
Properties of Dietary Flavone Glycosides, Aglycones, and Metabolites on the Catalysis of Human Endoplasmic Reticulum Uridine Diphosphate Glucuronosyltransferase 2B7 (UGT2B7)


Highlights
- Dietary flavone O-glycosides compounds have obvious inhibitory effects on the activity of uridine diphosphate glucuronosyltransferases (UGTs).
- Flavone O-glycosides generally have a stronger effect on UGT2B7 activity than their phenolic acid metabolites.
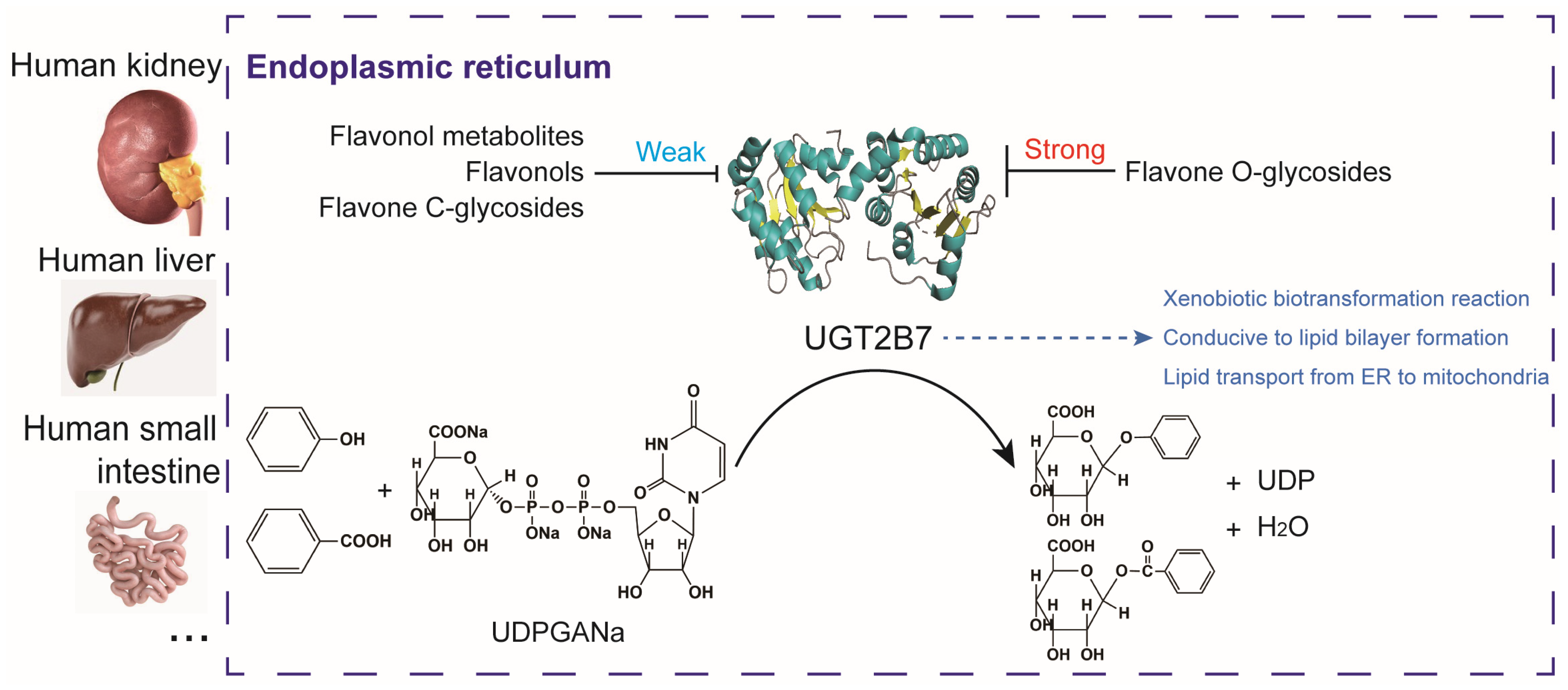
- The main finding identified a differential site for the absorption and utilization of flavone O-glycosides and flavone C-glycosides located in the endoplasmic reticulum.
- The main finding demonstrated the potential application of flavone glycosides, their aglycones and metabolites in regulating the synthesis of lipid-linked precursors and lipid transport.
Abstract
:1. Introduction
2. Methods
2.1. Chemicals
2.2. Gene Annotation Analysis of Uridine Diphosphate Glucuronosyltransferase 2B7
2.3. Molecular Docking to Explain the Interaction of Flavone Glycosides, Phenolic Acids Metabolites, and Iridoids towards Uridine Diphosphate Glucuronosyltransferase 2B7 Activities
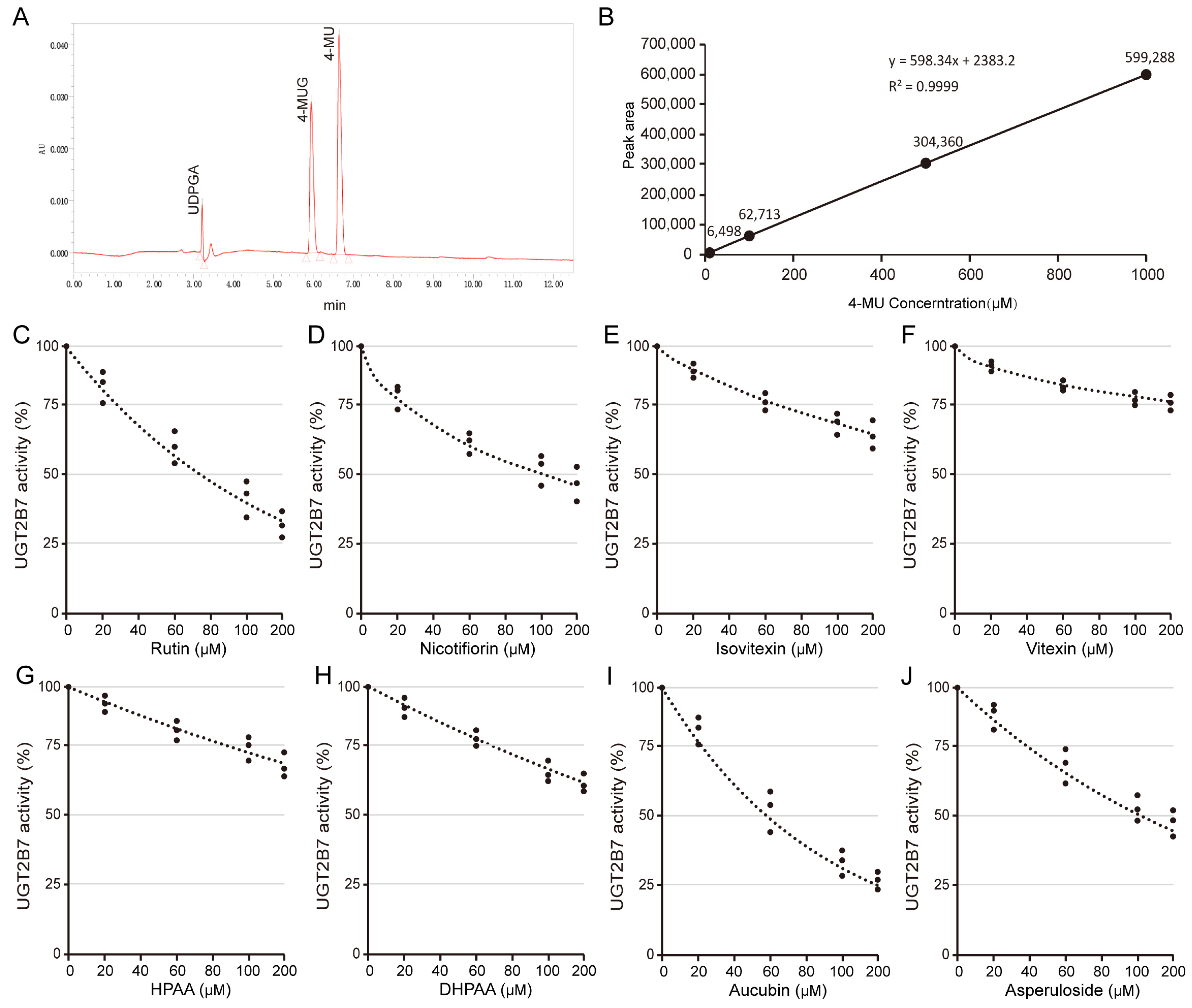
2.4. HPLC Assessment of Inhibitory Effects of Flavones against Human Uridine Diphosphate Glucuronosyltransferase 2B7
2.5. Cell Culture and Treatment
2.6. Transcription Analysis of Flavones on Uuridine Diphosphate Glucuronosyltransferase Protein Expression
2.7. Statistical Analysis
3. Results
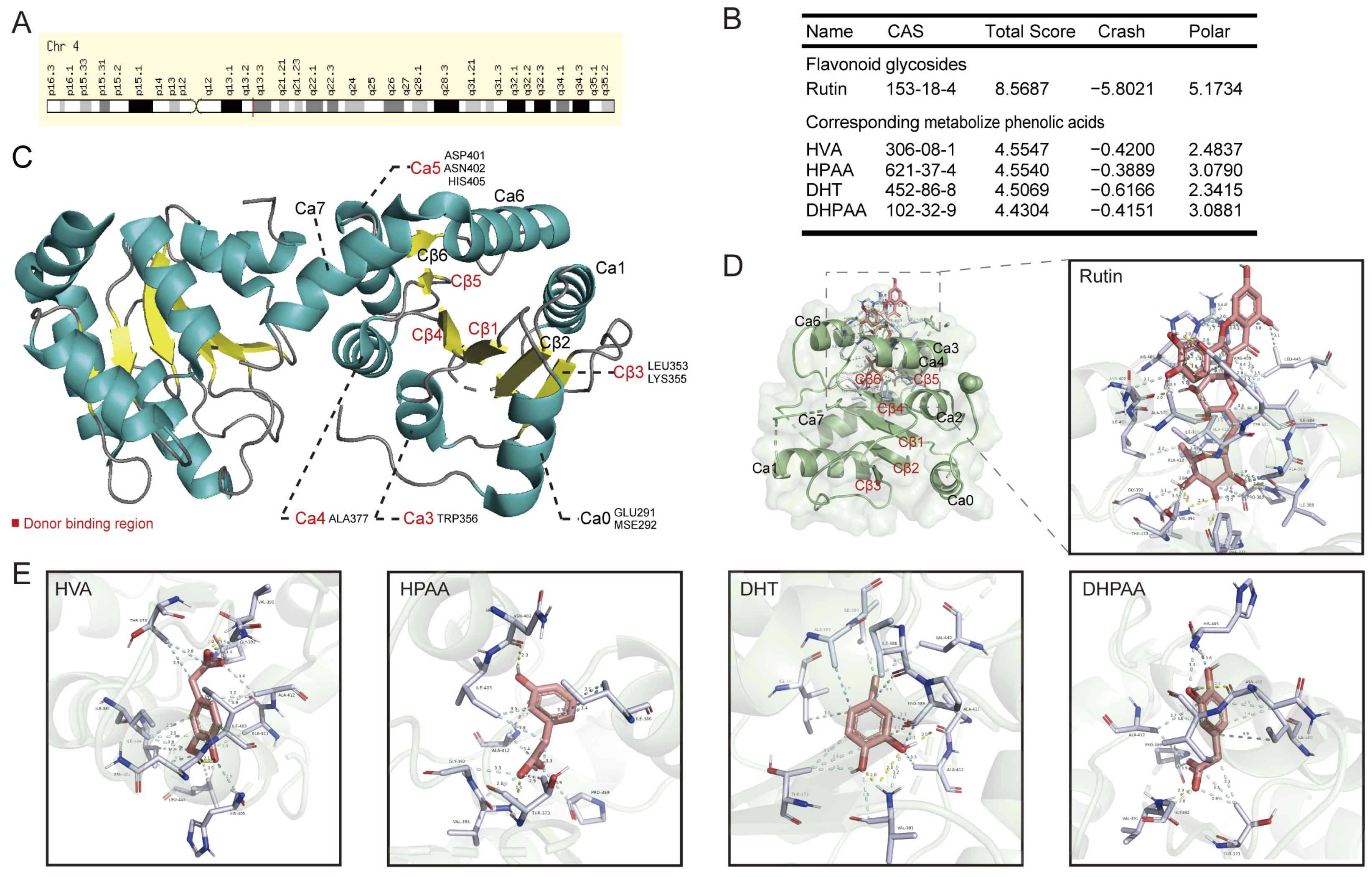
3.1. Overall Function of the Human Uridine Diphosphate Glucuronosyltransferase 2B7
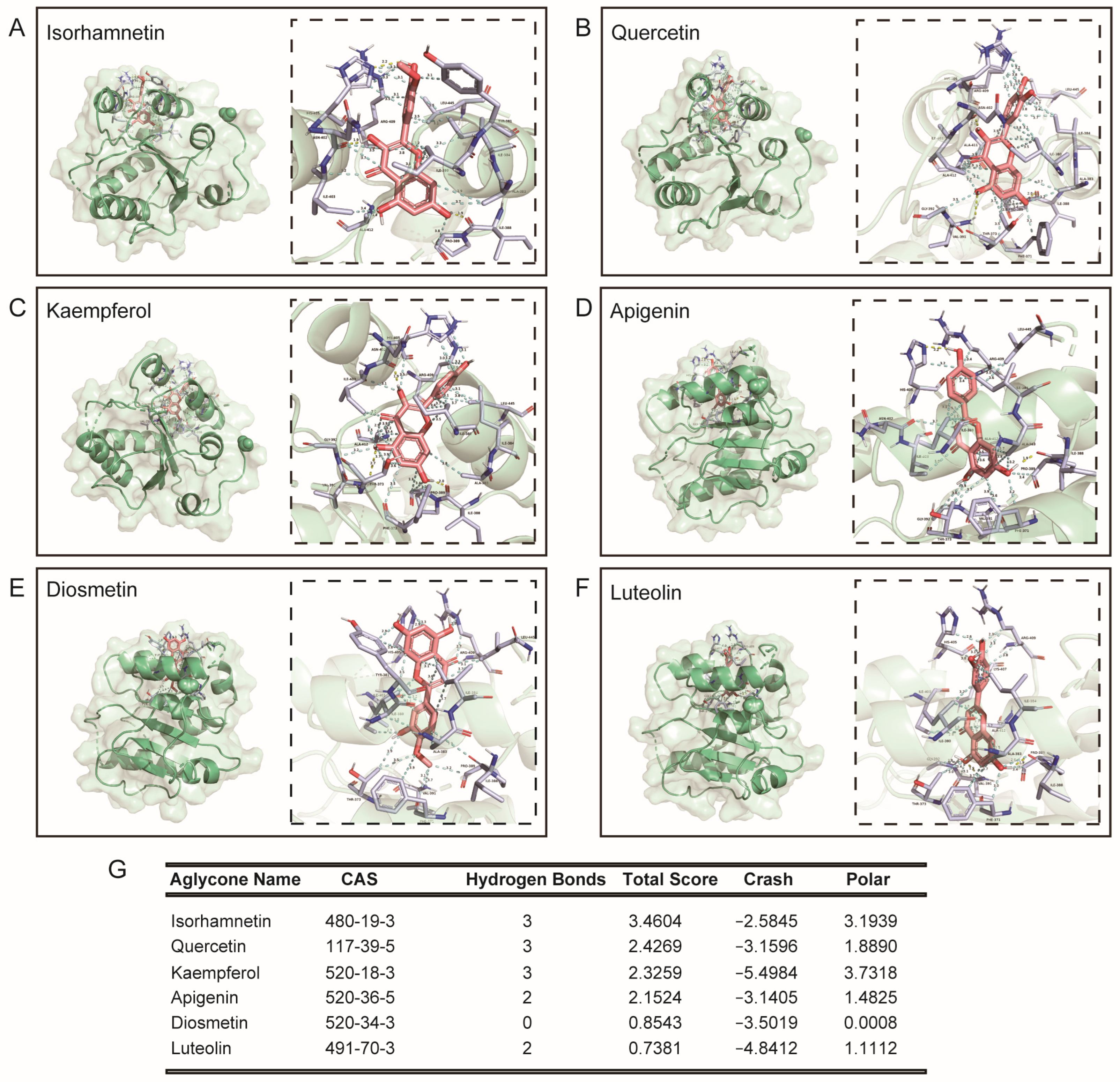
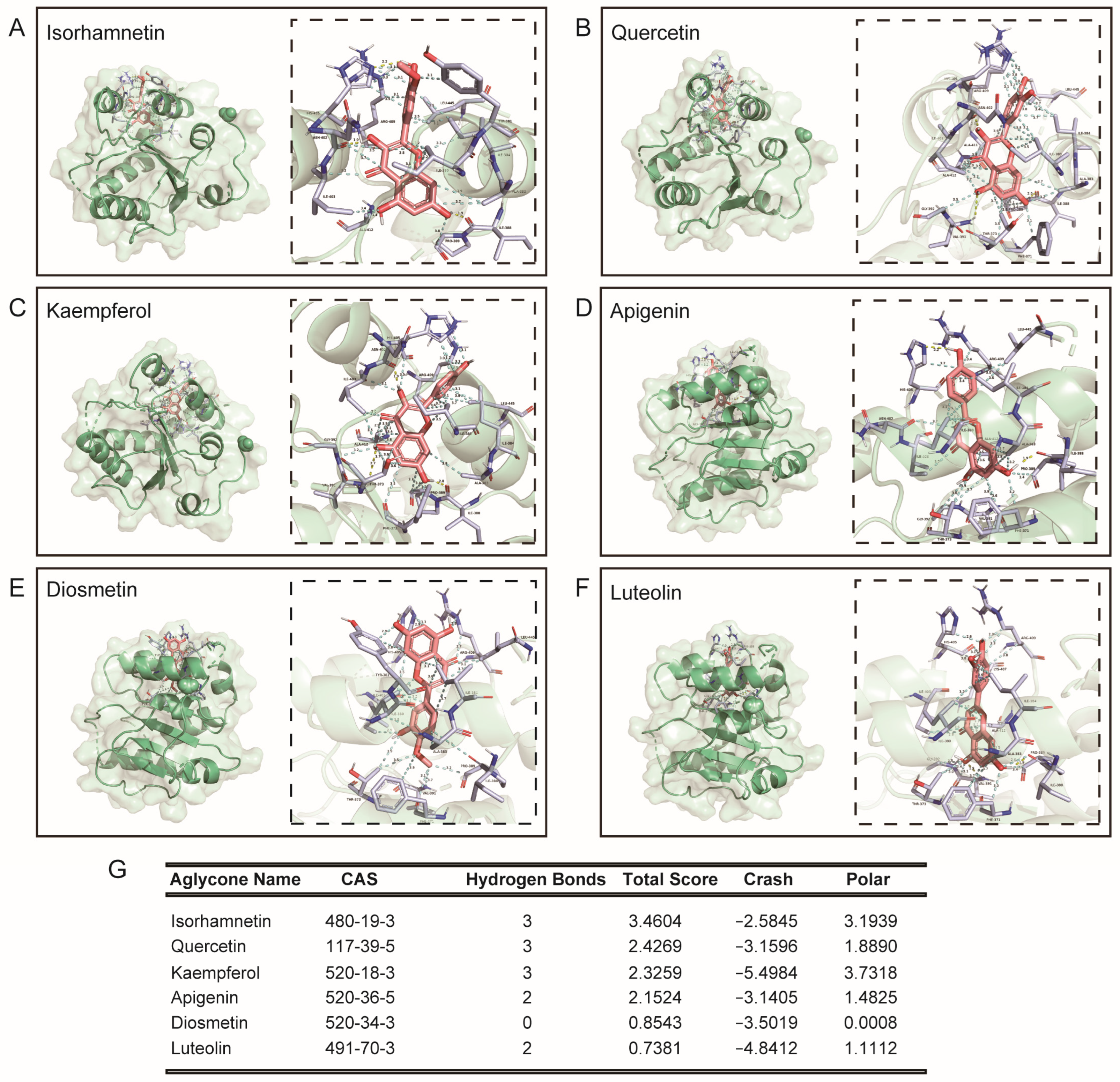
3.2. Molecular Docking of Rutin, Its Phenolic Acid Metabolites, O-Glycosylflavones, and C-Glycosylflavones towards Uridine Diphosphate Glucuronosyltransferase 2B7 (2O6L)
3.3. O-Glycosylflavone Compounds Generally Inhibit the Activity of Uridine Diphosphate Glucuronosyltransferase 2B7
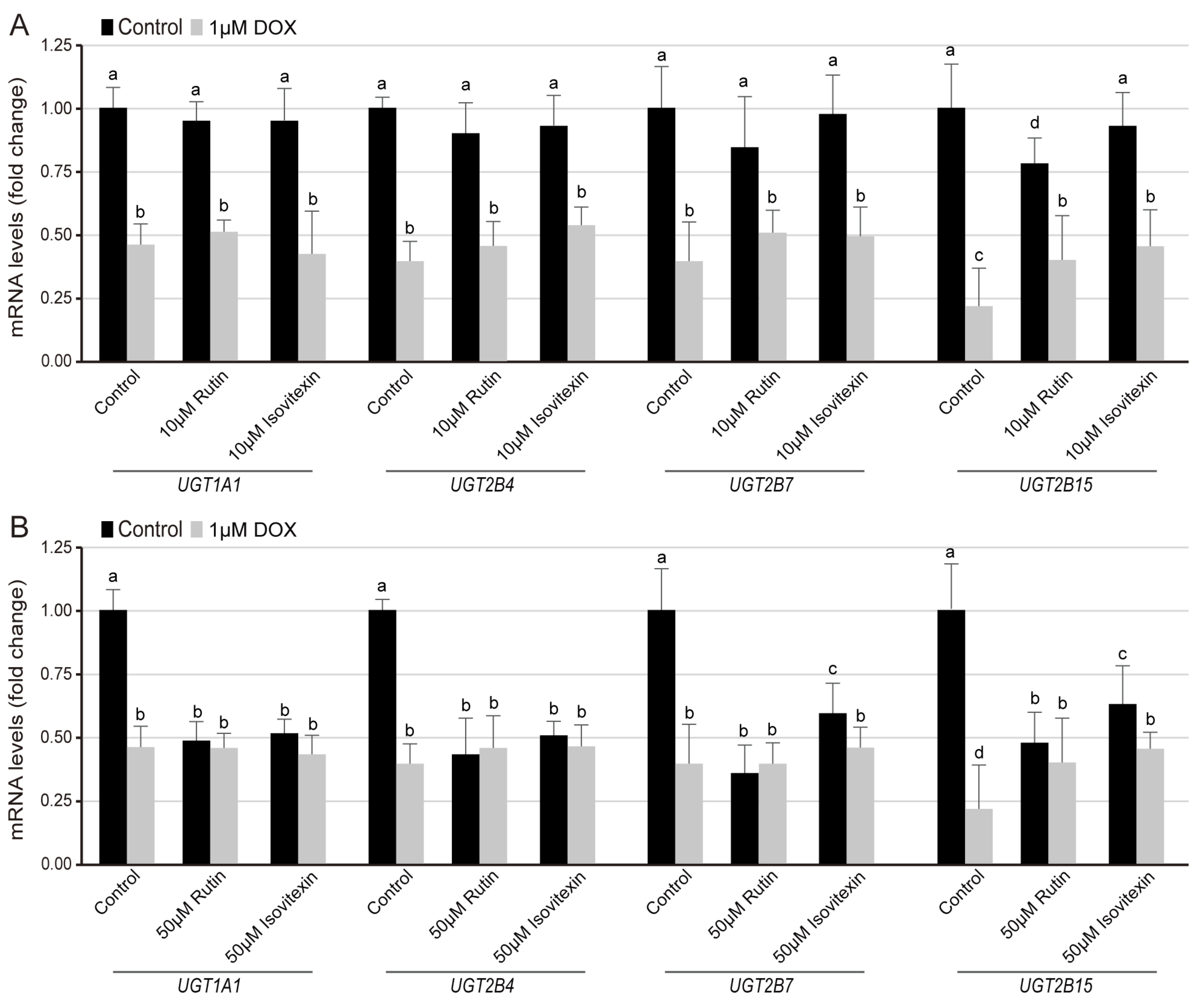
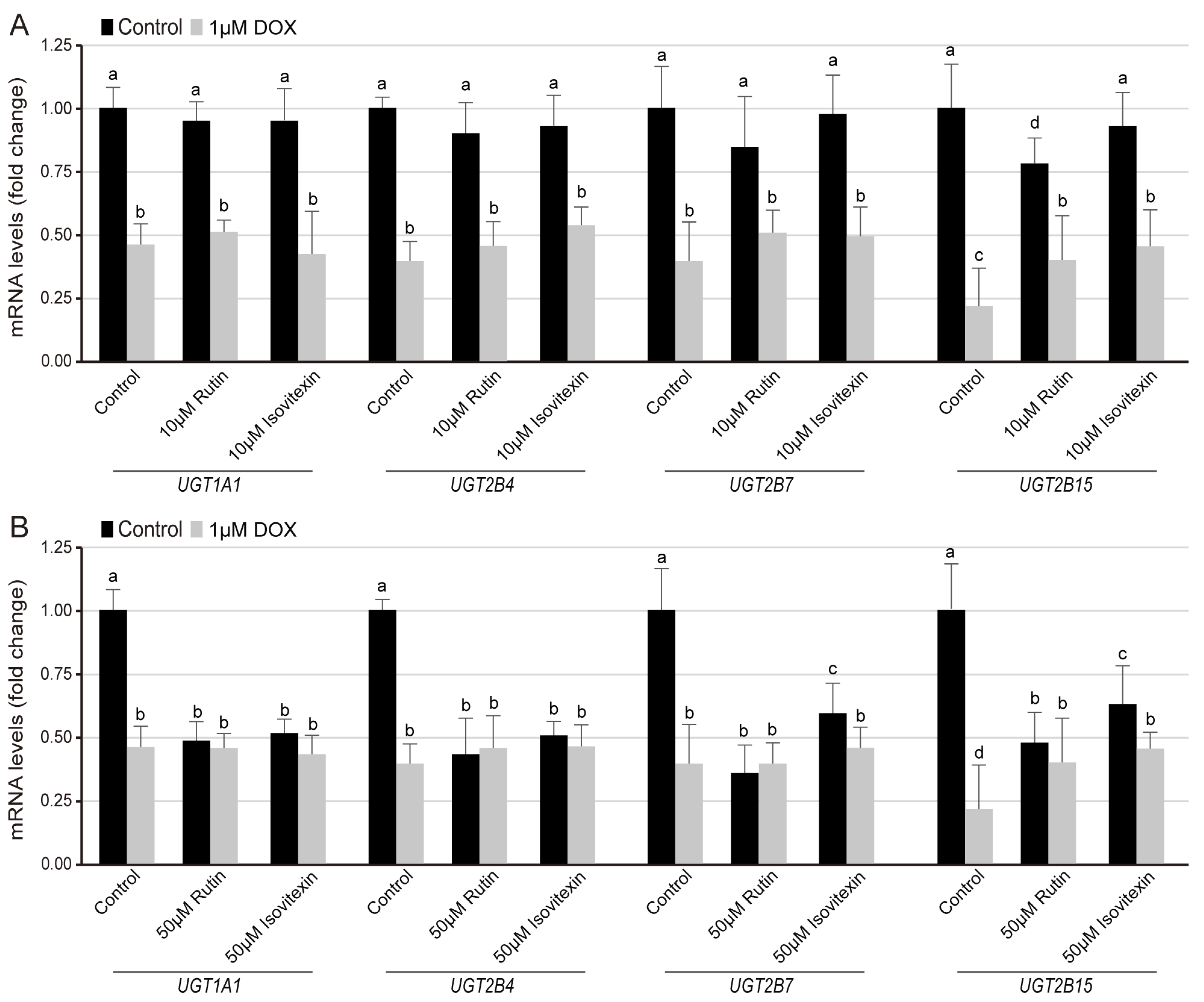
3.4. Regulation of Flavone Glycosides on the Transcription Level of UGT Families in Normal and DOX-Induced Lipid Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| UDPGA | Uridine diphosphate glucuronic acid |
| UGT | Uridine diphosphate glucuronosyltransferase |
| UGT2B7 | UDP-Glucuronosyltransferase Family 2 Member B7 |
| DHT | 3,4-dihydroxytoluene |
| HVA | 4-hydroxy-3-methoxyphenylacetic acid |
| DHPAA | 3,4-dihydroxyphenylacetic acid |
| HPAA | 3-hydroxyphenylacetic acid |
