

Mechanistic Understanding of D-Glucaric Acid to Support Liver Detoxification Essential to Muscle Health Using a Computational Systems Biology Approach

Abstract
1. Introduction
2. Materials and Methods
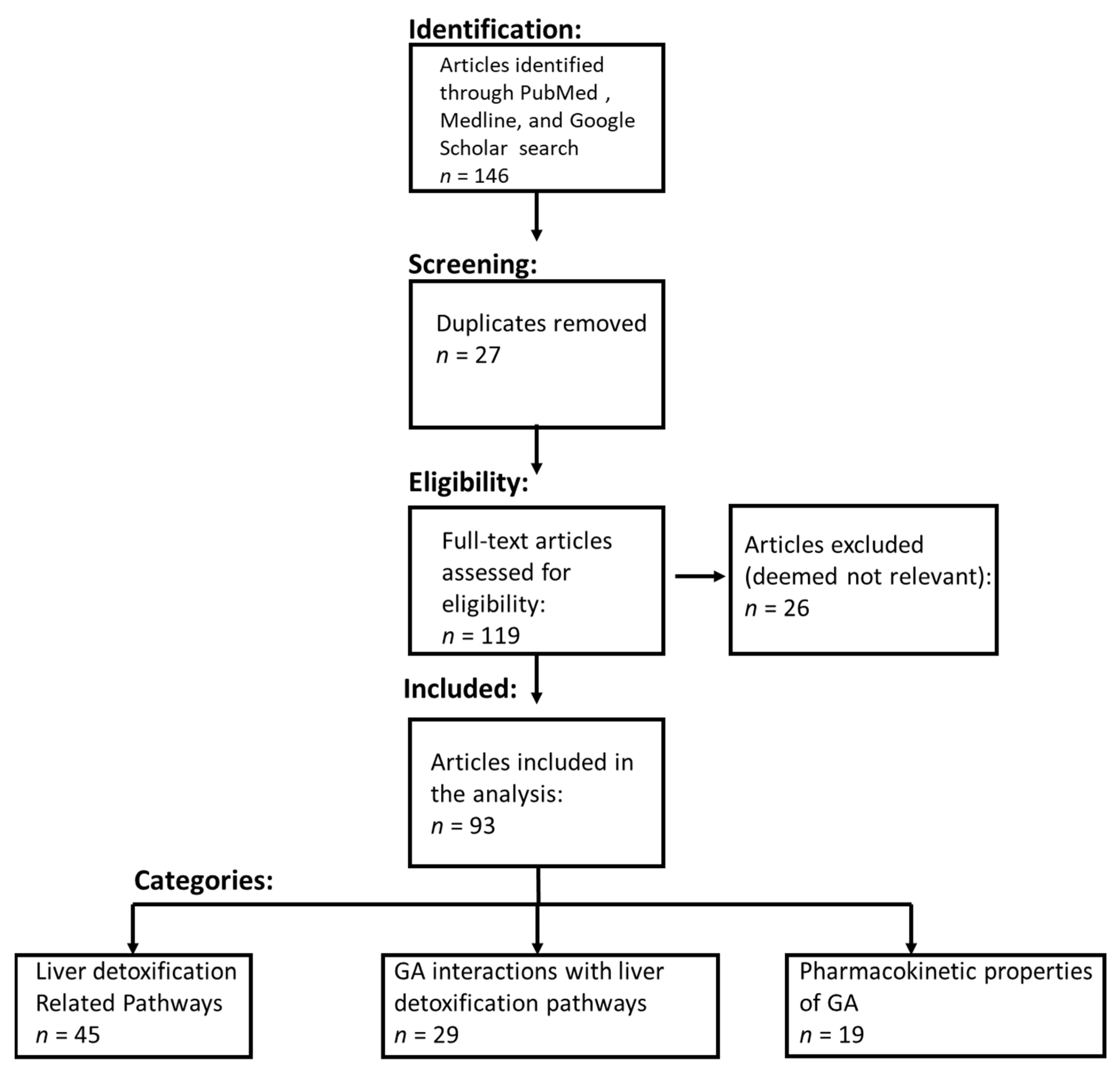
2.1. Systematic Literature Review Process and Inclusion Criteria
2.2. CytoSolve in Silico Modeling Protocol
2.2.1. Control Conditions
2.2.2. Computer Simulations to Study the Effect of GA on Integrated Model of Liver Detoxification
- Effect of GA on ROS levels
- Effect of GA on glucuronide deconjugate levels
- Effect of GA on C-PARP levels
- Effect of GA on β-glucuronidase synthesis
3. Results
3.1. Systematic Literature Review
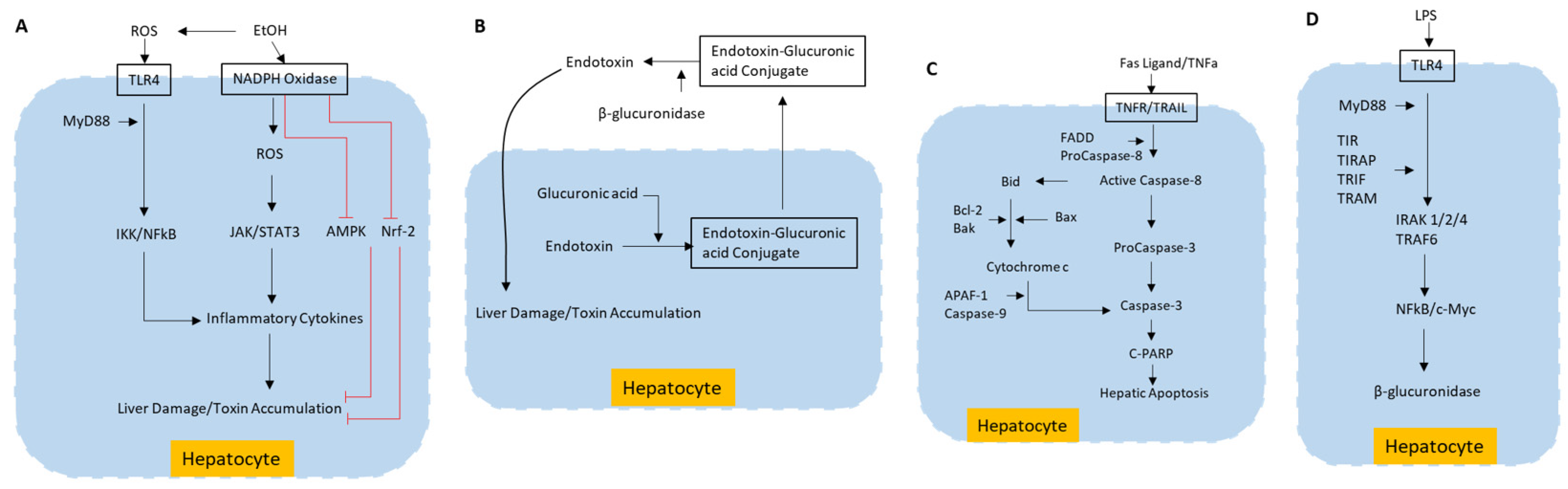
3.2. Molecular Pathways Involved in Liver Toxicity
3.2.1. ROS Production Pathway Involved in Liver Toxicity
3.2.2. Deconjugation/Deglucuronidation Pathway in Liver Toxicity
3.2.3. Hepatic Apoptosis Pathway in Liver Toxicity
3.2.4. β-Glucuronidase Synthesis Pathway in Liver Toxicity
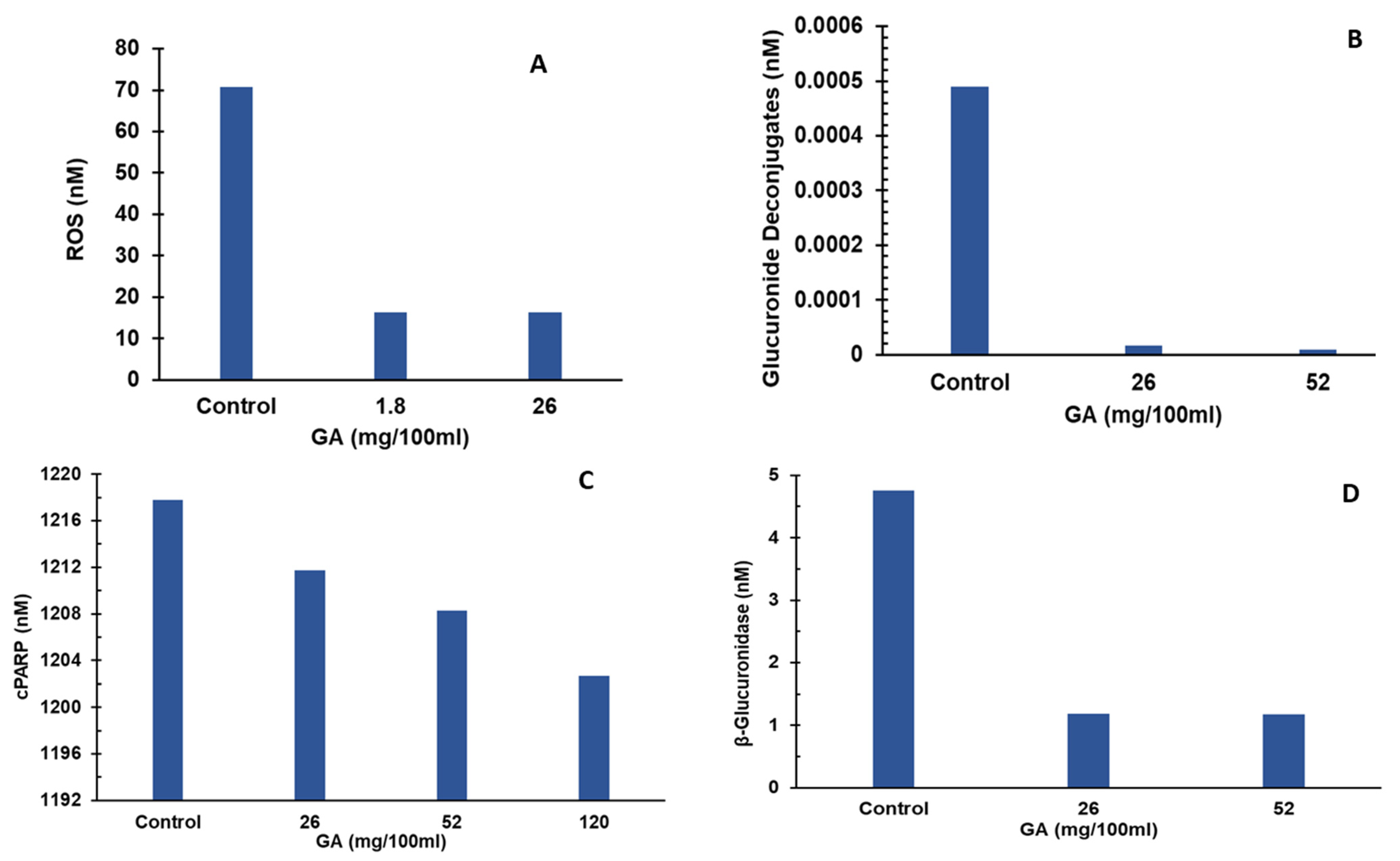
3.3. Simulation Results
3.3.1. Effect of Glucaric Acid on ROS Production
3.3.2. Effect of Glucaric Acid on Deconjugation/Deglucuronidation
3.3.3. Effect of GA on Hepatic Apoptosis
3.3.4. Effect of GA on β-Glucuronidase Synthesis
4. Discussion
5. Conclusions and Future Work
5.1. Conclusions
5.2. Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smart, N.A.; King, N.; McFarlane, J.R.; Graham, P.L.; Dieberg, G. Effect of exercise training on liver function in adults who are overweight or exhibit fatty liver disease: A systematic review and meta-analysis. Br. J. Sports Med. 2018, 52, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Van Der Windt, D.J.; Sud, V.; Zhang, H.; Tsung, A.; Huang, H. The Effects of Physical Exercise on Fatty Liver Disease. Gene Expr. 2018, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Mañas, L. Skeletal Muscle Regulates Metabolism via Interorgan Crosstalk: Roles in Health and Disease. J. Am. Med. Dir. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Laitselart, P.; Derely, J.; Daban, J.L.; De Rudnicki, S.; Libert, N. Relationship between creatine kinase and liver enzymes in war wounded with rhabdomyolysis. Injury 2022, 53, 166–170. [Google Scholar] [CrossRef]
- Lash, T.; Kraemer, R.R. Elevated liver enzymes indicating a diagnosis of limb-girdle muscular dystrophy. J. Gen. Intern. Med. 2014, 29, 813–815. [Google Scholar] [CrossRef][Green Version]
