Extract from Aronia melanocarpa L. Berries Prevents Cadmium-Induced Oxidative Stress in the Liver: A Study in A Rat Model of Low-Level and Moderate Lifetime Human Exposure to this Toxic Metal

 , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cd Diets
2.3. A. Melanocarpa Extract
2.4. Animal Model
2.5. Determination of the Oxidative/Antioxidative Status of the Liver
2.5.1. Preparation of Homogenates of the Liver Tissue
2.5.2. Determination of the Parameters of the Oxidative/Antioxidative Status and the Level of Oxidative Stress
2.6. Estimation of the Liver Morphology and the Activities of Liver Enzymes Markers in the Serum
2.7. Statistical Analysis
3. Results
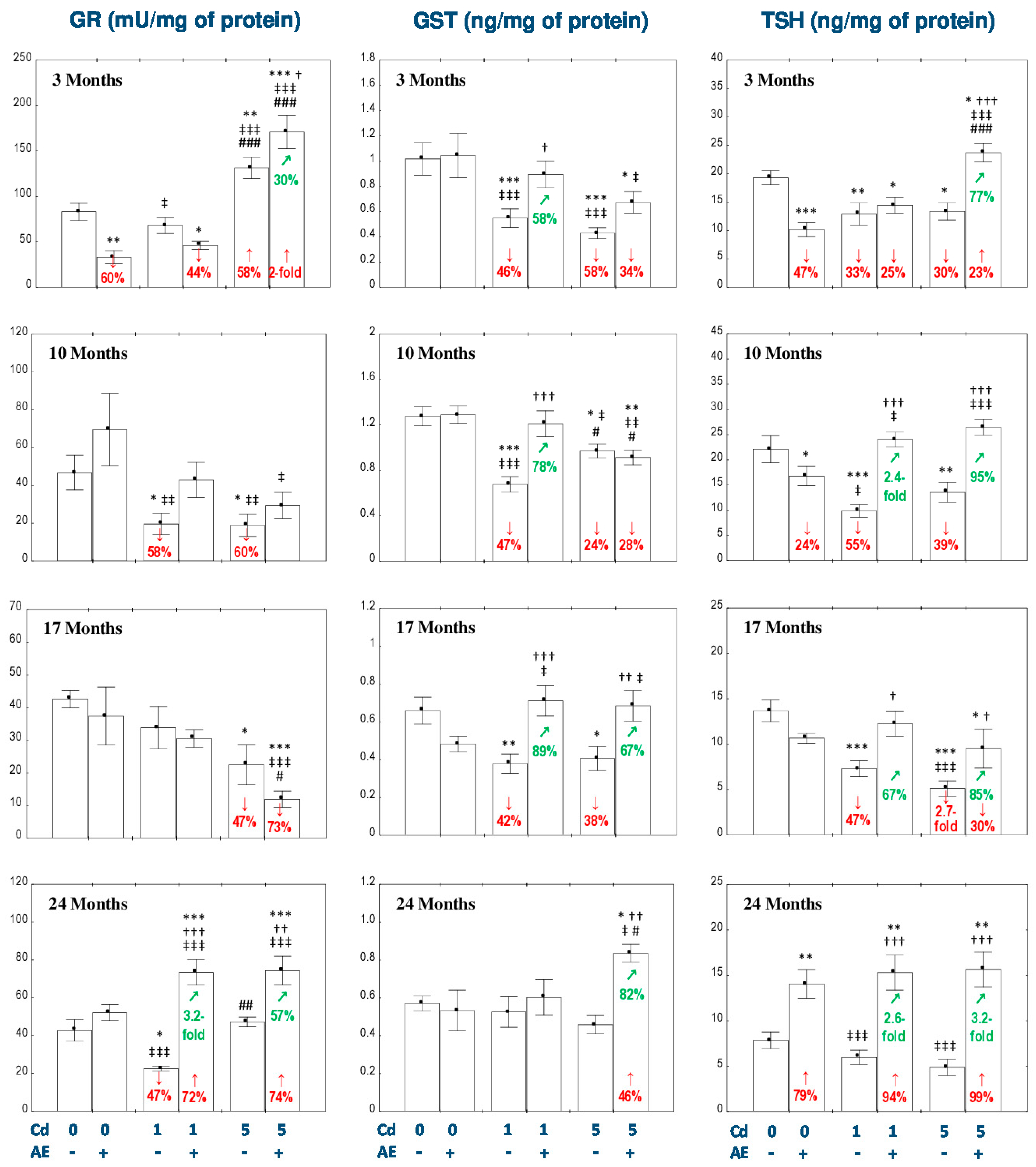
3.1. Effect of AE and/or Cd on the Enzymatic Antioxidative Barrier of the Liver
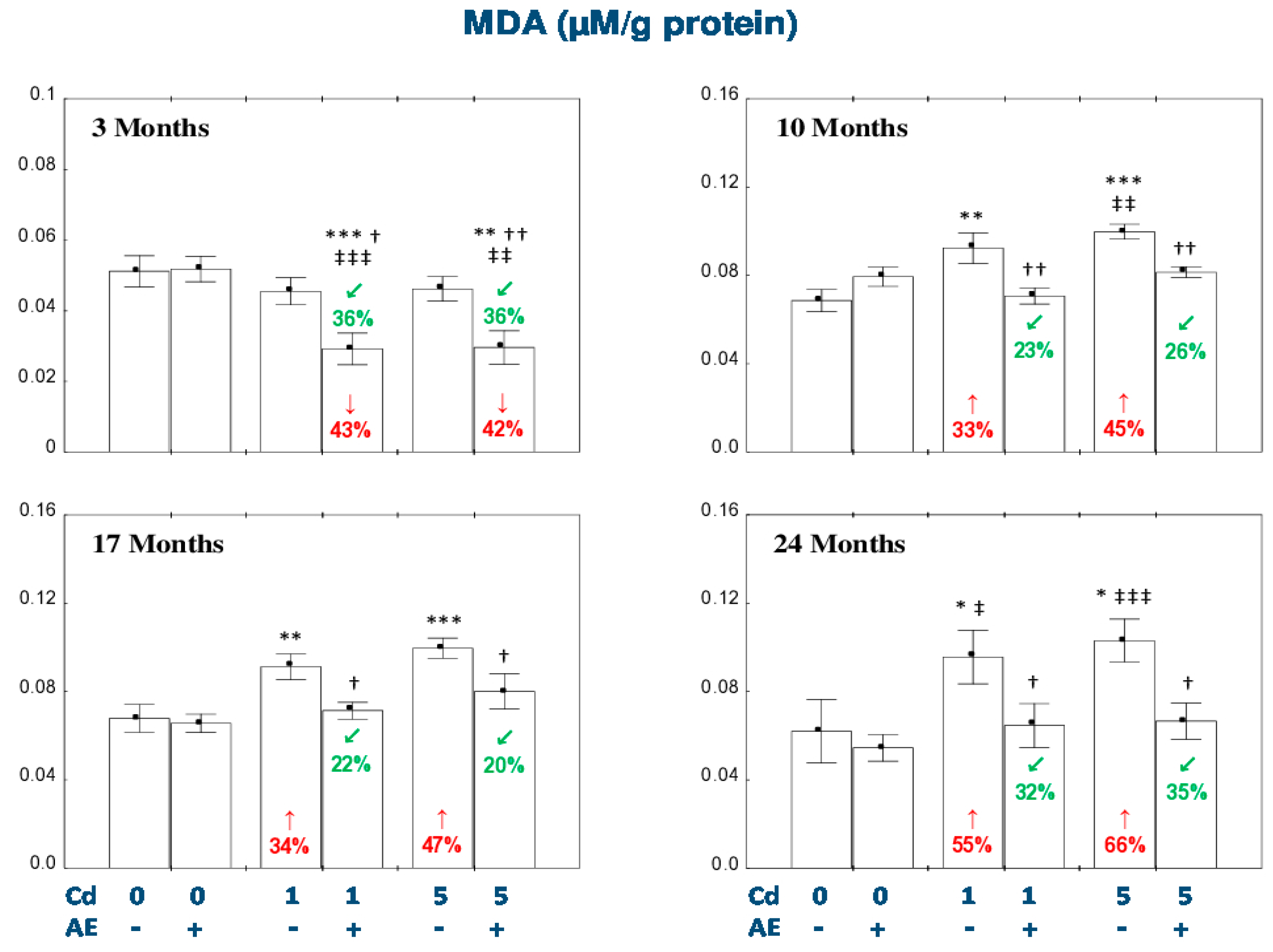
3.2. Effect of AE and/or Cd on the Nonenzymatic Antioxidative Barrier of the Liver
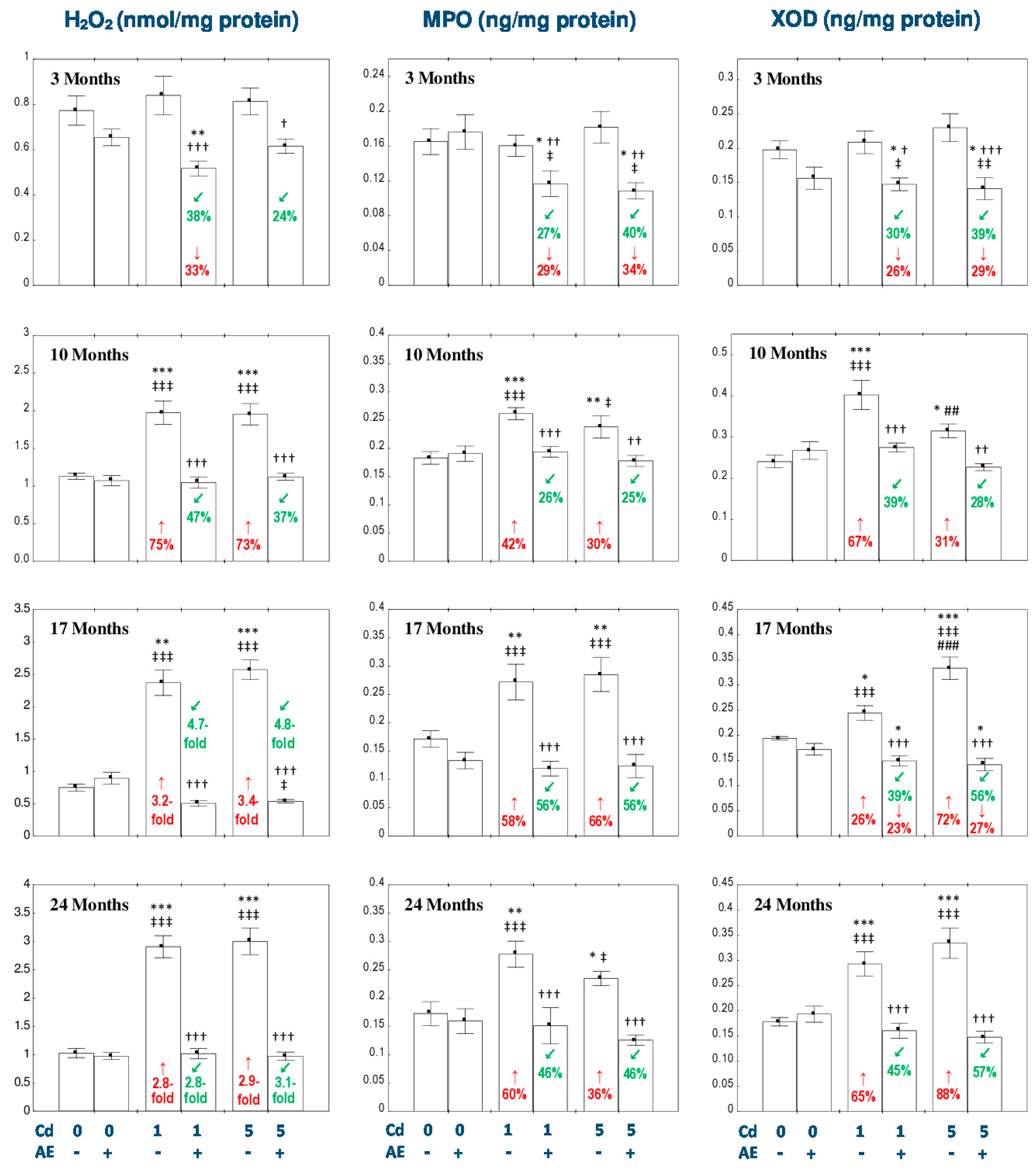
3.3. Effect of AE and/or Cd on the Concentration of H2O2 in the Liver
3.4. Effect of AE and/or Cd on the Concentrations of MPO and XOD in the Liver
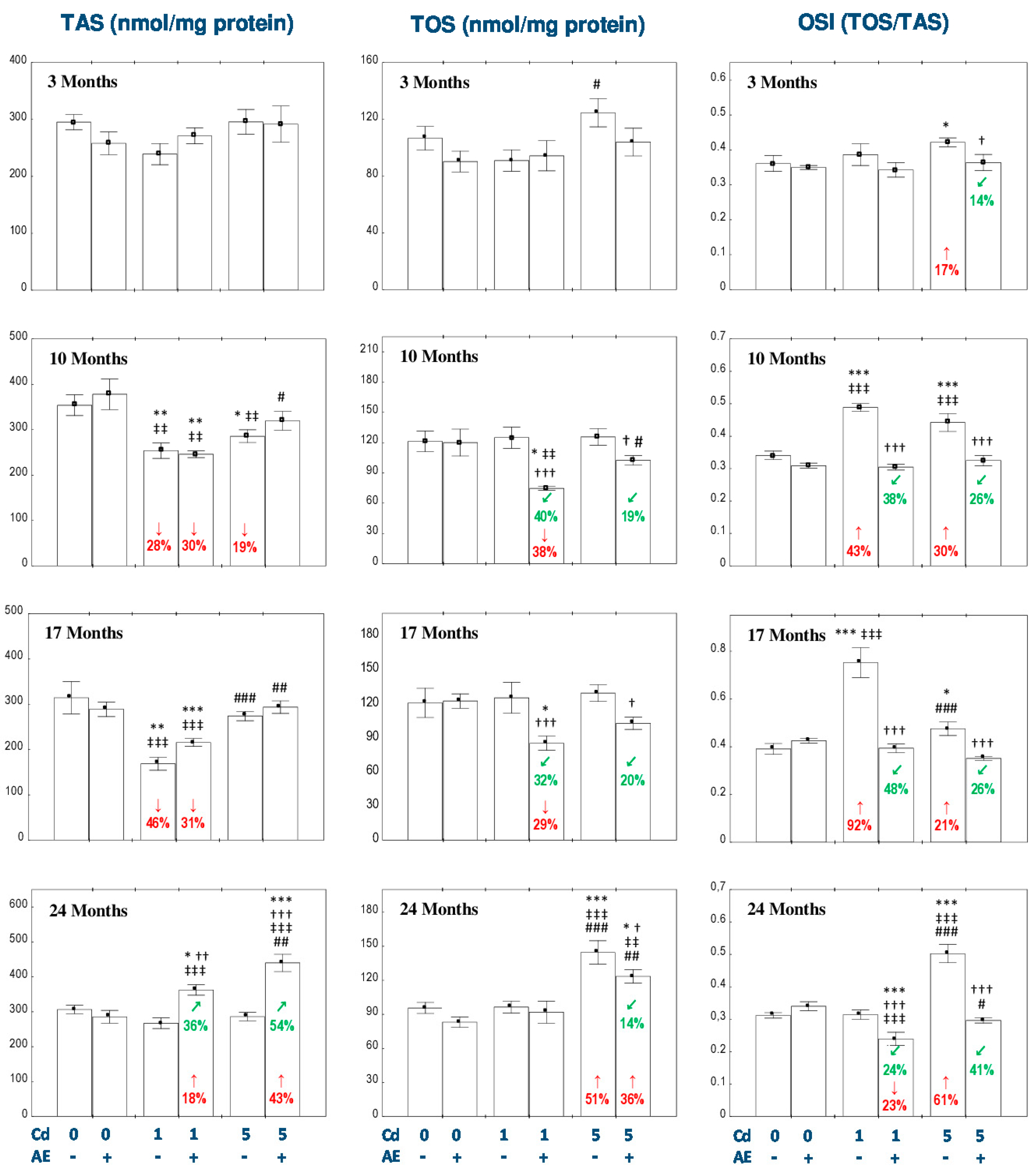
3.5. Effect of AE and/or Cd on TAS and TOS and the Level of Oxidative Stress in the Liver
3.6. Morphological Structure of the Liver
3.7. Effect of AE and/or Cd on the Activities of ALT and AST in the Serum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AE | extract from the berries of Aronia melanocarpa L. |
| ALT | alanine aminotransferase |
| ANOVA | a one-way analysis of variance |
| ANOVA/MANOVA | a two-way analysis of variance |
| AST | aspartate transaminase |
| CAT | catalase |
| Cd | cadmium |
| Cd2+ | divalent ion of cadmium |
| Cu | copper |
| DTNB | 5,5’-dithio-bis-2-nitrobenzoic acid |
| ELISA | enzyme-linked immunosorbent assay |
| FR | free radicals |
| GPx | glutathione peroxidase |
| GR | glutathione reductase |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| GST | glutathione S-transferase |
| H + E | hematoxylin and eosin |
| H2O2 | hydrogen peroxide |
| MDA | malondialdehyde |
| MPO | myeloperoxidase |
| MT | metallothionein |
| NADPH | reduced nicotinamide adenine dinucleotide phosphate |
| NADP+ | oxidized nicotinamide adenine dinucleotide phosphate |
| -OH group | hydroxyl group |
| OSI | oxidative stress index |
| O2·- | superoxide radicals |
| r | correlation coefficient |
| ROS | reactive oxygen species |
| SD | standard deviation |
| SE | standard error |