| 4-MU | 4-methylumbelliferone |
| 4-MUG | 4-methyl-2-oxo-2H-1-benzopyran-7-yl-β-D-glucopyranosiduronic acid |
| DOX | Doxycycline |
| HKC | Human renal proximal tubular cell line |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ANOVA | One-way analysis of variance |
| SULT | Sulfotransferase |
| COMT | Catechol-O-methyltransferase |
| CYP | Cytochrome P450 |
References
- DiMarco-Crook, C.; Xiao, H. Diet-Based Strategies for Cancer Chemoprevention: The Role of Combination Regimens Using Dietary Bioactive Components. Annu. Rev. Food Sci. Technol. 2015, 6, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Amrouche, T.A.; Yang, X.; Capanoglu, E.; Huang, W.; Chen, Q.; Wu, L.; Zhu, Y.; Liu, Y.; Wang, Y.; Lu, B. Contribution of edible flowers to the Mediterranean diet: Phytonutrients, bioactivity evaluation and applications. Food Front. 2022, 3, 592–630. [Google Scholar] [CrossRef]
- Saul, N.; Pietsch, K.; Stürzenbaum, S.R.; Menzel, R.; Steinberg, C.E. Diversity of polyphenol action in Caenorhabditis elegans: Between toxicity and longevity. J. Nat. Prod. 2011, 74, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Radavelli-Bagatini, S.; Sim, M.; Blekkenhorst, L.C.; Bondonno, N.P.; Bondonno, C.P.; Woodman, R.; Dickson, J.M.; Harms, C.; Magliano, D.J.; Shaw, J.E.; et al. Higher Consumption of Fruit and Vegetables Is Associated with Lower Worries, Tension and Lack of Joy Across the Lifespan. Front. Nutr. 2022, 9, 837066. [Google Scholar] [CrossRef]
- Bermont, F.; Hermant, A.; Benninga, R.; Chabert, C.; Jacot, G.; Santo-Domingo, J.; Kraus, M.R.; Feige, J.N.; De Marchi, U. Targeting Mitochondrial Calcium Uptake with the Natural Flavonol Kaempferol, to Promote Metabolism/Secretion Coupling in Pancreatic beta-cells. Nutrients 2020, 12, 538. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, R.; Nakahira, K.; Gu, Z. Mitochondrial DNA Mutation, Diseases, and Nutrient-Regulated Mitophagy. Annu. Rev. Nutr. 2019, 39, 201–226. [Google Scholar] [CrossRef]
- Gerges, S.H.; Wahdan, S.A.; Elsherbiny, D.A.; El-Demerdash, E. Pharmacology of Diosmin, a Citrus Flavone Glycoside: An Updated Review. Eur. J. Drug Metab. Pharmacokinet. 2022, 47, 1–18. [Google Scholar] [CrossRef]
- An, J.P.; Dang, L.H.; Ha, T.K.Q.; Pham, H.T.T.; Lee, B.W.; Lee, C.H.; Oh, W.K. Flavone glycosides from Sicyos angulatus and their inhibitory effects on hepatic lipid accumulation. Phytochemistry 2019, 157, 53–63. [Google Scholar] [CrossRef]
