepatoprotective Activity of the Fruits of Eleutherococcus senticosus in Acetaminophen-Induced Liver Injury in Mice and Their Chemical Composition

 , , , and
, , , and
Abstract
1. Introduction
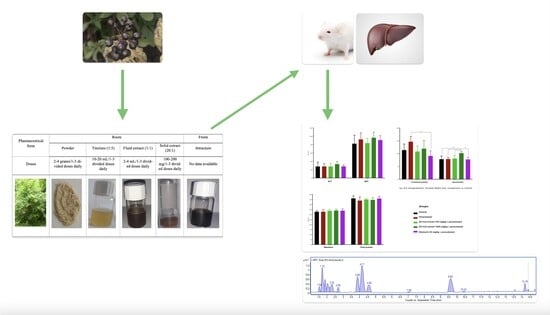
2. Results and Discussion
2.1. Evaluation of the Hepatoprotective Effect of the Eleutherococcus senticosus Fruit Extract in a Model of Liver Damage with Paracetamol
2.2. Histopathological Studies
2.3. Phytochemical Characterization-Amino Acid Profile
2.4. Low Molecular Weight Organic Acid
2.5. Secondary Metabolites
2.6. Mineral Composition
3. Materials and Methods
3.1. Plant Material
3.2. Evaluation of the Hepatoprotective Effect of the Eleutherococcus senticosus Fruit Extract in a Model of Liver Damage with Paracetamol
- Group 1 (Control): Water (per os (PO), 0.1 mL/10 g) for 7 days + vehicle (EtOH/PEG/NaCl) intraperitoneal (IP) on day 8,
- Group 2: Paracetamol (300 mg/kg, IP, day 8),
- Group 3: Eleutherococcus senticosus fruit extract (750 mg/kg, PO, twice daily for 7 days and once on day 8) + paracetamol (300 mg/kg, IP, day 8),
- Group 4: Eleutherococcus senticosus fruit extract (1500 mg/kg, PO, twice daily for 7 days and once on day 8) + paracetamol (300 mg/kg, IP, day 8),
- Group 5: Silymarin (50 mg/kg, PO, twice daily for 7 days and once on day 8) + paracetamol (300 mg/kg, IP, day 8).
3.3. Histopathological Studies
3.4. Ultra-High Performance Liquid Chromatography (UHPLC-DAD-MS)
3.5. Elemental Analysis
3.6. Statistical Analysis
- on the complete dataset, including all biochemical and hematological parameters,
- on a subset of data containing only injury markers (ALT, AST, creatinine, urea),
- on a subset of data comprising hematological and immune response parameters.
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| ATP | adenosine triphosphate |
| COX-2 | cyclooxygenase-2 |
| CV | coefficient of variation |
| ES | Eleutherococcus senticosus |
| EtOH/PEG/NaCl | ethanol/polyethylene glycol/sodium chloride |
| GFAP | glial fibrillary acidic protein |
| H&E | hematoxylin and eosin |
| HGB | hemoglobin |
| HPLC-DAD | high-performance liquid chromatography with diode-array detection |
| ICP-OES | inductively coupled plasma optical emission spectrometry |
| IP | intraperitoneal |
| MVP | mean platelet volume |
| NAFLD | non-alcoholic fatty liver disease |
| NAPQI | N-acetyl-p-benzoquinoneimine |
| PCT | plateletcrit |
| PDW | platelet distribution width (platelet anisocytosis) |
| PO | per os (by mouth) |
| RBC | red blood cells |
| TCA | tricarboxylic acid |
| UHPLC | ultra-high-performance liquid chromatography |
References
- Schmidt, M.; Thomsen, M.; Kelber, O.; Kraft, K. Myths and Facts in Herbal Medicines: Eleutherococcus senticosus (Siberian Ginseng) and Its Contraindication in Hypertensive Patients. hypertensive patients. Bot. Targets Ther. 2014, 27, 27–32. [Google Scholar] [CrossRef]
- Li, W.; Luo, Q.; Jin, L.H. Acanthopanax senticosus Extracts Have a Protective Effect on Drosophila Gut Immunity. J. Ethnopharmacol. 2013, 146, 257–263. [Google Scholar] [CrossRef]