| SOD | superoxide dismutase |
| -SH group | thiol group |
| TAS | total antioxidative status |
| TNT | 5-thio-2-nitrobenzoic acid |
| Trx | thioredoxin |
| TOS | total oxidative status |
| TSH | total thiol groups |
| XOD | xanthine oxidase |
| Zn | zinc |
References
- Mężyńska, M.; Brzóska, M.M. Review of polyphenol-rich products as potential protective and therapeutic factors against cadmium hepatotoxicity. J. Appl. Toxicol. 2018, 39, 117–145. [Google Scholar] [CrossRef] [PubMed]
- Mężyńska, M.; Brzóska, M.M. Environmental exposure to cadmium—A risk for health of the general population in industrialized countries and preventive strategies. Environ. Sci. Pollut. Res. 2018, 25, 3211–3232. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, M.M.; Borowska, S.; Tomczyk, M. Antioxidants as a potential preventive and therapeutic strategy for cadmium. Curr. Drug Targets 2016, 17, 1350–1384. [Google Scholar] [CrossRef] [PubMed]
- Borowska, S.; Brzóska, M.M. Chokeberries (Aronia melanocarpa) and their products as a possible means for the prevention and treatment of noncommunicable diseases and unfavorable health effects due to exposure to xenobiotics. Compr. Rev. Food Sci. Food Saf. 2016, 15, 982–1017. [Google Scholar] [CrossRef]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavova, J. Fruits of black chokeberry Aronia melanocarpa in the prevention of chronic diseases. Molecules 2017, 22, 944. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.S.; Staessen, J.A.; Roels, H.A. Cadmium exposure in the population: From health risks to strategies of prevention. Biometals 2010, 23, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Health risk assessment of dietary cadmium intake: Do current guidelines indicate how much is safe? Environ. Health Perspect. 2017, 125, 284–288. [Google Scholar] [CrossRef]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Current health risk assessment practice for dietary cadmium: Data from different countries. Food Chem. Toxicol. 2017, 106, 430–445. [Google Scholar] [CrossRef]
- Satarug, S.; Swaddiwudhipong, W.; Ruangyuttikarn, W.; Nishijo, M.; Ruiz, P. Modeling cadmium exposures in low- and high-exposure areas in Thailand. Environ. Health Perspect. 2013, 121, 531–536. [Google Scholar] [CrossRef]
- Gałażyn-Sidorczuk, M.; Brzóska, M.M.; Moniuszko-Jakoniuk, J. Estimation of Polish cigarettes contamination with cadmium and lead, and exposure to these metals via smoking. Environ. Monit. Assess. 2008, 137, 481–493. [Google Scholar] [CrossRef]
