
A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Attenuates Hypergravity-Induced Disruptions to the Microbiome in Female Mice
 and
and
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Experimental Design
2.3. NASA Centrifuge
2.4. Diets
2.5. Fecal Sample Collection Procedures
2.6. 16S rRNA Gene Sequencing
2.7. Hematology
2.8. Statistical Analysis
3. Results
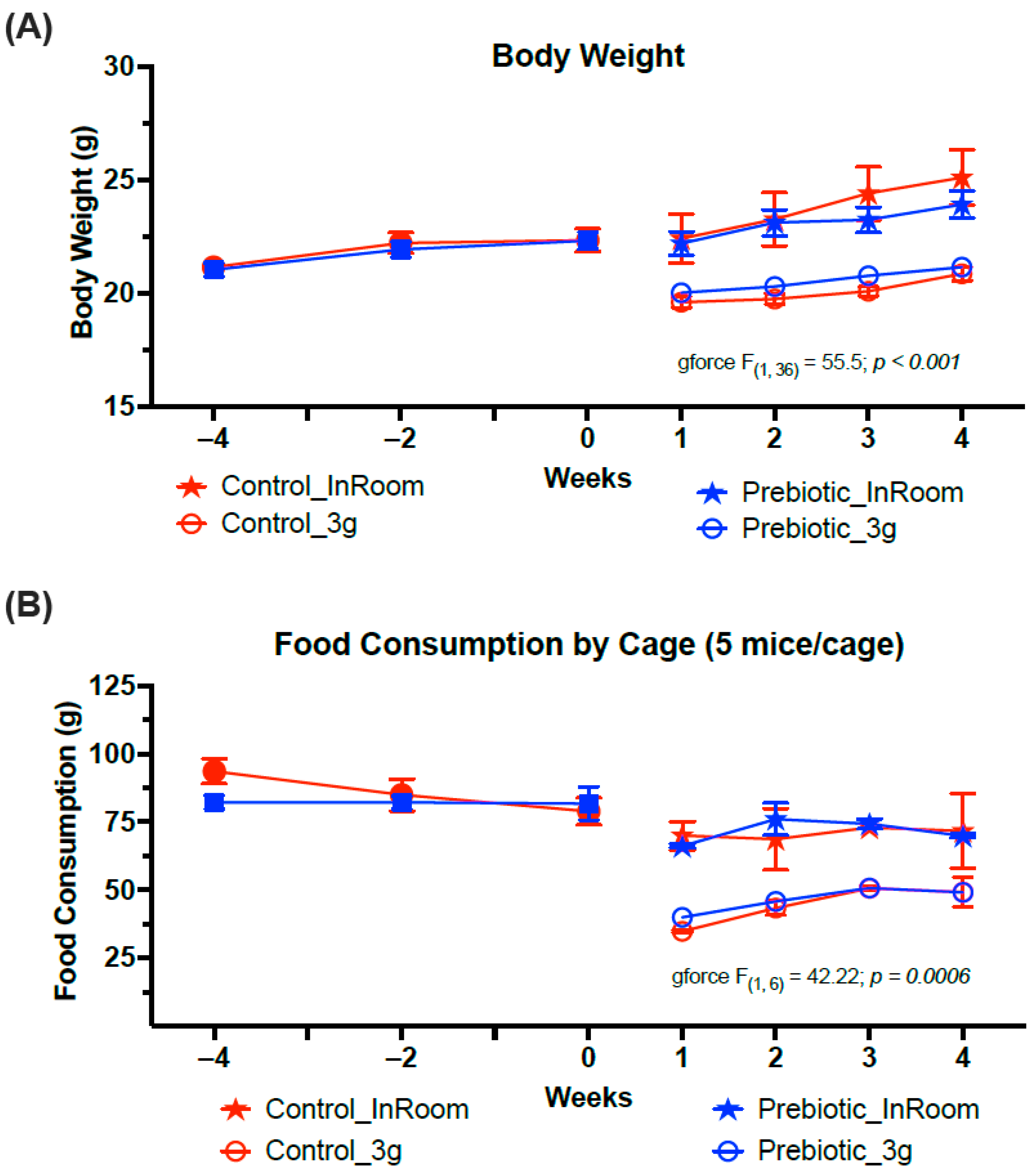
3.1. Body/Food Weight–3g Reduced Body Weight and Food Consumption
3.2. Microbiome
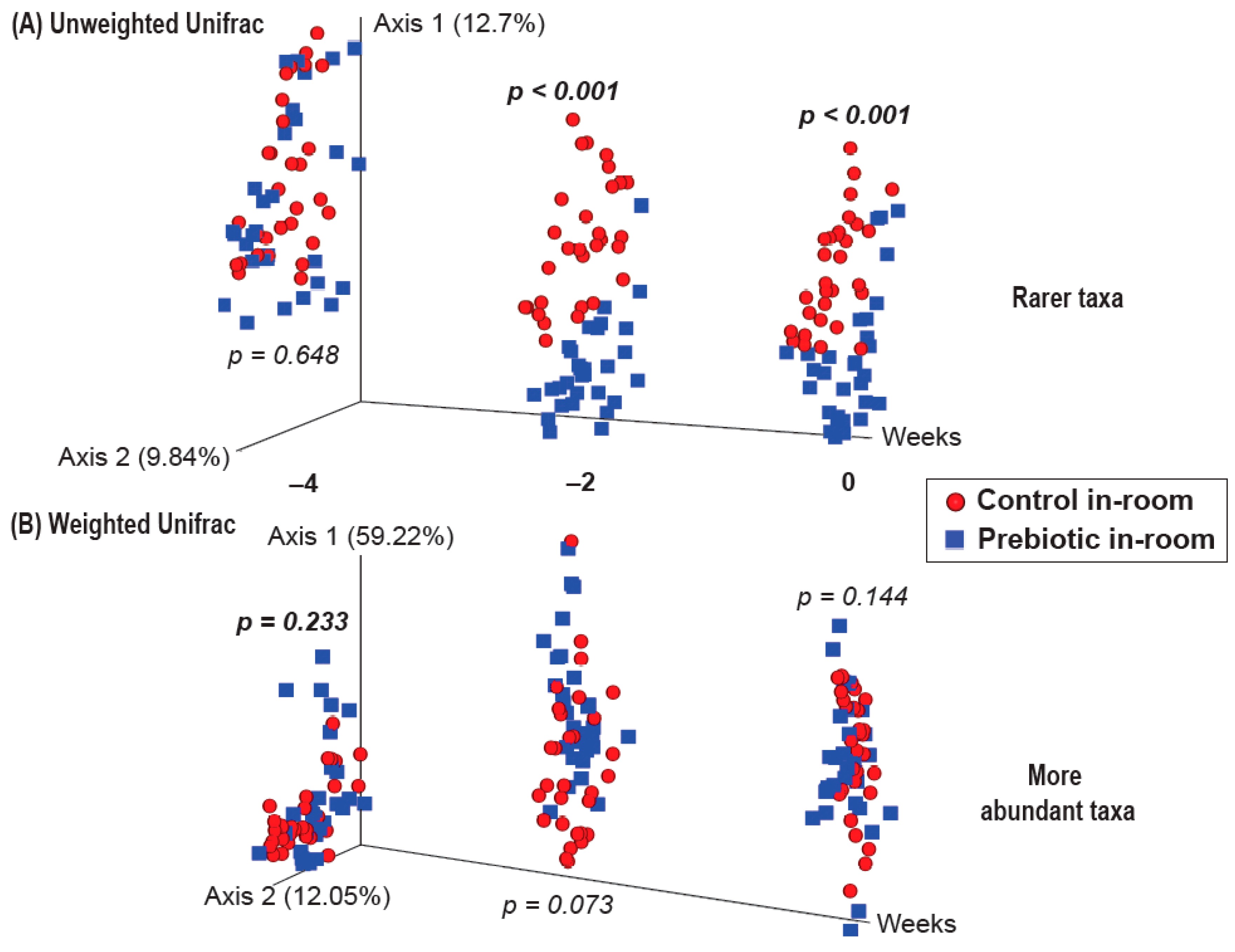
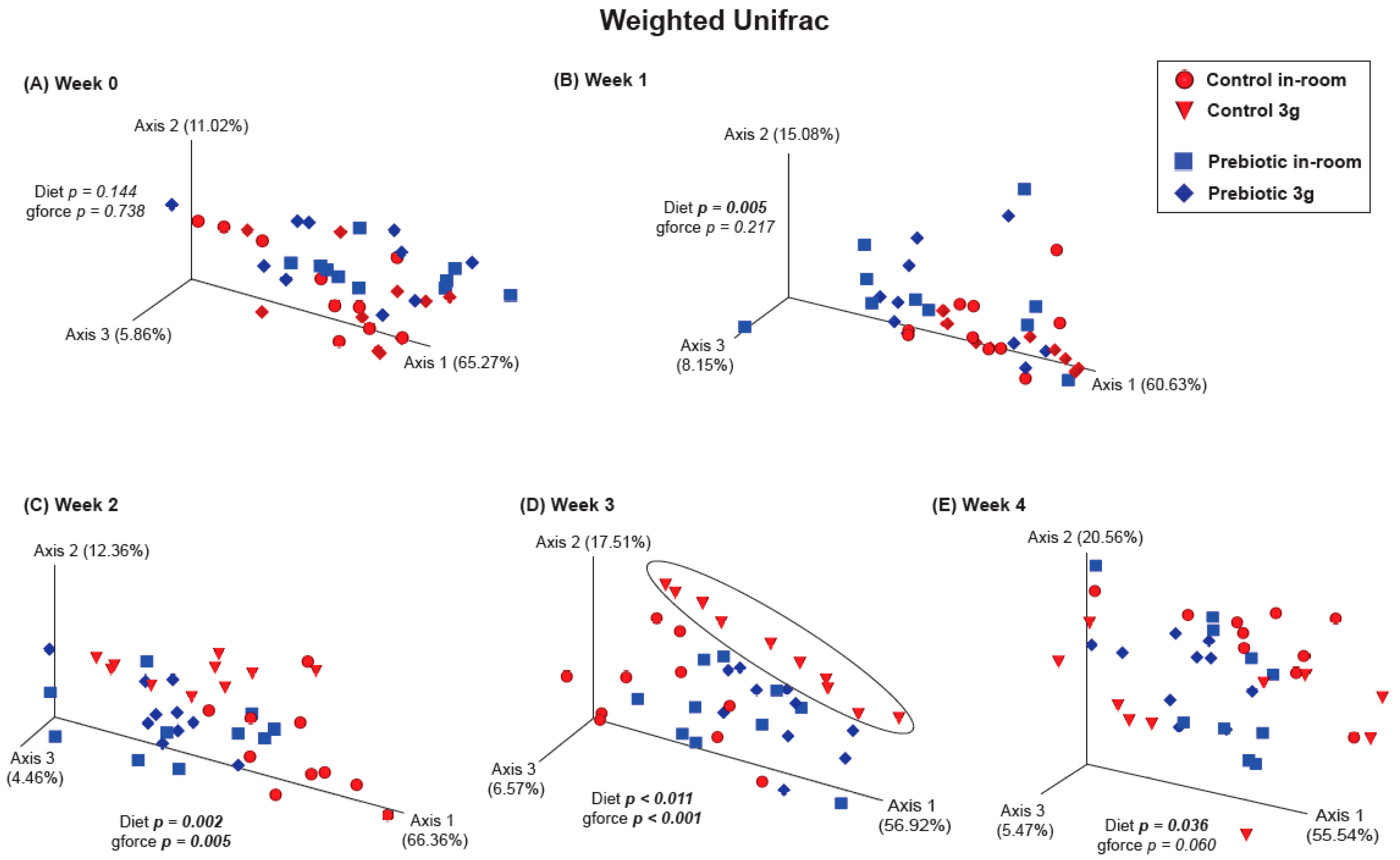
3.2.1. β-Diversity–Prebiotic Diet Attenuated 3g-Induced Changes
3.2.2. α-Diversity–Prebiotic Diet Reduced Measures of Alpha Diversity
3.3. Taxonomy–Prebiotic Diet Attenuated 3g-Induced Increases in Several Taxa
3.4. Organ Weights/Blood Panel Are Altered by Prebiotic Diet and 3g
3.5. Stepwise Multiple Regression Analysis Reveals Significant Relationships Between NLR, the Lungs, and the Gut Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bowers, S.J.; Summa, K.C.; Thompson, R.S.; Gonzalez, A.; Vargas, F.; Olker, C.; Jiang, P.; Lowry, C.A.; Dorrestein, P.C.; Knight, R.; et al. A Prebiotic Diet Alters the Fecal Microbiome and Improves Sleep in Response to Sleep Disruption in Rats. Front. Neurosci. 2022, 16, 889211. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Gaffney, M.; Hopkins, S.; Kelley, T.; Gonzalez, A.; Bowers, S.J.; Vitaterna, M.H.; Turek, F.W.; Foxx, C.L.; Lowry, C.A.; et al. Ruminiclostridium 5, Parabacteroides distasonis, and bile acid profile are modulated by prebiotic diet and associate with facilitated sleep/clock realignment after chronic disruption of rhythms. Brain Behav. Immun. 2021, 97, 150–166. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Roller, R.; Mika, A.; Greenwood, B.N.; Knight, R.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Dietary Prebiotics and Bioactive Milk Fractions Improve NREM Sleep, Enhance REM Sleep Rebound and Attenuate the Stress-Induced Decrease in Diurnal Temperature and Gut Microbial Alpha Diversity. Front. Behav. Neurosci. 2017, 10, 240. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Vargas, F.; Dorrestein, P.C.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Dietary prebiotics alter novel microbial dependent fecal metabolites that improve sleep. Sci. Rep. 2020, 10, 3848. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, J.L.; Hong, B.Y. Dysbiosis and Immune Dysregulation in Outer Space. Int. Rev. Immunol. 2016, 35, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Green, S.J.; Chlipala, G.E.; Turek, F.W.; Vitaterna, M.H. Reproducible changes in the gut microbiome suggest a shift in microbial and host metabolism during spaceflight. Microbiome 2019, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Tierney, B.T.; Kim, J.; Overbey, E.G.; Ryon, K.A.; Foox, J.; Sierra, M.A.; Bhattacharya, C.; Damle, N.; Najjar, D.; Park, J.; et al. Longitudinal multi-omics analysis of host microbiome architecture and immune responses during short-term spaceflight. Nat. Microbiol. 2024, 9, 1661–1675. [Google Scholar] [CrossRef] [PubMed]
