
Molecular Basis of IgE-Mediated Shrimp Allergy and Heat Desensitization



Abstract
:
1. Introduction
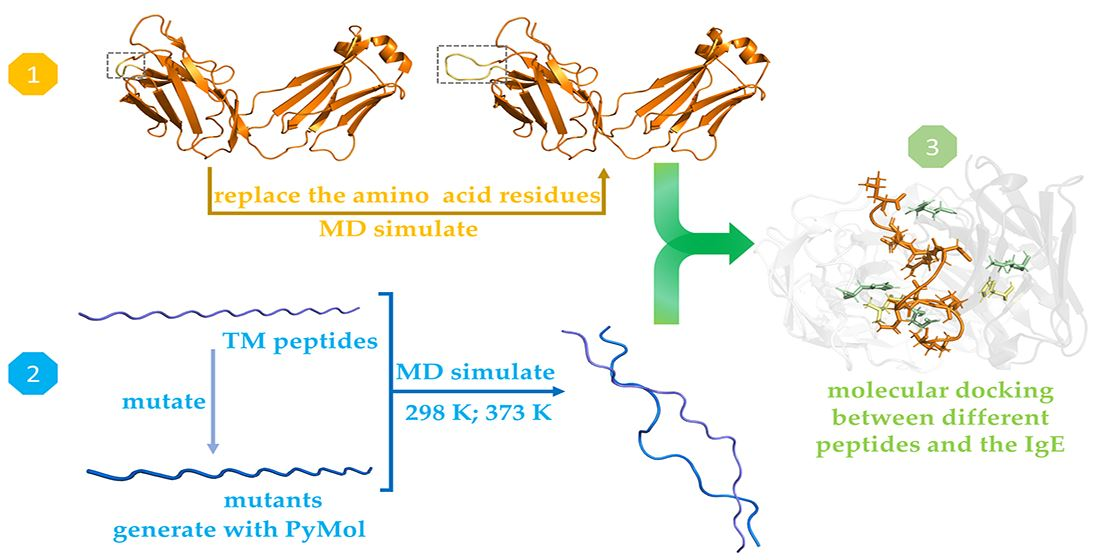
2. Materials and Methods
2.1. Preparation of the Shrimp-Allergic IgE Structure
2.2. Preparation and Heat Treatment of the Allergenic Peptides
2.3. Assessment of the Allergenicity of Heat-Treated Peptides
3. Results
3.1. Sequence Analysis of TM Epitope Peptides
3.2. Heat Treatment on the Conformational Changes of TM Peptides
3.3. IgE Interaction with Native and Heat-Treated TM Peptides
3.4. Critical Amino Acid Residues Involved in IgE Recognition of TM Allergenic Peptides
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandes, T.; Costa, J.; Oliveira, M.; Mafra, I. An overview on fish and shellfish allergens and current methods of detection. Food Agric. Immunol. 2015, 26, 848–869. [Google Scholar] [CrossRef]
- Nwaru, B.; Hickstein, L.; Panesar, S.; Roberts, G.; Muraro, A.; Sheikh, A.; Allergy, E.F.; Group, A.G. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 2014, 69, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Pascal, M.; El Kharbouchi, O.; Sabato, V.; Hagendorens, M.; Decuyper, I.; Bridts, C.; Ebo, D. Shellfish allergens: Tropomyosin and beyond. Allergy 2017, 72, 842–848. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.J.; Lin, Y.F.; Chiang, B.L.; Chow, L.P. Proteomics and immunological analysis of a novel shrimp allergen, Pen m 2. J. Immunol. 2003, 170, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayuso, R.; Grishina, G.; Bardina, L.; Carrillo, T.; Blanco, C.; Ibáñez, M.D.; Sampson, H.A.; Beyer, K. Myosin light chain is a novel shrimp allergen, Lit v 3. J. Allergy Clin. Immunol. 2008, 122, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, R.; Grishina, G.; Ibáñez, M.; Blanco, C.; Carrillo, T.; Sampson, H. Identification and Cloning of a Sarcoplasmic Calcium-binding Protein, a new Shrimp Allergen. J. Allergy Clin. Immunol. 2009, 123, S179. [Google Scholar] [CrossRef]
- Bauermeister, K.; Wangorsch, A.; Garoffo, L.P.; Reuter, A.; Conti, A.; Taylor, S.L.; Lidholm, J.; DeWitt, Å.M.; Enrique, E.; Vieths, S. Generation of a comprehensive panel of crustacean allergens from the North Sea Shrimp Crangon crangon. Mol. Immunol. 2011, 48, 1983–1992. [Google Scholar] [CrossRef]