- Mastali, V.P.; Hoseini, R.; Azizi, M. The short-term effect of vitamin D supplementation on the response to muscle and liver damages indices by exhaustive aerobic exercise in untrained men: A quasi-experimental study. BMC Sport. Sci. Med. Rehabil. 2022, 14, 7. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Ono, A.; Yoshida, M.; Matsumoto, K.; Saito, M. Toxicological significance of increased serum alkaline phosphatase activity in dog studies of pesticides: Analysis of toxicological data evaluated in Japan. Regul. Toxicol. Pharmacol. 2019, 109, 104482. [Google Scholar] [CrossRef]
- De Bandt, J.P.; Jegatheesan, P.; Tennoune-El-Hafaia, N. Muscle Loss in Chronic Liver Diseases: The Example of Nonalcoholic Liver Disease. Nutrients 2018, 10, 1195. [Google Scholar] [CrossRef]
- Huang, C.C.; Lee, M.C.; Ho, C.S.; Hsu, Y.J.; Ho, C.C.; Kan, N.W. Protective and Recovery Effects of Resveratrol Supplementation on Exercise Performance and Muscle Damage following Acute Plyometric Exercise. Nutrients 2021, 13, 3217. [Google Scholar] [CrossRef]
- Abdulkadir, M.; Londono, D.; Gordon, D.; Fernandez, T.V.; Brown, L.W.; Cheon, K.A.; Coffey, B.J.; Elzerman, L.; Fremer, C.; Fründt, O.; et al. Investigation of previously implicated genetic variants in chronic tic disorders: A transmission disequilibrium test approach. Eur. Arch. Psychiatry Clin. Neurosci. 2018, 268, 301–316. [Google Scholar] [CrossRef]
- Zoltaszek, R.; Kowalczyk, P.; Kowalczyk, M.C.; Hanausek, M.; Kilianska, Z.M.; Slaga, T.J.; Walaszek, Z. Dietary D-glucarate effects on the biomarkers of inflammation during early post-initiation stages of benzo[a]pyrene-induced lung tumorigenesis in A/J mice. Oncol. Lett. 2011, 2, 145–154. [Google Scholar]
- Bhattacharya, S.; Gachhui, R.; Sil, P.C. The prophylactic role of d-saccharic acid-1,4-lactone against hyperglycemia-induced hepatic apoptosis via inhibition of both extrinsic and intrinsic pathways in diabetic rats. Food Funct. 2013, 4, 283–296. [Google Scholar] [CrossRef]
- Yao, D.; Dong, Q.; Tian, Y.; Dai, C.; Wu, S. Lipopolysaccharide stimulates endogenous β-glucuronidase via PKC/NF-κB/c-myc signaling cascade: A possible factor in hepatolithiasis formation. Mol. Cell. Biochem. 2018, 444, 93–102. [Google Scholar] [CrossRef]
- Cline, J.C. Nutritional aspects of detoxification in clinical practice. Altern. Ther. Health Med. 2015, 21, 54–63. [Google Scholar]
- He, Y.; Ma, J.; Fan, X.; Ding, L.; Ding, X.; Zhang, Q.Y.; Lin, G. The key role of gut-liver axis in pyrrolizidine alkaloid-induced hepatotoxicity and enterotoxicity. Acta Pharm. Sin. B 2021, 11, 3820–3835. [Google Scholar] [CrossRef]
- Pandit, A.; Sachdeva, T.; Bafna, P. Drug-induced hepatotoxicity: A review. J. Appl. Pharm. Sci. 2012, 2, 233–243. [Google Scholar] [CrossRef]