- Markina, E.A.; Andrianova, I.V.; Shtemberg, A.S.; Buravkova, L.B. Effect of 30-Day Hindlimb Unloading and Hypergravity on Bone Marrow Stromal Progenitors in C57Bl/6N Mice. Bull. Exp. Biol. Med. 2018, 166, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Alauzet, C.; Cunat, L.; Wack, M.; Lozniewski, A.; Busby, H.; Agrinier, N.; Cailliez-Grimal, C.; Frippiat, J.P. Hypergravity disrupts murine intestinal microbiota. Sci. Rep. 2019, 9, 9410. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.M.; Mhatre, S.D.; Cekanaviciute, E.; Schreurs, A.S.; Tahimic, C.G.T.; Globus, R.K.; Anand, S.; Crucian, B.E.; Bhattacharya, S. Neutrophil-to-Lymphocyte Ratio: A Biomarker to Monitor the Immune Status of Astronauts. Front. Immunol. 2020, 11, 564950. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. NPJ Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Miller, G.M.; Nelson, G.A.; Gridley, D.S. Hypergravity-induced immunomodulation in a rodent model: Hematological and lymphocyte function analyses. J. Appl. Physiol. (1985) 2004, 97, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Pecaut, M.J.; Slater, J.M.; Gridley, D.S. Spaceflight modulates expression of extracellular matrix, adhesion, and profibrotic molecules in mouse lung. J. Appl. Physiol. (1985) 2010, 108, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Poyraz, T.; Cilengiroglu, O.V. Blood biomarkers in acute ischemic stroke: The prognostic value of neutrophil-to-lymphocyte ratio and mean platelet volume. Adv. Clin. Exp. Med. 2024, 33, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.Y.; Kim, H.N.; Lee, S.H.; Kim, S.J.; Chang, Y.; Ryu, S.; Shin, H.; Kim, H.L.; Lee, J.H. Association between Neutrophil-to-Lymphocyte Ratio and Gut Microbiota in a Large Population: A Retrospective Cross-Sectional Study. Sci. Rep. 2018, 8, 16031. [Google Scholar] [CrossRef] [PubMed]
- Lane, H.W.; Alfrey, C.P.; Driscoll, T.B.; Smith, S.M.; Nyquist, L.E. Control of red blood cell mass during spaceflight. J. Gravit. Physiol. 1996, 3, 87–88. [Google Scholar] [PubMed]
- Alfrey, C.P.; Udden, M.M.; Leach-Huntoon, C.; Driscoll, T.; Pickett, M.H. Control of red blood cell mass in spaceflight. J. Appl. Physiol. (1985) 1996, 81, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Salvagno, G.L.; Sanchis-Gomar, F.; Picanza, A.; Lippi, G. Red blood cell distribution width: A simple parameter with multiple clinical applications. Crit. Rev. Clin. Lab. Sci. 2015, 52, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Nelson, G.A.; Peters, L.L.; Kostenuik, P.J.; Bateman, T.A.; Morony, S.; Stodieck, L.S.; Lacey, D.L.; Simske, S.J.; Pecaut, M.J. Genetic models in applied physiology: Selected contribution: Effects of spaceflight on immunity in the C57BL/6 mouse. II. Activation, cytokines, erythrocytes, and platelets. J. Appl. Physiol. (1985) 2003, 94, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Aldosari, K.H.; Ahmad, G.; Al-Ghamdi, S.; Alsharif, M.H.K.; Elamin, A.Y.; Musthafa, M.; Abbas, M.Y.; Alqarni, A.A.; Alqudeebi, S.K.; Binsaqer, A.A.; et al. The influence and impact of smoking on red blood cell morphology and buccal microflora: A case-control study. J. Clin. Lab. Anal. 2020, 34, e23212. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, A.; Wysokinski, A.; Strzelecki, D.; Gawlik-Kotelnicka, O. Limited Changes in Red Blood Cell Parameters After Probiotic Supplementation in Depressive Individuals: Insights from a Secondary Analysis of the PRO-DEMET Randomized Controlled Trial. J. Clin. Med. 2025, 14, 265. [Google Scholar] [CrossRef] [PubMed]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Day, H.E.; Martinez, A.; Rumian, N.L.; Greenwood, B.N.