- Liang, Z.B.; Zhang, B.; Su, W.W.; Williams, P.G.; Li, Q.X. C-Glycosylflavones Alleviate Tau Phosphorylation and Amyloid Neurotoxicity through GSK3 beta Inhibition. ACS Chem. Neurosci. 2016, 7, 912–923. [Google Scholar] [CrossRef]
- Jancova, P.; Anzenbacher, P.; Anzenbacherova, E. Phase II drug metabolizing enzymes. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub. 2010, 154, 103–116. [Google Scholar] [CrossRef]
- Jaladanki, C.K.; Gahlawat, A.; Rathod, G.; Sandhu, H.; Jahan, K.; Bharatam, P.V. Mechanistic studies on the drug metabolism and toxicity originating from cytochromes P450. Drug Metab. Rev. 2020, 52, 366–394. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, G.; Wang, G.; Hao, H. Regulation of Mammalian UDP-Glucuronosyltransferases. Curr. Drug Metab. 2018, 19, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Little, J.M.; Williams, L.; Xu, J.; Radominska-Pandya, A. Glucuronidation of the dietary fatty acids, phytanic acid and docosahexaenoic acid, by human UDP-glucuronosyltransferases. Drug Metab. Dispos. 2002, 30, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Marri, S.; Mackenzie, P.I.; Hulin, J.A.; McKinnon, R.A.; Meech, R. The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes. Cancers 2021, 13, 4491. [Google Scholar] [CrossRef] [PubMed]
- Rowland, A.; Miners, J.O.; Mackenzie, P.I. The UDP-glucuronosyltransferases: Their role in drug metabolism and detoxification. Int. J. Biochem. Cell Biol. 2013, 45, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Lammers, L.A.; Achterbergh, R.; Romijn, J.A.; Mathot, R.A.A. Nutritional Status Differentially Alters Cytochrome P450 3A4 (CYP3A4) and Uridine 5′-Diphospho-Glucuronosyltransferase (UGT) Mediated Drug Metabolism: Effect of Short-Term Fasting and High Fat Diet on Midazolam Metabolism. Eur. J. Drug Metab. Pharmacokinet. 2018, 43, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, P.; Little, J.M.; Radominska-Pandya, A. Glucosidation of hyodeoxycholic acid by UDP-glucuronosyltransferase 2B7. Biochem. Pharmacol. 2003, 65, 417–421. [Google Scholar] [CrossRef]
- Barre, L.; Fournel-Gigleux, S.; Finel, M.; Netter, P.; Magdalou, J.; Ouzzine, M. Substrate specificity of the human UDP-glucuronosyltransferase UGT2B4 and UGT2B7. Identification of a critical aromatic amino acid residue at position 33. FEBS J. 2007, 274, 1256–1264. [Google Scholar] [CrossRef]
- Antonio, L.; Xu, J.; Little, J.M.; Burchell, B.; Magdalou, J.; Radominska-Pandya, A. Glucuronidation of catechols by human hepatic, gastric, and intestinal microsomal UDP-glucuronosyltransferases (UGT) and recombinant UGT1A6, UGT1A9, and UGT2B7. Arch. Biochem. Biophys. 2003, 411, 251–261. [Google Scholar] [CrossRef]
