

Sustainable Fisheries and Non-Target Species Management: A Seasonal and Depth-Based Study in the Deep-Sea Fisheries of Antalya Bay


Abstract
1. Introduction
2. Materials and Methods
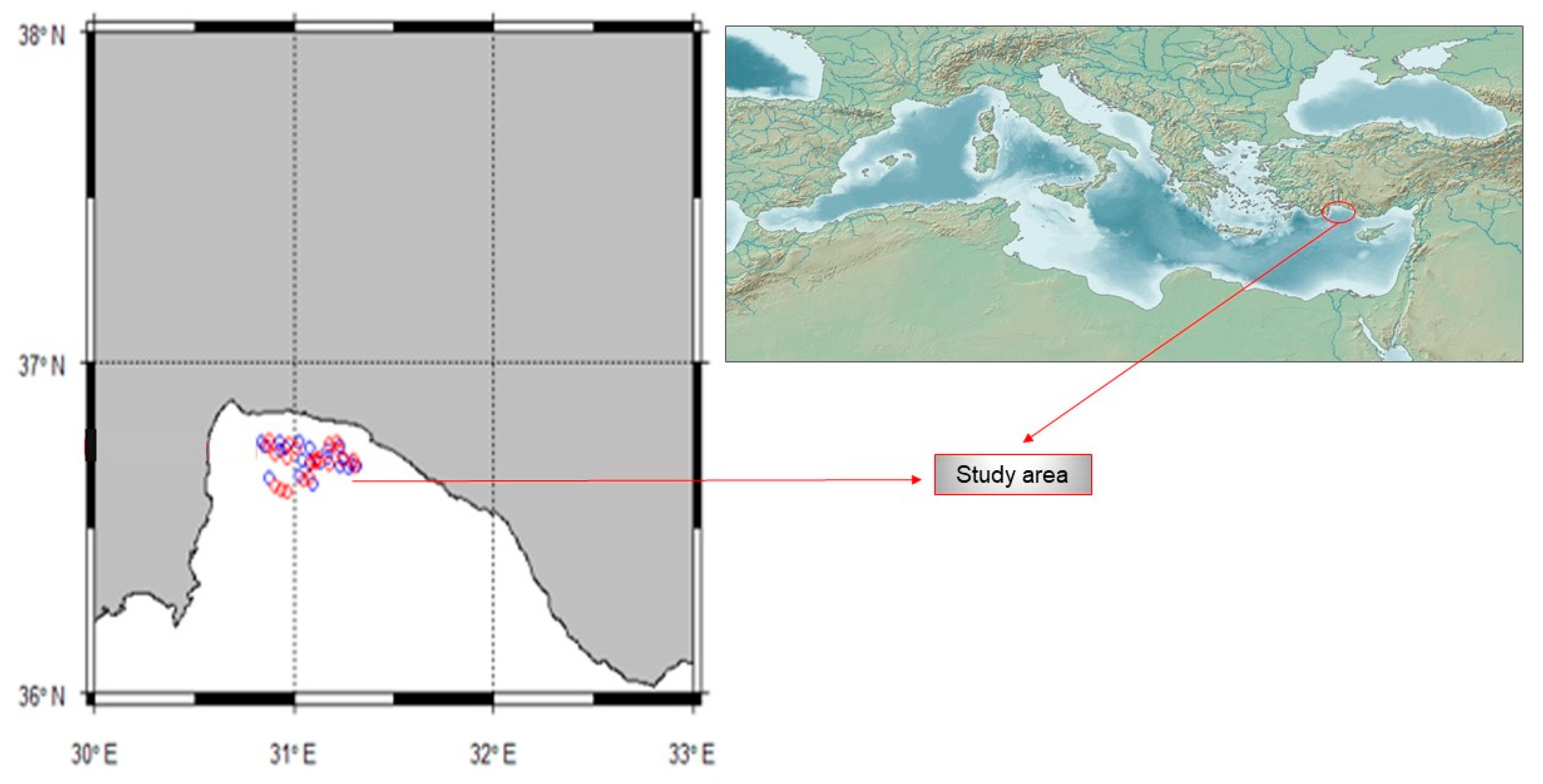
2.1. Study Area
2.2. Sampling and Laboratory Analysis
2.3. Data Analysis
Species Diversity and Community Structure
- Shannon–Wiener diversity index (H′):
- Pielou’s evenness index (J′):
- Simpson dominance index (1 − λ):
- Margalef’s species richness index (D′):
- Catch Composition and Effort Standardization
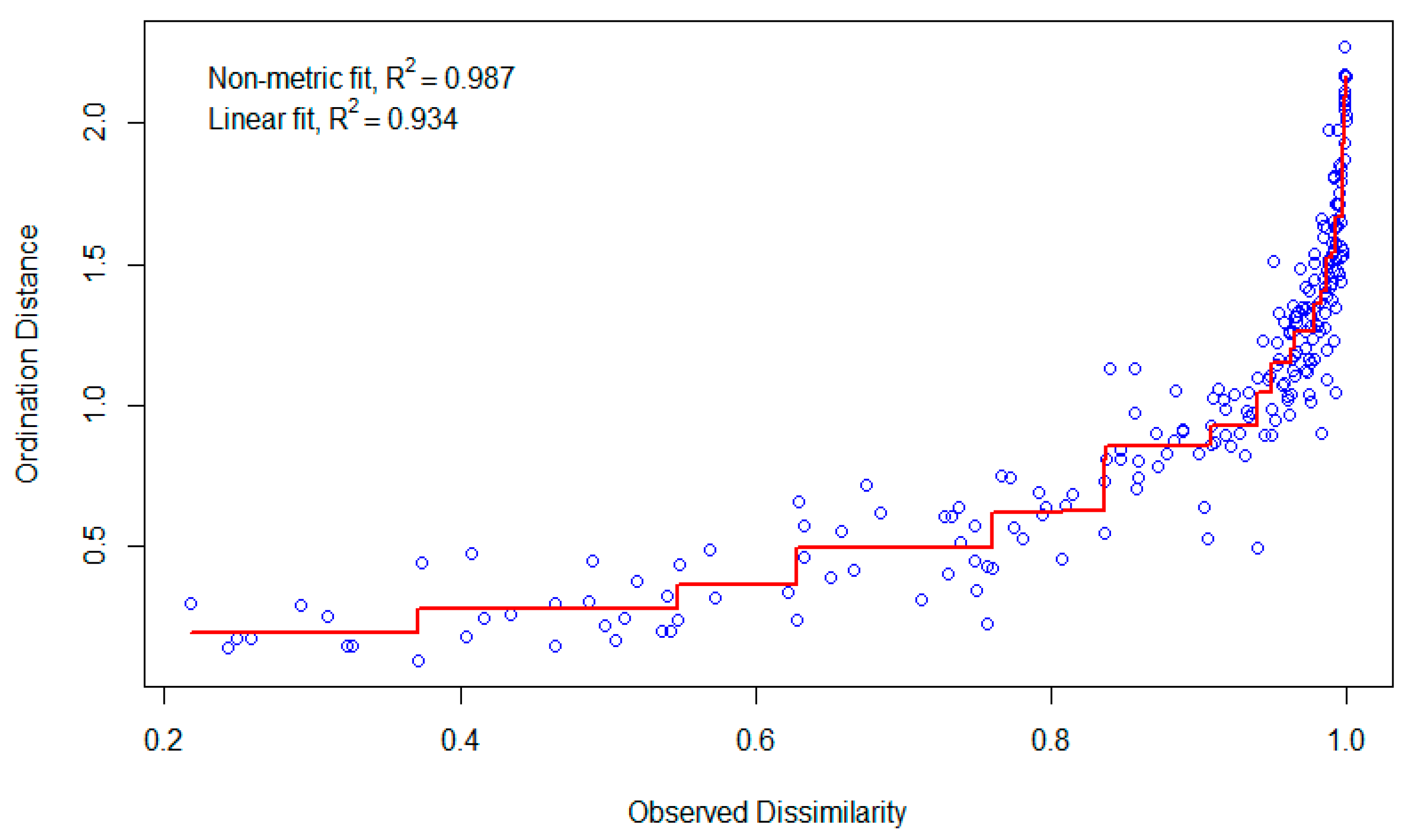
- Multivariate Analysis of Community Structure
- Statistical Hypothesis Testing
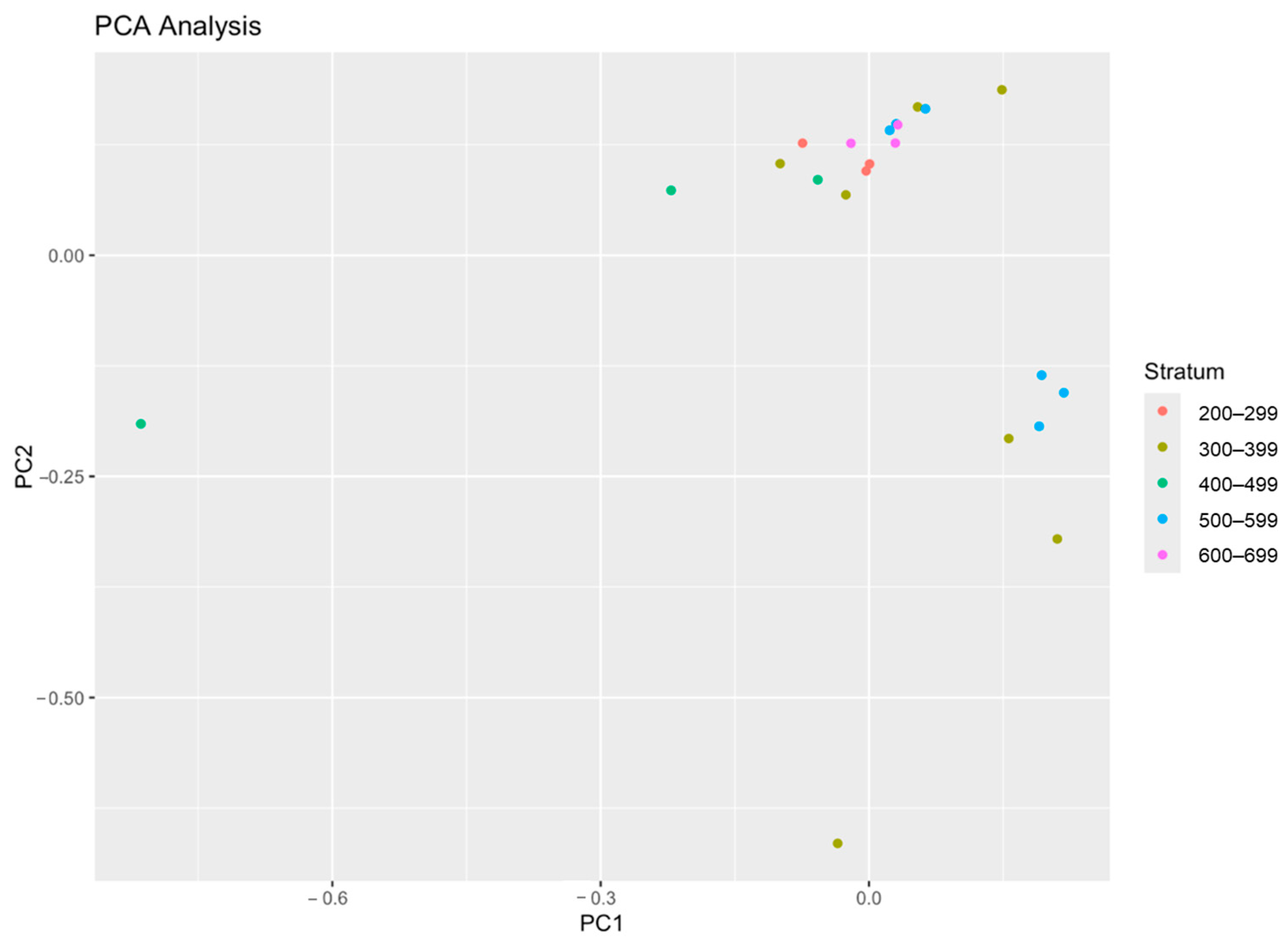
- Principal Component Analysis (PCA)
- Software and Statistical Tools
3. Results
3.1. Catch Composition
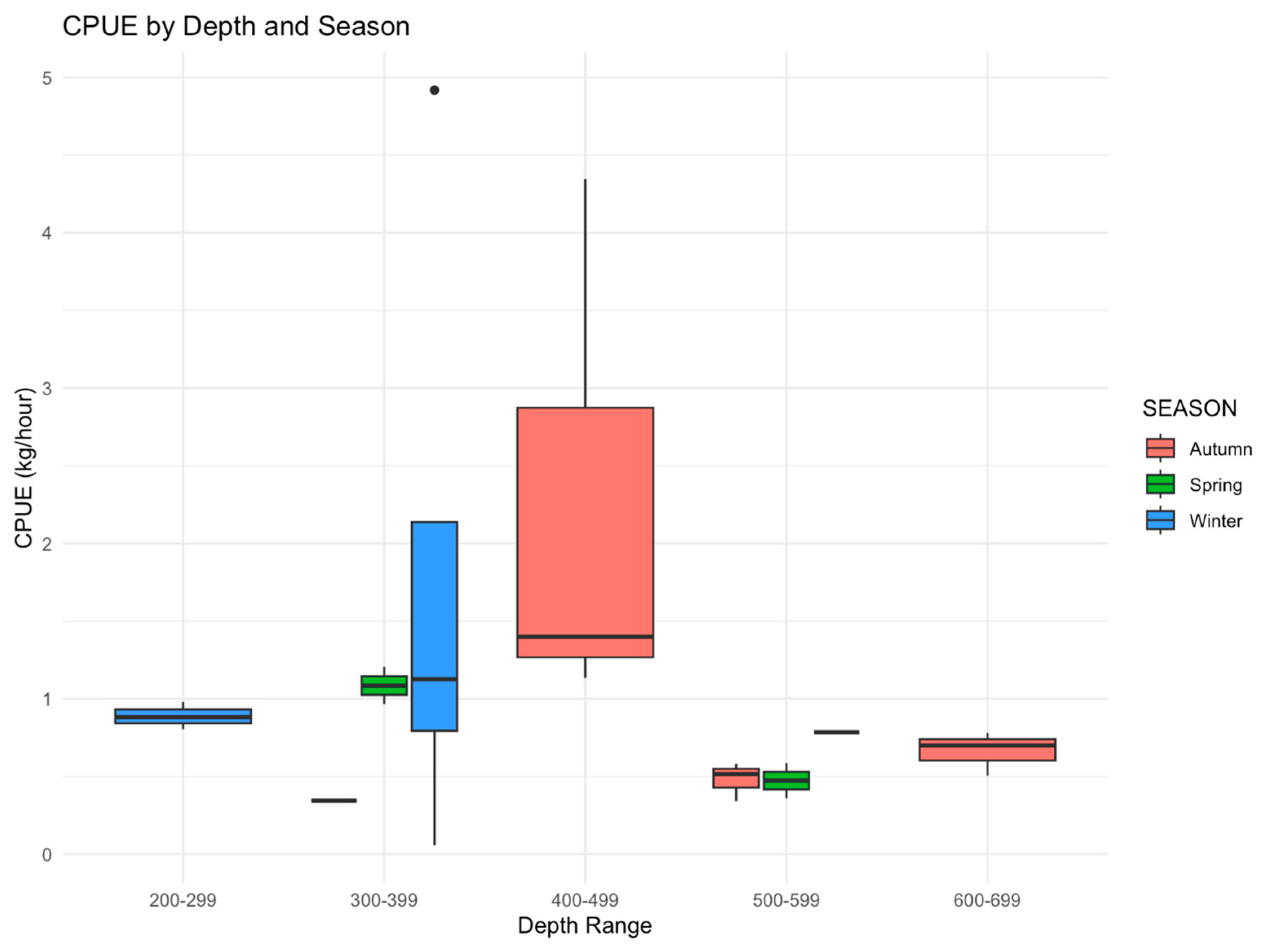