- Piboonpocanun, S.; Jirapongsananuruk, O.; Tipayanon, T.; Boonchoo, S.; Goodman, R.E. Identification of hemocyanin as a novel non-cross-reactive allergen from the giant freshwater shrimp Macrobrachium rosenbergii. Mol. Nutr. Food Res. Int. 2011, 55, 1492–1498. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.X.; Liu, M.; Maleki, S.J.; Zhang, M.L.; Liu, Q.M.; Cao, M.J.; Su, W.J.; Liu, G.M. Triosephosphate isomerase and filamin C share common epitopes as novel allergens of Procambarus clarkii. J. Agric. Food Chem. 2017, 65, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.C.; Tyan, Y.C.; Yu, W.T.; Huang, E.S.; Yu, H.S. Identification of pyruvate kinase as a novel allergen in whiteleg shrimp (Litopenaeus vannamei) by specific-IgE present in patients with shrimp allergy. Food Chem. 2018, 258, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Gámez, C.; Sánchez García, S.; Ibáñez, M.; López, R.; Aguado, E.; López, E.; Sastre, B.; Sastre, J.; Del Pozo, V. Tropomyosin IgE-positive results are a good predictor of shrimp allergy. Allergy 2011, 66, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.M.; Cheng, H.; Nesbit, J.B.; Su, W.J.; Cao, M.J.; Maleki, S.J. Effects of boiling on the IgE-binding properties of tropomyosin of shrimp (Litopenaeus vannamei). J. Food Sci. 2010, 75, T1–T5. [Google Scholar] [CrossRef] [PubMed]
- Mejrhit, N.; Azdad, O.; Aarab, L. Effect of industrial processing on the IgE reactivity of three commonly consumed Moroccan fish species in Fez region. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, X.; Chen, W.; Zhou, P. Conformation stability, in vitro digestibility and allergenicity of tropomyosin from shrimp (Exopalaemon modestus) as affected by high intensity ultrasound. Food Chem. 2018, 245, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, J.; Raghavan, V. Impact of microwave processing on the secondary structure, in-vitro protein digestibility and allergenicity of shrimp (Litopenaeus vannamei) proteins. Food Chem. 2021, 337, 127811. [Google Scholar] [CrossRef]
- Fu, L.; Song, J.; Wang, C.; Fu, S.; Wang, Y. Bifidobacterium infantis potentially alleviates shrimp tropomyosin-induced allergy by tolerogenic dendritic cell-dependent induction of regulatory T cells and alterations in gut microbiota. Front. Immunol. 2017, 8, 1536. [Google Scholar] [CrossRef]
- Fu, L.; Xie, M.; Wang, C.; Qian, Y.; Huang, J.; Sun, Z.; Zhang, H.; Wang, Y. Lactobacillus Casei Zhang Alleviates Shrimp Tropomyosin-Induced Food Allergy by Switching Antibody Isotypes through the NF-κB-Dependent Immune Tolerance. Mol. Nutr. Food Res. Int. 2020, 64, 1900496. [Google Scholar] [CrossRef]