- Załuski, D.; Olech, M.; Verpoorte, R.; Khan, I.; Kuźniewski, R.; Nowak, R. Phytoconstituents and Nutritional Properties of the Fruits of Eleutherococcus divaricatus and Eleutherococcus sessiliflorus: A Study of Non-European Species Cultivated in Poland. Oxid. Med. Cell. Longev. 2017, 2017, 8374295. [Google Scholar] [CrossRef]
- Chang, Y.; Jiang, Y.; Chen, J.; Li, S.; Wang, Y.; Chai, L.; Ma, J.; Wang, Z. Comprehensive Analysis of Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. Fruits Based on UPLC–MS/MS and GC–MS: A Rapid Qualitative Analysis. Food Sci. Nutr. 2024, 12, 1911–1927. [Google Scholar] [CrossRef]
- Gębalski, J.; Małkowska, M.; Kiełkowska, E.; Graczyk, F.; Wnorowska, S.; Hołyńska-Iwan, I.; Strzemski, M.; Wójciak, M.; Załuski, D. Chemical Composition and In Vitro Biological Activity of the Polar and Non-Polar Fractions Obtained from the Roots of Eleutherococcus senticosus (Rupr. et Maxim.) Maxim. Int. J. Mol. Sci. 2025, 26, 5619. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.; Ivanov, K.; Delattre, C.; Nalbantova, V.; Karcheva-Bahchevanska, D.; Ivanova, S. Plant Adaptogens—History and Future Perspectives. Nutrients 2021, 13, 2861. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Meng, Y.; Zhai, C.; Wang, M.; Avula, B.; Yuk, J.; Smith, K.M.; Isaac, G.; Khan, I.A. The Chemical Characterization of Eleutherococcus senticosus and Ci-Wu-Jia Tea Using UHPLC-UV-QTOF/MS. Int. J. Mol. Sci. 2019, 20, 475. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Cho, M.L.; Kim, D.B.; Shin, G.H.; Lee, J.H.; Lee, J.S.; Park, S.O.; Lee, S.J.; Shin, H.M.; Lee, O.H. The Antioxidant Activity and Their Major Antioxidant Compounds from Acanthopanax senticosus and A. koreanum. Molecules 2015, 20, 13281–13295. [Google Scholar] [CrossRef]
- Graczyk, F.; Orzechowska, B.; Franz, D.; Strzemski, M.; Verpoorte, R.; Załuski, D. The Intractum from the Eleutherococcus senticosus Fruits Affects the Innate Immunity in Human Leukocytes: From the Ethnomedicinal Use to Contemporary Evidence-Based Research. J. Ethnopharmacol. 2021, 268, 113636. [Google Scholar] [CrossRef]
- Załuski, D.; Janeczko, Z. Variation in Phytochemicals and Bioactivity of the Fruits of Eleutherococcus Species Cultivated in Poland. Nat. Prod. Res. 2015, 29, 2207–2211. [Google Scholar] [CrossRef]
- Graczyk, F.; Gębalski, J.; Sulejczak, D.; Małkowska, M.; Wójciak, M.; Gawenda-Kempczyńska, D.; Piskorska, E.; Krolik, K.; Markiewicz, M.; Kondrzycka-Dąda, A.; et al. UHPLC-DAD/ESI-TOF-MS Phytochemical Characterization and Evaluation of the Impact of Eleutherococcus senticosus Fruit Intractum on Biochemical, Hepatological, and Blood Parameters in Balb/c Mice. Int. J. Mol. Sci. 2024, 25, 9295. [Google Scholar] [CrossRef]
- Miyauchi-Wakuda, S.; Kagota, S.; Maruyama-Fumoto, K.; Shiokawa, Y.; Yamada, S.; Shinozuka, K. Acanthopanax senticosus Root Extract Exerts Dual Action on Mouse Ileal Smooth Muscle Function, Leading to Modulation of Gastrointestinal Motility. Biol. Pharm. Bull. 2020, 43, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Van Crombruggen, K.; Holtappels, G.; Bachert, C. A Herbal Composition of Scutellaria Baicalensis and Eleutherococcus senticosus Shows Potent Anti-Inflammatory Effects in an Ex Vivo Human Mucosal Tissue Model. Evid.-Based Complement. Altern. Med. 2012, 2012, 673145. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Al-Harbi, M.S. Amelioration of Paracetamol Hepatotoxicity and Oxidative Stress on Mice Liver with Silymarin and Nigella Sativa Extract Supplements. Asian Pac. J. Trop. Biomed. 2015, 5, 521–531. [Google Scholar] [CrossRef]