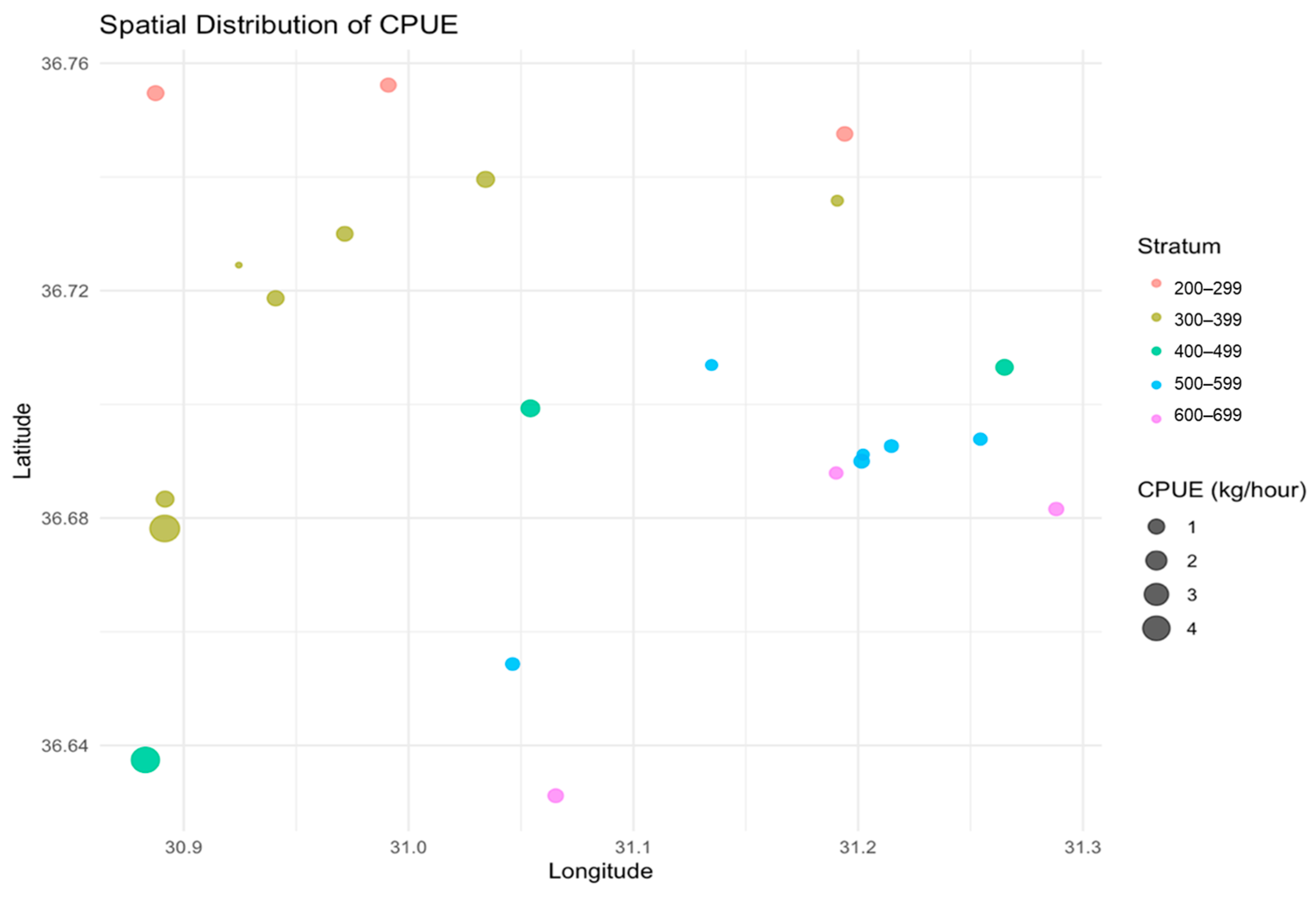
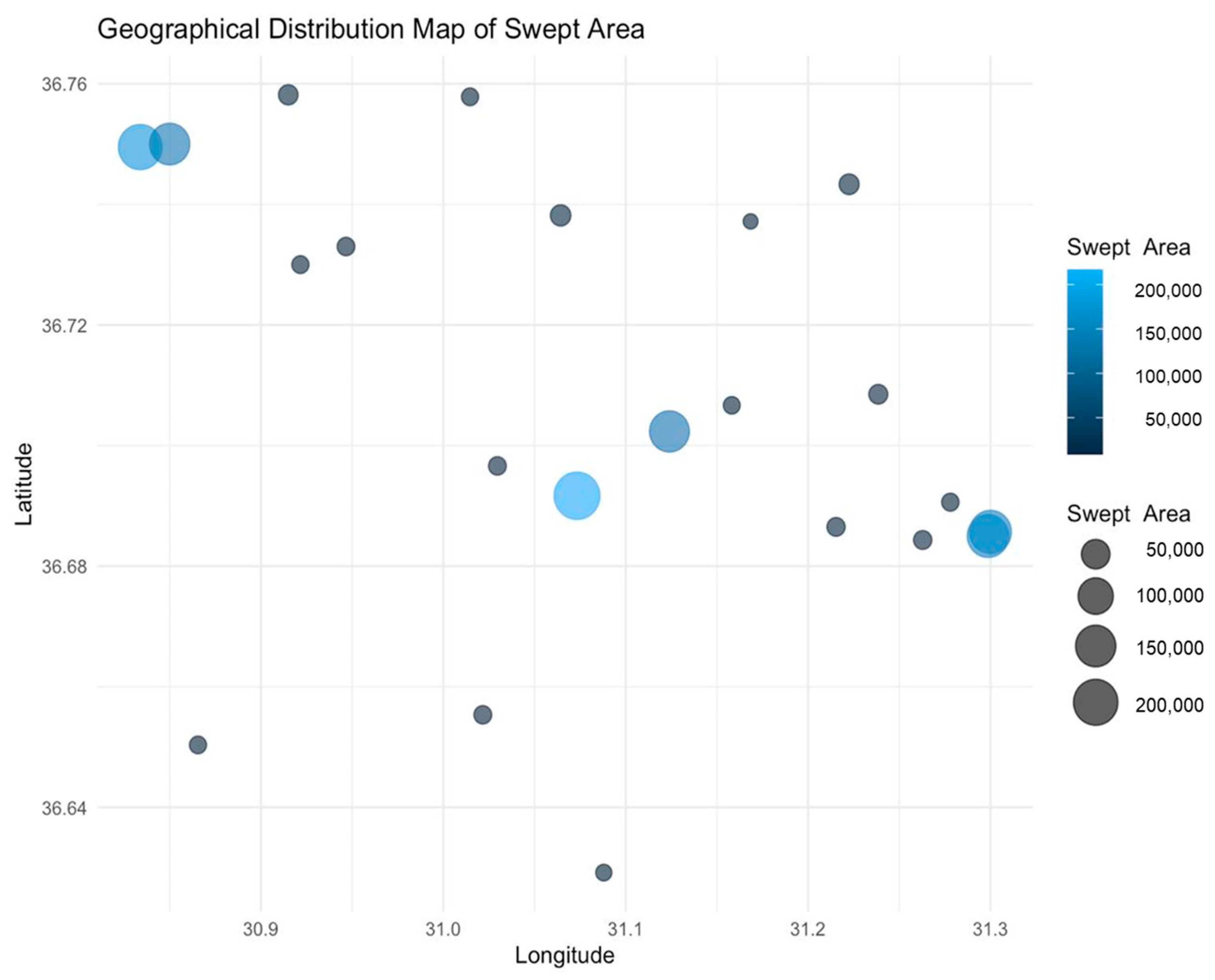
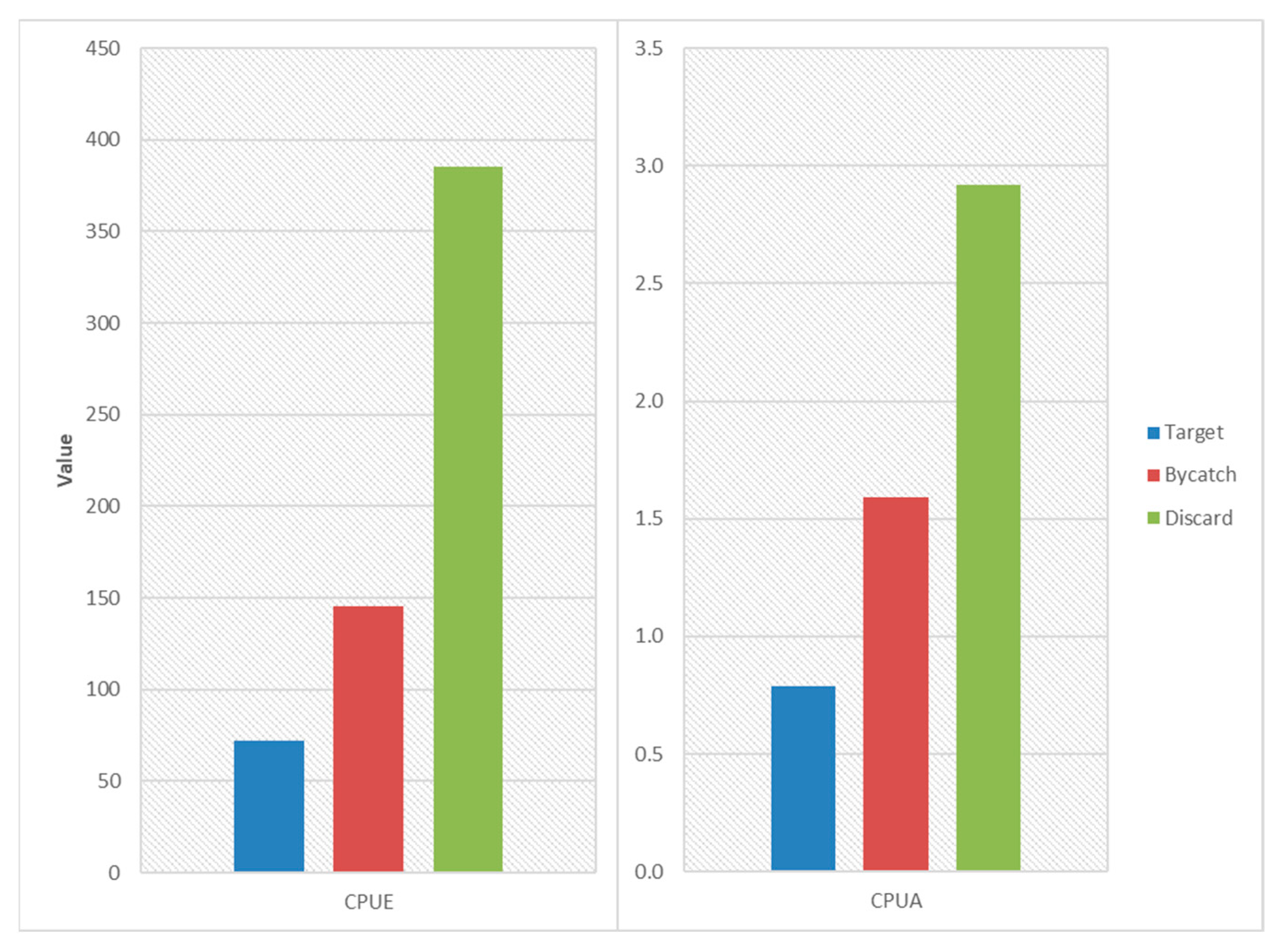
3.2. CPUE and CPUA Analyses
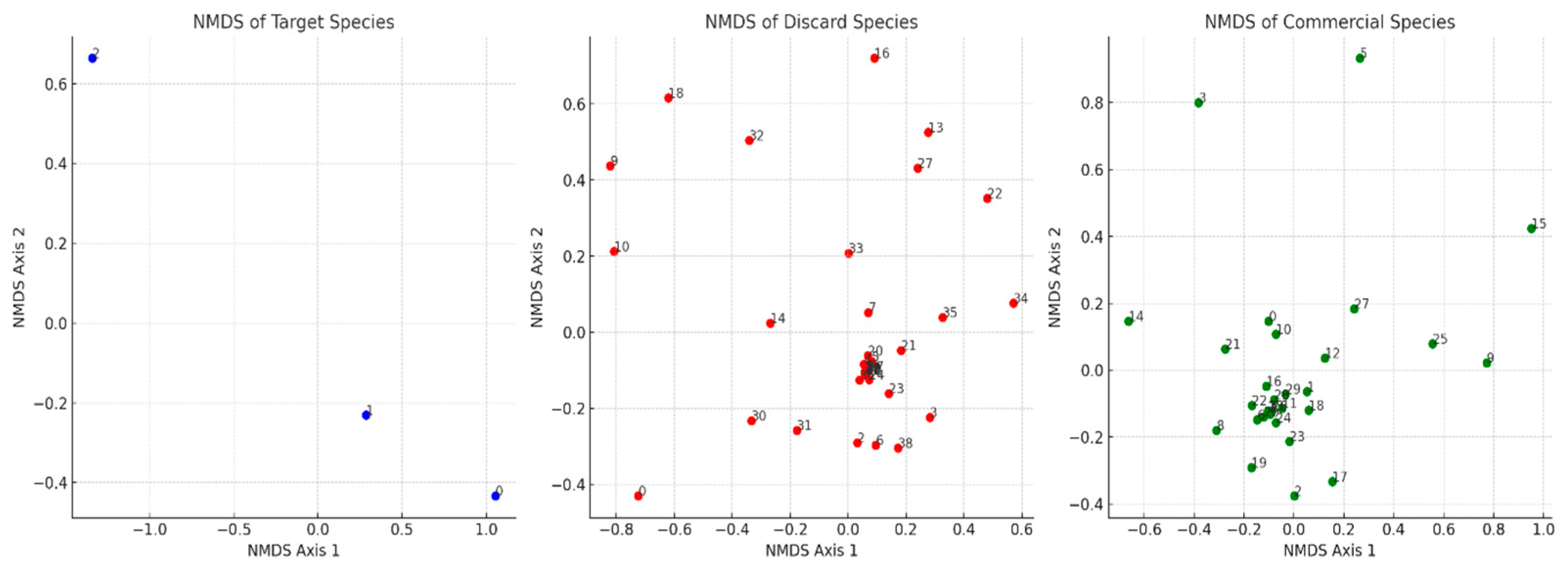
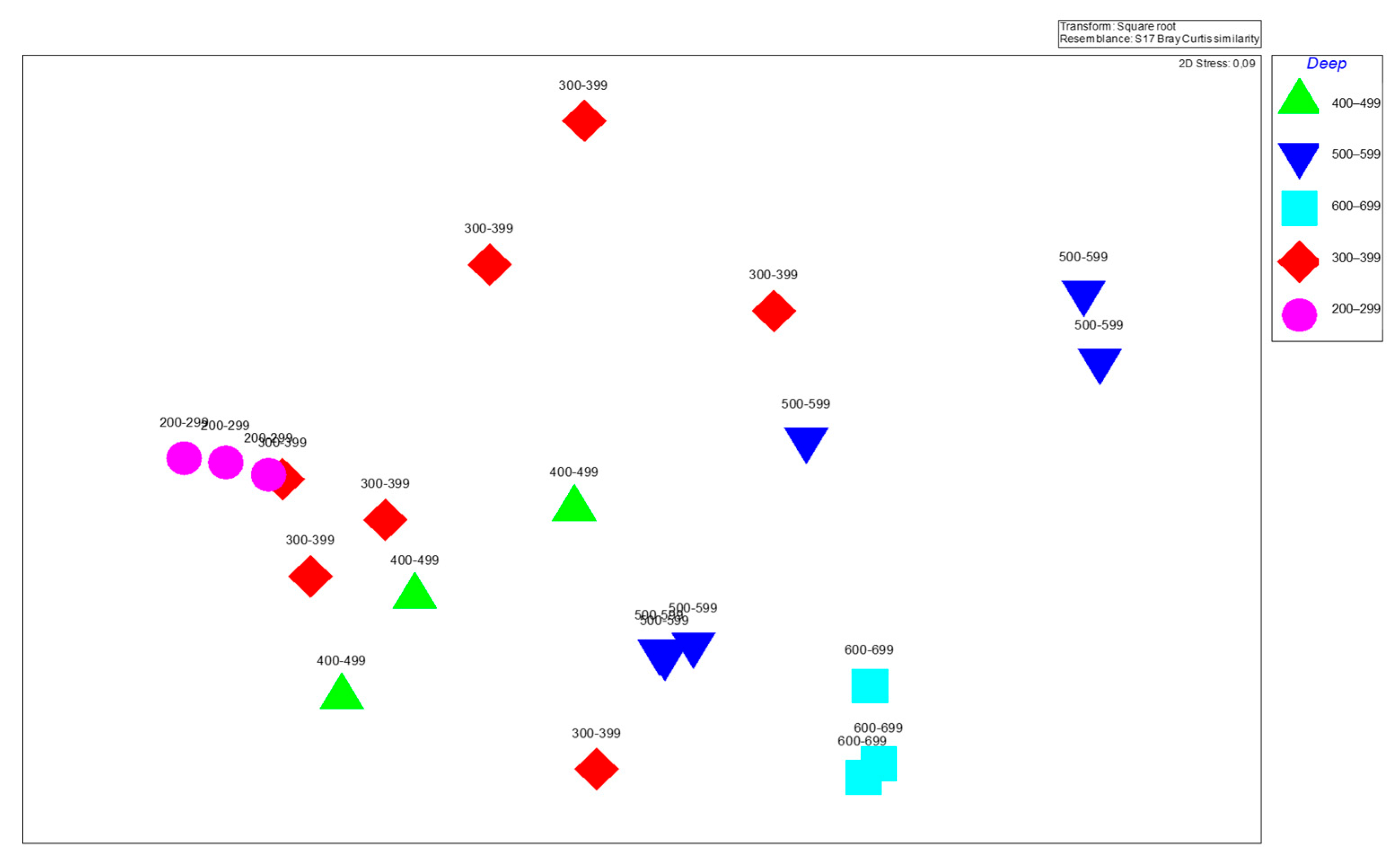
3.3. Biodiversity and Species Composition in Trawl Operations
Diversity Indices and Depth-Based Variations
4. Discussion
4.1. Discard and Bycatch Patterns in Deep-Sea Trawling
4.2. Ecological Impacts and Stock Status Concerns
4.3. Management Challenges and Policy Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EU | European Union |
| CFP | Common Fisheries Policy |
| IUCN | International Union for the Conservation of Nature |
| RFMOs | Regional Fisheries Management Organizations |
| GFCM | General Fisheries Commission for the Mediterranean |
| CPUE | Catch Per Unit Effort |
| CPUA | Catch Per Unit Area |