- Wang, D.; Sun, H.; Wu, Y.; Zhou, Z.; Ding, Z.; Chen, X.; Xu, Y. Tubular and glomerular kidney effects in the Chinese general population with low environmental cadmium exposure. Chemosphere 2016, 147, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Wallin, M.; Barregard, L.; Sallsten, G.; Lundh, T.; Karlsson, M.K.; Lorentzon, M.; Ohlsson, C.; Mellström, D. Low-level cadmium exposure is associated with decreased bone mineral density and increased risk of incident fractures in elderly men: The MrOSSweden Study. J. Bone Miner. Res. 2016, 31, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, B.; Barregard, L.; Sallsten, G.; Forsgard, N.; Ostling, G.; Persson, M.; Borné, Y.; Engström, G.; Hedblad, B. Cadmium exposure and atherosclerotic carotid plaques—Results from the Malmö diet and cancer study. Environ. Res. 2015, 136, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.W.; Schaumberg, D.A.; Park, S.K. Environmental cadmium and lead exposures and age-related macular degeneration in U.S. adults: The National Health and Nutrition Examination Survey 2005 to 2008. Environ. Res. 2014, 133, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Hu, H.; Mukherjee, B.; Miller, J.; Park, S.K. Environmental cadmium and lead exposures and hearing loss in U.S. adults: The National Health and Nutrition Examination Survey, 1999 to 2004. Environ. Health Perspect. 2012, 120, 1544–1550. [Google Scholar] [CrossRef]
- Chen, C.; Xun, P.; Nishijo, M.; Sekikawa, A.; He, K. Cadmium exposure and risk of pancreatic cancer: A meta-analysis of prospective cohort studies and case-control studies among individuals without occupational exposure history. Environ. Sci. Pollut. Res. 2015, 22, 17465–17474. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, F.; Lei, Y. Dietary intake and urinary level of cadmium and breast cancer risk: A meta-analysis. Cancer Epidemiol. 2016, 42, 101–107. [Google Scholar] [CrossRef]
- Hyder, O.; Chung, M.; Cosgrove, D.; Herman, J.M.; Li, Z.; Firoozmand, A.; Gurakar, A.; Koteish, A.; Pawlik, T.M. Cadmium exposure and liver disease among US adults. J. Gastrointest. Surg. 2013, 17, 1265–1273. [Google Scholar] [CrossRef]