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Early life diets with prebiotics and bioactive milk fractions attenuate the impact of stress on learned helplessness behaviours and alter gene expression within neural circuits important for stress resistance. Eur. J. Neurosci. 2017, 45, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Bowers, S.J.; Vargas, F.; Hopkins, S.; Kelley, T.; Gonzalez, A.; Lowry, C.A.; Dorrestein, P.C.; Vitaterna, M.H.; Turek, F.W.; et al. A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Produces Dynamic and Reproducible Changes in the Gut Microbial Ecosystem in Male Rats. Nutrients 2024, 16, 1790. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Ahmed, W.; Zafar, S.; Farooq, U.; Abid, J.; Shah, H.B.U.; Akram, S.; Ghazanfar, M.; Ahmad, A.M.R. Effect of inulin, galacto oligosaccharides and iron fortification on iron deficiency anemia among women of reproductive age; a randomized controlled trial. Front. Nutr. 2022, 9, 1028956. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Fleshner, M. Early-life exercise may promote lasting brain and metabolic health through gut bacterial metabolites. Immunol. Cell Biol. 2016, 94, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Maslanik, T.; Mahaffey, L.; Tannura, K.; Beninson, L.; Greenwood, B.N.; Fleshner, M. The inflammasome and danger associated molecular patterns (DAMPs) are implicated in cytokine and chemokine responses following stressor exposure. Brain Behav. Immun. 2013, 28, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Bowers, S.J.; Vargas, F.; Gonzalez, A.; He, S.; Jiang, P.; Dorrestein, P.C.; Knight, R.; Wright, K.P., Jr.; Lowry, C.A.; Fleshner, M.; et al. Repeated sleep disruption in mice leads to persistent shifts in the fecal microbiome and metabolome. PLoS ONE 2020, 15, e0229001. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef] [PubMed]
- Janssen, S.; McDonald, D.; Gonzalez, A.; Navas-Molina, J.A.; Jiang, L.; Xu, Z.Z.; Winker, K.; Kado, D.M.; Orwoll, E.; Manary, M.; et al. Phylogenetic Placement of Exact Amplicon Sequences Improves Associations with Clinical Information. mSystems 2018, 3, e00021-18. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and "All-species Living Tree Project (LTP)" taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed]
- Knights, D.; Kuczynski, J.; Koren, O.; Ley, R.E.; Field, D.; Knight, R.; DeSantis, T.Z.; Kelley, S.T. Supervised classification of microbiota mitigates mislabeling errors. ISME J. 2011, 5, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project, C. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Phylogenetic pattern and the quantification of organismal biodiversity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1994, 345, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbo, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Baqai, F.P.; Gridley, D.S. Impact of total-body irradiation on the response to a live bacterial challenge. Int. J. Radiat. Biol. 2014, 90, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.J.; Gross, R.; Bittinger, K.; Sherrill-Mix, S.; Lewis, J.D.; Collman, R.G.; Bushman, F.D.; Li, H. Power and sample-size estimation for microbiome studies using pairwise distances and PERMANOVA. Bioinformatics 2015, 31, 2461–2468. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.Z.; Chen, G.; Alekseyenko, A.V. PERMANOVA-S: Association test for microbial community composition that accommodates confounders and multiple distances. Bioinformatics 2016, 32, 2618–2625. