
Lactiplantibacillus plantarum 06CC2 Enhanced the Expression of Intestinal Uric Acid Excretion Transporter in Mice

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of LP06CC2
2.3. Animal Experiments
2.4. Western Blot Analysis
2.5. Measurement of UA
2.5.1. UA Extraction from Feces
2.5.2. Sample Preparation
2.5.3. Measurement
2.6. Measurement of XOD Activity
2.7. Measurement of Protein-Derived Gut Microbiota Metabolites in the Feces
2.8. Measurement of Short-Chain Fatty Acids in the Cecal Contents and Feces
2.9. Statistical Analysis
3. Results
3.1. Growth Parameters and Tissue Weights
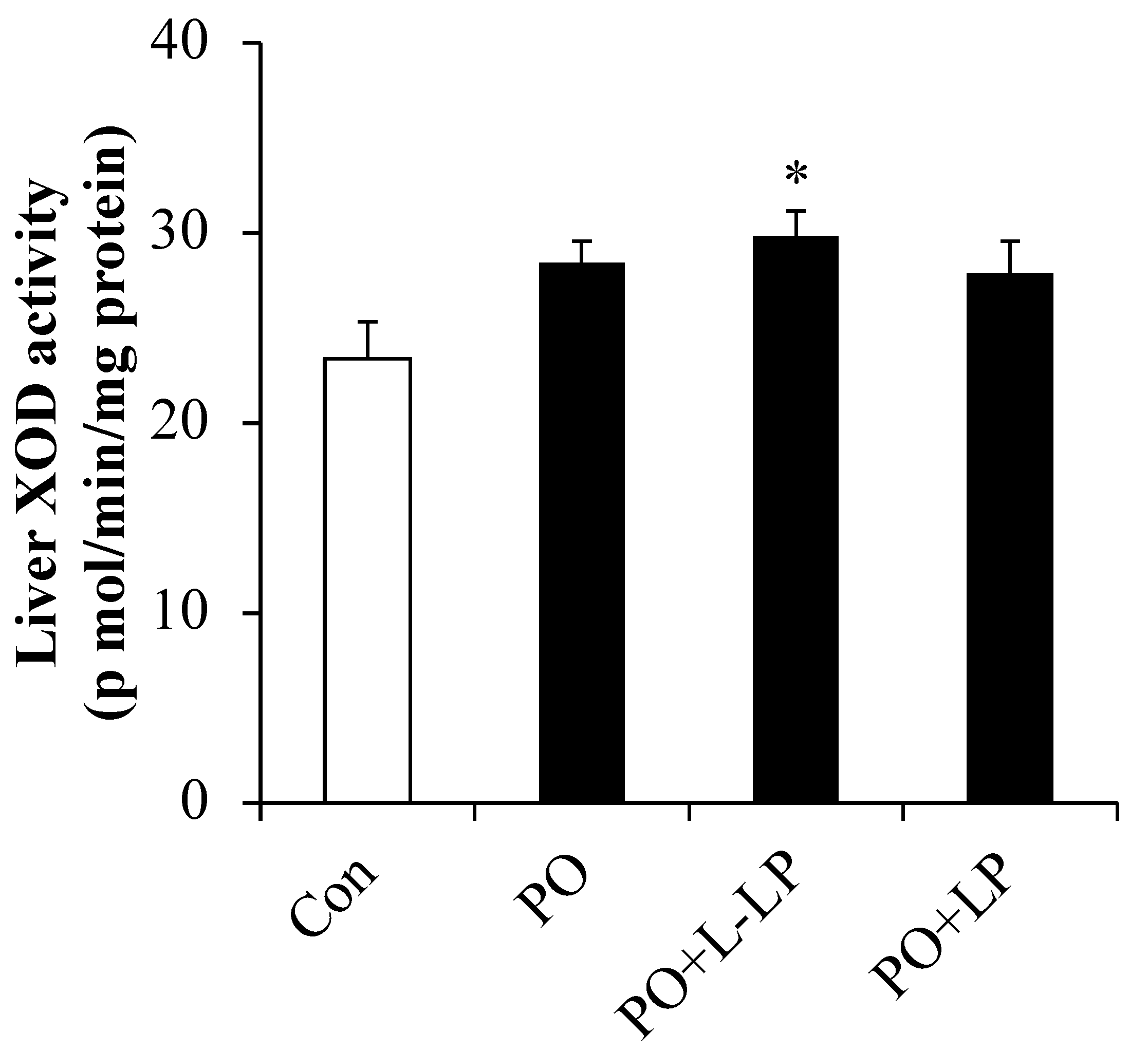
3.2. UA and Liver XOD Activity
3.3. Intestinal UA Excretion Transporter Expression Level
3.4. UA Excretion Transporter Expression Level in Kidney
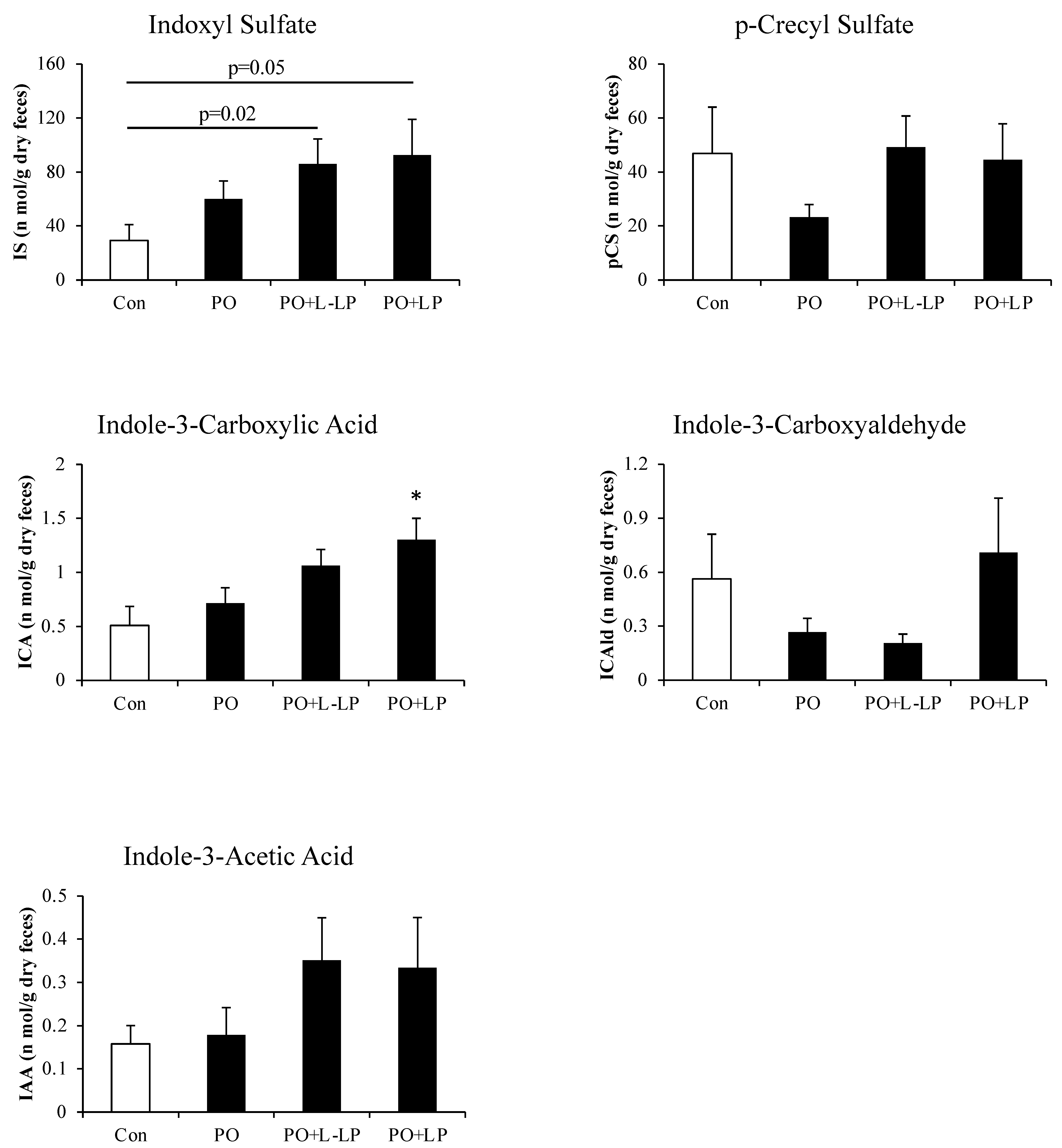
3.5. Fecal Excretion of Protein-Derived Gut Microbiota Metabolites
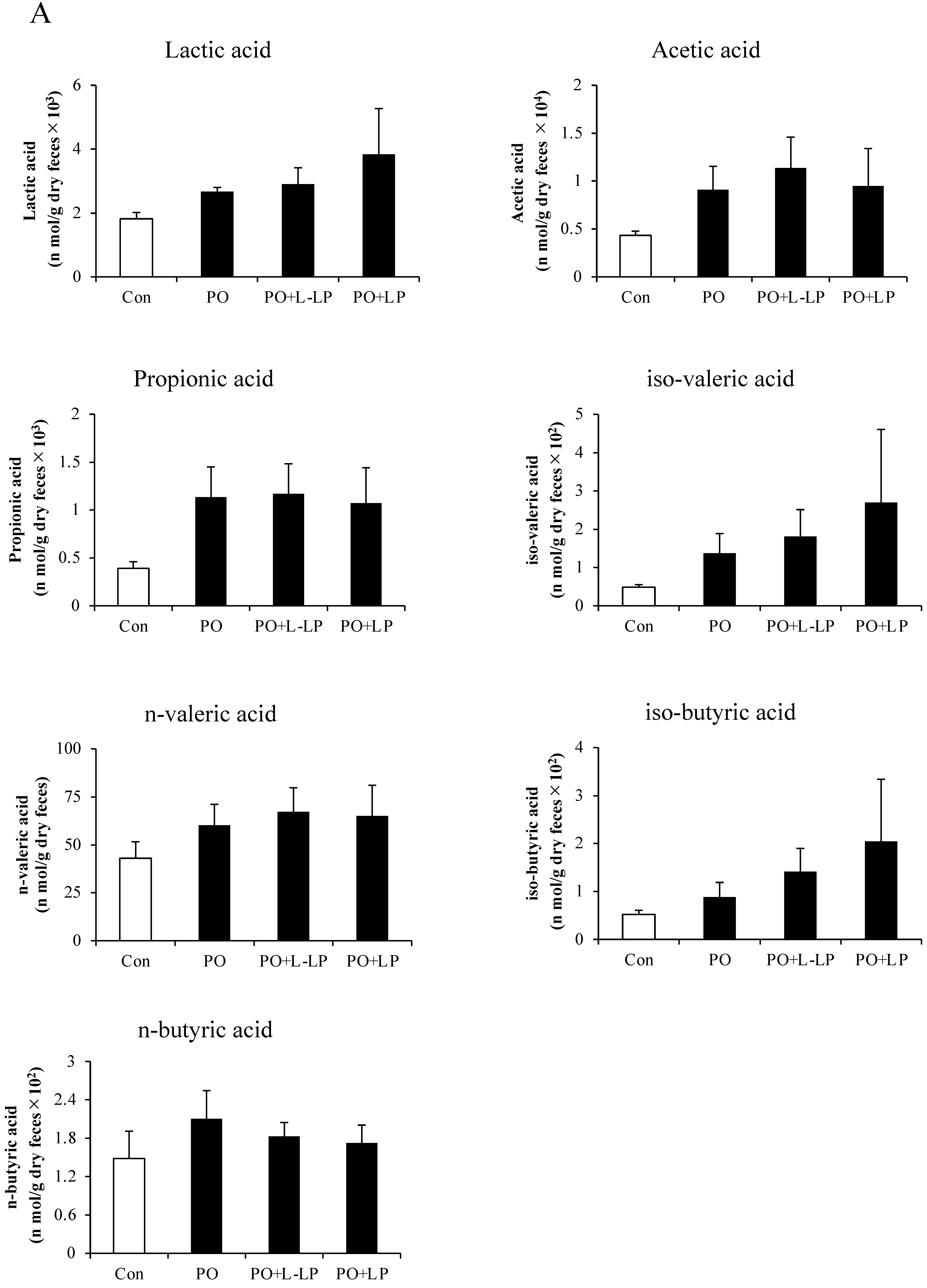
3.6. Fecal and Cecal SCFAs Content
3.7. Correlation between Fecal UA Levels and Fecal Indoxyl Sulphate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, S.; Yang, H.; Guo, Y.; Wei, F.; Yang, X.; Li, D.; Li, M.; Xu, W.; Li, W.; Sun, L.; et al. Comparative efficacy and safety of urate-lowering therapy for the treatment of hyperuricemia: A systematic review and network meta-analysis. Sci. Rep. 2016, 6, 33082. [Google Scholar] [CrossRef]
- Stamp, L.K.; Chapman, P.T.; Palmer, S.C. Allopurinol and kidney function: An update. Joint Bone Spine 2016, 83, 19–24. [Google Scholar] [CrossRef] [PubMed]
- White, W.B.; Saag, K.G.; Becker, M.A.; Borer, J.S.; Gorelick, P.B.; Whelton, A.; Hunt, B.; Castillo, M.; Gunawardhana, L. Cardiovascular Safety of Febuxostat or Allopurinol in Patients with Gout. N. Engl. J. Med. 2018, 378, 1200–1210. [Google Scholar] [CrossRef]