- Owens, I.S.; Basu, N.K.; Banerjee, R. UDP-glucuronosyltransferases: Gene structures of UGT1 and UGT2 families. Methods Enzym. 2005, 400, 1–22. [Google Scholar] [CrossRef]
- Maruo, Y.; Iwai, M.; Mori, A.; Sato, H.; Takeuchi, Y. Polymorphism of UDP-glucuronosyltransferase and drug metabolism. Curr. Drug Metab. 2005, 6, 91–99. [Google Scholar] [CrossRef]
- Turgeon, D.; Carrier, J.S.; Levesque, E.; Hum, D.W.; Belanger, A. Relative enzymatic activity, protein stability, and tissue distribution of human steroid-metabolizing UGT2B subfamily members. Endocrinology 2001, 142, 778–787. [Google Scholar] [CrossRef]
- Nair, P.C.; Chau, N.; McKinnon, R.A.; Miners, J.O. Arginine-259 of UGT2B7 Confers UDP-Sugar Selectivity. Mol. Pharmacol. 2020, 98, 710–718. [Google Scholar] [CrossRef]
- Radominska-Pandya, A.; Czernik, P.J.; Little, J.M.; Battaglia, E.; Mackenzie, P.I. Structural and functional studies of UDP-glucuronosyltransferases. Drug Metab. Rev. 1999, 31, 817–899. [Google Scholar] [CrossRef]
- Liu, Y.; Coughtrie, M.W.H. Revisiting the Latency of Uridine Diphosphate-Glucuronosyltransferases (UGTs)-How Does the Endoplasmic Reticulum Membrane Influence Their Function? Pharmaceutics 2017, 9, 32. [Google Scholar] [CrossRef]
- Mackenzie, P.I.; Rodbourne, L.; Stranks, S. Steroid UDP glucuronosyltransferases. J. Steroid Biochem. Mol. Biol. 1992, 43, 1099–1105. [Google Scholar] [CrossRef]
- He, B.X.; Qiao, B.; Lam, A.K.; Zhao, X.L.; Zhang, W.Z.; Liu, H. Association between UDP-glucuronosyltransferase 2B7 tagSNPs and breast cancer risk in Chinese females. Clin. Exp. Pharmacol. Physiol. 2018, 45, 437–443. [Google Scholar] [CrossRef]
- Miley, M.J.; Zielinska, A.K.; Keenan, J.E.; Bratton, S.M.; Radominska-Pandya, A.; Redinbo, M.R. Crystal structure of the cofactor-binding domain of the human phase II drug-metabolism enzyme UDP-glucuronosyltransferase 2B7. J. Mol. Biol. 2007, 369, 498–511. [Google Scholar] [CrossRef]
- Zhang, R.; Wei, Y.; Yang, T.; Huang, X.; Zhou, J.; Yang, C.; Zhou, J.; Liu, Y.; Shi, S. Inhibitory effects of quercetin and its major metabolite quercetin-3-O-β-D-glucoside on human UDP-glucuronosyltransferase 1A isoforms by liquid chromatography-tandem mass spectrometry. Exp. Ther. Med. 2021, 22, 842. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, C.; Chen, J.; Hu, X.; Zhang, H.; Li, Z.; Lan, B.; Zhang, W.; Su, Y.; Zhang, C. Potential interactions among myricetin and dietary flavonols through the inhibition of human UDP-glucuronosyltransferase in vitro. Toxicol. Lett. 2022, 358, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, S.; Yang, K.; Liu, Y.Z.; Yang, K.; Chen, Y.; Fang, Z.Z. Inhibition of UDP-glucuronosyltransferases (UGTs) by bromophenols (BPs). Chemosphere 2020, 238, 124645. [Google Scholar] [CrossRef]