| JICA | Japan International Cooperation Agency |
| MCRS | Minimum Conservation Reference Size |
References
- Koslow, J.; Boehlert, G.W.; Gordon, J.D.; Haedrich, R.L.; Lorance, P.; Parin, N. Continental slope and deep-sea fisheries: Implications for a fragile ecosystem. ICES J. Mar. Sci. 2000, 57, 548–557. [Google Scholar] [CrossRef]
- FAO. Deep-Ocean Climate Change Impacts on Habitat, Fish and Fisheries; Levin, L., Baker, M., Thompson, A., Eds.; FAO: Rome, Italy, 2019; FAO Fisheries and Aquaculture Technical Paper No. 638; 186p. [Google Scholar]
- Morato, T.; Cheung, W.; Pitcher, T. Vulnerability of seamount fish to fishing: Fuzzy analysis of life-history attributes. J. Fish Biol. 2006, 68, 209–221. [Google Scholar] [CrossRef]
- Colloca, F.; Cardinale, M.; Belluscio, A.; Ardizzone, G. Pattern of distribution and diversity of demersal assemblages in the central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2003, 56, 469–480. [Google Scholar] [CrossRef]
- Rochet, M.J.; Trenkel, V.M. Factors for the variability of discards: Assumptions and field evidence. Can. J. Fish. Aquat. Sci. 2005, 62, 224–235. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. Discards of elasmobranchs in a trammel net fishery targeting cuttlefish, Sepia officinalis Linnaeus, 1758, along the coast of Sicily (central Mediterranean Sea). Reg. Stud. Mar. Sci. 2018, 20, 60–63. [Google Scholar] [CrossRef]
- Tsagarakis, K.; Palialexis, A.; Vassilopoulou, V. Mediterranean fishery discards: Review of the existing knowledge. ICES J. Mar. Sci. 2014, 71, 1219–1234. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Catchpole, T.L.; Frid, C.L.; Gray, T.S. Discards in North Sea fisheries: Causes, consequences and solutions. Mar. Policy 2005, 29, 421–430. [Google Scholar] [CrossRef]
- Johnsen, J.P.; Eliasen, S. Solving complex fisheries management problems What the EU can learn from the Nordic ex-periences of reduction of discards. Mar. Policy 2011, 35, 130–139. [Google Scholar] [CrossRef]
- Kelleher, K. Discards in the World’s Marine Fisheries. An Update; FAO: Rome, Italy, 2005; FAO Fisheries Technical Paper. No. 470; 131p, ISBN 92-5-105289-1. [Google Scholar]
- EU. Green Paper–Reform of the Common Fisheries Policy; CPMR: Brussels, Belgium, 2009.
- Bellido, J.M.; Carbonell, A.; Garcia, M.; Garcia, T.; González, M. The Obligation to Land All Catches–Consequences for the Mediterranean. In-Depth Analysis; European Parliament: Brussels, Belgium, 2014; Policy department B: Structural and cohesion policies.