- Yang, L.; Seki, E. Toll-like receptors in liver fibrosis: Cellular crosstalk and mechanisms. Front. Physiol. 2012, 3, 138. [Google Scholar] [CrossRef]
- Galicia-Moreno, M.; Gutiérrez-Reyes, G. The role of oxidative stress in the development of alcoholic liver disease. Rev. Gastroenterol. Mex. 2014, 79, 135–144. [Google Scholar] [CrossRef]
- Hodges, R.E.; Minich, D.M. Modulation of Metabolic Detoxification Pathways Using Foods and Food-Derived Components: A Scientific Review with Clinical Application. J. Nutr. Metab. 2015, 2015, 760689. [Google Scholar] [CrossRef]
- Perez, J.L.; Jayaprakasha, G.K.; Yoo, K.S.; Patil, B.S. Development of a method for the quantification of D-glucaric acid in different varieties of grapefruits by high-performance liquid chromatography and mass spectra. J. Chromatogr. A 2008, 1190, 394–397. [Google Scholar] [CrossRef]
- Xie, B.; Liu, Y.; Zou, H.; Son, Y.; Wang, H.; Wang, H.; Shao, J. Determination of D-glucaric acid and/or D-glucaro-1,4-lacton in different apple varieties through hydrophilic interaction chromatography. Food Chem. 2016, 203, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ayyadurai, V.A.S.; Dewey, C.F. CytoSolve: A Scalable Computational Method for Dynamic Integration of Multiple Molecular Pathway Models. Cell. Mol. Bioeng. 2011, 4, 28–45. [Google Scholar] [CrossRef]
- Koo, A.; Nordsletten, D.; Umeton, R.; Yankama, B.; Ayyadurai, S.; García-Cardeña, G.; Dewey, C.F. In Silico Modeling of Shear-Stress-Induced Nitric Oxide Production in Endothelial Cells through Systems Biology. Biophys. J. 2013, 104, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Oketch-Rabah, H.A.; Hardy, M.L.; Patton, A.P.; Chung, M.; Sarma, N.D.; Yoe, C.; Ayyadurai, V.A.S.; Fox, M.A.; Jordan, S.A.; Mwamburi, M.; et al. Multi-Criteria Decision Analysis Model for Assessing the Risk from Multi-Ingredient Dietary Supplements (MIDS). J. Diet. Suppl. 2021, 18, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Al-Lazikani, B.; Banerji, U.; Workman, P. Combinatorial drug therapy for cancer in the post-genomic era. Nat. Biotechnol. 2012, 30, 679–692. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the neurovascular unit: Key functions and signaling pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P.; McLure, K.G.; Sakamoto, K.M. Molecular Systems Architecture of Interactome in the Acute Myeloid Leukemia Microenvironment. Cancers 2022, 14, 756. [Google Scholar] [CrossRef]