- Yeşilyurt, N.; Yılmaz, B.; Ağagündüz, D.; Capasso, R. Involvement of Probiotics and Postbiotics in the Immune System Modulation. Biologics 2021, 1, 89–110. [Google Scholar] [CrossRef]
- Wai, C.Y.; Leung, N.Y.; Leung, P.S.; Chu, K.H. Modulating shrimp tropomyosin-mediated allergy: Hypoallergen DNA vaccines induce regulatory T cells to reduce hypersensitivity in mouse model. Int. J. Mol. Sci. 2019, 20, 4656. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Ji, H.; Zhang, L.; Wang, Y.; Zhou, H. Chitosan oligosaccharide exerts anti-allergic effect against shrimp tropomyosin-induced food allergy by affecting Th1 and Th2 cytokines. Int. Arch. Allergy Immunol. 2019, 180, 10–16. [Google Scholar] [CrossRef]
- Sathe, S.K.; Teuber, S.S.; Roux, K.H. Effects of food processing on the stability of food allergens. Biotechnol. Adv. 2005, 23, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, C.; Wang, J.; Ni, S.; Wang, Y. Maillard reaction with ribose, galacto-oligosaccharide or chitosan-oligosaccharide reduced the allergenicity of shrimp tropomyosin by inducing conformational changes. Food Chem. 2019, 274, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ni, S.; Wang, C.; Li, X.; Fu, L. Cross-linking of shrimp tropomyosin catalyzed by transglutaminase and tyrosinase produces hypoallergens for potential immunotherapy. Food Funct. 2019, 10, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, J.; Ni, S.; Wang, C.; Wang, Y. Identification of Allergenic Epitopes and Critical Amino Acids of Major Allergens in Chinese Shrimp ( Penaeus chinensis) by Immunoinformatics Coupled with Competitive-Binding Strategy. J. Agric. Food Chem. 2018, 66, 2944–2953. [Google Scholar] [CrossRef]
- Niemi, M.; Jylhä, S.; Laukkanen, M.L.; Söderlund, H.; Mäkinen-Kiljunen, S.; Kallio, J.M.; Hakulinen, N.; Haahtela, T.; Takkinen, K.; Rouvinen, J. Molecular interactions between a recombinant IgE antibody and the β-lactoglobulin allergen. Structure 2007, 15, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Yu, P.; Liang, H.; Fu, J.; Luo, Z.; Yang, D. The wPDI Redox Cycle Coupled Conformational Change of the Repetitive Domain of the HMW-GS 1Dx5-A Computational Study. Molecules 2020, 25, 4393. [Google Scholar] [CrossRef]
- Chu, K.H.; Wong, S.H.; Leung, P.S. Tropomyosin is the major mollusk allergen: Reverse transcriptase polymerase chain reaction, expression and IgE reactivity. Mar. Biotechnol. 2000, 2, 499–509. [Google Scholar] [CrossRef]