- Gilman, E.; Passfield, K.; Nakamura, K. Performance of regional fisheries management organizations: Ecosystem-based governance of bycatch and discards. Fish Fish. 2014, 15, 327–351. [Google Scholar] [CrossRef]
- Maravelias, C.D.; Tserpes, G.; Pantazi, M.; Peristeraki, P. Habitat selection and temporal abundance fluctuations of demer-sal cartilaginous species in the Aegean Sea (eastern Mediterranean). PLoS ONE 2012, 7, e35474. [Google Scholar] [CrossRef] [PubMed]
- Tserpes, G.; Maravelias, C.D.; Pantazi, M.; Peristeraki, P. Distribution of relatively rare demersal elasmobranchs in the eastern Mediterranean. Estuar. Coast. Shelf Sci. 2013, 117, 48–53. [Google Scholar] [CrossRef]
- Deval, M.C.; Özgen, G.; Özbilgin, H. Selectivity of 50 mm T0 and T90 codends for commercial shrimp species in the Turkish deepwater trawl fishery, Eastern Mediterranean. J. Appl. Ichthyol. 2016, 32, 1041–1057. [Google Scholar] [CrossRef]
- Deval, M.C.; Kapiris, K. A review of biological patterns of the blue-red shrimp Aristeus antennatus in the Mediterra-nean Sea: A case study of the population of Antalya Bay, eastern Mediterranean Sea. Sci. Mar. 2016, 80, 339–348. [Google Scholar] [CrossRef]
- Deval, M.C.; Yılmaz, S.; Kapiris, K. Spatio temporal variations in decapod crustacean assemblages of bathyal ground in the Antalya Bay (Eastern Mediterranean). Turk. J. Fish. Aquat. Sci. 2017, 17, 967–979. [Google Scholar] [CrossRef]
- Deval, M.C. Population dynamics and biological patterns of commercial crustacean species in the Antalya Bay, Eastern Mediterranean Sea: III. The giant red shrimp Aristaeomorpha foliacea Risso, 1827. Turk. J. Fish. Aquat. Sci-Ences 2019, 20, 311–323. [Google Scholar] [CrossRef]
- Deval, M.C.; Mutlu, E. Spatio-temporal density of the demersal Chondrichthyes assemblage in an upper bathyal of the eastern Mediterranean Sea. Mar. Biodivers. 2024, 54, 31. [Google Scholar] [CrossRef]
- Bayhan, Y.K.; Ergüden, D.; Cartes, J.E. Deep Sea Fisheries in Mersin Bay, Turkey, Eastern Mediterranean: Diversity and Abundance of Shrimps and Benthic Fish Fauna. Acta Zool. Bulg. 2018, 70, 259–268. [Google Scholar]
- Yemisken, E.; Dalyan, C.; Eryilmaz, L. Catch and discard fish species of trawl fisheries in the Iskenderun Bay (North-eastern Mediterranean) with emphasis on lessepsian and chondricthyan species. Mediterr. Mar. Sci. 2014, 15, 380–389. [Google Scholar] [CrossRef]
- Holden, M.J.; Raitt, D.F.S. Manual of Fisheries Science Part 2-Methods of Resource Investigation and Their Application; FAO Fisheries Technical Paper No. 115; FAO: Rome, Italy, 1974; 255p. [Google Scholar]
- Avşar, D. Fisheries Biology and Population Dynamics; Adana Nobel Kitabevi: Adana, Türkiye, 2005; p. 332. ISBN 978-975-856-144-5. [Google Scholar]
- Whitehead, P.J.P.; Bauchot, M.L.; Hureau, J.C.; Nielsen, J.; Tortonese, E. Fishes of the North-Eastern Atlantic and the Medi-Terranean; United Nations Educational, Scientific and Cultural Organization: Paris, France, 1986; Volume 2. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: https://fishbase.org/search.php (accessed on 13 February 2025).
- Fischer, W.; Schneider, M.; Bauchot, M.-L. Fiches FAO D’identification des Especes Pour les Besoins de la Pêche. Mé-Diterranée et Mer Noire (Zone De Pêche 37), Révision 1, Volume 2; FAO: Rome, Italy, 1987. [Google Scholar]
- Shannon, C.; Weiner, W. A Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1948. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Margalef, R. La teoria de la informacion en ecologia. Mem. Real Acad. De Cienc. Artes Barc. 1957, 32, 373–436. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Beisel, J.N.; Moreteau, J.C. A simple formula for calculating the lower limit of Shannon’s diversity index. Ecol. Model. 1997, 99, 289–292. [Google Scholar] [CrossRef]
- Sparre, P. Introduction to tropical fish stock assessment. Part 1: Manual. FAO Fish. Tech. Paper 1993, 306, 192–218. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.; et al. Package ‘vegan’. Community Ecology Package, Version 2013; Volume 2, pp. 1–295. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 23 January 2024).
- RStudio Team. RStudio: Integrated Development for R; PBC: Boston, MA, USA, 2024. [Google Scholar]
- Clarke, K.; Gorley, R. Primer. PRIMER-e, Plymouth; Saint Mary’s University: Halifax, NS, Canada, 2006; p. 866. [Google Scholar]
- Cicek, E.; Avsar, D.; Yeldan, H.; Ozutok, M. Length–weight relationships for 31 teleost fishes caught by bottom trawl net in the Babadillimani Bight (northeastern Mediterranean). J. Appl. Ichthyol. 2006, 22, 290–292. [Google Scholar] [CrossRef]
- Çiçek, E.; Avşar, D.; Yeldan, H.; Meltem Özütok, M. General characteristics of teleost fish fauna trawled from Babadılli-manı Bight (Mersin, Türkiye). Ege J. Fish. Aquat. Sci. 2004, 21, 223–227. [Google Scholar]
- Deval, M.C.; Bök, T.; Ateş, C.; Ulutürk, T.; Tosunoğlu, T. Comparison of the size selectivity of diamond (PA) and square (PE) mesh codends for deepwater crustacean species in the Antalya Bay, eastern Mediterranean. J. Appl. Ichthyol. 2009, 25, 372–380. [Google Scholar] [CrossRef]
- Eryaşar, A.R.; Özbilgin, H.; Gökçe, G.; Özbilgin, Y.D.; Saygu, İ.; Bozaoğlu, A.S.; Kalecik, E. The effect of codend circumfer-ence on selectivity of hand-woven slack knotted codend in the North Eastern Mediterranean demersal trawl fishery. Turk. J. Fish. Aquat. Sci. 2014, 14, 463–470. [Google Scholar] [CrossRef]
- Kebapçıoğlu, T. Interaction Between the Coastal Bottom Trawl Fishery and the Small-Scale Fisheries in the Northeast Mediterranean (Antalya). Ph.D. Thesis, Akdeniz University, Antalya, Turkey, 2014. [Google Scholar]
- Soykan, O. Seasonal Distribution of By-Catch Species in Sığacık Bay by Demersal Trawl. Ph.D. Thesis, Ege University, Bornova, Turkey, 2011. [Google Scholar]
- Yemişken, E. Comparison of Trawl Fisheries Effects Onchondrichthyan Species in Fishing Areas of Eastern Mediterranean Sea. Ph.D. Thesis, İstanbul Üniversitesi, Fatih, Turkey, 2017. [Google Scholar]
- Yeşilçimen, H.Ö.; Kuşat, M. Monthly change of economic fish species caught by bottom trawl fishing from Antalya bay. J. FisheriesSciences.com 2011, 5, 115–121. [Google Scholar] [CrossRef]
- Biagi, F.; Sartor, P.; Ardizzone, G.D.; Belcari, P.; Belluscio, A.; Serena, F. Analysis of demersal fish assemblages of the Tus-cany and Latium coasts (north-western Mediterranean). Sci. Mar. 2002, 66, 233–242. [Google Scholar] [CrossRef]
- Busalacchi, B.; Rinelli, P.; De Domenico, F.; Profeta, A.; Perdichizzi, F.; Bottari, T. Analysis of demersal fish assemblages off the Southern Tyrrhenian Sea (central Mediterranean). Hydrobiologia 2010, 654, 111–124. [Google Scholar] [CrossRef]
- Cartes, J.E.; Maynou, F.; Fanelli, E.; Romano, C.; Mamouridis, V.; Papiol, V. The distribution of megabenthic, invertebrate epifauna in the Balearic Basin (western Mediterranean) between 400 and 2300 m: Environmental gradients influencing assem-blages composition and biomass trends. J. Sea Res. 2009, 61, 244–257. [Google Scholar] [CrossRef]
- Colloca, F.; Crespi, V.; Cerasi, S.; Coppola, S.R. Evolution of the Artisanal Fishery in Cilento, Italy: Case Study; FAO: Rome, Italy, 2003. [Google Scholar]
- Demestre, M.; Sanchez, P.; Abello, P. Demersal fish assemblages and habitat characteristics on the continental shelf and upper slope of the north-western Mediterranean. J. Mar. Biol. Assoc. U. K. 2000, 80, 981–988. [Google Scholar] [CrossRef]
- Gaertner, D.; Menard, F.; Develter, C.; Javier, A. Bycatch of billfishes by the European tuna purse-seine fishery in the Atlantic Ocean. Fish. Bull. 2002, 100, 683–690. [Google Scholar]
- Soykan, O.; Bakır, K.; Kınacıgil, H.T. Demersal trawl discards with spatial and bathymetric emphasis in the Turkish coast of the Aegean Sea. Mar. Biol. Res. 2019, 15, 113–123. [Google Scholar] [CrossRef]
- Japonya Uluslararası İşbirliği Ajansı (Jıca). Marmara, Ege ve Akdeniz’de Demersal Balıkçılık Kaynakları Sörvey Raporu (Survey Report on Demersal Fisheries Resources in Sea of Marmara, Aegean Sea and Mediterranean Sea). Tarım ve Köyişleri Bakanlığı, Tarımsal Üretim ve Geliştirme Genel Müdürlüğü, Japonya Uluslararası İşbirliği Ajansı: Ankara, Türkiye, 1993; 579p. Available online: https://kutuphane.tarimorman.gov.tr/vufind/Record/11040 (accessed on 26 May 2024).