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ishima, T.; Qu, Y.; Shan, J.; Chang, L.; Wei, Y.; Zhang, J.; Pu, Y.; Fujita, Y.; Tan, Y.; et al. Ingestion of Faecalibaculum rodentium causes depression-like phenotypes in resilient Ephx2 knock-out mice: A role of brain-gut-microbiota axis via the subdiaphragmatic vagus nerve. J. Affect. Disord. 2021, 292, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, C.B.; Lichatz, R.; Pich, A.; Muhlfeld, C.; Woltemate, S.; Vital, M.; Brandenberger, C. Short-chain fatty acids improve inflamm-aging and acute lung injury in old mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2023, 324, L480–L492. [Google Scholar] [CrossRef] [PubMed]
- Zagato, E.; Pozzi, C.; Bertocchi, A.; Schioppa, T.; Saccheri, F.; Guglietta, S.; Fosso, B.; Melocchi, L.; Nizzoli, G.; Troisi, J.; et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 2020, 5, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Qaisar, R.; Khan, N.A.; Alharbi, A.M.; Alfahemi, H.; Elmoselhi, A. Effect of Microgravity on the Gut Microbiota Bacterial Composition in a Hindlimb Unloading Model. Life 2022, 12, 1865. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhang, R.; Huang, F.; Dong, L.; Liu, L.; Jia, X.; Chi, J.; Ma, Y.; Deng, M.; Chen, Y.; et al. Hydrolyzed Bound Phenolics from Rice Bran Alleviate Hyperlipidemia and Improve Gut Microbiota Dysbiosis in High-Fat-Diet Fed Mice. Nutrients 2022, 14, 1277. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, N.E.; Yoon, H.; Shin, C.M.; Kim, N.; Lee, D.H.; Park, J.Y.; Choi, C.H.; Kim, J.G.; Kim, Y.K.; et al. Fecal Microbiota and Gut Microbe-Derived Extracellular Vesicles in Colorectal Cancer. Front. Oncol. 2021, 11, 650026. [Google Scholar] [CrossRef] [PubMed]
- Tajasuwan, L.; Kettawan, A.; Rungruang, T.; Wunjuntuk, K.; Prombutara, P. Role of Dietary Defatted Rice Bran in the Modulation of Gut Microbiota in AOM/DSS-Induced Colitis-Associated Colorectal Cancer Rat Model. Nutrients 2023, 15, 1528. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Zhang, D.; Wu, J.; Liu, J.; Tan, Y.; Feng, W.; Peng, C. Atractylodes macrocephala Koidz. volatile oil relieves acute ulcerative colitis via regulating gut microbiota and gut microbiota metabolism. Front. Immunol. 2023, 14, 1127785. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Liu, S.; Zhang, W.; Zhu, H.; Tao, Q.; Wang, H.; Yan, H. Impact of traditional Chinese medicine treatment on chronic unpredictable mild stress-induced depression-like behaviors: Intestinal microbiota and gut microbiome function. Food Funct. 2019, 10, 5886–5897. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wang, Q.; Yu, T.; Hu, H.; Wu, G.; Duan, X.; Jiang, R.; Xu, Y.; Huang, Y. Quercetin ameliorates bone loss in OVX rats by modulating the intestinal flora-SCFAs-inflammatory signaling axis. Int. Immunopharmacol. 2024, 136, 112341. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Shao, D.; Han, C.; Huang, Q.; Zhao, W. Response of human gut microbiota under simulated microgravity. Appl. Microbiol. Biotechnol. 2022, 106, 5221–5231. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Feng, S.Y.; Li, X.J.; Liu, T.H.; Ye, S.Y.; Ma, L.; Man, S.L. Litchi procyanidins inhibit colon cancer proliferation and metastasis by triggering gut-lung axis immunotherapy. Cell Death Dis. 2023, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.Q.; Martinez-Alvaro, M.; Lima, J.; Auffret, M.D.; Rutherford, K.M.D.; Simm, G.; Dewhurst, R.J.; Baima, E.T.; Roehe, R. Identification of intestinal and fecal microbial biomarkers using a porcine social stress model. Front. Microbiol. 