
Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study

 , , , ,
, , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Clinical Parameters
2.2.1. Anthropometric Measurements
2.2.2. Blood Pressure Measurements
2.2.3. Lipid Profile and Glucose Measurement
2.3. Dietary Intake
2.4. Faecal Sample Collection
2.5. Sample Size
2.6. Faecal Microbiota Analysis
2.6.1. DNA Purification and Sequencing
2.6.2. Fecal Microbiota Analysis: Sequence Analysis
2.7. Statistical Analysis
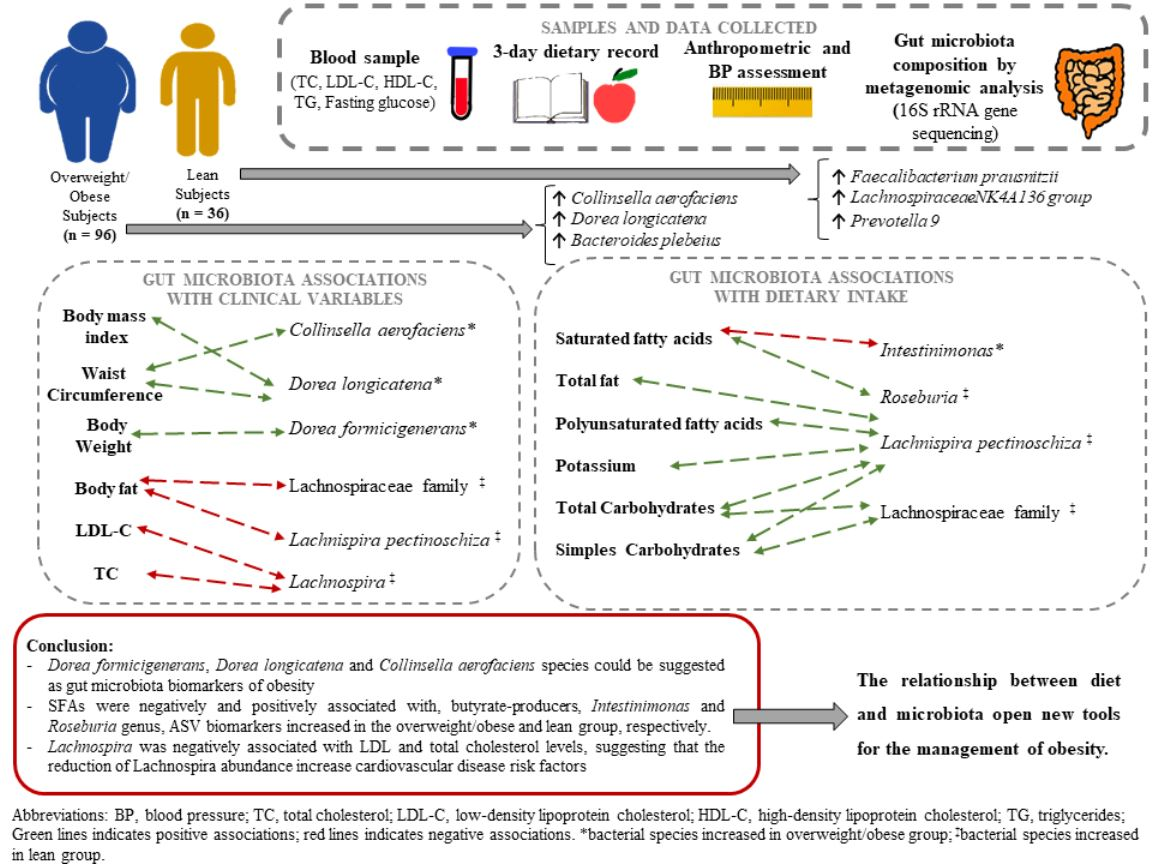
3. Results
3.1. Baseline Characteristics of the Participants
3.2. Dietary Assessment
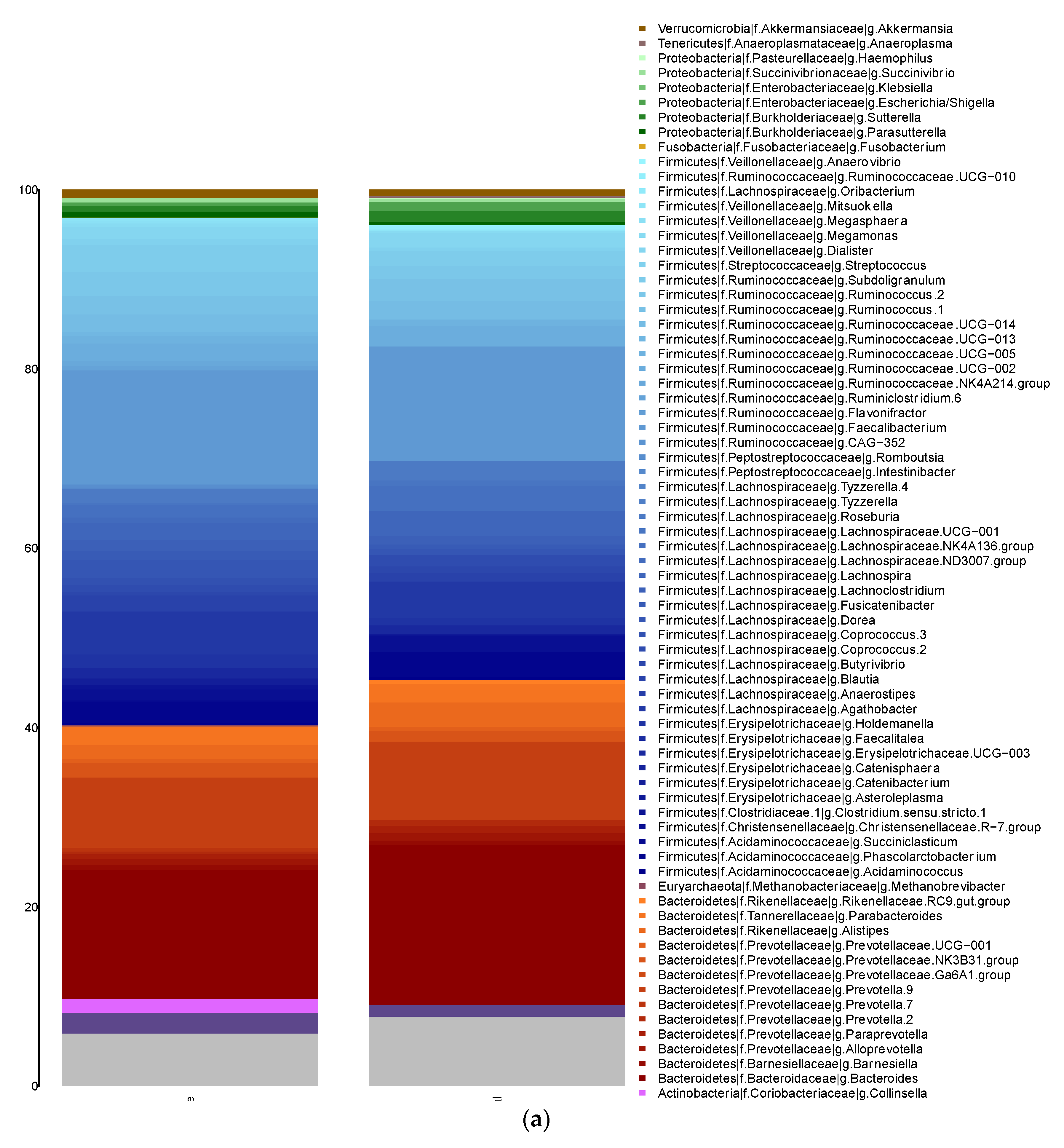
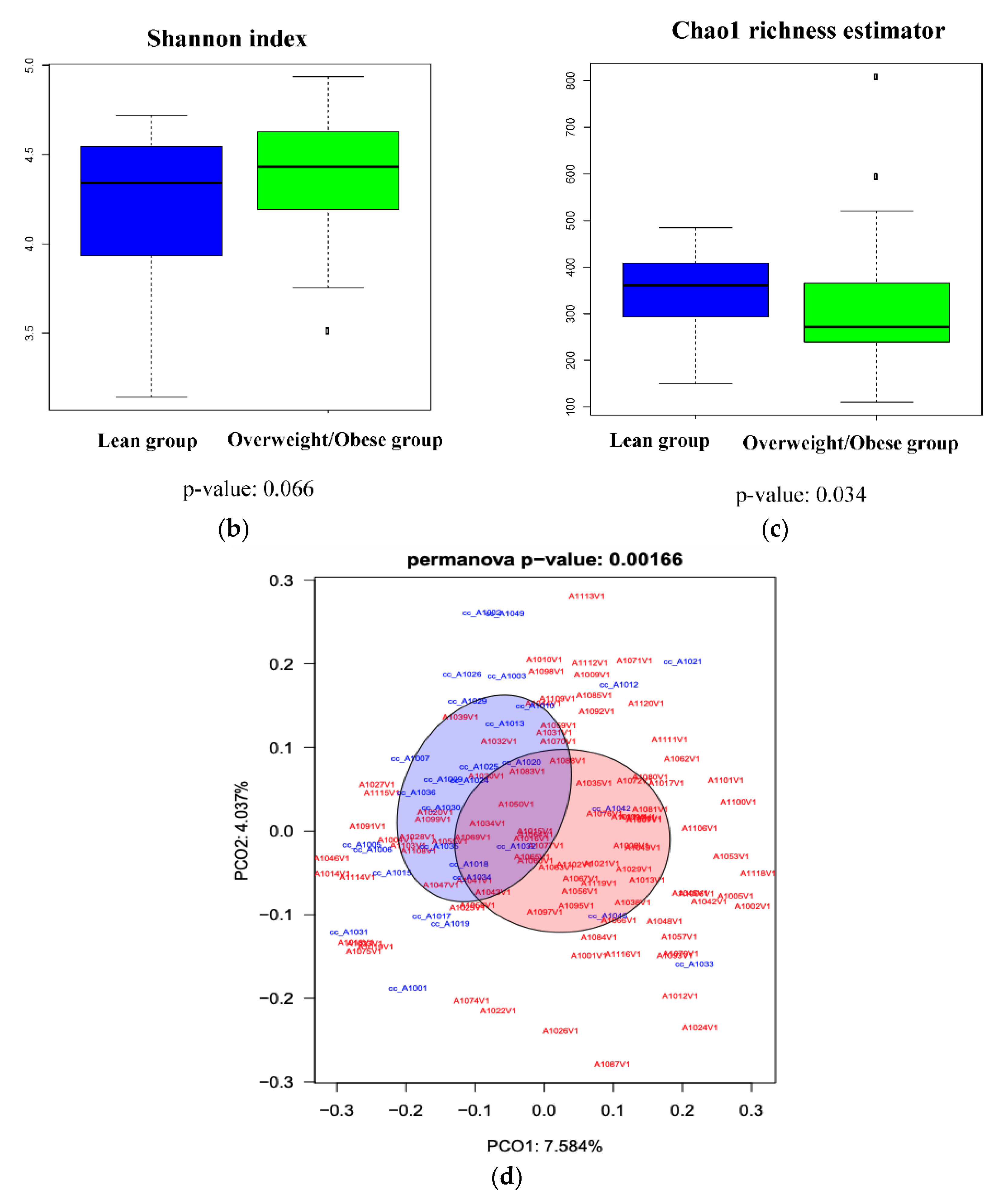
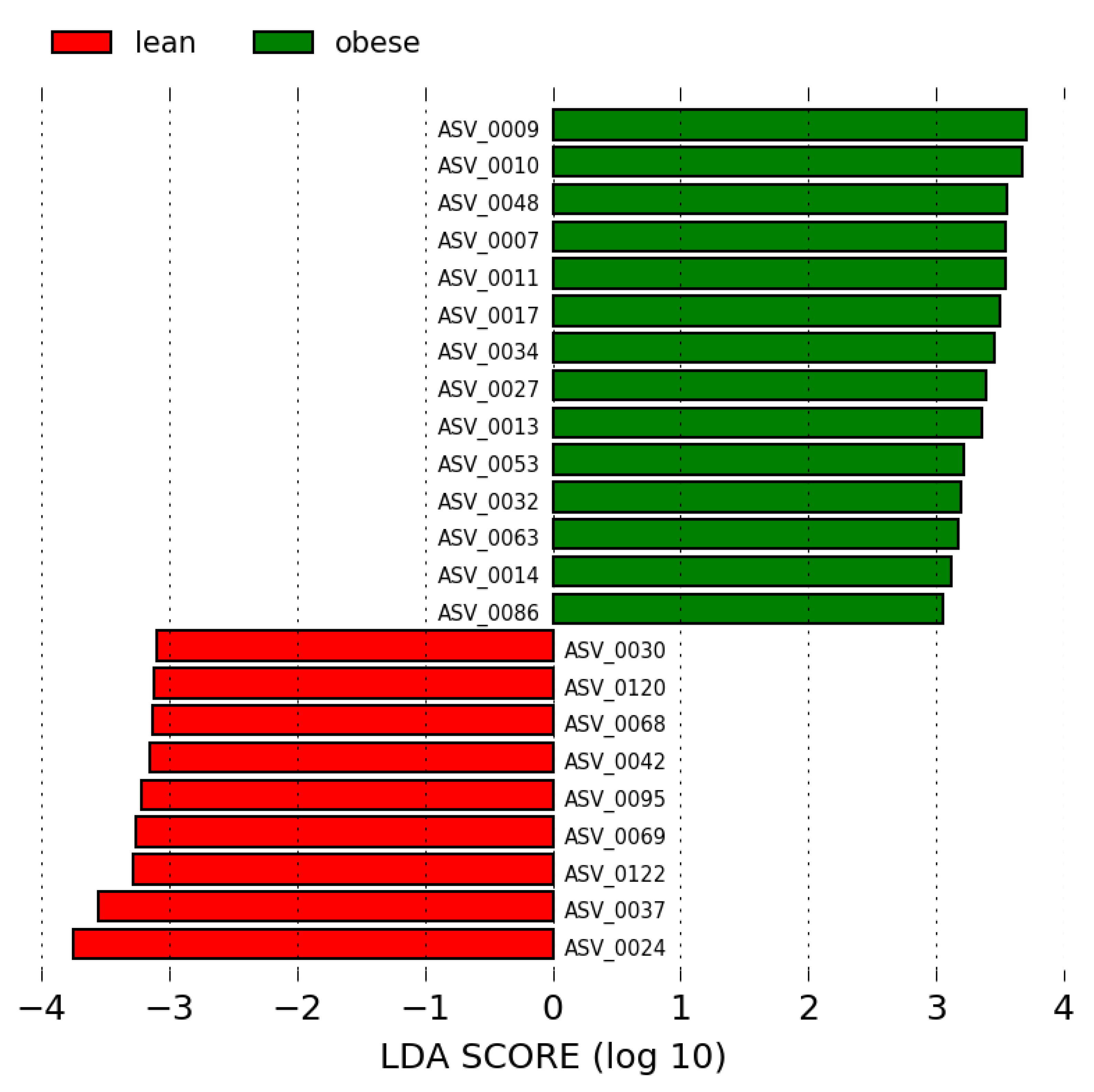