- Stamp, L.K.; Barclay, M.L. How to prevent allopurinol hypersensitivity reactions? Rheumatology 2018, 57 (Suppl. S1), i35–i41. [Google Scholar] [CrossRef] [PubMed]
- Ichida, K.; Matsuo, H.; Takada, T.; Nakayama, A.; Murakami, K.; Shimizu, T.; Yamanashi, Y.; Kasuga, H.; Nakashima, H.; Nakamura, T.; et al. Decreased extra-renal urate excretion is a common cause of hyperuricemia. Nat. Commun. 2012, 3, 764. [Google Scholar] [CrossRef]
- Mao, Q. BCRP/ABCG2 in the placenta: Expression, function and regulation. Pharm. Res. 2008, 25, 1244–1255. [Google Scholar] [CrossRef]
- Matsuo, H.; Takada, T.; Ichida, K.; Nakamura, T.; Nakayama, A.; Ikebuchi, Y.; Ito, K.; Kusanagi, Y.; Chiba, T.; Tadokoro, S.; et al. Common defects of ABCG2, a high-capacity urate exporter, cause gout: A function-based genetic analysis in a Japanese population. Sci. Transl. Med. 2009, 1, 5ra11. [Google Scholar] [CrossRef] [PubMed]
- Takada, T.; Yamamoto, T.; Matsuo, H.; Tan, J.K.; Ooyama, K.; Sakiyama, M.; Miyata, H.; Yamanashi, Y.; Toyoda, Y.; Higashino, T.; et al. Identification of ABCG2 as an Exporter of Uremic Toxin Indoxyl Sulfate in Mice and as a Crucial Factor Influencing CKD Progression. Sci. Rep. 2018, 8, 11147. [Google Scholar] [CrossRef]
- Wang, Z.; Song, L.; Li, X.; Xiao, Y.; Huang, Y.; Zhang, Y.; Li, J.; Li, M.; Ren, Z. Lactiplantibacillus pentosus P2020 protects the hyperuricemia and renal inflammation in mice. Front. Nutr. 2023, 10, 1094483. [Google Scholar] [CrossRef]