- Nordsletten, D.A.; Yankama, B.; Umeton, R.; Ayyadurai, V.V.S.; Dewey, C.F. Multiscale Mathematical Modeling to Support Drug Development. IEEE Trans. Biomed. Eng. 2011, 58, 3508–3512. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P. Bioactive compounds in green tea may improve transplant tolerance: A computational systems biology analysis. Clin. Nutr. ESPEN 2021, 46, 439–452. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P.; Bannuru, R.R. Attenuation of low-grade chronic inflammation by phytonutrients: A computational systems biology analysis. Clin. Nutr. ESPEN 2022, 49, 425–435. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P. In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients. Appl. Sci. 2022, 12, 10013. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339, 332–336. [Google Scholar] [CrossRef]
- Muthumani, M.; Prabu, S.M. Silibinin potentially attenuates arsenic-induced oxidative stress mediated cardiotoxicity and dyslipidemia in rats. Cardiovasc. Toxicol. 2014, 14, 83–97. [Google Scholar] [CrossRef]
- Harte, A.L.; da Silva, N.F.; Creely, S.J.; McGee, K.C.; Billyard, T.; Youssef-Elabd, E.M.; Tripathi, G.; Ashour, E.; Abdalla, M.S.; Sharada, H.M.; et al. Elevated endotoxin levels in non-alcoholic fatty liver disease. J. Inflamm. 2010, 7, 15. [Google Scholar] [CrossRef]
- Yang, Y.L.; Li, J.P.; Xu, X.P.; Dou, K.F.; Yue, S.Q.; Li, K.Z. Protective effects of tumor necrosis factor alpha antibody and ulinastatin on liver ischemic reperfusion in rats. World J. Gastroenterol. 2004, 10, 3161–3164. [Google Scholar] [CrossRef]
- Walaszek, Z.; Szemraj, J.; Narog, M.; Adams, A.K.; Kilgore, J.; Sherman, U.; Hanausek, M. Metabolism, uptake, and excretion of a D-glucaric acid salt and its potential use in cancer prevention. Cancer Detect. Prev. 1997, 21, 178–190. [Google Scholar]
- Liu, Y.; Wen, P.H.; Zhang, X.X.; Dai, Y.; He, Q. Breviscapine ameliorates CCl4-induced liver injury in mice through inhibiting inflammatory apoptotic response and ROS generation. Int. J. Mol. Med. 2018, 42, 755–768. [Google Scholar] [CrossRef]
- Kurup, R.K.; Kurup, P.A. Hypothalamic digoxin and hemispheric chemical dominance: Relation to speech and language dysfunction. Int. J. Neurosci. 2003, 113, 797–814. [Google Scholar] [CrossRef]
- Liao, Y.J.; Wang, Y.H.; Wu, C.Y.; Hsu, F.Y.; Chien, C.Y.; Lee, Y.C. Ketogenic Diet Enhances the Cholesterol Accumulation in Liver and Augments the Severity of CCl 4 and TAA-Induced Liver Fibrosis in Mice. Int. J. Mol. Sci. 2021, 22, 2934. [Google Scholar] [CrossRef]
- G. Bardallo, R.; Panisello-Roselló, A.; Sanchez-Nuno, S.; Alva, N.; Roselló-Catafau, J.; Carbonell, T. Nrf2 and oxidative stress in liver ischemia/reperfusion injury. FEBS J. 2021, 289, 5463–5479. [Google Scholar] [CrossRef]