- Ayuso, R.; Reese, G.; Leong Kee, S.; Plante, M.; Lehrer, S.B. Molecular basis of arthropod cross-reactivity: IgE-binding cross-reactive epitopes of shrimp, house dust mite and cockroach tropomyosins. Int. Arch. Allergy Immunol. 2002, 129, 38–48. [Google Scholar] [CrossRef]
- Reese, G.; Viebranz, J.; Leong-Kee, S.M.; Plante, M.; Lauer, I.; Randow, S.; Moncin, M.S.-M.; Ayuso, R.; Lehrer, S.B.; Vieths, S. Reduced allergenic potency of VR9-1, a mutant of the major shrimp allergen Pen a 1 (tropomyosin). J. Immunol. 2005, 175, 8354–8364. [Google Scholar] [CrossRef]
- Leung, P.S.; Chu, K.H.; Chow, W.K.; Ansari, A.; Bandea, C.I.; Kwan, H.S.; Nagy, S.M.; Gershwin, M.E. Cloning, expression, and primary structure of Metapenaeus ensis tropomyosin, the major heat-stable shrimp allergen. J. Allergy Clin. Immunol. 1994, 94, 882–890. [Google Scholar] [CrossRef]
- Rahman, A.M.A.; Kamath, S.; Lopata, A.L.; Helleur, R.J. Analysis of the allergenic proteins in black tiger prawn (Penaeus monodon) and characterization of the major allergen tropomyosin using mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2462–2470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, W.; Zhang, R.; Zhao, X.; Fang, L.; Qin, X.; Gu, R.; Lu, J.; Li, G. Hypoallergenic mutants of the major oyster allergen Cra g 1 alleviate oyster tropomyosin allergenic potency. Int. J. Biol. Macromol. 2020, 164, 1973–1983. [Google Scholar] [CrossRef]
- Mykles, D.L.; Cotton, J.L.; Taniguchi, H.; Sano, K.i.; Maeda, Y. Cloning of tropomyosins from lobster (Homarus americanus) striated muscles: Fast and slow isoforms may be generated from the same transcript. J. Muscle Res. Cell Motil. 1998, 19, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Medler, S.; Lilley, T.; Mykles, D.L. Fiber polymorphism in skeletal muscles of the American lobster, Homarus americanus: Continuum between slow-twitch (S1) and slow-tonic (S2) fibers. J. Exp. Biol. 2004, 207, 2755–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, P.S.; Chen, Y.C.; Mykles, D.L.; Chow, W.K.; Li, C.P.; Chu, K.H. Molecular identification of the lobster muscle protein tropomyosin as a seafood allergen. Mol. Mar. Biol. Biotechnol. Adv. 1998, 7, 12–20. [Google Scholar] [CrossRef]
- Leung, P.S.; Chen, Y.c.; Gershwin, M.E.; Wong, S.H.; Kwan, H.S.; Chu, K.H. Identification and molecular characterization of Charybdis feriatus tropomyosin, the major crab allergen. J. Allergy Clin. Immunol. 1998, 102, 847–852. [Google Scholar] [CrossRef]
