
Aralia continentalis Root Enhances Non-Rapid Eye Movement Sleep by Activating GABAA Receptors

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of KS-126
2.3. High-Performance Liquid Chromatography Analysis
2.4. Animals
2.5. Pentobarbital-Induced Sleep Accelerated Test
2.6. EEG Surgery, Recording and Data Analysis
2.7. Patch-Clamp Electrophysiology
2.7.1. Primary Neuron Culture
2.7.2. Electrophysiology
2.8. Statistical Analysis
3. Results
3.1. Quantitative Analysis of CA and KA in KS-126
3.2. Hypnotic Effects of KS-126
3.3. Effects of KS-126 on Sleep Quantity and Sleep Quality of NREMS in C57BL/6N Mice
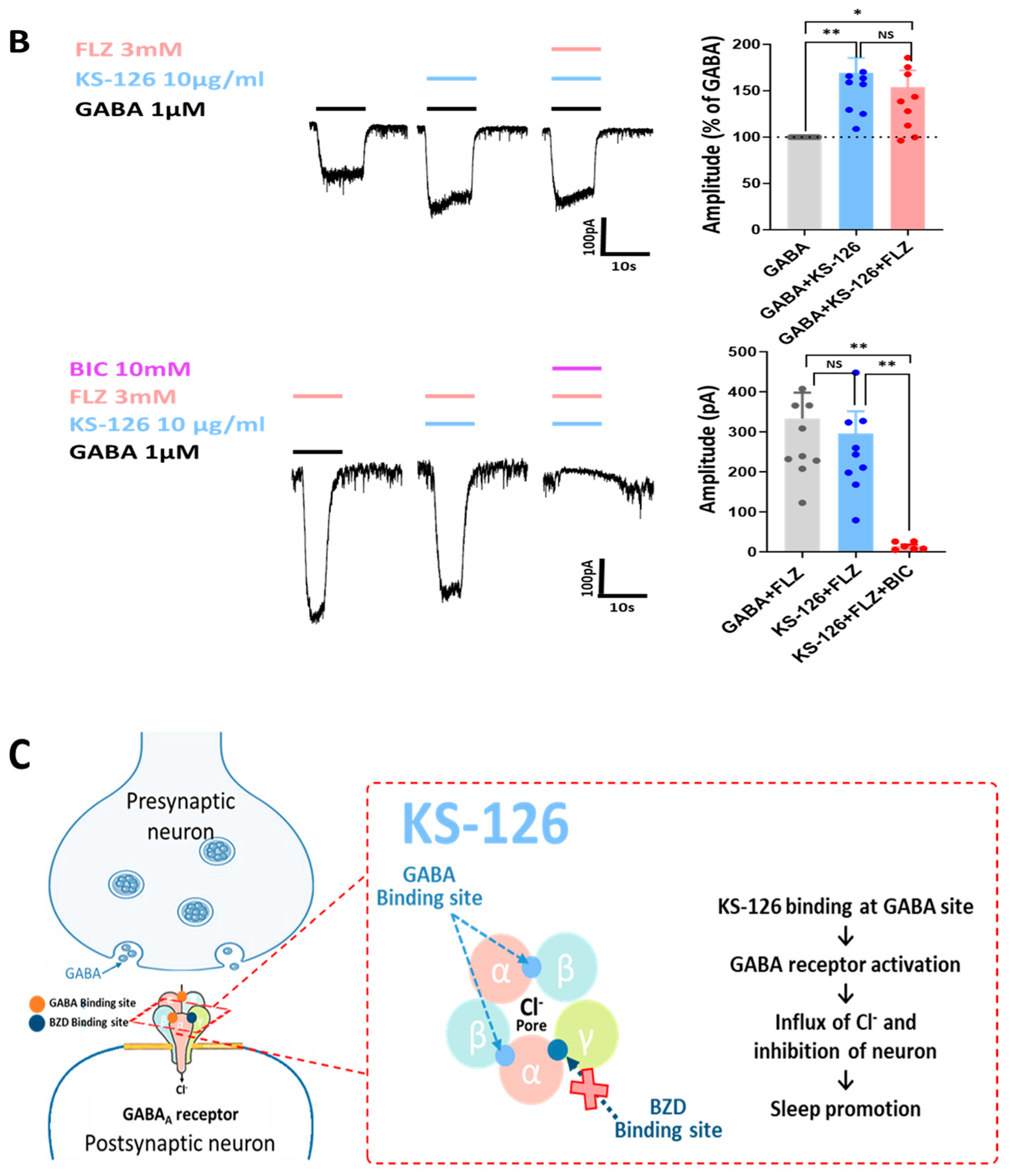
3.4. Possible Mechanism of Action for KS-126
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Behar, J.A. From sleep medicine to medicine during sleep: A new paradigm. Sleep 2020, 43, zsz279. [Google Scholar] [CrossRef]
- Ohayon, M.M. Epidemiology of insomnia: What we know and what we still need to learn. Sleep Med. Rev. 2002, 6, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Mendoza, J.; Vgontzas, A.N. Insomnia and its Impact on Physical and Mental Health. Curr. Psychiatry Rep. 2013, 15, 418. [Google Scholar] [CrossRef]
- Buscemi, N.; Vandermeer, B.; Friesen, C.; Bialy, L.; Tubman, M.; Ospina, M.; Klassen, T.P.; Witmans, M. The efficacy and safety of drug treatments for chronic insomnia in adults: A meta-analysis of RCTs. J. Gen. Intern. Med. 2007, 22, 1335–1350. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Aitchison, K.; Bateson, A.; Curran, H.V.; Davies, S.; Leonard, B.; Nutt, D.J.; Stephens, D.N.; Wilson, S. Benzodiazepines: Risks and benefits. A reconsideration. J. Psychopharmacol. 2013, 27, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ortuno, M.M.; Belanger, L.; Ivers, H.; LeBlanc, M.; Morin, C.M. The use of natural products for sleep: A common practice? Sleep Med. 2009, 10, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Um, M.Y.; Kim, J.Y.; Han, J.K.; Kim, J.; Yang, H.; Yoon, M.; Kim, J.; Kang, S.W.; Cho, S. Phlorotannin supplement decreases wake after sleep onset in adults with self-reported sleep disturbance: A randomized, controlled, double-blind clinical and polysomnographic study. Phytother. Res. 2018, 32, 698–704. [Google Scholar] [CrossRef]
- Jiang, J.G.; Huang, X.J.; Chen, J.; Lin, Q.S. Comparison of the sedative and hypnotic effects of flavonoids, saponins, and polysaccharides extracted from Semen Ziziphus jujube. Nat. Prod. Res. 2007, 21, 310–320. [Google Scholar] [CrossRef]
- Wang, M.Y.; Liu, J.L.; Sun, W.; Li, N.; Yu, Z.P.; Zhu, K.; Gao, J.Q.; Wang, C.M.; Sun, J.H.; Chen, J.G.; et al. Schisantherin A Exerts Sedative and Hypnotic Effects Through Regulating GABA and its Receptor in Mice and Rats. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef]