- Zhang, X.X.; Cao, Y.F.; Wang, L.X.; Yuan, X.L.; Fang, Z.Z. Inhibitory effects of tanshinones towards the catalytic activity of UDP-glucuronosyltransferases (UGTs). Pharm. Biol. 2017, 55, 1703–1709. [Google Scholar] [CrossRef]
- Gao, W.F.; Li, Y.X.; Zhang, W.H.; Tao, R.; Yin, T.T.; Wang, Y.J.; Liu, L.N.; Fu, Z.W.; Li, S.N.; Liu, N.R.; et al. Comparison of the inhibition potential of parthenolide and micheliolide on various UDP-glucuronosyltransferase isoforms. Xenobiotica 2019, 49, 1158–1163. [Google Scholar] [CrossRef]
- Mano, Y.; Usui, T.; Kamimura, H. Substrate-dependent modulation of UDP-glucuronosyltransferase 1A1 (UGT1A1) by propofol in recombinant human UGT1A1 and human liver microsomes. Basic Clin. Pharmacol. Toxicol. 2007, 101, 211–214. [Google Scholar] [CrossRef]
- Chengcheng, G.; Rui, X.; Tianheng, M.; Wei, Y.; Liqun, P. Probe substrate and enzyme source-dependent inhibition of UDP-glucuronosyltransferase (UGT) 1A9 by wogonin. Afr. Health Sci. 2013, 13, 551–555. [Google Scholar] [CrossRef]
- Miners, J.O.; Lillywhite, K.J.; Matthews, A.P.; Jones, M.E.; Birkett, D.J. Kinetic and inhibitor studies of 4-methylumbelliferone and 1-naphthol glucuronidation in human liver microsomes. Biochem. Pharmacol. 1988, 37, 665–671. [Google Scholar] [CrossRef]
- Fang, Z.Z.; Cao, Y.F.; Hu, C.M.; Hong, M.; Sun, X.Y.; Ge, G.B.; Liu, Y.; Zhang, Y.Y.; Yang, L.; Sun, H.Z. Structure-inhibition relationship of ginsenosides towards UDP-glucuronosyltransferases (UGTs). Toxicol. Appl. Pharmacol. 2013, 267, 149–154. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Zheng, L.; He, X.; Luo, Y.; Huang, K.; Xu, W. Glucose-regulated protein 75 in foodborne disease models induces renal tubular necrosis. Food Chem. Toxicol. 2019, 133, 110720. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, S.; Huang, C.; Wang, H.; Luo, Y.; Xu, W.; Huang, K. Ochratoxin A induced premature senescence in human renal proximal tubular cells. Toxicology 2017, 382, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Pashikanti, S.; de Alba, D.R.; Boissonneault, G.A.; Cervantes-Laurean, D. Rutin metabolites: Novel inhibitors of nonoxidative advanced glycation end products. Free Radic. Biol. Med. 2010, 48, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Pfeiffer, M.; Martini, M.; Campbell, D.; Slavin, J.; Potter, J. The