- Motoyama, K.; Suma, Y.; Ishizaki, S.; Nagashima, Y.; Shiomi, K. Molecular cloning of tropomyosins identified as allergens in six species of crustaceans. J. Agric. Food Chem. 2012, 55, 985–991. [Google Scholar] [CrossRef]
- Daul, C.B.; Morgan, J.E.; Waring, N.P.; McCants, M.L.; Hughes, J.; Lehrer, S.B. Immunologic evaluation of shrimp-allergic individuals. J. Allergy Clin. Immunol. 1987, 80, 716–722. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Yuan, F.; Gao, Q.; Ma, J. Effect of 4-hydroxy-2-nonenal treatment on the IgE binding capacity and structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem. 2016, 212, 313–322. [Google Scholar] [CrossRef]
- Leung, N.Y.; Wai, C.Y.; Ho, M.H.; Liu, R.; Lam, K.S.; Wang, J.J.; Shu, S.A.; Chu, K.H.; Leung, P.S. Screening and identification of mimotopes of the major shrimp allergen tropomyosin using one-bead-one-compound peptide libraries. Cell. Mol. Immunol. 2017, 14, 308–318. [Google Scholar] [CrossRef] [Green Version]
- James, J.K.; Pike, D.H.; Khan, I.J.; Nanda, V. Structural and Dynamic Properties of Allergen and Non-Allergen Forms of Tropomyosin. Structure 2018, 26, 997–1006.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Fernández, J.; Rodero, M.; Daschner, A.; Cuéllar, C. New insights into the allergenicity of tropomyosin: A bioinformatics approach. Mol. Biol. Rep. 2014, 41, 6509–6517. [Google Scholar] [CrossRef] [PubMed]

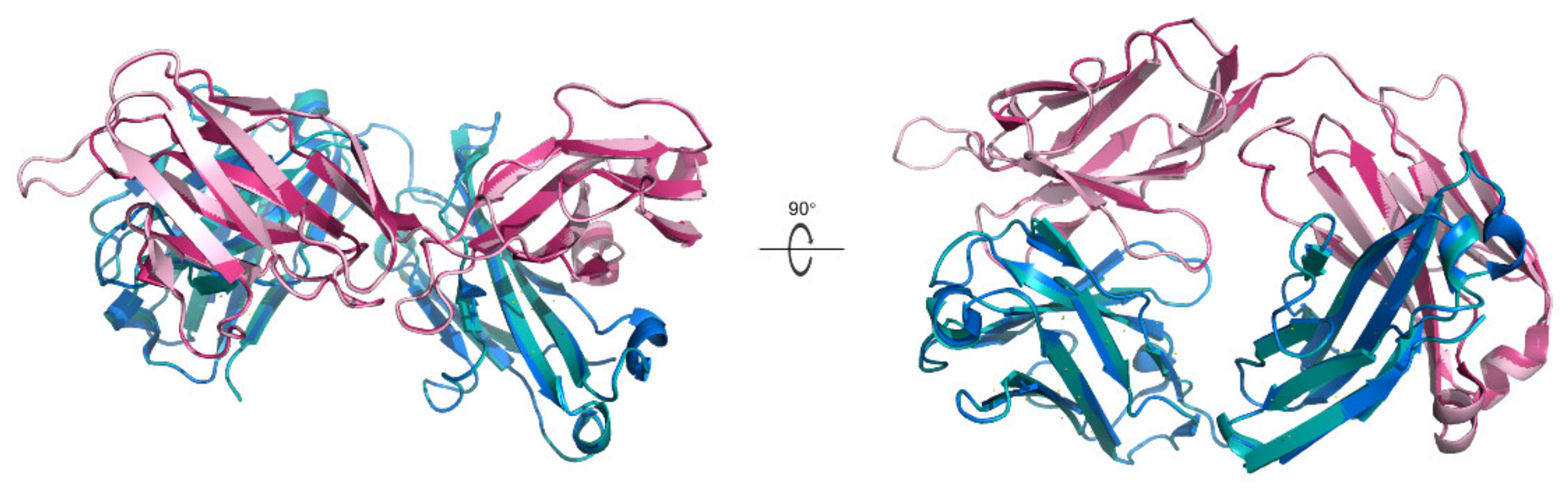


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide 1 | Sequence | Position 2 | Mutation 3 |
|---|---|---|---|
| T1 | KLEKDNAMDRADTL | 12−25 | |
| T3 | QKRMQQLENDLDQV | 47−60 | |
| T5 | ALNRRIQLLEEDLERSEER | 87−105 | |
| T6 | KLAEASQAADESERMRK | 112−128 | |
| T7 | ENRSLSDEERMDALENQ | 131−147 | |
| T7-E131 | ANRSLSDEERMDALENQ | E131 | |
| T7-R140 | ENRSLSDEEAMDALENQ | R140 | |
| T7-D142 | ENRSLSDEERMAALENQ | D142 | |
| T7-Q147 | ENRSLSDEERMDALENA | Q147 | |
| T8 | AEEADRKYDEVAR | 155−167 | |
| T9 | ESKIVELEEELRVVGNN | 187−203 | |
| T10 | YKEQIKTLTNKLKAAEARA | 221−239 | |
| T11 | ERSVQKLQKEVDRLEDE | 243−259 |
| Peptide | RMSD (Å) 1 | ΔSASA (Å2) 2 | Terminal Distance (Å) 3 | Number of H-Bonds 4 | ||
|---|---|---|---|---|---|---|
| Native | Heated | Native | Heated | |||
| T1 | 2.469 | 11.40 | 38.0 | 31.2 | 5 | 5 |
| T3 | 3.376 | −1.57 | 37.7 | 39.1 | 2 | 5 |
| T5 | 2.542 | 4.79 | 57.1 | 54.9 | 4 | 4 |