- Papackova, Z.; Heczkova, M.; Dankova, H.; Sticova, E.; Lodererova, A.; Bartonova, L.; Poruba, M.; Cahova, M. Silymarin Prevents Acetaminophen-Induced Hepatotoxicity in Mice. PLoS ONE 2018, 13, e0191353. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Madrigal-Santillán, E.; Morales-González, Á.; Esquivel-Soto, J.; Esquivel-Chirino, C.; García-Luna y González-Rubio, M.G.; Gayosso-de-Lucio, J.A.; Morales-González, J.A. Hepatoprotective Effect of Silymarin. World J. Hepatol. 2014, 6, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Rasool, M.K.; Sabina, E.P.; Ramya, S.R.; Preety, P.; Patel, S.; Mandal, N.; Mishra, P.P.; Samuel, J. Hepatoprotective and Antioxidant Effects of Gallic Acid in Paracetamol-Induced Liver Damage in Mice. J. Pharm. Pharmacol. 2010, 62, 638–643. [Google Scholar] [CrossRef]
- Graczyk, F.; Gębalski, J.; Makuch-Kocka, A.; Gawenda-Kempczyńska, D.; Ptaszyńska, A.A.; Grzyb, S.; Bogucka-Kocka, A.; Załuski, D. Phenolic Profile, Antioxidant, Anti-Enzymatic and Cytotoxic Activity of the Fruits and Roots of Eleutherococcus senticosus (Rupr. et Maxim.) Maxim. Molecules 2022, 27, 5579. [Google Scholar] [CrossRef]
- Facchinetti, F.; Neri, I.; Tarabusi, M. Eleutherococcus senticosus Reduces Cardiovascular Stress Response in Healthy Subjects: A Randomized, Placebo-Controlled Trial. Stress. Health 2002, 18, 11–17. [Google Scholar] [CrossRef]
- Lim, S.H.; Park, Y.H.; Kwon, C.J.; Ham, H.J.; Jeong, H.N.; Kim, K.H.; Ahn, Y.S. Anti-Diabetic and Hypoglycemic Effect of Eleutherococcus Spp. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1761–1768. [Google Scholar] [CrossRef]
- Hart, J.G.; Timbrell, J.A. The Effect of Age on Paracetamol Hepatotoxicity in Mice. Biochem. Pharmacol. 1979, 28, 3015–3017. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Huang, P.C. Antioxidant and Hepatoprotective Effects of Acathopanax Senticosus. Phytother. Res. 2000, 14, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Elblehi, S.S.; Gad El-Karim, D.R.S. Potential Amelorative Effect of Panax Ginseng on Hepatorenal Damage Ocuured in Hydroxurea-Treated Rats. Slov. Vet. Res. 2019, 56, 595–606. [Google Scholar] [CrossRef]
- Hu, J.N.; Liu, Z.; Wang, Z.; Li, X.D.; Zhang, L.X.; Li, W.; Wang, Y.P. Ameliorative Effects and Possible Molecular Mechanism of Action of Black Ginseng (Panax Ginseng) on Acetaminophen-Mediated Liver Injury. Molecules 2017, 22, 664. [Google Scholar] [CrossRef]
- Jang, D.; Lee, J.; Eom, S.H.; Lee, S.M.; Gil, J.; Lim, H.B.; Hyun, T.K. Composition, Antioxidant and Antimicrobial Activities of Eleutherococcus Senticosus Fruit Extracts. J. Appl. Pharm. Sci. 2016, 6, 125–130. [Google Scholar] [CrossRef]
- Załuski, D.; Olech, M.; Galanty, A.; Verpoorte, R.; Kuźniewski, R.; Nowak, R.; Bogucka-Kocka, A. Phytochemical Content and Pharma-Nutrition Study on Eleutherococcus senticosus Fruits Intractum. Oxid. Med. Cell. Longev. 2016, 2016, 9270691. [Google Scholar] [CrossRef]