- Cho, S.; Yang, H.; Jeon, Y.J.; Lee, C.J.; Jin, Y.H.; Baek, N.I.; Kim, D.; Kang, S.M.; Yoon, M.; Yong, H.; et al. Phlorotannins of the edible brown seaweed Ecklonia cava Kjellman induce sleep via positive allosteric modulation of gamma-aminobutyric acid type A-benzodiazepine receptor: A novel neurological activity of seaweed polyphenols. Food Chem. 2012, 132, 1133–1142. [Google Scholar] [CrossRef]
- Cho, S.; Yoon, M.; Pae, A.N.; Jin, Y.H.; Cho, N.C.; Takata, Y.; Urade, Y.; Kim, S.; Kim, J.S.; Yang, H.; et al. Marine polyphenol phlorotannins promote non-rapid eye movement sleep in mice via the benzodiazepine site of the GABA(A) receptor. Psychopharmacology 2014, 231, 2825–2837. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.J.; Kwak, C.H.; Chung, T.W.; Park, S.J.; Cheeeei, M.; Park, S.S.; Seo, C.S.; Son, J.K.; Chang, Y.C.; Park, Y.G.; et al. Pimaric acid from Aralia cordata has an inhibitory effect on TNF-alpha-induced MMP-9 production and HASMC migration via down-regulated NF-kappa B and AP-1. Chem.-Biol. Interact. 2012, 199, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.D.; Lee, J.Y.; Cho, B.J.; Park, T.W.; Kim, C.J. The Analgesic and Anti-inflammatory Effects of 7-Oxosandaracopimaric Acid Isolated from the Roots of Aralia cordata. Arch. Pharm. Res. 2010, 33, 509–514. [Google Scholar] [CrossRef]
- Cho, S.O.; Ban, J.Y.; Kim, J.Y.; Ju, H.S.; Lee, I.S.; Song, K.S.; Bae, K.; Seong, Y.H. Anti-ischemic activities of Aralia cordata and its active component, oleanolic acid. Arch. Pharm. Res. 2009, 32, 923–932. [Google Scholar] [CrossRef]
- Park, D.S.; Huh, J.E.; Baek, Y.H. Therapeutic effect of Aralia cordata extracts on cartilage protection in collagenase-induced inflammatory arthritis rabbit model. J. Ethnopharmacol. 2009, 125, 207–217. [Google Scholar] [CrossRef]
- Baek, Y.H.; Huh, J.E.; Lee, J.D.; Choi, D.Y.; Park, D.S. Effect of Aralia cordata extracts on cartilage protection and apoptosis inhibition. Biol. Pharm. Bull. 2006, 29, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Khan, A.; Ali, J.; Ullah, H.; Khan, A.; Ali, H.; Irshad, N.; Khan, S. Attenuation of LPS-induced acute lung injury by continentalic acid in rodents through inhibition of inflammatory mediators correlates with increased Nrf2 protein expression. BMC Pharmacol. Toxicol. 2020, 21, 81. [Google Scholar] [CrossRef]
- Paiva, L.A.F.; Gurgel, L.A.; Silva, R.M.; Tome, A.R.; Gramosa, N.V.; Silveira, E.R.; Santos, F.A.; Rao, V.S.N. Anti-inflammatory effect of kaurenoic acid, a diterpene from Copaifera langsdorffii on acetic acid-induced colitis in rats. Vasc. Pharmacol. 2002, 39, 303–307. [Google Scholar] [CrossRef]
- Jeon, B.E.; Kwon, C.S.; Lee, J.E.; Moon, K.; Cha, J.; Park, I.; Koh, S.; Yoon, M.; Kim, S.W.; Kim, J.N. Anticancer Activity of Continentalic Acid in B-Cell Lymphoma. Molecules 2021, 26, 6845. [Google Scholar] [CrossRef]