- Hanausek, M.; Walaszek, Z.; Slaga, T.J. Detoxifying Cancer Causing Agents to Prevent Cancer. Integr. Cancer Ther. 2003, 2, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Peyrol, J.; Meyer, G.; Obert, P.; Dangles, O.; Pechère, L.; Amiot, M.J.; Riva, C. Involvement of bilitranslocase and beta-glucuronidase in the vascular protection by hydroxytyrosol and its glucuronide metabolites in oxidative stress conditions. J. Nutr. Biochem. 2018, 51, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Mandrekar, P.; Ambade, A. Cellular Signaling Pathways in Alcoholic Liver Disease. In Trends in Alcoholic Liver Disease Research—Clinical and Scientific Aspects; IntechOpen: London, UK, 2012. [Google Scholar]
- Wang, K. Molecular mechanisms of hepatic apoptosis. Cell Death Dis. 2014, 5, e996. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Gores, G.J. Death receptor-mediated apoptosis and the liver. J. Hepatol. 2002, 37, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Rampal, G.; Khanna, N.; Thind, T.S.; Arora, S.; Vig, A.P. Role of isothiocyanates as anticancer agents and their contributing molecular and cellular mechanisms. Med. Chem. Drug Discov. 2012, 3, 79–93. [Google Scholar]
- Loegering, D.J.; Lennartz, M.R. Protein kinase C and toll-like receptor signaling. Enzyme Res. 2011, 2011, 537821. [Google Scholar] [CrossRef]
- Walaszek, Z. Potential use of d-glucaric acid derivatives in cancer prevention. Cancer Lett. 1990, 54, 1–8. [Google Scholar] [CrossRef]
- Lampe, J.W.; Li, S.S.; Potter, J.D.; King, I.B. Serum β-Glucuronidase Activity Is Inversely Associated with Plant-Food Intakes in Humans. J. Nutr. 2002, 132, 1341–1344. [Google Scholar] [CrossRef]
- Subramanian, G.; Madras, G. Introducing saccharic acid as an efficient iron chelate to enhance photo-Fenton degradation of organic contaminants. Water Res. 2016, 104, 168–177. [Google Scholar] [CrossRef]
- Wang, K.; Gan, X.; Tang, X.; Wang, S.; Tan, H. Determination of d-saccharic acid-1,4-lactone from brewed kombucha broth by high-performance capillary electrophoresis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 371–374. [Google Scholar] [CrossRef]
- Jaeschke, H.; Gores, G.J.; Cederbaum, A.I.; Hinson, J.A.; Pessayre, D.; Lemasters, J.J. Mechanisms of Hepatotoxicity. Toxicol. Sci. 2002, 65, 166–176. [Google Scholar] [CrossRef]
- Food and Drug Administration Center for Drug Evaluation and Research. Request for Determination of Exempt Status of Investigational New Drug Application (IND) for Cyto-001 as Treatment for Patients with Pancreatic Cancer (PIND: 118833); Food and Drug Administration Center for Drug Evaluation and Research: Silver Spring, MD, USA, 2013.