- Takeda, S.; Yamasaki, K.; Takeshita, M.; Kikuchi, Y.; Tsend-Ayush, C.; Dashnyam, B.; Ahhmed, A.M.; Kawahara, S.; Muguruma, M. The investigation of probiotic potential of lactic acid bacteria isolated from traditional Mongolian dairy products. Anim. Sci. J. 2011, 82, 571–579. [Google Scholar] [CrossRef]
- Tanaka, A.; Kanmura, S.; Morinaga, Y.; Kawabata, K.; Arima, S.; Sasaki, F.; Nasu, Y.; Tanoue, S.; Hashimoto, S.; Takeshita, M.; et al. Oral administration of Lactobacillus plantarum 06CC2 prevents experimental colitis in mice via an anti inflammatory response. Mol. Med. Rep. 2020, 21, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, A.; Mavel, S.; Nadal-Desbarats, L.; Galineau, L.; Attucci, S.; Dufour, D.; Sokol, H.; Emond, P. Validation of a global quantitative analysis methodology of tryptophan metabolites in mice using LC-MS. Talanta 2019, 195, 593–598. [Google Scholar] [CrossRef]
- Yamasaki, M.; Minesaki, M.; Iwakiri, A.; Miyamoto, Y.; Ogawa, K.; Nishiyama, K.; Tsend-Ayush, C.; Oyunsuren, T.; Li, Y.; Nakano, T.; et al. Lactobacillus plantarum 06CC2 reduces hepatic cholesterol levels and modulates bile acid deconjugation in Balb/c mice fed a high-cholesterol diet. Food Sci. Nutr. 2020, 8, 6164–6173. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Miyamoto, Y.; Ogawa, K.; Nishiyama, K.; Tsend-Ayush, C.; Li, Y.; Matsusaki, T.; Nakano, T.; Takeshita, M.; Arima, Y. Lactiplantibacillus plantarum 06CC2 upregulates intestinal ZO-1 protein and bile acid metabolism in Balb/c mice fed high-fat diet. Biosci. Microbiota Food Health 2024, 43, 13–22. [Google Scholar] [CrossRef]
- De Oliveira, E.P.; Burini, R.C. High plasma uric acid concentration: Causes and consequences. Diabetol. Metab. Syndr. 2012, 4, 12. [Google Scholar] [CrossRef]