2023, 14, 1197371. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, H.; Cao, Y.; Wang, C.; Zhao, C.; Wang, H.; Cui, G.; Wang, M.; Pan, Y.; Shi, Y.; et al. Fluoxetine ameliorates dysbiosis in a depression model induced by chronic unpredicted mild stress in mice. Int. J. Med. Sci. 2019, 16, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Zhao, Z.; Xiao, C.; Li, L.; Liu, Y.E.; Fu, J.; Liao, H.; Zhou, T.; Zhang, J. Gut microbiome promotes mice recovery from stress-induced depression by rescuing hippocampal neurogenesis. Neurobiol. Dis. 2024, 191, 106396. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Wang, Q.; Bai, F.; Li, Z.; Li, Y.; Liu, W.; Yan, Y.; Zhang, S.; Gao, C.; Yu, Y. Role of microbiota-gut-brain axis in natural aging-related alterations in behavior. Front. Neurosci. 2024, 18, 1362239. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.H.; Wang, J.; Zhang, C.Y.; Zhao, L.; Sheng, Y.Y.; Tao, G.S.; Xue, Y.Z. Gut microbial characteristical comparison reveals potential anti-aging function of Dubosiella newyorkensis in mice. Front. Endocrinol. 2023, 14, 1133167. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Han, X.; Ruan, M.; Huang, F.; Yang, L.; Xu, T.; Wang, H.; Wu, H.; Shi, S.; Wang, Y.; et al. Prebiotic inulin controls Th17 cells mediated central nervous system autoimmunity through modulating the gut microbiota and short chain fatty acids. Gut Microbes 2024, 16, 2402547. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, L.; Jiang, T.; Liang, T.; Yang, L.; Li, Y.; Gao, H.; Zhang, J.; Xie, X.; Wu, Q. Lactiplantibacillus plantarum 124 Modulates Sleep Deprivation-Associated Markers of Intestinal Barrier Dysfunction in Mice in Conjunction with the Regulation of Gut Microbiota. Nutrients 2023, 15, 4002. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Wu, Y.; Liang, J.; Yang, S.; Huang, Z.; Hu, J.; Yang, Q.; Liu, F.; Li, S. Gut microbiota has important roles in the obstructive sleep apnea-induced inflammation and consequent neurocognitive impairment. Front. Microbiol. 2024, 15, 1457348. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Wu, J.L.; Huang, W.C. Effects of prebiotics on intestinal physiology, neuropsychological function, and exercise capacity of mice with sleep deprivation. Food Res. Int. 2023, 165, 112568. [Google Scholar] [CrossRef] [PubMed]
- Manuel, C.R.; Latuga, M.S.; Ashby, C.R., Jr.; Reznik, S.E. Immune tolerance attenuates gut dysbiosis, dysregulated uterine gene expression and high-fat diet potentiated preterm birth in mice. Am. J. Obstet. Gynecol. 2019, 220, 596.e1–596.e28. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, Z.; Ye, B.; Ma, J.H.; Ji, S.; Sheng, W.; Ye, S.; Ou, Y.; Peng, Y.; Yang, X.; et al. Sodium butyrate ameliorates diabetic retinopathy in mice via the regulation of gut microbiota and related short-chain fatty acids. J. Transl. Med. 2023, 21, 451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, F.; Shen, Y.; Chen, Y.; Ma, J. Sleep apnea is associated with the increase of certain genera of Ruminococcaceae and Lachnospiraceae in the gut microbiome of hypertensive patients. Expert. Rev. Respir. Med. 2022, 16, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Kang, Z.; He, Y.; Qiu, Y.; Song, Y.; Liu, W. Association between gut microbiota and common overlapping gastrointestinal disorders: A bidirectional two-sample Mendelian randomization study. Front. Microbiol. 2024, 15, 1343564. [Google Scholar] [CrossRef] [PubMed]