- Ayyadurai, V.A.S.; Deonikar, P. Do GMOs Accumulate Formaldehyde and Disrupt Molecular Systems Equilibria? Systems Biology May Provide Answers. Agric. Sci. 2015, 6, 630–662. [Google Scholar] [CrossRef]
- Kothandaram, S.; Deonikar, P.; Mohan, M.; Venugopal, V.; Ayyadurai, V.A.S. In Silico Modeling of C1 Metabolism. Am. J. Plant Sci. 2015, 6, 1444–1465. [Google Scholar] [CrossRef]
- Mohan, M.; Kothandaram, S.; Venugopal, V.; Deonikar, P.; Ayyadurai, V.A.S. Integrative Modeling of Oxidative Stress and C1 Metabolism Reveals Upregulation of Formaldehyde and Downregulation of Glutathione. Am. J. Plant Sci. 2015, 6, 1527–1542. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Hansen, M.; Fagan, J.; Deonikar, P. In-Silico Analysis & In-Vivo Results Concur on Glutathione Depletion in Glyphosate Resistant GMO Soy, Advancing a Systems Biology Framework for Safety Assessment of GMOs. Am. J. Plant Sci. 2016, 7, 1571–1589. [Google Scholar]
- Thanh, V.H.; Zunino, R.; Priami, C. Efficient finite-difference method for computing sensitivities of biochemical reactions. Proc. R. Soc. A Math. Phys. Eng. Sci. 2018, 474, 20180303. [Google Scholar] [CrossRef]
- Ainscough, J.S.; Gerberick, G.F.; Zahedi-Nejad, M.; Lopez-Castejon, G.; Brough, D.; Kimber, I.; Dearman, R.J. Dendritic cell IL-1α and IL-1β are polyubiquitinated and degraded by the proteasome. J. Biol. Chem. 2014, 289, 35582–35592. [Google Scholar] [CrossRef]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes (Reprinted from Molecular Mechanisms in Spermatogenesis, 2007). Oxidative Med. Cell. Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef]
- Albeck, J.G.; Burke, J.M.; Spencer, S.L.; Lauffenburger, D.A.; Sorger, P.K. Modeling a Snap-Action, Variable-Delay Switch Controlling Extrinsic Cell Death. PLoS Biol. 2008, 6, 2831–2852. [Google Scholar] [CrossRef]
- Ambrosio, G.; Zweier, J.L.; Jacobus, W.E.; Weisfeldt, M.L.; Flaherty, J.T. Improvement of postischemic myocardial function and metabolism induced by administration of deferoxamine at the time of reflow: The role of iron in the pathogenesis of reperfusion injury. Circulation 1987, 76, 906–915. [Google Scholar] [CrossRef]
- Van Arsdell, S.W.; Murphy, K.P.; Pazmany, C.; Erickson, D.; Burns, C.; Moody, M.D. Xplore® mRNA Assays for the Quantification of IL-1 β and TNF- α mRNA in Lipopolysaccharide- Induced Mouse Macrophages. BioTechniques 2000, 28, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Atunes, F.; Salvador, A.; Marinho, H.S.; Alves, R.; Pinto, R.E. Lipid Peroxidation in Mitochondrial inner membranes. An integrative kinetic model. Free. Radic. Biol. Med. 1996, 21, 917–943. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y.; Jones, D.P. Direct determination of UDP-glucuronic acid in cell extracts by high-performance liquid chromatography. Anal. Biochem. 1982, 127, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.; Kuru, K. Purification and Partial Characterization of Catalase from Chicken Erythrocytes and the Effect of Various Inhibitors on Enzyme Activity. Turk. J. Chem. 2003, 27, 85–97. [Google Scholar]
- Babbs, C.F.; Steiner, M.G. Simulation of free radical reactions in biology and medicine: A new two-compartment kinetic model of intracellular lipid peroxidation. Free. Radic. Biol. Med. 1990, 8, 471–485. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Chatterjee, S.; Manna, P.; Das, J.; Ghosh, J.; Gachhui, R.; Sil, P.C. Prophylactic role of D-saccharic acid-1,4-lactone in tertiary butyl hydroperoxide induced cytotoxicity and cell death of murine hepatocytes via mitochondria-dependent pathways. J. Biochem. Mol. Toxicol. 2011, 25, 341–354. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Manna, P.; Gachhui, R.; Sil, P.C. D-Saccharic acid 1, 4-lactone protects diabetic rat kidney by ameliorating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via NF-κB and PKC signaling. Toxicol. Appl. Pharmacol. 2013, 267, 16–29. [Google Scholar] [CrossRef]