- Kang, M.Y.; Cho, S.H.; Lim, Y.H.; Seo, J.C.; Hong, Y.C. Effects of environmental cadmium exposure on liver function in adults. Occup. Environ. Med. 2013, 70, 268–273. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Zobeiri, M.; Parvizi, F.; El-Senduny, F.F.; Marmouzi, I.; Coy-Barrera, E.; Naseri, R.; Nabavi, S.M.; Rahimi, R.; Abdollahi, M. Curcumin in liver diseases: A systematic review of the cellular mechanisms of oxidative stress and clinical perspective. Nutrients 2018, 10, 855. [Google Scholar] [CrossRef]
- Brea, A.; Pintó, X.; Ascaso, J.S.; Blasco, M.; Díaz, A.; González-Santos, P.; Hernández-Mijares, A.; Mantilla, T.; Millán, J.; Pedro-Botet, J. Nonalcoholic fatty liver disease, association with cardiovascular disease and treatment (II). The treatment of nonalcoholic fatty liver disease. Clín. Investig. Arterioscler. 2017, 29, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Chenb, F.; Wang, L.; Wang, J.; Jina, S.; Ma, Y. Protective effects of blueberries (Vaccinium corymbosum L.) extract against cadmium-induced hepatotoxicity in mice. Environ. Toxicol. Pharmacol. 2014, 37, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, R.; Elangovan, P.; Pari, L. Protective role of tetrahydrocurcumin: An active polyphenolic curcuminoid on cadmium-induced oxidative damage in rats. Appl. Biochem. Biotechnol. 2017, 183, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Elkhadragy, M.F.; Abdel Moneim, A.E. Protective effect of Fragaria ananassa methanolic extract on cadmium chloride (CdCl2)-induced hepatotoxicity in rats. Toxicol. Mech. Methods. 2017, 27, 335–345. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Rogalska, J.; Roszczenko, A.; Gałażyn-Sidorczuk, M.; Tomczyk, M. The mechanism of the osteoprotective action of a polyphenol-rich Aronia melanocarpa extract during chronic exposure to cadmium is mediated by the oxidative defense system. Planta Med. 2016, 82, 621–631. [Google Scholar] [CrossRef]
- Borowska, S.; Brzóska, M.M.; Gałażyn-Sidorczuk, M.; Rogalska, J. Effect of an extract from Aronia melanocarpa L. berries on the body status of zinc and copper under chronic exposure to cadmium: An in vivo experimental study. Nutrients 2017, 9, 1374. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Gałażyn-Sidorczuk, M.; Jurczuk, M.; Tomczyk, M. Protective effect of Aronia melanocarpa polyphenols on cadmium accumulation in the body: A study in a rat model of human exposure to this metal. Curr. Drug Targets 2015, 16, 1470–1487. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Jurczuk, M.; Roszczenko, A.; Tomczyk, M. Protective effect of Aronia melanocarpa polyphenols against cadmium-induced disorders in bone metabolism: A study in a rat model of lifetime human exposure to this heavy metal. Chem. Biol. Interact. 