- Damalas, D.; Ligas, A.; Tsagarakis, K.; Vassilopoulou, V.; Stergiou, K.I.; Kallianotis, A.; Sbrana, M.; Maynou, F. The” dis-card problem” in Mediterranean fisheries, in the face of the European Union landing obligation: The case of bottom trawl fishery and implications for management. Mediterr. Mar. Sci. 2018, 19, 459–476. [Google Scholar] [CrossRef]
- Sánchez, F.; Olaso, I. Effects of fisheries on the Cantabrian Sea shelf ecosystem. Ecol. Model. 2004, 172, 151–174. [Google Scholar] [CrossRef]
- Sartor, P.; Sbrana, M.; Reale, B.; Belcari, P. Impact of the deep sea trawl fishery on demersal communities of the northern Tyrrhenian Sea (Western Mediterranean). J. Northwest Atl. Fish. Sci. 2003, 31, 275–284. [Google Scholar] [CrossRef]
- Monteiro, P.; Araújo, A.; Erzini, K.; Castro, M. Discards of the Algarve (southern Portugal) crustacean trawl fishery. Hydrobiologia 2001, 449, 267–277. [Google Scholar] [CrossRef]
- Castriota, L.; Falautano, M.; Romeo, T.; Florio, J.; Pelusi, P.; Finoia, M.G.; Andaloro, F. Crustacean fishery with bottom traps in an area of the southern Tyrrhenian Sea: Species composition, abundance and biomass. Mediterr. Mar. Sci. 2004, 5, 15–22. [Google Scholar] [CrossRef]
- Danovaro, R.; Company, J.B.; Corinaldesi, C.; D’Onghia, G.; Galil, B.; Gambi, C.; Gooday, A.J.; Lampadarious, N.; Luna, G.M.; Morigi, C.; et al. Deep-sea biodiversity in the Mediterranean Sea: The known, the unknown, and the unknowable. PLoS ONE 2010, 5, e11832. [Google Scholar] [CrossRef] [PubMed]
- Clavel-Henry, M.; Bahamon, N.; Solé, J.; Gorelli, G.; Garcia del Arco, J.A.; Carretón, M.; Rotllant, G.; Company, J.B. Mod-eling the spatiotemporal distribution of the deep-sea shrimp Aristeus antennatus (Crustacea: Decapoda) on the northwestern Mediterranean continental margin crossed by submarine canyons. J. Mar. Syst. 2020, 209, 103372. [Google Scholar] [CrossRef]
- Pipitone, C.; Agnetta, D.; Zenone, A.; Giacalone, V.M.; Badalamenti, F.; Fiorentino, F.; D’Anna, G. When the trawl ban is a good option: Opportunities to restore fish biomass and size structure in a Mediterranean fisheries restricted area. Sustainbility 2023, 15, 2425. [Google Scholar] [CrossRef]
- Demirci, A. Non-Target Demersal Species Inhabiting İskenderun Bay and Them Biomass Estimation. Master’s Thesis, Mustafa Kemal University, Antakya, Turkey, 2003. [Google Scholar]
- Tursi, A.; Corbelli, V.; Cipriano, G.; Capasso, G.; Velardo, R.; Chimienti, G. Mega-litter and remediation: The case of Mar Piccolo of Taranto (Ionian Sea). Rend. Lincei. Sci. Fis. Nat. 2018, 29, 817–824. [Google Scholar] [CrossRef]
- Moranta, J.; Massutí, E.; Morales-Nin, B. Fish catch composition of the deep-sea decapod crustacean fisheries in the Balearic Islands (western Mediterranean). Fish. Res. 2000, 45, 253–264. [Google Scholar] [CrossRef]
- Lucchetti, A.; Melli, V.; Brcic’, J. Editorial: Innovations in fishing technology aimed at achieving sustainable fishing. Front. Mar. Sci. 2023, 10, 6. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Katsanevakis, S.; Micheli, F.; Sala, E.; Rilov, G.; Sarà, G.; Malak, D.A.; Abdulla, A.; Grerovasileiou, V.; Gissi, E.; et al. The status of coastal benthic ecosystems in the Mediterranean Sea: Evidence from ecological indicators. Front. Mar. Sci. 2020, 7, 475. [Google Scholar] [CrossRef]
- Quignard, J.P.; Tomasini, J. Mediterranean fish biodiversity. Biol. Mar. Mediterr. 2000, 7, 1–66. [Google Scholar]
- Bilecenoğlu, M.; Kaya, M.; Cihangir, B.; Çiçek, E. An updated checklist of the marine fishes of Turkey. Turk. J. Zool. 2014, 38, 901–929. [Google Scholar] [CrossRef]
- Carbonell, A.; Alemany, F.; Merella, P.; Quetglas, A.; Román, E. The by-catch of sharks in the western Mediterranean (Bal-earic Islands) trawl fishery. Fish. Res. 2003, 61, 7–18. [Google Scholar] [CrossRef]
- Gorelli, G.; Blanco, M.; Sardà, F.; Carretón, M.; Company, J.B. Spatio-temporal variability of discards in the fishery of the deep-sea red shrimp Aristeus antennatus in the northwestern Mediterranean Sea: Implications for management. Sci. Mar. 2016, 80, 79–88. [Google Scholar] [CrossRef]
- Martínez-Abraín, A.; Maestre, R.; Oro, D. Demersal trawling waste as a food source for Western Mediterranean sea-birds during the summer. ICES J. Mar. Sci. 2002, 59, 529–537. [Google Scholar] [CrossRef]
- Sánchez, P.; Sartor, P.; Recasens, L.; Ligas, A.; Martin, J.; De Ranieri, S.; Demestre, M. Trawl catch composition during dif-ferent fishing intensity periods in two Mediterranean demersal fishing grounds. Sci. Mar. 2007, 71, 765–773. [Google Scholar] [CrossRef]
- Blanco, M.; Nos, D.; Lombarte, A.; Recasens, L.; Company, J.B.; Galimany, E. Characterization of discards along a wide bathymetric range from a trawl fishery in the NW Mediterranean. Fish. Res. 2023, 258, 106552. [Google Scholar] [CrossRef]
- Gönülal, O.; Özcan, T.; Katagan, T. A contrubition on the distribution of the giant red shrimp Aristaeomorpha foliacea (Risso, 1827) alond the Aegen Sea and Mediterranean part of Turkey. Rapp. Comm. Int. Mer. Médit. 2010, 39, 534. [Google Scholar]
- Republic of Türkiye Ministry of Agriculture and Forestry. Available online: https://www.tarimorman.gov.tr/Konular/Su-Urunleri/Su-Urunleri-Avciligi (accessed on 25 December 2024).