- Yin, H.; Liu, N.; Chen, J. The role of the intestine in the development of hyperuricemia. Front. Immunol. 2022, 13, 845684. [Google Scholar] [CrossRef]
- Chen, R.J.; Chen, M.H.; Chen, Y.L.; Hsiao, C.M.; Chen, H.M.; Chen, S.J.; Wu, M.D.; Yech, Y.J.; Yuan, G.W.; Wang, Y.J. Evaluating the urate-lowering effects of different microbial fermented extracts in hyperuricemic models accompanied with a safety study. J. Food Drug Anal. 2016, 25, 597–606. [Google Scholar] [CrossRef]
- Lee, Y.; Werlinger, P.; Suh, J.W.; Cheng, J. Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms 2022, 10, 851. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, X.; Zhao, H.; Yang, E.; Wang, Y.; Cheng, N.; Cao, W. Impact of Camellia japonica bee pollen polyphenols on hyperuricemia and gut microbiota in potassium Oxonate-induced mice of the article. Nutrients 2021, 13, 2665. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Zhong, H.; Chen, F.; Regenstein, J.; Hu, X.; Cai, L.; Feng, F. The gut microbiota as a target to control hyperuricemia pathogenesis: Potential mechanisms and therapeutic strategies of the article. Crit. Rev. Food Sci. Nutr. 2022, 62, 3979–3989. [Google Scholar] [CrossRef]
- Chien, C.Y.; Chien, Y.J.; Lin, Y.H.; Lin, Y.H.; Chan, S.T.; Hu, W.C.; Wu, H.F.; Chiang, C.F.; Hsu, C.L. Supplementation of Lactobacillus plantarum (TCI227) Prevented Potassium-Oxonate-Induced Hyperuricemia in Rats. Nutrients 2022, 14, 4832. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, H.; Wang, R.; Yu, Y.; Liu, X.; Tian, Z. Protective effect of sodium butyrate on intestinal barrier damage and uric acid reduction in hyperuricemia mice. Biomed. Pharmacother. 2023, 161, 114568. [Google Scholar] [CrossRef]