- Su, J.; Zhang, X.; Kan, Q.; Chu, X. Antioxidant Activity of Acanthopanax senticosus Flavonoids in H2O2-Induced RAW 264.7 Cells and DSS-Induced Colitis in Mice. Molecules 2022, 27, 2872. [Google Scholar] [CrossRef]
- Su, J.; Fu, X.; Liu, R.; Zhang, X.; Xue, J.; Li, Y.; Zhang, R.; Li, X.; Wang, X.; Wang, X.; et al. Antioxidant and Antiapoptotic Properties of N-Butanol Fraction the Acanthopanax senticosus Extracts in H2O2-RAW264.7 Cells and CCl4-Induced Liver Injury in Mice. Evid.-Based Complement. Altern. Med. 2023, 2023, 9190198. [Google Scholar] [CrossRef]
- Song, Z.; Jiao, C.; Chen, B.; Xu, W.; Wang, M.; Zou, J.; Xu, W.; Xu, Z.; Wang, Q. Dietary Acanthopanax senticosus Extracts Modulated the Inflammatory and Apoptotic Responses of Yellow Catfish to Protect against Edwardsiella ictaluri Infection. Aquac. Res. 2021, 52, 5078–5092. [Google Scholar] [CrossRef]
- Rostami, S.; Arefhosseini, S.; Tutunchi, H.; Khoshbaten, M.; Ebrahimi-Mameghani, M. Does Myo-Inositol Supplementation Influence Oxidative Stress Biomarkers in Patients with Non-Alcoholic Fatty Liver Disease? Food Sci. Nutr. 2024, 12, 1279–1289. [Google Scholar] [CrossRef]
- Pan, S.; Yan, X.; Dong, X.; Li, T.; Suo, X.; Tan, B.; Zhang, S.; Li, Z.; Yang, Y.; Zhang, H. The Positive Effects of Dietary Inositol on Juvenile Hybrid Grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatu) Fed High-Lipid Diets: Growthperformance, Antioxidant Capacity and Immunity. Fish Shellfish Immunol. 2022, 126, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.S.; Li, N.R.; Zhao, W.L.; Liu, H.; Ge, M.X.; Zhang, Y.X.; Zhao, L.Y.; You, X.F.; He, H.W.; Shao, R.G. D-Chiro-Inositol Effectively Attenuates Cholestasis in Bile Duct Ligated Rats by Improving Bile Acid Secretion and Attenuating Oxidative Stress. Acta Pharmacol. Sin. 2018, 39, 213–221. [Google Scholar] [CrossRef]
- Lee, E.; Lim, Y.; Kwon, S.W.; Kwon, O. Pinitol Consumption Improves Liver Health Status by Reducing Oxidative Stress and Fatty Acid Accumulation in Subjects with Non-Alcoholic Fatty Liver Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Nutr. Biochem. 2019, 68, 33–41. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Li, Y.; Zhang, Z.; Liang, Y. Antioxidant and Prooxidant Activities of Phenolic Acids Commonly Existed in Vegetables and Their Relationship with Structures. Food Sci. Technol. 2022, 42, e07622. [Google Scholar] [CrossRef]
- Kanane, A.; Rouaki, F.; Errahmani, M.B.; Laraba, A.; Mesbah, H.; Ammouche, A. Original Communication Pro-Oxidant Effects of a High α-Tocopherol Dose on Kidney Antioxidant Biomarkers and Histopathological Aspects. Int. J. Vitam. Nutr. Res. 2017, 87, 179–190. [Google Scholar] [CrossRef]
- Forester, S.C.; Lambert, J.D. The Role of Antioxidant versus Pro-Oxidant Effects of Green Tea Polyphenols in Cancer Prevention. Mol. Nutr. Food Res. 2011, 55, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Tryniszewski, W.; Sarniak, A.; Wlodarczyk, A.; Nowak, P.J.; Nowak, D. Concentration Dependence of Anti- and Pro-Oxidant Activity of Polyphenols as Evaluated with a Light-Emitting Fe2+-Egta-H2O2 System. Molecules 2022, 27, 3453. [Google Scholar] [CrossRef]