- Boase, S.; Miners, J.O. In vitro–in vivo correlations for drugs eliminated by glucuronidation: Investigations with the model substrate zidovudine. Br. J. Clin. Pharmacol. 2002, 54, 493–503. [Google Scholar] [CrossRef]
- Bracey, L.T.; Paigen, K. Changes in translational yield regulate tissue-specific expression of β-glucuronidase. Proc. Natl. Acad. Sci. USA 1987, 84, 9020–9024. [Google Scholar] [CrossRef]
- Buettner, G.R.; Ng, C.F.; Wang, M.; Rodgers, V.G.J.; Schafer, F.Q. A new paradigm: Manganese superoxide dismutase influences the production of H2O2 in cells and thereby their biological state. Free. Radic. Biol. Med. 2006, 41, 1338–1350. [Google Scholar] [CrossRef]
- Dargemont, C.; Kuhn, L.C. Export of mRNA from Microinjected Nuclei of Xenopus laevis Oocytes. J. Cell Biol. 1992, 118, 1–9. [Google Scholar] [CrossRef]
- Dwivedi, C.; Heck, W.J.; Downie, A.A.; Larroya, S.; Webb, T.E. Effect of calcium glucarate on β-glucuronidase activity and glucarate content of certain vegetables and fruits. Biochem. Med. Metab. Biol. 1990, 43, 83–92. [Google Scholar] [CrossRef]
- Edwards, A.; Cao, C.; Pallone, T.L. Cellular mechanisms underlying nitric oxide-induced vasodilation of descending vasa recta. American Journal of Physiology. Ren. Physiol. 2011, 300, F441–F456. [Google Scholar] [CrossRef]
- Feng, S.; Song, J.D. Determination of β-glucuronidase in human colorectal carcinoma cell lines. World J. Gastroenterol. 1997, 3, 251–252. [Google Scholar] [CrossRef][Green Version]
- Gutiérrez, J.; St Laurent, G.; Urcuqui-Inchima, S. Propagation of Kinetic Uncertainties through a Canonical Topology of the TLR4 Signaling Network in Different Regions of Biochemical Reaction Space. Theor. Biol. Med. Model. 2010, 7, 7. [Google Scholar] [CrossRef]
- Stillinger, F.H. Proton Transfer Reactions and Kinetics in Water. In Theoretical Chemistry, Advances and Perspedives; Elsevier: Amsterdam, The Netherlands, 1978; pp. 178–234. [Google Scholar]
- Henle, E.S.; Luo, Y.; Linn, S. Fe2+, Fe3+, and Oxygen React with DNA-Derived Radicals Formed during iron mediated Fenton reactions. Biochemistry 1996, 35, 12212–12219. [Google Scholar] [CrossRef]
- Hoffmann, A.; Levchenko, A.; Scott, M.L.; Baltimore, D. The IkappaB-NF-kappaB signaling module: Temporal control and selective gene activation. Science 2002, 298, 1241–1245. [Google Scholar] [CrossRef]
- Kavida, M. Mathematical and Computational Models of Oxidative and Nitrosative Stress. Crit. Rev. Biomed. Eng. 2011, 39, 461–472. [Google Scholar] [CrossRef]
- Li, C.P.; Li, J.H.; He, S.Y.; Li, P.; Zhong, X.L. Roles of Fas/Fasl, Bcl-2/Bax, and Caspase-8 in rat nonalcoholic fatty liver disease pathogenesis. Genet. Mol. Res. 2014, 13, 3991–3999. [Google Scholar] [CrossRef]
- Li, J.; Ke, W.; Zhou, Q.; Wu, Y.; Luo, H.; Zhou, H.; Yang, B.; Guo, Y.; Zheng, Q.; Zhang, Y. Tumour necrosis factor-α promotes liver ischaemia-reperfusion injury through the PGC-1α/Mfn2 pathway. J. Cell. Mol. Med. 2014, 18, 1863–1873. [Google Scholar] [CrossRef]
- Macfarlane, N.G.; Miller, D.J. Depression of Peak Force Without Altering Calcium Sensitivit by the Superoxide Anion in Chemically Skinned Cardiac Muscle of Rat. Circ. Res. 1992, 70, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.W.; Ladd, M.A.; Fitzsimmons, P.N. Measurement of Kinetic Parameters for Biotransformation of Polycyclic Aromatic Hydrocarbons by Trout Liver S9 Fractions: Implications for Bioaccumulation Assessment. Appl. In Vitro Toxicol. 2017, 4, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Nolan, J.P.; Hare, D.K.; McDevitt, J.J.; Vilayat Ali, M. In Vitro Studies of Intestinal Endotoxin Absorption: I. Kinetics of absorption in the isolated everted gut sac. Gastroenterology 1977, 72, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Schliemann, M.; Bullinger, E.; Borchers, S.; Allgöwer, F.; Findeisen, R.; Scheurich, P. Heterogeneity reduces sensitivity of cell death for TNF-Stimuli. BMC Syst. Biol. 2011, 5, 204. [Google Scholar] [CrossRef] [PubMed]