quantitation of metabolites of quercetin flavonols in human urine. Cancer Epidemiol. Biomarks Prev. 1996, 5, 711–720. [Google Scholar]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, J.; Shi, X.; Miao, S.; Li, Y.; Wen, A. Absorption and metabolism characteristics of rutin in Caco-2 cells. Sci. World J. 2013, 2013, 382350. [Google Scholar] [CrossRef]
- de Moura, P.H.B.; Porzel, A.; Nunes, R.M.; Baratto, L.C.; Wessjohann, L.A.; Martins, R.C.C.; Leal, I.C.R. Antioxidant capacity and fragmentation features of C-glycoside isoflavones by high-resolution electrospray ionization tandem mass spectrometry using collision-induced and high-energy collisional dissociation techniques. J. Mass. Spectrom. 2021, 56, e4793. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.L.; Chang, W.S.; Lu, W.C.; Wei, G.J.; Wang, Y.; Ho, C.T.; Hwang, L.S. Pharmacokinetics, bioavailability, tissue distribution and excretion of tangeretin in rat. J. Food Drug Anal. 2018, 26, 849–857. [Google Scholar] [CrossRef]
- Gao, H.; Jiang, Q.; Han, Y.; Peng, J.; Wang, C. Hispidulin potentiates the antitumor effect of sunitinib against human renal cell carcinoma in laboratory models. Cell Biochem. Biophys. 2015, 71, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Basu, S.; Meng, S.; Wang, X.; Hu, M. Regioselective sulfation and glucuronidation of phenolics: Insights into the structural basis. Curr. Drug Metab. 2011, 12, 900–916. [Google Scholar] [CrossRef]
- Wang, R.; Wang, L.; Zhang, L.; Wan, S.; Li, C.; Liu, S. Solvents effect on phenolics, iridoids, antioxidant activity, antibacterial activity, and pancreatic lipase inhibition activity of noni (Morinda citrifolia L.) fruit extract. Food Chem. 2022, 377, 131989. [Google Scholar] [CrossRef]
- Phuong Thao, T.T.; Bui, T.Q.; Quy, P.T.; Bao, N.C.; Van Loc, T.; Van Chien, T.; Chi, N.L.; Van Tuan, N.; Van Sung, T.; Ai Nhung, N.T. Isolation, semi-synthesis, docking-based prediction, and bioassay-based activity of Dolichandrone spathacea iridoids: New catalpol derivatives as glucosidase inhibitors. RSC Adv. 2021, 11, 11959–11975. [Google Scholar] [CrossRef]
- Rensvold, J.W.; Shishkova, E.; Sverchkov, Y.; Miller, I.J.; Cetinkaya, A.; Pyle, A.; Manicki, M.; Brademan, D.R.; Alanay, Y.; Raiman, J.; et al. Defining mitochondrial protein functions through deep multiomic profiling. Nature 2022, 606, 382–388. [Google Scholar] [CrossRef]