- Mercier-Bonin, M.; Chapot-Chartier, M.P. Surface Proteins of Lactococcus lactis: Bacterial Resources for Muco-adhesion in the Gastrointestinal Tract. Front. Microbiol. 2017, 8, 2247. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, F.; Guo, Y.; Du, Q.; Zeng, X.; Wu, Z.; Guo, Y.; Tu, M.; Pan, D. Potential prebiotic properties and proliferation mechanism of fermented milk-derived polypeptides. Food Chem. 2025, 463, 141335. [Google Scholar] [CrossRef] [PubMed]
- Azcarate-Peril, M.A.; Roach, J.; Marsh, A.; Chey, W.D.; Sandborn, W.J.; Ritter, A.J.; Savaiano, D.A.; Klaenhammer, T.R. A double-blind, 377-subject randomized study identifies Ruminococcus, Coprococcus, Christensenella, and Collinsella as long-term potential key players in the modulation of the gut microbiome of lactose intolerant individuals by galacto-oligosaccharides. Gut Microbes 2021, 13, 1957536. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.B.; Chen, H.; Oliver, B.G.G. The Lungs in Space: A Review of Current Knowledge and Methodologies. Cells 2024, 13, 1154. [Google Scholar] [CrossRef] [PubMed]
- Limper, U.; Ahnert, T.; Maegele, M.; Froehlich, M.; Grau, M.; Gauger, P.; Bauerfeind, U.; Gorlinger, K.; Potzsch, B.; Jordan, J. Simulated Hypergravity Activates Hemostasis in Healthy Volunteers. J. Am. Heart Assoc. 2020, 9, e016479. [Google Scholar] [CrossRef] [PubMed]
- Trudel, G.; Shahin, N.; Ramsay, T.; Laneuville, O.; Louati, H. Hemolysis contributes to anemia during long-duration space flight. Nat. Med. 2022, 28, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Waisberg, E.; Ong, J.; Masalkhi, M.; Lee, A.G. Anemia and Spaceflight Associated Neuro-Ocular Syndrome (SANS). Prehosp. Disaster Med. 2023, 38, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Gaffney, M.; Roller, R.; Hills, A.; Bouchet, C.A.; Hulen, K.A.; Thompson, R.S.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Feeding the developing brain: Juvenile rats fed diet rich in prebiotics and bioactive milk fractions exhibit reduced anxiety-related behavior and modified gene expression in emotion circuits. Neurosci. Lett. 2018, 677, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Tokgoz, S.; Keskin, S.; Kayrak, M.; Seyithanoglu, A.; Ogmegul, A. Is neutrophil/lymphocyte ratio predict to short-term mortality in acute cerebral infarct independently from infarct volume? J. Stroke Cerebrovasc. Dis. 2014, 23, 2163–2168. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Huang, W.; Chen, X.; Li, Q.; Cai, Z.; Yu, T.; Shao, B. Neutrophil-to-Lymphocyte Ratio Is a Prognostic Marker in Acute Ischemic Stroke. J. Stroke Cerebrovasc. Dis. 2017, 26, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Giede-Jeppe, A.; Madzar, D.; Sembill, J.A.; Sprugel, M.I.; Atay, S.; Hoelter, P.; Lucking, H.; Huttner, H.B.; Bobinger, T. Increased Neutrophil-to-Lymphocyte Ratio is Associated with Unfavorable Functional Outcome in Acute Ischemic Stroke. Neurocrit. Care 2020, 33, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Switonska, M.; Piekus-Slomka, N.; Slomka, A.; Sokal, P.; Zekanowska, E.; Lattanzi, S. Neutrophil-to-Lymphocyte Ratio and Symptomatic Hemorrhagic Transformation in Ischemic Stroke Patients Undergoing Revascularization. Brain Sci. 2020, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Demirci, S.; Demirci, S.; Kutluhan, S.; Koyuncuoglu, H.R.; Yurekli, V.A. The clinical significance of the neutrophil-to-lymphocyte ratio in multiple sclerosis. Int. J. Neurosci. 