- Xie, Q.S.; Zhang, J.X.; Liu, M.; Liu, P.H.; Wang, Z.J.; Zhu, L.; Jiang, L.; Jin, M.M.; Liu, X.N.; Liu, L.; et al. Short-chain fatty acids exert opposite effects on the expression and function of p-glycoprotein and breast cancer resistance protein in rat intestine. Acta Pharmacol. 2021, 42, 470–481. [Google Scholar] [CrossRef]
- Yantong, Z.; Xiaofei, P.; Guanghui, L. An update on the animal models in hyperuricaemia research. Clin. Exp. Rheumatol. 2017, 35, 860–864. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g) | Con | PO | PO + L-LP | PO + LP |
|---|---|---|---|---|
| Casein | 200 | 200 | 200 | 200 |
| Corn starch | 397.5 | 362.5 | 362.3 | 361.5 |
| α-Corn starch | 132 | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 | 100 |
| Soybean oil | 70 | 70 | 70 | 70 |
| Cellulose | 50 | 50 | 50 | 50 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 | 2.5 |
| L-cystine | 3 | 3 | 3 | 3 |
| Mineral mix | 35 | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 | 10 |
| tert-Butylhydroquinone | 0.014 | 0.014 | 0.014 | 0.014 |
| RNA from yeast | 0 | 10 | 10 | 10 |
| Potassium oxonate | 0 | 25 | 25 | 25 |
| LP06CC2 | 0 | 0 | 0.2 | 1 |
| Total | 1000.014 | 1000.014 | 1000.014 | 1000.014 |
| Con | PO | PO + L-LP | PO + LP | |
|---|---|---|---|---|
| Final body weight (g) | 24.1 ± 0.39 | 23.5 ± 0.25 | 23.3 ± 0.56 | 23.2 ± 0.45 |
| Food intake (g/day) | 3.11 ± 0.09 | 3.20 ± 0.11 | 3.40 ± 0.17 | 3.01 ± 0.07 |
| Urine volume (mL/day) | 1.01 ± 0.06 | 1.17 ± 0.11 | 0.81 ± 0.11 | 0.91 ± 0.05 |
| (mg/g Body Weight) | Con | PO | PO + L-LP | PO + LP |
|---|---|---|---|---|
| Liver | 47.2 ± 0.8 | 47.5 ± 0.9 | 46.9 ± 1.2 | 46.3 ± 0.6 |