2015, 229, 132–146. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Roszczenko, A.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Mężyńska, M. Protective effect of chokeberry (Aronia melanocarpa l.) extract against cadmium impact on the biomechanical properties of the femur: A study in a rat model of low and moderate lifetime women exposure to this heavy metal. Nutrients 2017, 9, 543. [Google Scholar] [CrossRef]
- Kowalczyk, E.; Kopff, A.; Fijalkowski, P.; Kopff, M.; Niedworok, J.; Blaszczyk, J.; Kedziora, J.; Tyslerowicz, P. Effect of anthocyanins on selected biochemical parameters in rats exposed to cadmium. Acta Biochim. Pol. 2003, 50, 543–548. [Google Scholar] [PubMed]
- Dei Cas, M.; Ghidoni, R. Cancer prevention and therapy with polyphenols: Sphingolipid-mediated mechanisms. Nutrients 2018, 10, 940. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.P.; Dong, X.J.; Ma, H.H. Antioxidative and chelating properties of anthocyanins in Azolla umbricata induced by cadmium. Pol. J. Environ. Stud. 2012, 21, 837–844. [Google Scholar]
- Borowska, S.; Brzoska, M.M.; Tomczyk, M. Complexation of bioelements and toxic metals by polyphenolic compounds—Implications for health. Curr. Drug Targets 2018, 19, 1612–1638. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.D. Liver function: Test selection and interpretation of results. Clin. Lab. Med. 2002, 22, 377–390. [Google Scholar] [CrossRef]
- Şahin, E.; Gümüşlü, S. Immobilization stress in rat tissues: Alterations in protein oxidation, lipid peroxidation and antioxidant defense system. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2007, 144, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H.E. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Gałażyn-Sidorczuk, M.; Brzóska, M.M.; Rogalska, J.; Roszczenko, A.; Jurczuk, M. Effect of zinc supplementation on glutathione peroxidase activity and selenium concentration in the serum, liver and kidney of rats chronically exposed to cadmium. J. Trace Elem. Med. Biol. 2012, 26, 46–52. [Google Scholar] [CrossRef]
- El-Boshy, M.E.; Risha, E.F.; Abdelhamid, F.M.; Mubarak, M.S.; Hadda, T.B. Protective effects of selenium against cadmium induced hematological disturbances, immunosuppressive, oxidative stress and hepatorenal damage in rats. J. Trace. Elem. Med. Biol. 2015, 29, 104–110. [Google Scholar] [CrossRef]