- Tan, S.; Wu, T.; Zhang, D.; Zhang, Z. Cell or cell membrane-based drug delivery systems. Theranostics 2015, 5, 863–881. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, L.F.; Chen, X.C.; Huang, X.R.; Li, H.Y.; An, N.; Tang, J.X.; Liu, H.F.; Yang, C. Research progress on endoplasmic reticulum homeostasis in kidney diseases. Cell Death Dis. 2023, 14, 473. [Google Scholar] [CrossRef] [PubMed]
- Koleini, N.; Kardami, E. Autophagy and mitophagy in the context of doxorubicin-induced cardiotoxicity. Oncotarget 2017, 8, 46663–46680. [Google Scholar] [CrossRef] [PubMed]
- Boer, J.; Young-Sciame, R.; Lee, F.; Bowman, K.J.; Yang, X.; Shi, J.G.; Nedza, F.M.; Frietze, W.; Galya, L.; Combs, A.P.; et al. Roles of UGT, P450, and Gut Microbiota in the Metabolism of Epacadostat in Humans. Drug Metab. Dispos. 2016, 44, 1668–1674. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wang, H.L.; Yu, H.L.; Cui, X.H.; Xu, M.T.; Xu, X.; Gao, J.P. Doxorubicin toxicity changes myocardial energy metabolism in rats. Chem. Biol. Interact. 2016, 244, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, W.; Huang, K.; Zhang, B.; Wang, H.; Zhang, X.; Gong, L.; Luo, Y.; He, X. Precision toxicology shows that troxerutin alleviates ochratoxin A-induced renal lipotoxicity. FASEB J. 2019, 33, 2212–2227. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Xia, Y.M.; Zhang, Y.D.; Zhang, T.T.; Peng, Q.R.; Fang, Y. Influence of substrates on the in vitro kinetics of steviol glucuronidation and interaction between steviol glycosides metabolites and UGT2B7. Int. J. Food Sci. Nutr. 2018, 69, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450 research and The Journal of Biological Chemistry. J. Biol. Chem. 2019, 294, 1671–1680. [Google Scholar] [CrossRef]
- Knights, K.M.; Rowland, A.; Miners, J.O. Renal drug metabolism in humans: The potential for drug-endobiotic interactions involving cytochrome P450 (CYP) and UDP-glucuronosyltransferase (UGT). Br. J. Clin. Pharmacol. 2013, 76, 587–602. [Google Scholar] [CrossRef]
- Reed, J.R.; Backes, W.L. The functional effects of physical interactions involving cytochromes P450: Putative mechanisms of action and the extent of these effects in biological membranes. Drug Metab. Rev. 2016, 48, 453–469. [Google Scholar] [CrossRef]
- Omura, T. Structural diversity of cytochrome P450 enzyme system. J. Biochem. 2010, 147, 297–306. [Google Scholar] [CrossRef]