2016, 126, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.N.; Tong, M.S.; Sung, P.H.; Chen, Y.L.; Chen, C.H.; Tsai, N.W.; Huang, C.J.; Chang, Y.T.; Chen, S.F.; Chang, W.N.; et al. Higher neutrophil counts and neutrophil-to-lymphocyte ratio predict prognostic outcomes in patients after non-atrial fibrillation-caused ischemic stroke. Biomed. J. 2017, 40, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Hasselbalch, I.C.; Sondergaard, H.B.; Koch-Henriksen, N.; Olsson, A.; Ullum, H.; Sellebjerg, F.; Oturai, A.B. The neutrophil-to-lymphocyte ratio is associated with multiple sclerosis. Mult. Scler. J. Exp. Transl. Clin. 2018, 4, 2055217318813183. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Cook, E.J.; Goulder, F.; Justin, T.A.; Keeling, N.J. Neutrophil-lymphocyte ratio as a prognostic factor in colorectal cancer. J. Surg. Oncol. 2005, 91, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Seifi, N.; Sedaghat, A.; Nematy, M.; Khadem-Rezaiyan, M.; Shirazinezhad, R.; Ranjbar, G.; Safarian, M. Effects of synbiotic supplementation on the serum endotoxin level, inflammatory status, and clinical outcomes of adult patients with critical illness: A randomized controlled trial. Nutr. Clin. Pract. 2022, 37, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Nakaoka, K.; Ohno, E.; Kuramitsu, K.; Kuzuya, T.; Funasaka, K.; Tochio, T.; Fujii, T.; Takahashi, H.; Kondo, N.; Miyahara, R.; et al. Efficacy of 1-Kestose Supplementation in Patients with Pancreatic Ductal Adenocarcinoma: A Randomized Controlled Pilot Study. Nutrients 2024, 16, 2889. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nonparametric Longitudinal Data (nparLD) Table: ANOVA-Type Statistics (ATSs) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Altered Mainly by 3g | Weeks −4–0 (Pre 3g) | Weeks 1–4 (Post 3g) | ||||||
| Diet | Diet × Time | Diet | Gforce (3g) | Diet × 3g | Diet × Time | 3g × Time | Diet × 3g × Time | |
| Faecalibaculum | ns | F(2,29.87) = 3.36; p = 0.039 | F(1,2.11) = 10.94; p = 0.0009 | F(1,2.11) = 57.41; p = 3.54 × 10−14 | F(1,2.11) = 4.02; p = 0.045 | ns | F(3,26.70) = 6.27; p = 0.002 | F(3,26.70) = 4.62; p = 0.009 |
| Bacteroides | F(1,1.92) = 3.23; p = 0.072 | ns | ns | F(1,2.59) = 16.21; p = 0.00006 | ns | ns | ns | ns |
| Clostridium_sensu_stricto_1 | ns | F(2,33.45) = 8.35; p = 0.0003 | F(1,2.96) = 23.12; p = 0.000002 | F(1,2.96) = 8.77; p = 0.003 | F(1,2.96) = 6.91; p = 0.009 | ns | F(3,20.14) = 3.41; p = 0.02 | ns |
| Turicibacter | ns | F(2,30.37) = 12.95; p = 0.000003 | F(1,2.54) = 22.12; p = 0.000003 | F(1,2.54) = 29.11; p = 6.83 × 10−8 | ns | F(3,20.82) = 3.96; p = 0.012 | ns | F(3,20.82) = 3.55; p = 0.019 |
| Alistipes | F(1,1.83) = 10.00; p = 0.0009 | ns | F(1,2.62) = 24.28; p = 8.32 × 10−7 | F(1,2.62) = 10.97; p = 0.0009 | F(1,2.62) = 10.29; p = 0.001 | ns | ns | F(3,27.49) = 3.31; p = 0.024 |
| Lachnospiraceae_UCG-006 | F(1,1.97) = 23.59; p = 0.000001 | ns | F(1,2.74) = 46.94; p = 7.30 × 10−12 | F(1,2.74) = 8.95; p = 0.003 | F(1,2.74) = 7.10; p = 0.008 | F(3,21.76) = 17.45; p = 0.0006 | ns | F(3,21.76) = 2.97; p = 0.035 |
| Romboutsia | F(1,1.80) = 21.80; p = 0.000003 | F(2,28.48) = 20.12; p = 9.68 × 10−9 | F(1,2.87) = 100.80; p = 1.02 × 10−23 | F(1,2.87) = 4.46; p = 0.035 | ns | ns | F(3,22.64) = 3.42; p = 0.018 | F(3,22.64) = 3.62; p = 0.014 |