- Rogalska, J.; Pilat-Marcinkiewicz, B.; Brzóska, M.M. Protective effect of zinc against cadmium hepatotoxicity depends on this bioelement intake and level of cadmium exposure: A study in a rat model. Chem. Biol. Interact. 2011, 193, 191–203. [Google Scholar] [CrossRef]
- Li, X.; Jiang, X.; Sun, J.; Zhu, C.; Li, X.; Tian, L.; Liu, L.; Bai, W. Cytoprotective effects of dietary flavonoids against cadmium-induced toxicity. Ann. N. Y. Acad. Sci. 2017, 1398, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Haouem, S.; El Hani, A. Effect of cadmium on lipid peroxidation and on some antioxidants in the liver, kidneys and testes of rats given diet containing cadmium-polluted radish bulbs. J. Toxicol. Pathol. 2013, 26, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Renugadevi, J.; Prabu, S.M. Cadmium-induced hepatotoxicity in rats and the protective effect of naringenin. Exp. Toxicol. Pathol. 2010, 62, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Refaie, M.M.M.; El-Hussieny, M.; Zenhom, N.M. Protective role of nebivolol in cadmium-induced hepatotoxicity via downregulation of oxidative stress, apoptosis and inflammatory pathways. Environ. Toxicol. Pharmacol. 2018, 58, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Njoku, R.C.; Lawrence, C.J.; Charles, I.A.; Ikewuchi, J.C. Rutin ameliorates oxidative stress and preserves hepatic and renal functions following exposure to cadmium and ethanol. Pharm. Biol. 2017, 55, 2161–2169. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, Y.S.; El-Gazzar, A.M.; El-Nahas, A.F.; Ashry, K.M. Vitamin C modulates cadmium-induced hepatic antioxidants’ gene transcripts and toxicopathic changes in Nile tilapia, Oreochrom isniloticus. Environ. Sci. Pollut. Res. 2016, 23, 1664–1670. [Google Scholar] [CrossRef] [PubMed]
- Adi, P.D.; Burra, S.P.; Vataparti, A.R.; Matcha, B. Calcium, zinc and vitamin E ameliorate cadmium-induced renal oxidative damage in albino Wistar rats. Toxicol. Rep. 2016, 3, 591–597. [Google Scholar] [CrossRef]
- Nawirska, A. Binding of heavy metals to pomace fibers. Food Chem. 2005, 90, 395–400. [Google Scholar] [CrossRef]
- Borycka, B. Frakcje włókna pokarmowego z wytłoków aroniowych w relacjach z jonami Pb i Cd oraz Ca i Mg. ŻNTJ 2012, 6, 31–40. (In Polish) [Google Scholar]
- Smith, G.J.; Thomsen, S.J.; Markham, R.R.; Andary, C.; Cardon, D. The photostabilities of naturally occurring 5-hydroxyflavones, flavonols, their glycosides and their aluminium complexes. J. Photochem. Photobiol. A Chem. 2000, 136, 87–91. [Google Scholar] [CrossRef]