- Yoo, G.; Lee, T.H.; Lee, J.H.; Kang, K.B.; Yang, H.J.; Park, Y.K.; Kim, S.Y.; Kim, S.H. Simultaneous determination of two diterpenoids, continentalic acid and kaurenoic acid, in the water extract of Aralia continentalis and their wound-healing activity. Pharmacogn. Mag. 2020, 16, 745–749. [Google Scholar] [CrossRef]
- Okoye, T.C.; Akah, P.A. Anticonvulsant Effects of Kaurenoic Acid Isolated from the Root Bark of Annona Senegalensis. Basic. Clin. Pharmacol. 2014, 115, 99. [Google Scholar]
- Kim, H.D.; Hong, K.B.; Noh, D.O.; Suh, H.J. Sleep-inducing effect of lettuce (Lactuca sativa) varieties on pentobarbital-induced sleep. Food Sci. Biotechnol. 2017, 26, 807–814. [Google Scholar] [CrossRef]
- Shah, V.K.; Choi, J.J.; Han, J.Y.; Lee, M.K.; Hong, J.T.; Oh, K.W. Pachymic Acid Enhances Pentobarbital-Induced Sleeping Behaviors via GABAA-ergic Systems in Mice. Biomol. Ther. 2014, 22, 314–320. [Google Scholar] [CrossRef]
- Kwon, Y.O.; Hong, J.T.; Oh, K.W. Rosmarinic Acid Potentiates Pentobarbital-Induced Sleep Behaviors and Non-Rapid Eye Movement (NREM) Sleep through the Activation of GABA(A)-ergic Systems. Biomol. Ther. 2017, 25, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Cheng, S.X.; Sun, H.T.; Ma, T.Z.; Zhang, S. Ferulic acid potentiates pentobarbital-induced sleep via the serotonergic system. Neurosci. Lett. 2012, 525, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Um, M.Y.; Yoon, M.; Lee, J.; Jung, J.; Cho, S. A Novel Potent Sleep-Promoting Effect of Turmeric: Turmeric Increases Non-Rapid Eye Movement Sleep in Mice Via Histamine H1Receptor Blockade. Mol. Nutr. Food Res. 2021, 65, e2100100. [Google Scholar] [CrossRef]
- Lee, B.; Hong, R.; Lim, P.; Cho, D.; Yeom, M.; Lee, S.; Kang, K.S.; Lee, S.C.; Shim, I.; Lee, H.; et al. The ethanolic extract of Aralia continentalis ameliorates cognitive deficits via modifications of BDNF expression and anti-inflammatory effects in a rat model of post-traumatic stress disorder. BMC Complement. Altern. Med. 2019, 19, 11. [Google Scholar] [CrossRef]
- Yang, D.K.; Lee, S.-J.; Adam, G.O.; Kim, S.-J. Aralia continentalis kitagawa extract attenuates the fatigue induced by exhaustive exercise through inhibition of oxidative stress. Antioxidants 2020, 9, 379. [Google Scholar] [CrossRef]
- Kang, S.K.; Ammanuel, S.; Thodupunuri, S.; Adler, D.A.; Johnston, M.V.; Kadam, S.D. Sleep dysfunction following neonatal ischemic seizures are differential by neonatal age of insult as determined by qEEG in a mouse model. Neurobiol. Dis. 2018, 116, 1–12. [Google Scholar] [CrossRef]
- Bush, B.J.; Donnay, C.; Andrews, E.A.; Lewis-Sanders, D.; Gray, C.L.; Qiao, Z.; Brager, A.J.; Johnson, H.; Brewer, H.C.S.; Sood, S.; et al. Non-rapid eye movement sleep determines resilience to social stress. Elife 2022, 11, e80206. [Google Scholar] [CrossRef]