3.3. Analysis of the Gut Microbiota Composition
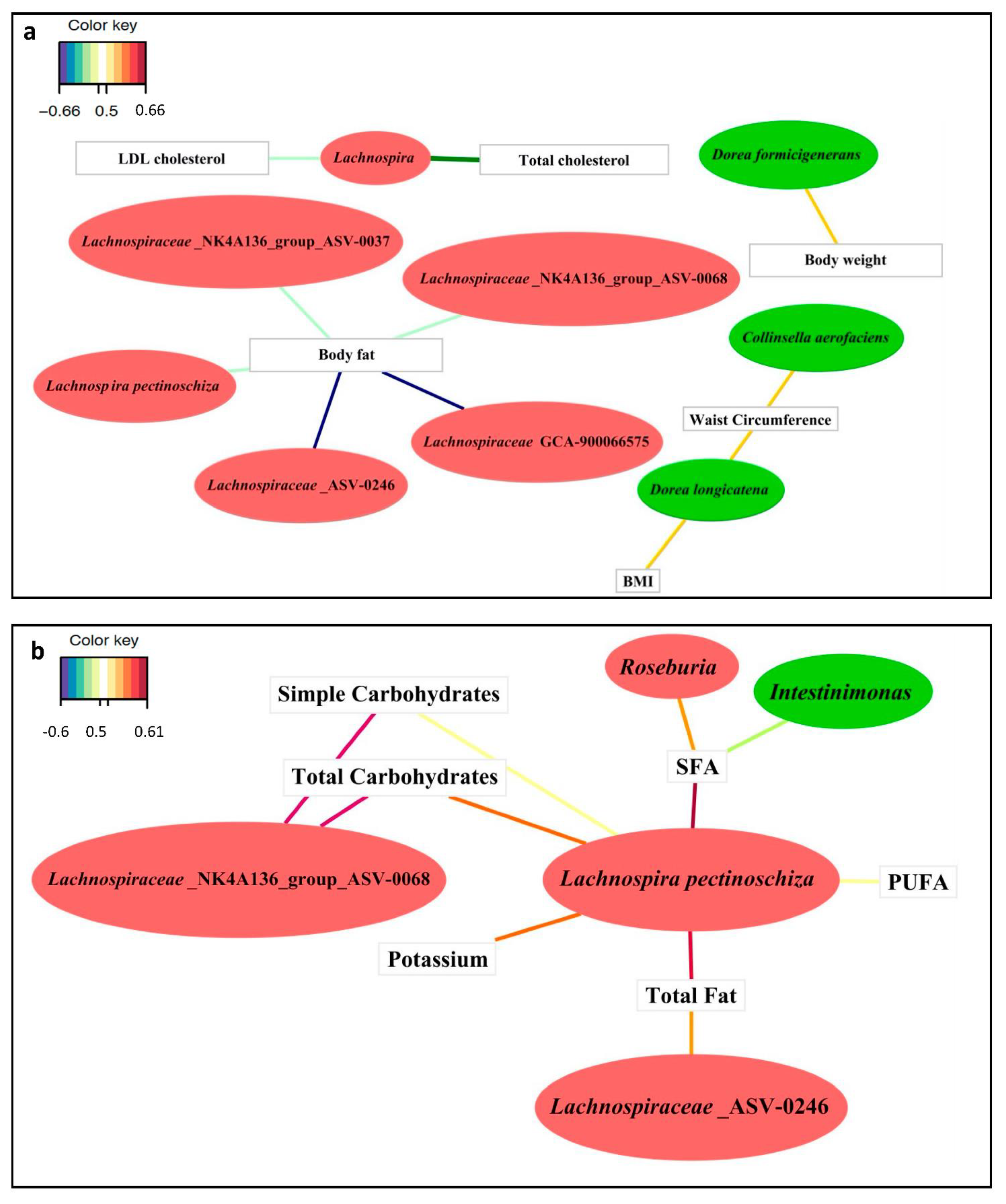
3.3.1. Associations between Clinical Variables and the Gut Microbiota
3.3.2. Associations between Diet and the Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 15 July 2020).
- National Heart Lung and Blood Institute Overweight and Obesity | NHLBI, NIH. Available online: https://www.nhlbi.nih.gov/health-topics/overweight-and-obesity (accessed on 15 July 2020).