| Altered Mainly by Diet | ||||||||
| Dubosiella | F(1,1.99) = 7.05; p = 0.008 | ns | F(1,2.63) = 35.25; p = 2.89 × 10−9 | ns | ns | ns | F(3,24.33) = 3.33; p = 0.024 | ns |
| Muribaculaceae | ns | ns | ns | ns | ns | F(3,27.40) = 4.02; p = 0.008 | ns | ns |
| Lachnospiraceae_NK4A136 group | F(1,1.91) = 5.81; p = 0.016 | ns | F(1,2.66) = 28.33; p = 1.02 × 10−7 | ns | ns | F(3,29.80) = 3.94; p = 0.011 | F(3,29.80) = 5.09; p = 0.003 | ns |
| Lactococcus | F(1,1.89) = 14.57; p = 0.00013 | ns | F(1,2.54) = 104.91; p = 1.28 × 10−24 | ns | ns | ns | F(3,34.30) = 8.01; p = 0.00008 | ns |
| Bifidobacterium | ns | ns | F(1,2.52) = 8.36; p = 0.004 | F(1,2.52) = 6.36; p = 0.012 | ns | ns | ns | ns |
| Diet | 3g | Mean ± SEM (Units) | |
|---|---|---|---|
| Organ Weights/Body Weight | |||
| Liver | ns | ns | 0.043 ± 0.0007 (g) |
| Lungs (Figure 9A) | ns | F(1,36) = 4.62; p = 0.039 | 0.009 ± 0.0006 (g) |
| Heart | ns | ns | 0.007± 0.0004 (g) |
| Spleen | ns | ns | 0.0035 ± 0.0003 (g) |
| [Cort] & [Glucose] & Heart Rate | |||
| Corticosterone | ns | ns | 3078 ± 221 (pg/mL) |
| Glucose | ns | ns | 266.8 ± 12.81 (mg/dL) |
| Heart Rate | ns | ns | 528 ± 5.87 (bpm) |
| Hematology | |||
| White Blood Cells | ns | ns | 2.10 ± 0.19 (103/μL) |
| Neutrophil % (Figure 9D) | F(1,36) = 3.13; p = 0.086 | ns | 24.22 ± 1.48 (%) |
| Lymphocyte % (Figure 9E) | F(1,36) = 3.53; p = 0.068 | ns | 65.94 ± 2.24 (%) |
| NLR (Figure 9F) | F(1,36) = 4.12; p = 0.049 | ns | 0.429 ± 0.05 (a.u.) |
| Monocyte % (Figure 9G) | F(1,36) = 3.65; p = 0.064 | ns | 3.78 ± 0.45 (%) |
| Eosinophils % (Figure 9H) | ns | ns | 4.28 ± 0.38 (%) |
| Basophils % (Figure 9I) | F(1,36) = 3.19; p = 0.082 | ns | 1.78 ± 0.33 (%) |
| Red Blood Cells | ns | ns | 7.34 ± 0.27 (106/μL) |
| Hemoglobin | ns | ns | 11.24 ± 0.40 (g/dL) |
| HCT % | ns | ns | 35.47 ± 1.18 (%) |
| MCV | ns | ns | 48.46 ± 0.34 (fL) |
| MCH (Figure 9B) | F(1,36) = 7.76; p = 0.008 | F(1,36) = 10.47; p = 0.003 | 15.34 ± 0.05 (pg) |
| [MCHC] | ns | ns | 31.59 ± 0.18 (g/dL) |
| RDW-CV % (Figure 9C) | F(1,36) = 8.65; p = 0.006 | F(1,36) = 9.15; p = 0.005 | 14.09 ± 0.14 (%) |
| Platelets | ns | ns | 546 ± 36.72 (103/μL) |
| Mean Platelet Vol. | ns | ns | 5.14 ± 0.03 (fL) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, R.S.; Hopkins, S.; Kelley, T.; Wilson, C.G.; Pecaut, M.J.; Fleshner, M. A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Attenuates Hypergravity-Induced Disruptions to the Microbiome in Female Mice. Nutrients 2025, 17, 2417. https://doi.org/10.3390/nu17152417
Thompson RS, Hopkins S, Kelley T, Wilson CG, Pecaut MJ, Fleshner M. A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Attenuates Hypergravity-Induced Disruptions to the Microbiome in Female Mice. Nutrients. 2025; 17(15):2417. https://doi.org/10.3390/nu17152417
Chicago/Turabian StyleThompson, Robert S., Shelby Hopkins, Tel Kelley, Christopher G. Wilson, Michael J. Pecaut, and Monika Fleshner. 2025. "A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Attenuates Hypergravity-Induced Disruptions to the Microbiome in Female Mice" Nutrients 17, no. 15: 2417. https://doi.org/10.3390/nu17152417
APA StyleThompson, R. S., Hopkins, S., Kelley, T., Wilson, C. G., Pecaut, M. J., & Fleshner, M. (2025). A Prebiotic Diet Containing Galactooligosaccharides and Polydextrose Attenuates Hypergravity-Induced Disruptions to the Microbiome in Female Mice. Nutrients, 17(15), 2417. https://doi.org/10.3390/nu17152417