- Kwon, S.; Jung, J.H.; Cho, S.; Moon, K.-D.; Lee, J. Dieckol is a natural positive allosteric modulator of GABAA-benzodiazepine receptors and enhances inhibitory synaptic activity in cultured neurons. Nutr. Neurosci. 2021, 24, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.J.; Page, A.T. Herbal medicine for insomnia: A systematic review and meta-analysis. Sleep Med. Rev. 2015, 24, 1–12. [Google Scholar] [CrossRef]
- Ghorbani, A.; Rakhshandeh, H.; Sadeghnia, H.R. Potentiating Effects of Lactuca sativa on Pentobarbital-Induced Sleep. Iran. J. Pharm. Res. 2013, 12, 401–406. [Google Scholar] [PubMed]
- Yoon, M.; Cho, S. Triphlorethol A, a Dietary Polyphenol from Seaweed, Decreases Sleep Latency and Increases Non-Rapid Eye Movement Sleep in Mice. Mar. Drugs 2018, 16, 139. [Google Scholar] [CrossRef] [PubMed]
- Masaki, M.; Aritake, K.; Tanaka, H.; Shoyama, Y.; Huang, Z.L.; Urade, Y. Crocin promotes non-rapid eye movement sleep in mice. Mol. Nutr. Food Res. 2012, 56, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Winsky-Sommerer, R. Role of GABAA receptors in the physiology and pharmacology of sleep. Eur. J. Neurosci. 2009, 29, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Lancel, M.; Faulhaber, J.; Schiffelholz, T.; Mathias, S.; Deisz, R.A. Muscimol and midazolam do not potentiate each other’s effects on sleep EEG in the rat. J. Neurophysiol. 1997, 77, 1624–1629. [Google Scholar] [CrossRef]
- Tobler, I.; Kopp, C.; Deboer, T.; Rudolph, U. Diazepam-induced changes in sleep: Role of the alpha 1 GABA(A) receptor subtype. Proc. Natl. Acad. Sci. USA 2001, 98, 6464–6469. [Google Scholar] [CrossRef]
- Kopp, C.; Rudolph, U.; Tobler, I. Sleep EEG changes after zolpidem in mice. Neuroreport 2004, 15, 2299–2302. [Google Scholar] [CrossRef]
- Kopp, C.; Rudolph, U.; Low, K.; Tobler, I. Modulation of rhythmic brain activity by diazepam: GABA(A) receptor subtype and state specificity. Proc. Natl. Acad. Sci. USA 2004, 101, 3674–3679. [Google Scholar] [CrossRef]
- van Lier, H.; Drinkenburg, W.H.; van Eeten, Y.J.; Coenen, A.M. Effects of diazepam and zolpidem on EEG beta frequencies are behavior-specific in rats. Neuropharmacology 2004, 47, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, C.; Dordal, A.; Aixendri, R.; Guzman, A.; Hamon, M.; Adrien, J. Sleep-stabilizing effects of E-6199, compared to zopiclone, zolpidem and THIP in mice. Sleep 2008, 31, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Xu, X.H.; Qu, W.M.; Lazarus, M.; Urade, Y.; Huang, Z.L. A mouse model mimicking human first night effect for the evaluation of hypnotics. Pharmacol. Biochem. Behav. 2014, 116, 129–136. [Google Scholar] [CrossRef]
- Luo, Y.; Balle, T. GABAA receptors as targets for anaesthetics and analgesics and promising candidates to help treat coronavirus infections: A mini-review. Basic. Clin. Pharmacol. Toxicol. 2022, 131, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, W. Structure and pharmacology of gamma-aminobutyric acidA receptor subtypes. Pharmacol. Rev. 1995, 47, 181–234. [Google Scholar] [PubMed]