- González-Rodríguez, L.G.; Perea Sánchez, J.M.; Aranceta-Bartrina, J.; Gil, Á.; González-Gross, M.; Serra-Majem, L.; Varela-Moreiras, G.; Ortega, R.M. Intake and Dietary Food Sources of Fibre in Spain: Differences with Regard to the Prevalence of Excess Body Weight and Abdominal Obesity in Adults of the ANIBES Study. Nutrients 2017, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Klabunde, R.A.; Lazar Neto, F.; Louzada, A.; de Moura, R.F.; Calixto, F.M.; Danaei, G.; Castro, M.C. Prevalence and predictors of overweight and obesity in Brazilian immigrants in Massachusetts. BMC Public Health 2020, 20, 42. [Google Scholar] [CrossRef]
- Wolters, M.; Ahrens, J.; Romaní-Pérez, M.; Watkins, C.; Sanz, Y.; Benítez-Páez, A.; Stanton, C.; Günther, K. Dietary fat, the gut microbiota, and metabolic health—A systematic review conducted within the MyNewGut project. Clin. Nutr. 2019, 38, 2504–2520. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20–76 years of age. NPJ Biofilms Microbiomes 2019, 5. [Google Scholar] [CrossRef]
- Osborne, G.; Wu, F.; Yang, L.; Kelly, D.; Hu, J.; Li, H.; Jasmine, F.; Kibriya, M.G.; Parvez, F.; Shaheen, I.; et al. The association between gut microbiome and anthropometric measurements in Bangladesh. Gut Microbes 2020, 11, 63–76. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. The microbiota–gut–brain axis in obesity. Lancet Gastroenterol. Hepatol. 2017, 2, 747–756. [Google Scholar] [CrossRef]
- Lachat, C.; Hawwash, D.; Ocké, M.C.; Berg, C.; Forsum, E.; Hörnell, A.; Larsson, C.; Sonestedt, E.; Wirfält, E.; Åkesson, A.; et al. Strengthening the Reporting of Observational Studies in Epidemiology—Nutritional Epidemiology (STROBE-nut): An Extension of the STROBE Statement. PLoS Med. 2016, 13. [Google Scholar] [CrossRef]
- Hercberg, S.; Deheeger, M.; Preziosi, P. Portions Alimentaires: Manuel-Photos Pour L’estimation des Quantités; Economica: Paris, France, 2002. [Google Scholar]
- Reyes, L.M.; Vázquez, R.G.; Arroyo, S.M.C.; Avalos, A.M.; Castillo, P.A.R.; Pérez, D.A.C.; Terrones, I.R.; Ibáñez, N.R.; Magallanes, M.M.R.; Langella, P.; et al. Correlation between diet and gut bacteria in a population of young adults. Int. J. Food Sci. Nutr. 2016, 67, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: A cross-sectional study. Gut Pathog. 2013, 5, 10. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R A Lang. R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Best, D.J.; Roberts, D.E. Algorithm AS 89: The Upper Tail Probabilities of Spearman’s Rho. Appl. Stat. 1975. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Das, M.; Jeffery, I.B.; O’Toole, P.W. Adjusting for age improves identification of gut microbiome alterations in multiple diseases. elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, V.; Engertsberger, L.; Komarova, I.; Feldbacher, N.; Leber, B.; Pichler, G.; Fink, N.; Scarpatetti, M.; Schippinger, W.; Schmidt, R.; et al. Dysbiosis, gut barrier dysfunction and inflammation in dementia: A pilot study. BMC Geriatr. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Machado Arruda, S.P.; da Silva, A.A.M.; Kac, G.; Vilela, A.A.F.; Goldani, M.; Bettiol, H.; Barbieri, M.A. Dietary patterns are associated with excess weight and abdominal obesity in a cohort of young Brazilian adults. Eur. J. Nutr. 2016, 55, 2081–2091. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Wagner, B.D.; Sontag, M.K.; Lozupone, C.A.; Eggesbø, M. Pre-pregnancy weight, gestational weight gain, and the gut microbiota of mothers and their infants. Microbiome 2017, 5, 113. [Google Scholar] [CrossRef]