- Smalinskiene, A.; Lesauskaite, V.; Zitkevicius, V.; Savickiene, N.; Savickas, A.; Ryselis, S.; Sadauskiene, I.; Ivanov, L. Estimation of the Combined Effect of Eleutherococcus senticosus Extract and Cadmium on Liver Cells. Ann. N. Y. Acad. Sci. 2009, 1171, 314–320. [Google Scholar] [CrossRef]
- Fu, J.; Fu, J.; Yuan, J.; Zhang, N.; Gao, B.; Fu, G.; Tu, Y.; Zhang, Y. Anti-Diabetic Activities of Acanthopanax senticosus Polysaccharide (ASP) in Combination with Metformin. Int. J. Biol. Macromol. 2012, 50, 619–623. [Google Scholar] [CrossRef]
- Lee, D.; Park, J.; Yoon, J.; Kim, M.Y.; Choi, H.Y.; Kim, H. Neuroprotective Effects of Eleutherococcus senticosus Bark on Transient Global Cerebral Ischemia in Rats. J. Ethnopharmacol. 2012, 139, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Kim, S.W.; Seo, H.W.; Hyun, S.H.; Kyung, J.S.; Youn, S.H.; So, S.H.; In, G.; Park, C.K.; Yi, E.C.; et al. Long-Term Evaluation of Safety and Biological Effects of Korean Red Ginseng (Panax ginseng): A Long-Term in Vivo Study. BMC Complement. Med. Ther. 2022, 22, 284. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione Synthesis. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef]
- Kurokawa, Y.; Matsuzawa, A.; Ogawa, H.; Hirata, Y.; Noguchi, T.; Kawashima, H. Adverse Roles of Citric Acid and L-Histidine in the Transition Metal-Dependent Generation of Hydroxyl Radical at Circumneutral PH. Res. Sq. 2024. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, N.; Zhou, J.; Sun, P.; Zhao, L.; Zhou, F. Protective Effects of Several Common Amino Acids, Vitamins, Organic Acids, Flavonoids and Phenolic Acids against Hepatocyte Damage Caused by Alcohol. Foods 2022, 11, 3014. [Google Scholar] [CrossRef]
- Kos, G.; Czarnek, K.; Sadok, I.; Krzyszczak-Turczyn, A.; Kubica, P.; Fila, K.; Emre, G.; Tatarczak-Michalewska, M.; Latalska, M.; Blicharska, E.; et al. Eleutherococcus senticosus (Acanthopanax senticosus): An Important Adaptogenic Plant. Molecules 2025, 30, 2512. [Google Scholar] [CrossRef] [PubMed]
- Himoto, T.; Masaki, T. Current Trends of Essential Trace Elements in Patients with Chronic Liver Diseases. Nutrients 2020, 12, 2084. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, F.; Gębalski, J.; Piskorska, E.; Małkowska, M.; Słomka, A.; Gawenda-Kempczyńska, D.; Kondrzycka-Dąda, A.; Olszewska-Słonina, D.; Styczyński, J.; Taglialatela-Scafati, O.; et al. The Eleutherococcus senticosus Fruits’ Intractum Affects Changes in the Transepithelial Electric Potential in the Distal Section of the Rabbit’s Large Intestine and Inhibits Hyaluronidase. J. Ethnopharmacol. 2024, 325, 117847. [Google Scholar] [CrossRef] [PubMed]
- Dresler, S.; Wójciak, M.; Sowa, I.; Sawicki, J.; Strzemski, M.; Hawrylak-Nowak, B.; Hanaka, A. Accumulation Ability of Trace Metals by Silver Birch Leaves in Areas Contaminated by Zn–Pb Ore Processing: Effects of Excessive Trace Metal Accumulation on Specialized Metabolism. Chemosphere 2024, 362, 142719. [Google Scholar] [CrossRef]
- ISO 17294-1:2004; Water Quality—Applications of Inductively Coupled Plasma Mass Spectrometry (ICP-MS)—Part 1: General Guidance. Available online: https://www.iso.org/standard/32957.html (accessed on 30 September 2020).