- Sharp, G.C.; Ma, H.; Saunders PT, K.; Norman, J.E. A Computational Model of Lipopolysaccharide-Induced Nuclear Factor Kappa B Activation: A Key Signalling Pathway in Infection-Induced Preterm Labour. PLoS ONE 2013, 8, e70180. [Google Scholar] [CrossRef]
- Shin, H.J.; Lee, H.; Park, J.D.; Hyun, H.C.; Sohn, H.O.; Lee, D.W.; Kim, Y.S. Molecules and Kinetics of Binding of LPS to Recombinant CD14, TLR4, and MD-2 Proteins. Mol. Cells 2007, 24, 119–124. [Google Scholar]
- Taylor, D.J.; Styles, P.; Matthews, P.M.; Gadian, D.G. Energetics of Human Muscle: Exercise-Induced ATP Depletion. Magn. Reson. Med. 1986, 3, 44–54. [Google Scholar] [CrossRef]
- Tomasic, J.; Keglevic, D. The kinetics of hydrolysis of synthetic glucuronic esters and glucuronic ethers by bovine liver and Escherichia coli b-glucuronidase. Biochem. J. 1973, 133, 789–795. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Watson, G.; Daveyl, R.A.; Labarca, C.; Paigen, K. Genetic determination of kinetic parameters in β-glucuronidase induction by androgen. J. Biol. Chem. 1981, 256, 3005–3011. [Google Scholar] [CrossRef]
- Xue, C.; Chou, C.-S.; Kao, C.-Y.; Sen, C.K.; Friedman, A. Propagation of Cutaneous Thermal Injury: A Mathematical Model. Wound Repair Regener. 2012, 20, 114–122. [Google Scholar] [CrossRef]
- Xue, L.; He, J.; Gao, N.; Lu, X.; Li, M.; Wu, X.; Liu, Z.; Jin, Y.; Liu, J.; Xu, J.; et al. Probiotics may delay the progression of nonalcoholic fatty liver disease by restoring the gut microbiota structure and improving intestinal endotoxemia. Sci. Rep. 2017, 7, 45176. [Google Scholar] [CrossRef]
- Yang, H.; Hreggvidsdottir, H.S.; Palmblad, K.; Wang, H.; Ochani, M.; Li, J.; Lu, B.; Chavan, S.; Rosas-Ballina, M.; Al-Abed, Y.; et al. A Critical Cysteine Is Required for HMGB1 Binding to Toll-like Receptor 4 and Activation of Macrophage Cytokine Release. Proc. Natl. Acad. Sci. USA 2010, 107, 11942–11947. [Google Scholar] [CrossRef]
- Yoza, B.K.; Wells, J.D.; McCall, C.E. Interleukin-1β Expression after Inhibition of Protein Phosphatases in Endotoxin-Tolerant Cells. Clin. Vaccine Immunol. 1998, 5, 281–287. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucaric Acid Dose (mg/100 mL) | Serum Concentration (µM) |
|---|---|
| 1.8 | 2.7 |
| 26 | 39 |
| 52 | 78 |
| 120 | 180 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayyadurai, V.A.S.; Deonikar, P.; Fields, C. Mechanistic Understanding of D-Glucaric Acid to Support Liver Detoxification Essential to Muscle Health Using a Computational Systems Biology Approach. Nutrients 2023, 15, 733. https://doi.org/10.3390/nu15030733
Ayyadurai VAS, Deonikar P, Fields C. Mechanistic Understanding of D-Glucaric Acid to Support Liver Detoxification Essential to Muscle Health Using a Computational Systems Biology Approach. Nutrients. 2023; 15(3):733. https://doi.org/10.3390/nu15030733
Chicago/Turabian StyleAyyadurai, V. A. Shiva, Prabhakar Deonikar, and Christine Fields. 2023. "Mechanistic Understanding of D-Glucaric Acid to Support Liver Detoxification Essential to Muscle Health Using a Computational Systems Biology Approach" Nutrients 15, no. 3: 733. https://doi.org/10.3390/nu15030733
APA StyleAyyadurai, V. A. S., Deonikar, P., & Fields, C. (2023). Mechanistic Understanding of D-Glucaric Acid to Support Liver Detoxification Essential to Muscle Health Using a Computational Systems Biology Approach. Nutrients, 15(3), 733. https://doi.org/10.3390/nu15030733