- Li, H.; Fang, Q.; Nie, Q.; Hu, J.; Yang, C.; Huang, T.; Li, H.; Nie, S. Hypoglycemic and Hypolipidemic Mechanism of Tea Polysaccharides on Type 2 Diabetic Rats via Gut Microbiota and Metabolism Alteration. J. Agric. Food Chem. 2020, 68, 10015–10028. [Google Scholar] [CrossRef]
- Dugas, L.R.; Bernabé, B.P.; Priyadarshini, M.; Fei, N.; Park, S.J.; Brown, L.; Plange-Rhule, J.; Nelson, D.; Toh, E.C.; Gao, X.; et al. Decreased microbial co-occurrence network stability and SCFA receptor level correlates with obesity in African-origin women. Sci. Rep. 2018, 8, 17135. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Kedenko, I.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut microbiota dysbiosis associated with glucose metabolism disorders and the metabolic syndrome in older adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Becerra, L.; Cornejo-Granados, F.; García-López, R.; Valdez-Lara, A.; Bikel, S.; Canizales-Quinteros, S.; López-Contreras, B.E.; Mendoza-Vargas, A.; Nielsen, H.; Ochoa-Leyva, A. Metatranscriptomic analysis to define the Secrebiome, and 16S rRNA profiling of the gut microbiome in obesity and metabolic syndrome of Mexican children. Microb. Cell Fact. 2020, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Astrup, A.; Magkos, F.; Bier, D.M.; Brenna, J.T.; de Oliveira Otto, M.C.; Hill, J.O.; King, J.C.; Mente, A.; Ordovas, J.M.; Volek, J.S.; et al. Saturated Fats and Health: A Reassessment and Proposal for Food-Based Recommendations. J. Am. Coll. Cardiol. 2020, 76, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Thingholm, L.B.; Rühlemann, M.C.; Koch, M.; Fuqua, B.; Laucke, G.; Boehm, R.; Bang, C.; Franzosa, E.A.; Hübenthal, M.; Rahnavard, A.; et al. Obese Individuals with and without Type 2 Diabetes Show Different Gut Microbial Functional Capacity and Composition. Cell Host Microbe 2019, 26, 252–264.e10. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Ritari, J.; Boeren, S.; De Waard, P.; Plugge, C.M.; De Vos, W.M. Production of butyrate from lysine and the Amadori product fructoselysine by a human gut commensal. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 fatty acids correlate with gut microbiome diversity and production of N-carbamylglutamate in middle aged and elderly women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.E.; Hsu, T.M.; Jones, R.B.; Fodor, A.A.; Goran, M.I.; Kanoski, S.E. Early-Life Sugar Consumption Affects the Rat Microbiome Independently of Obesity. J. Nutr. 2017, 147, 20–28. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Overweight and Obese Subjects (n = 96) | Lean Subjects (n = 32) | p Value |
|---|---|---|---|
| Age, y | 52.2 ± 9.7 | 40.2 ± 8.9 | <0.001 |
| Female, % | 38.5 | 50.0 | 0.254 |
| Systolic blood pressure, mm Hg | 129.8 ± 15.9 | 109.7 ± 7.1 | <0.001 |
| Diastolic blood pressure, mm Hg | 81.1 ± 9.6 | 65.8 ± 5.9 | <0.001 |
| Pulse pressure, mm Hg | 68.4 ± 8.8 | 43.8 ± 6.8 | <0.001 |
| Physical Activity, % | 0.180 | ||
| Inactive | 7.3 | 0.0 | |
| Very low activity | 3.1 | 10.0 | |
| Low activity | 14.6 | 6.7 | |
| Moderate activity | 20.8 | 20.0 | |
| High activity | 54.2 | 63.3 | |
| Anthropometric parameters | |||
| Body mass index, kg/m2 | 31.2 ± 3.4 | 23.9 ± 2.6 | <0.001 |
| Waist circumference, cm | |||