- Hamden, K.; Carreau, S.; Marki, F.A.; Masmoudi, H.; El Feki, A. Positive effects of green tea on hepatic dysfunction, lipid peroxidation and antioxidant defence depletion induced by cadmium. Biol. Res. 2008, 41, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Winiarska-Mieczan, A. The potential protective effect of green, black, red and white tea infusions against adverse effect of cadmium and lead during chronic exposure—A rat model study. Regul. Toxicol. Pharmacol. 2015, 73, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Baiomy, A.A.; Mansour, A.A. Genetic and histopathological responses to cadmium toxicity in rabbit’s kidney and liver: Protection by ginger (Zingiber officinale). Biol. Trace. Elem. Res. 2016, 170, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Amamou, F.; Nemmiche, S.; Meziane, R.K.; Didi, A.; Yazit, S.A.; Chabane-Sari, D. Protective effect of olive oil and colocynth oil against cadmium-induced oxidative stress in the liver of Wistar rats. Food Chem. Toxicol. 2015, 78, 177–184. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
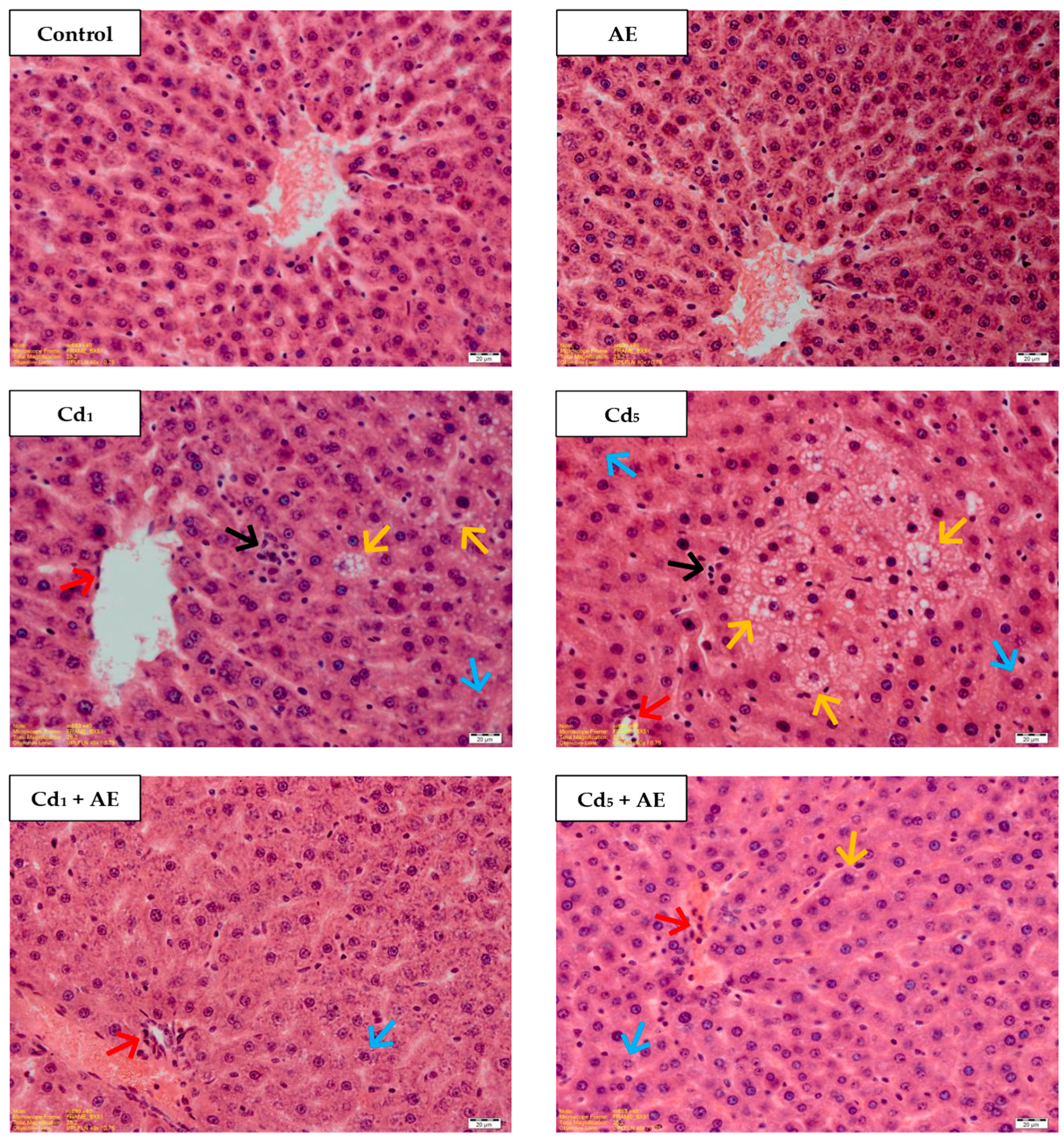{kind=link}
| Group | Blurred Trabecular Structure of the Lobes | Microvascular Steatosis | Colliquative Necrosis | Vacuolization, Enlarged Dimensions of Cells | Mononuclear Cell Infiltrations |
|---|---|---|---|---|---|
| 3 Months | |||||
| Control | – | – | – | – | – |
| AE | – | – | – | – | – |
| Cd1 | ± b | – | – | ± d | ± d |
| Cd1 + AE | ± b | – | – | ± b | ± d |
| Cd5 | ± d | ± d | ± b | ± d | ± b |
| Cd5 + AE | ± b | ± b | – | ± b | ± b |
| 10 Months | |||||
| Control | – | – | – | – | – |
| AE | – | – | – | – | – |
| Cd1 | ± d | ± b | ± a | ± b | ± b |
| Cd1 + AE | ± b | – | – | ± a | ± b |
| Cd5 | + d | ± d | ± d | ± d | ± d |
| Cd5 + AE | ± d | ± b | ± b | ± b | ± b |
| 17 Months | |||||
| Control | – | – | – | – | – |
| AE | – | – | – | – | – |
| Cd1 | + d | + d | + d | + d | + d |
| Cd1 + AE | ± d | ± d | ± d | + b | ± d |
| Cd5 | ++ d | + d | ++ d | ++ d | + d |
| Cd5 + AE | + d | ± d | + d | + c | ± d |
| 24 Months | |||||
| Control | – | – | – | – | – |
| AE | – | – | – | – | – |
| Cd1 | + d | ++ d | + d | ++ d | + d |
| Cd1 + AE | ± d | + d | ± d | + b | ± d |
| Cd5 | ++ d | ++ d | ++ d | ++ b | + d |
| Cd5 + AE | + d | + d | + d | + b | ± d |
| Group | Duration of the Experiment | |||
|---|---|---|---|---|