- ISO 17294-2:2016; Water Quality—Applications of Inductively Coupled Plasma Mass Spectrometry (ICP-MS)—Part 2: Determination of Selected Elements, Including Isotopes of Uranium. Available online: https://www.iso.org/standard/62962.html (accessed on 30 September 2020).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | % Body Mass | ||||
|---|---|---|---|---|---|
| Liver | Kidneys | Spleen | Lungs | Brain | |
| 1. control | 4.43 | 1.19 | 0.44 | 0.64 | 2.09 |
| 2. paracetamol 300 mg/kg | 4.45 | 1.28 | 0.44 | 0.76 | 2.23 |
| 3. ES fruit extract 750 mg/kg + paracetamol | 4.20 | 1.3 | 0.41 | 0.74 | 2.27 |
| 4. ES fruit extract 1500 mg/kg + paracetamol | 4.25 | 1.28 | 0.4 | 0.73 | 2.37 |
| 5. Silymarin 50 mg/kg + paracetamol | 4.19 | 1.31 | 0.39 | 0.74 | 2.21 |
| RT (min.) | Extraction Window [m/z+H] | Mass Error ppm Range | Formula | Compound | Content (mg/g ± SD) |
|---|---|---|---|---|---|
| 1.57 | 147.112–147.114 | −5.5 to 8.18 | C6H14N2O2 | lysine | 0.36 ± 0.02 |
| 1.69 | 156.076–156.078 | −4.86 to 8.04 | C6H9N3O2 | histidine | 0.09 ± 0.01 |
| 1.69 | 175.118–175.12 | −5.47 to 6.02 | C6H14N4O2 | arginine | 1.15 ± 0.07 |
| 1.69 | 76.039–76.04 | −4.06 to 9.26 | C2H5NO2 | glycine | 0.27 ± 0.02 |
| 1.70 | 106.049–106.051 | −8.28 to 10.76 | C3H7NO3 | serine | 1.54 ± 0.11 |
| 1.72 | 134.044–134.046 | −5.89 to 9.14 | C4H7NO4 | aspartic acid | 0.08 ± 0.01 |
| 1.73 | 120.065–120.067 | −4.36 to 12.43 | C4H9NO3 | threonine | 1.01 ± 0.08 |
| 1.72 | 90.054–90.056 | −10.72 to 11.54 | C3H7NO2 | alanine | 0.98 ± 0.06 |
| 1.74 | 147.075–147.077 | −9.71 to 3.98 | C5H10N2O3 | glutamine | 0.69 ± 0.04 |
| 1.77 | 148.059–148.061 | −9.75 to 3.85 | C5H9NO4 | glutamic acid | 1.36 ± 0.09 |
| 1.89 | 116.07–116.072 | −5.26 to 12.12 | C5H9NO2 | proline | 0.63 ± 0.04 |
| 2.05 | 118.085–118.087 | −10.72 to 6.36 | C5H11NO2 | valine | 1.25 ± 0.07 |
| 2.24 | 241.03–241.033 | −4.69 to 7.81 | C6H12N2O4S2 | cystine | detected |
| 2.30 | 118.085–118.0087 | −10.72 to 6.36 | C5H11NO2 | norvaline | 1.13 ± 0.08 |
| 2.69 | 150.057–150.059 | −8.89 to 4.52 | C5H11NO2S | methionine | 0.42 ± 0.31 |
| 3.87 | 132.101–132.103 | −6.9 to 8.35 | C6H13NO2 | isoleucine | 2.13 ± 0.17 |
| 4.17 | 132.101–132.103 | −6.9 to 8.35 | C6H13NO2 | leucine | 3.55 ± 0.24 |
| 4.60 | 182.080–182.082 | −6.46 to 4.59 | C9H11NO3 | tyrosine | 3.98 ± 0.21 |
| 9.61 | 166.085–166.087 | −7.6 to 4.51 | C9H11NO2 | phenylalanine | 2.75 ± 0.11 |
| 14.19 | 205.096–205.098 | −5.65 to 4.14 | C11H12N2O2 | tryptophan | 1.05 ± 0.07 |
| Total amino acids | 24.42 | ||||
| RT (min) | Observed Ion Mass [M–H]− | Mass Error (ppm) | Formula | Identified | Content (µg/g ± SD) |
|---|---|---|---|---|---|
| 1.77 | 195.05153 | 2.57 | C6H12O7 | gluconic acid | 2.11 ± 0.14 |