| Epididymal fat | 14.0 ± 0.8 | 11.9 ± 0.8 | 11.9 ± 1.7 | 13.0 ± 0.9 |
| Small intestine | 19.8 ± 1.2 | 29.6 ± 1.3 | 29.1 ± 1.5 | 24.7 ± 1.9 |
| Large intestine | 6.3 ± 0.2 | 6.6 ± 0.2 | 6.9 ± 0.2 | 6.3 ± 0.3 |
| Kidney | 18.0 ± 0.3 | 18.2 ± 0.5 | 18.0 ± 0.6 | 17.2 ± 0.2 |
| Perinephric fat | 2.7 ± 0.2 | 2.4 ± 0.2 | 2.2 ± 0.2 | 2.4 ± 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nei, S.; Matsusaki, T.; Kawakubo, H.; Ogawa, K.; Nishiyama, K.; Tsend-Ayush, C.; Nakano, T.; Takeshita, M.; Shinyama, T.; Yamasaki, M. Lactiplantibacillus plantarum 06CC2 Enhanced the Expression of Intestinal Uric Acid Excretion Transporter in Mice. Nutrients 2024, 16, 3042. https://doi.org/10.3390/nu16173042
Nei S, Matsusaki T, Kawakubo H, Ogawa K, Nishiyama K, Tsend-Ayush C, Nakano T, Takeshita M, Shinyama T, Yamasaki M. Lactiplantibacillus plantarum 06CC2 Enhanced the Expression of Intestinal Uric Acid Excretion Transporter in Mice. Nutrients. 2024; 16(17):3042. https://doi.org/10.3390/nu16173042
Chicago/Turabian StyleNei, Shunsuke, Tatsuya Matsusaki, Hibiki Kawakubo, Kenjirou Ogawa, Kazuo Nishiyama, Chuluunbat Tsend-Ayush, Tomoki Nakano, Masahiko Takeshita, Takuo Shinyama, and Masao Yamasaki. 2024. "Lactiplantibacillus plantarum 06CC2 Enhanced the Expression of Intestinal Uric Acid Excretion Transporter in Mice" Nutrients 16, no. 17: 3042. https://doi.org/10.3390/nu16173042
APA StyleNei, S., Matsusaki, T., Kawakubo, H., Ogawa, K., Nishiyama, K., Tsend-Ayush, C., Nakano, T., Takeshita, M., Shinyama, T., & Yamasaki, M. (2024). Lactiplantibacillus plantarum 06CC2 Enhanced the Expression of Intestinal Uric Acid Excretion Transporter in Mice. Nutrients, 16(17), 3042. https://doi.org/10.3390/nu16173042