| Male | 111.2 ± 8.1 | 87.7 ± 6.6 | <0.001 |
| Female | 99.3 ± 7.6 | 80.5 ± 9.7 | <0.001 |
| Fat mass, % | 34.9 ± 8.3 | 22.3 ± 8.1 | <0.001 |
| Fat mass, kg | 30.6 ± 7.8 | 14.9 ± 6.3 | <0.001 |
| Lean mass, kg | 58.0 ± 13.8 | 53.6 ± 11.0 | 0.146 |
| Muscle mass, kg | 55.1 ± 13.1 | 50.9 ± 10.5 | 0.157 |
| Bone mass, kg | 2.8 ± 0.6 | 2.6 ± 0.5 | 0.162 |
| Total water, % | 46.6 ± 6.4 | 55.1 ± 8.5 | <0.001 |
| Total water, kg | 41.8 ± 10.3 | 37.8 ± 8.7 | 0.073 |
| Blood parameters | |||
| Fasting glucose, mg/dl | 94.5 ± 10.2 | 80.3 ± 7.2 | <0.001 |
| Total cholesterol, mg/dl | 205.1 ± 30.8 | 179.6 ± 34.6 | <0.001 |
| LDL-cholesterol, mg/dl | 127.4 ± 26.3 | 99.4 ± 33.5 | <0.001 |
| HDL-cholesterol, mg/dl | 52.7 ± 13.0 | 63.1 ± 17.6 | 0.001 |
| Triglycerides, mg/dl | 125.3 ± 56.8 | 85.5 ± 43.0 | <0.001 |
| Energy, Macro- and Micronutrients | Overweight and Obese Group (n = 96) | Lean Group (n = 32) | p Value |
|---|---|---|---|
| Energy, kcal/day | 2055.9 ± 604.3 | 2095.9 ± 502.8 | 0.184 |
| CHO, % energy | 36.0 ± 6.3 | 39.9 ± 6.8 | 0.013 |
| CHO, grams | 181.2 ± 60.9 | 206.0 ± 53.1 | 0.005 |
| Simple CHO, % energy | 15.5 ± 4.8 | 17.5 ± 4.1 | 0.043 |
| Simple CHO, grams | 78.3 ± 35.8 | 92.1 ± 35.9 | 0.014 |
| Complex CHO, % energy | 20.6 ± 5.7 | 22.3 ± 5.1 | 0.301 |
| Complex CHO, grams | 103.8 ± 41.9 | 113.8 ± 24.8 | 0.017 |
| Protein, % energy | 18.1 ± 4.3 | 17.6 ± 3.3 | 0.702 |
| Protein, grams | 90.07 ± 24.9 | 90.3 ± 22.9 | 0.369 |
| Total fat, % energy | 41.6 ± 5.6 | 40.0 ± 7.0 | 0.472 |
| Total fat, grams | 97.4 ± 34.5 | 95.0 ± 33.1 | 0.429 |
| SFA, % energy | 12.4 ± 2.8 | 11.0 ± 2.7 | 0.083 |
| SFA, grams | 29.6 ± 13.1 | 26.5 ± 10.8 | 0.515 |
| MUFA, % energy | 19.0 ± 3.2 | 19.0 ± 4.5 | 0.506 |
| MUFA, grams | 44.0 ± 15.0 | 45.3 ± 17.7 | 0.162 |
| PUFA, % energy | 6.6 ± 2.0 | 6.6 ± 1.8 | 0.710 |
| PUFA, grams | 15.6 ± 7.4 | 15.6 ± 6.0 | 0.247 |
| Fibre, g/day | 18.8 ± 6.8 | 23.7 ± 9.1 | <0.001 |
| Alcohol, g/day | 12.9 ± 18.8 | 7.4 ± 9.2 | 0.327 |
| Dietary cholesterol, mg/day | 391.8 ± 188.5 | 341.1 ± 164.4 | 0.259 |
| Sodium, mg/day | 2580.8 ± 928.6 | 2353.2 ± 728.1 | 0.485 |
| Potassium, mg/day | 3042.4 ± 868.5 | 3448.3 ± 826.2 | 0.002 |
| Calcium, mg/day | 752.9 ± 354.2 | 789.3 ± 339.9 | 0.549 |
| Magnesium, mg/day | 309.5 ± 106.9 | 345.2 ± 74.0 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; del Bas, J.M.; et al. Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients 2021, 13, 2032. https://doi.org/10.3390/nu13062032
Companys J, Gosalbes MJ, Pla-Pagà L, Calderón-Pérez L, Llauradó E, Pedret A, Valls RM, Jiménez-Hernández N, Sandoval-Ramirez BA, del Bas JM, et al. Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients. 2021; 13(6):2032. https://doi.org/10.3390/nu13062032
Chicago/Turabian StyleCompanys, Judit, Maria José Gosalbes, Laura Pla-Pagà, Lorena Calderón-Pérez, Elisabet Llauradó, Anna Pedret, Rosa Maria Valls, Nuria Jiménez-Hernández, Berner Andrée Sandoval-Ramirez, Josep Maria del Bas, and et al. 2021. "Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study" Nutrients 13, no. 6: 2032. https://doi.org/10.3390/nu13062032
APA StyleCompanys, J., Gosalbes, M. J., Pla-Pagà, L., Calderón-Pérez, L., Llauradó, E., Pedret, A., Valls, R. M., Jiménez-Hernández, N., Sandoval-Ramirez, B. A., del Bas, J. M., Caimari, A., Rubió, L., & Solà, R. (2021). Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients, 13(6), 2032. https://doi.org/10.3390/nu13062032