- Saito, K.; Moriya, H.; Sawaguchi, T.; Hayakawa, T.; Nakahara, S.; Goto, A.; Arimura, Y.; Imai, K.; Kurosawa, N.; Owada, E.; et al. Haplotype analysis of UDP-glucuronocyltransferase 2B7 gene (UGT2B7) polymorphisms in healthy Japanese subjects. Clin. Biochem. 2006, 39, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.F.; Guo, W.C.; Chen, J.G.; Qin, Y.Q.; Golka, K.; Xiang, C.Q.; Ma, Q.W.; Lu, D.R.; Shen, J.H. An association of UDP-glucuronosyltransferase 2B7 C802T (His268Tyr) polymorphism with bladder cancer in benzidine-exposed workers in China. Toxicol. Sci. 2005, 85, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.S.; Lee, S.J.; Jeong, H.E.; Lee, S.; Yoo, M.A.; Shin, J.G. Genetic variations in UDP-glucuronosyltransferase 2B7 gene (UGT2B7) in a Korean population. Drug Metab. Pharmacokinet. 2010, 25, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Novillo, A.; Gaibar, M.; Romero-Lorca, A.; Chaabani, H.; Amir, N.; Moral, P.; Esteban, M.E.; Fernandez-Santander, A. UDP-glucuronosyltransferase genetic variation in North African populations: A comparison with African and European data. Ann. Hum. Biol. 2018, 45, 516–523. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′→3′) |
|---|---|
| UGT1A1-F | GCTTTTGTCTGGCTGTTCCCACT |
| UGT1A1-R | TCGAAGGTCATGTGATCTGAATGAGA |
| UGT2B4-F | GTATTGGCATCTTCAGCTTCCATTTC |
| UGT2B4-R | AAGTTCTGCCCATCTCTTAACCAGC |
| UGT2B7-F | TTTCACAAGTACAGGAAATCATGTCAAT |
| UGT2B7-R | CAGCAGCTCACTACAGGGAAAAAT |
| UGT2B15-F | GAAAATTCTCGATAGATGGATATATGGTG |
| UGT2B15-R | AACTGCATCTTTACAGAGCTTGTTACTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, T.; Lv, Y.; Cui, Y.; Liu, D.; Xu, T.; Lu, B.; Yang, X. Properties of Dietary Flavone Glycosides, Aglycones, and Metabolites on the Catalysis of Human Endoplasmic Reticulum Uridine Diphosphate Glucuronosyltransferase 2B7 (UGT2B7). Nutrients 2023, 15, 4941. https://doi.org/10.3390/nu15234941
Xu T, Lv Y, Cui Y, Liu D, Xu T, Lu B, Yang X. Properties of Dietary Flavone Glycosides, Aglycones, and Metabolites on the Catalysis of Human Endoplasmic Reticulum Uridine Diphosphate Glucuronosyltransferase 2B7 (UGT2B7). Nutrients. 2023; 15(23):4941. https://doi.org/10.3390/nu15234941
Chicago/Turabian StyleXu, Ting, Yangjun Lv, Yuhan Cui, Dongchen Liu, Tao Xu, Baiyi Lu, and Xuan Yang. 2023. "Properties of Dietary Flavone Glycosides, Aglycones, and Metabolites on the Catalysis of Human Endoplasmic Reticulum Uridine Diphosphate Glucuronosyltransferase 2B7 (UGT2B7)" Nutrients 15, no. 23: 4941. https://doi.org/10.3390/nu15234941
APA StyleXu, T., Lv, Y., Cui, Y., Liu, D., Xu, T., Lu, B., & Yang, X. (2023). Properties of Dietary Flavone Glycosides, Aglycones, and Metabolites on the Catalysis of Human Endoplasmic Reticulum Uridine Diphosphate Glucuronosyltransferase 2B7 (UGT2B7). Nutrients, 15(23), 4941. https://doi.org/10.3390/nu15234941