| 2.29 | 133.01489 | 4.80 | C4H6O5 | malic acid | 25.12 ± 1.14 |
| 3.39 | 191.01882 | −4.72 | C6H8O7 | citric acid | 8.39 ± 0.36 |
| Total acids | 61.46 |
| Observed Ion Mass [m/z−H]− | Formula | Identified | Content (mg/g ± SD) |
|---|---|---|---|
| 153.01932 | C7H6O4 | Protocatechuic acid | 0.43 ± 0.02 |
| 353.087–353.089 | C16H18O9 | Chlorogenic acids | 28.02 ± 1.21 |
| 335.077–335.079 | C16H16O8 | Caffeoylshikimic acids | 1.03 ± 0.05 |
| 367.103–367.104 | C17H20O9 | Feruloylquinic acids | 0.45 ± 0.02 |
| 515.118–515.121 | C25H24O12 | Dicaffeoylquinic acids | 13.42 ± 0.76 |
| 207.06715 | C11H12O4 | Caffeic acid derivative | 0.42 ± 0.02 |
| Total phenolic acids | 43.77 | ||
| 451.12473 | C21H24O11 | Catechin hexoside | 0.13 ± 0.01 |
| 289.07182 | C15H14O6 | Catechin | 0.28 ± 0.02 |
| 609.145–609.148 | C27H30O16 | Quercetin O-rutinosides | 0.74 ± 0.02 |
| 463.088–463.09 (300) | C21H20O12 | Quercetin 3-O-hexosides | 0.88 ± 0.03 |
| 301.03656 | C15H10O7 | Quercetin | 0.19 ± 0.01 |
| Total flavonoids | 2.22 |
| Element | Wavelength (nm) | Content (µg/g) | CV (%) |
|---|---|---|---|
| Ca | 315.8869 | 5788 | 0.08 |
| K | 766.4911 | 24,181 | 0.27 |
| Mg | 258.2162 | 1369 | 0.69 |
| Na | 589.5924 | 291.7 | 0.16 |
| Se | 196.0280 | 292.9 | 0.09 |
| Zn | 206.2000 | 33.12 | 3.15 |
| Fe | 259.9396 | nd | – |
| Cu | 327.3960 | nd | – |
| Mn | 257.6100 | nd | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graczyk, F.; Krolik, K.; Gawenda-Kempczyńska, D.; Wójciak, M.; Sowa, I.; Sulejczak, D. Hepatoprotective Activity of the Fruits of Eleutherococcus senticosus in Acetaminophen-Induced Liver Injury in Mice and Their Chemical Composition. Nutrients 2025, 17, 3456. https://doi.org/10.3390/nu17213456
Graczyk F, Krolik K, Gawenda-Kempczyńska D, Wójciak M, Sowa I, Sulejczak D. Hepatoprotective Activity of the Fruits of Eleutherococcus senticosus in Acetaminophen-Induced Liver Injury in Mice and Their Chemical Composition. Nutrients. 2025; 17(21):3456. https://doi.org/10.3390/nu17213456
Chicago/Turabian StyleGraczyk, Filip, Krystian Krolik, Dorota Gawenda-Kempczyńska, Magdalena Wójciak, Ireneusz Sowa, and Dorota Sulejczak. 2025. "Hepatoprotective Activity of the Fruits of Eleutherococcus senticosus in Acetaminophen-Induced Liver Injury in Mice and Their Chemical Composition" Nutrients 17, no. 21: 3456. https://doi.org/10.3390/nu17213456
APA StyleGraczyk, F., Krolik, K., Gawenda-Kempczyńska, D., Wójciak, M., Sowa, I., & Sulejczak, D. (2025). Hepatoprotective Activity of the Fruits of Eleutherococcus senticosus in Acetaminophen-Induced Liver Injury in Mice and Their Chemical Composition. Nutrients, 17(21), 3456. https://doi.org/10.3390/nu17213456