| 3 Months | 10 Months | 17 Months | 24 Months | |
| ALT (U/L) | ||||
| Control | 24.19 ± 1.306 | 22.55 ± 1.308 | 15.95 ± 1.686 | 24.11 ± 3.003 |
| AE | 22.70 ± 1.993 | 15.05 ± 1.004 * | 16.22 ± 1.453 | 31.86 ± 5.217 |
| Cd1 | 21.74 ± 3.480 | 29.29 ± 2.260 * ‡‡‡ | 24.44 ± 2.960 ** ‡‡ | 33.65 ± 3.825 * |
| Cd1 + AE | 21.82 ± 2.020 | 30.25 ± 3.320 * ‡‡‡ | 10.54 ± 0.397 ††† ‡ | 22.61 ± 2.184 † ‡ |
| Cd5 | 23.15 ± 3.442 | 36.77 ± 4.278 *** ‡‡‡ # | 24.64 ± 2.226 ** ‡‡ | 35.92 ± 2.078 * |
| Cd5 + AE | 23.35 ± 3.506 | 19.50 ± 0.846 ††† ## | 12.79 ± 1.314 ††† | 20.44 ± 1.006 †† ‡ |
| AST (U/L) | ||||
| Control | 82.28 ± 5.983 | 62.98 ± 2.645 | 79.48 ± 8.155 | 45.32 ± 4.260 |
| AE | 96.14 ± 4.549 | 64.39 ± 4.516 | 94.32 ± 9.591 | 47.54 ± 2.612 |
| Cd1 | 96.24 ± 7.035 | 80.68 ± 7.370 * | 139.02 ± 10.35 *** ‡‡‡ | 57.33 ± 4.174 * |
| Cd1 + AE | 101.5 ± 6.851 | 77.70 ± 7.296 | 127.5 ± 7.253 *** ‡ | 51.65 ± 5.743 |
| Cd5 | 113.6 ± 5.391 ** | 84.10 ± 7.487 * | 112.37 ± 8.591 * # | 60.02 ± 2.004 * |
| Cd5 + AE | 93.92 ± 7.822 | 83.37 ± 7.533 * | 104.10 ± 7.918 | 51.58 ± 2.396 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mężyńska, M.; Brzóska, M.M.; Rogalska, J.; Piłat-Marcinkiewicz, B. Extract from Aronia melanocarpa L. Berries Prevents Cadmium-Induced Oxidative Stress in the Liver: A Study in A Rat Model of Low-Level and Moderate Lifetime Human Exposure to this Toxic Metal. Nutrients 2019, 11, 21. https://doi.org/10.3390/nu11010021
Mężyńska M, Brzóska MM, Rogalska J, Piłat-Marcinkiewicz B. Extract from Aronia melanocarpa L. Berries Prevents Cadmium-Induced Oxidative Stress in the Liver: A Study in A Rat Model of Low-Level and Moderate Lifetime Human Exposure to this Toxic Metal. Nutrients. 2019; 11(1):21. https://doi.org/10.3390/nu11010021
Chicago/Turabian StyleMężyńska, Magdalena, Małgorzata M. Brzóska, Joanna Rogalska, and Barbara Piłat-Marcinkiewicz. 2019. "Extract from Aronia melanocarpa L. Berries Prevents Cadmium-Induced Oxidative Stress in the Liver: A Study in A Rat Model of Low-Level and Moderate Lifetime Human Exposure to this Toxic Metal" Nutrients 11, no. 1: 21. https://doi.org/10.3390/nu11010021
APA StyleMężyńska, M., Brzóska, M. M., Rogalska, J., & Piłat-Marcinkiewicz, B. (2019). Extract from Aronia melanocarpa L. Berries Prevents Cadmium-Induced Oxidative Stress in the Liver: A Study in A Rat Model of Low-Level and Moderate Lifetime Human Exposure to this Toxic Metal. Nutrients, 11(1), 21. https://doi.org/10.3390/nu11010021