- Aguayo, L.G.; Peoples, R.W.; Yeh, H.H.; Yevenes, G.E. GABA-A receptors as molecular sites of ethanol action. Direct or indirect actions? Curr. Top. Med. Chem. 2002, 2, 869–885. [Google Scholar] [CrossRef]
- Förstera, B.; Castro, P.A.; Moraga-Cid, G.; Aguayo, L.G. Potentiation of gamma aminobutyric acid receptors (GABAAR) by ethanol: How are inhibitory receptors affected? Front. Cell. Neurosci. 2016, 10, 114. [Google Scholar] [CrossRef]
- Cutler, A.J.; Mattingly, G.W.; Maletic, V. Understanding the mechanism of action and clinical effects of neuroactive steroids and GABAergic compounds in major depressive disorder. Transl. Psychiatry 2023, 13, 228. [Google Scholar] [CrossRef]
- Erman, M.K. Therapeutic options in the treatment of insomnia. J. Clin. Psychiatry 2005, 66 (Suppl. 9), 18–23, quiz 42–13. [Google Scholar]
- Stephenson, F.A. The GABAA receptors. Biochem. J. 1995, 310 Pt 1, 1–9. [Google Scholar] [CrossRef]
- Anaclet, C.; Zhang, M.; Zhao, C.; Buda, C.; Seugnet, L.; Lin, J.S. Effects of GF-015535-00, a novel alpha1 GABA A receptor ligand, on the sleep-wake cycle in mice, with reference to zolpidem. Sleep 2012, 35, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Johnston, G.A. GABA(A) receptor channel pharmacology. Curr. Pharm. Des. 2005, 11, 1867–1885. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.I.; Ong, J. GABA agonists and antagonists. Med. Res. Rev. 1992, 12, 593–636. [Google Scholar] [CrossRef] [PubMed]
- Khawaled, R.; Bruening-Wright, A.; Adelman, J.P.; Maylie, J. Bicuculline block of small-conductance calcium-activated potassium channels. Pflügers Arch. 1999, 438, 314–321. [Google Scholar] [CrossRef]
- Sharma, R.; Nakamura, M.; Neupane, C.; Jeon, B.H.; Shin, H.; Melnick, S.M.; Glenn, K.J.; Jang, I.S.; Park, J.B. Positive allosteric modulation of GABA(A) receptors by a novel antiepileptic drug cenobamate. Eur. J. Pharmacol. 2020, 879, 173117. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, M.; Lim, D.W.; Jung, J.; Jung, Y.S.; Lee, C.; Um, M.Y. Aralia continentalis Root Enhances Non-Rapid Eye Movement Sleep by Activating GABAA Receptors. Nutrients 2023, 15, 5020. https://doi.org/10.3390/nu15245020
Yoon M, Lim DW, Jung J, Jung YS, Lee C, Um MY. Aralia continentalis Root Enhances Non-Rapid Eye Movement Sleep by Activating GABAA Receptors. Nutrients. 2023; 15(24):5020. https://doi.org/10.3390/nu15245020
Chicago/Turabian StyleYoon, Minseok, Dong Wook Lim, Jonghoon Jung, Young Sung Jung, Changho Lee, and Min Young Um. 2023. "Aralia continentalis Root Enhances Non-Rapid Eye Movement Sleep by Activating GABAA Receptors" Nutrients 15, no. 24: 5020. https://doi.org/10.3390/nu15245020
APA StyleYoon, M., Lim, D. W., Jung, J., Jung, Y. S., Lee, C., & Um, M. Y. (2023). Aralia continentalis Root Enhances Non-Rapid Eye Movement Sleep by Activating GABAA Receptors. Nutrients, 15(24), 5020. https://doi.org/10.3390/nu15245020