| T6 | 2.680 | 37.02 | 49.9 | 49.5 | 1 | 4 |
| T7 | 3.278 | −149.64 | 40.6 | 36.6 | 1 | 3 |
| T8 | 2.880 | −29.47 | 34.1 | 33.0 | 2 | 2 |
| T9 | 2.570 | 35.04 | 44.6 | 46.1 | 1 | 1 |
| T10 | 3.448 | −106.49 | 55.3 | 51.2 | 0 | 2 |
| T11 | 2.190 | −10.65 | 45.7 | 38.3 | 8 | 11 |
| Peptide | Number of H-Bonds 1 | Number of Hydrophobic Interactions 2 | Binding Energy (kcal/mol) | RMSD (Å) 3 | ΔSASA (Å2) 4 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Native | Heated | Native | Heated | Native | Heated | Native | Heated | Native | Heated | |
| T1 | 14 | 5 | 64 | 35 | −17.28 | −7.83 | 6.62 | 5.61 | −171.37 | −255.80 |
| T3 | 12 | 6 | 58 | 64 | −12.16 | −21.11 | 5.47 | 3.69 | −209.82 | −176.00 |
| T5 | 8 | 9 | 52 | 73 | −19.01 | −18.21 | 6.11 | 7.27 | −89.79 | −313.73 |
| T6 | 9 | 6 | 46 | 46 | −2.89 | −12.53 | 9.03 | 5.62 | −82.85 | −0.50 |
| T7 | 12 | 11 | 62 | 72 | −24.16 | −2.56 | 5.06 | 4.45 | −165.80 | −170.14 |
| T8 | 8 | 5 | 59 | 42 | −10.19 | −0.084 | 7.38 | 6.63 | −89.50 | −77.24 |
| T9 | 4 | 5 | 54 | 59 | −4.52 | −1.53 | 5.72 | 5.15 | −121.52 | −119.43 |
| T10 | 7 | 15 | 42 | 85 | −10.08 | −20.27 | 4.10 | 5.75 | −229.70 | −189.70 |
| T11 | 8 | 4 | 62 | 47 | −30.49 | −3.10 | 7.23 | 3.17 | −69.17 | −87.17 |
| Peptide | RMSD 1 (Å) | Terminal Distance 2 (Å) | Number of H-Bonds 3 | Binding Energy (kcal/mol) | ΔSASA (Å2) 4 |
|---|---|---|---|---|---|
| T7-WT | - | 57.1 | 1 | −24.16 | −165.80 |
| T7-E131 | 2.283 | 41.5 | 7 | −12.49 | −228.30 |
| T7-R140 | 3.641 | 41.1 | 7 | −12.84 | −231.39 |
| T7-D142 | 3.294 | 44.9 | 11 | −14.05 | −108.21 |
| T7-Q147 | 3.303 | 37.9 | 13 | −17.91 | −101.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Gao, J.; Che, H.; Xue, W.; Yang, D. Molecular Basis of IgE-Mediated Shrimp Allergy and Heat Desensitization. Nutrients 2021, 13, 3397. https://doi.org/10.3390/nu13103397
Zhang P, Gao J, Che H, Xue W, Yang D. Molecular Basis of IgE-Mediated Shrimp Allergy and Heat Desensitization. Nutrients. 2021; 13(10):3397. https://doi.org/10.3390/nu13103397
Chicago/Turabian StyleZhang, PeiAo, Jihui Gao, Huilian Che, Wentong Xue, and Dong Yang. 2021. "Molecular Basis of IgE-Mediated Shrimp Allergy and Heat Desensitization" Nutrients 13, no. 10: 3397. https://doi.org/10.3390/nu13103397
APA StyleZhang, P., Gao, J., Che, H., Xue, W., & Yang, D. (2021). Molecular Basis of IgE-Mediated Shrimp Allergy and Heat Desensitization. Nutrients, 13(10), 3397. https://doi.org/10.3390/nu13103397