- Edelist, D.; Sonin, O.; Golani, D.; Rilov, G.; Spanier, E. Spatiotemporal patterns of catch and discards of the Israeli Mediter-ranean trawl fishery in the early 1990 s: Ecological and conservation perspectives. Sci. Mar. 2011, 75, 641–652. [Google Scholar] [CrossRef]
- Garcia, S.M.; Zerbi, A.; Aliaume, C.; Do Chi, T.; Lasserre, G. The Ecosystem Approach to Fisheries. Issues, Terminology, Prin-Ciples, Institutional Foundations, Implementation and Outlook; FAO: Rome, Italy, 2003; FAO Fisheries Technical Paper. No. 443; p. 71. ISBN 92-5-104960-2. [Google Scholar]
- Gücü, A.C. Impact of depth and season on the demersal trawl discard. Turk. J. Fish. Aquat. Sci. 2012, 12, 817–830. [Google Scholar] [CrossRef]
- Massutí, E.; Reñones, O. Demersal resource assemblages in the trawl fishing grounds off the Balearic Islands (western Mediterranean). Sci. Mar. 2005, 69, 167–181. [Google Scholar] [CrossRef]
- Milisenda, G.; Vitale, S.; Massi, D.; Enea, M.; Gancitano, V.; Giusto, G.B.; Badalucco, C.; Gristina, M.; Garofalo, G.; Fiorentino, F. Discard composition associated with the deep water rose shrimp fisheries (Parapenaeus longirostris, Lucas 1846) in the south-central Mediterranean Sea. Mediterr. Mar. Sci. 2017, 18, 53–63. [Google Scholar] [CrossRef]
- Pillai, S.L.; Kizhakudan, S.J.; Radhakrishnan, E.V.; Thirumilu, P. Crustacean bycatch from trawl fishery along north Tamil Nadu coast. Indian J. Fish. 2014, 61, 7–13. [Google Scholar]
- Jennings, S.; Kaiser, M.J. The effects of fishing on marine ecosystems. Adv. Mar. Biol. 1998, 34, 201–352. [Google Scholar] [CrossRef]
- Bellido, J.M.; Santos, M.B.; Pennino, M.G.; Valeiras, X.; Pierce, G.J. Fishery discards and bycatch: Solutions for an ecosys-tem approach to fisheries management? Hydrobiologia 2011, 670, 317–333. [Google Scholar] [CrossRef]
- Hilborn, R. Future directions in ecosystem based fisheries management: A personal perspective. Fish. Res. 2011, 108, 235–239. [Google Scholar] [CrossRef]
- Geraci, M.L.; Colloca, F.; Di Maio, F.; Falsone, F.; Fiorentino, F.; Sardo, G.; Scannella, D.; Gancitano; Vitale, S. How is artificial lighting affecting the catches in deep water rose shrimp trawl fishery of the Central Mediterranean Sea? Ocean Coast. Manag. 2021, 215, 105970. [Google Scholar] [CrossRef]
- Tsagarakis, T.K.; Libralato, S.; Giannoulaki, M.; Touloumis, K.; Somarakis, S.; Machias, A.; Frangoulis, C.; Papantoniou; Kavadas, S.; Stoumboudi, M.T. Drivers of the north Aegean Sea ecosystem (Eastern Mediterranean) through time: Insights from multidecadal retrospective analysis and future simulations. Front. Mar. Sci. 2022, 9, 919793. [Google Scholar] [CrossRef]
- Mytilineou, C.; Herrmann, B.; Mantopoulou-Palouka, D.; Sala, A.; Megalofonou, P. Escape, discard, and landing probabil-ity in multispecies Mediterranean bottom-trawl fishery. ICES J. Mar. Sci. 2023, 80, 542–555. [Google Scholar] [CrossRef]
- Rochet, M.J.; Collie, J.S.; Jennings, S.; Hall, S.J. Does selective fishing conserve community biodiversity? Predictions from a length-based multispecies model. Can. J. Fish. Aquat. Sci. 2011, 68, 469–486. [Google Scholar] [CrossRef]
- Gezelius, S.S. The Problem of Implementing Policies for Sustainable Fishing. In Making Fisheries Management Work: Implementation of Policies for Sustainable Fishing; Gezelius, S.S., Raakjaer, J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–25. ISBN 978-1-4020-8627-4. [Google Scholar]
- Fiorentino, F.; Badalamenti, F.; D’anna, G.; Garofalo, G.; Gianguzza, P.; Gristina, M.; Pipitone, C.; Rizzo, P.; Fortibuoni, T. Changes in spawning-stock structure and recruitment pattern of red mullet, Mullus barbatus, after a trawl ban in the Gulf of Castellammare (central Mediterranean Sea). ICES J. Mar. Sci. 2008, 65, 1175–1183. [Google Scholar] [CrossRef]
- Uhlmann, S.S.; Van Helmond, A.T.; Kemp Stefánsdóttir, E.; Sigurðardóttir, S.; Haralabous, J.; Bellido, J.M.; Catbonell, A.; Catchpole, T.; Damas, D.; Faucannet, L.; et al. Discarded fish in European waters: General patterns and contrasts. ICES J. Mar. Sci. 2014, 71, 1235–1245. [Google Scholar] [CrossRef]
- Voliani, A.; Abella, A.; Auteri, R. Some considerations on the growth performance of Mullus barbatus. Cah. Options Mediterr. 1998, 35, 93–106. [Google Scholar]
- Gökçe, G.; Saygu, I.; Eryaşar, A.R. Catch composition of trawl fisheries in Mersin Bay with emphasis on catch biodiver-sity. Turk. J. Zool. 2016, 40, 522–533. [Google Scholar] [CrossRef]
- Kaiser, M.J.; Ramsay, K.; Richardson, C.A.; Spence, F.E.; Brand, A.R. Chronic fishing disturbance has changed shelf sea benthic community structure. J. Anim. Ecol. 2000, 69, 494–503. [Google Scholar] [CrossRef]
- Lauria, V.; Garofalo, G.; Fiorentino, F.; Massi, D.; Milisenda, G.; Piraino, S.; Russo, T.; Gristina, M. Species distribution models of two critically endangered deep-sea octocorals reveal fishing impacts on vulnerable marine ecosystems in central Mediterranean Sea. Sci. Rep. 2017, 7, 8049. [Google Scholar] [CrossRef] [PubMed]
- Pinello, D.; Gee, J.; Accadia, P.; Sabatella, E.C.; Vitale, S.; Polymeros, K.; Fiorentino, F. Efficiency of shallow-and deep-water trawling in the Mediterranean and its implications for discard reduction. Sci. Mar. 2018, 82, 97–106. [Google Scholar] [CrossRef]
- Pitcher, C.R.; Ellis, N.; Althaus, F.; Williams, A.; McLeod, I.M.; Bustamante, R.H.; Kenyon, R.; Fuller, M. Implications of Currentspatial Management Measures for AFMA ERAs for Habitats—FRDC Project No 2014/204; CSIRO Oceans & Atmosphere: Brisbane, Australia, 2015; 50p, ISBN 978-1-4863-0685-5. [Google Scholar]
- Valentine, P.C.; Almeida, F.P. Effects of fishing on gravel habitats: Assessment and recovery of benthic megafauna on Georges Bank. Am. Fish. Soc. Symp. 2005, 41, 325–343. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Family | Species | Author | N% | W% | t/c/d |
|---|---|---|---|---|---|---|
| Elasmobranchii | Centrophoridae | Centrophorus granulosus | Bloch and Schneider, 1801 | 0.01 | 4.90 | d |
| Etmopteridae | Etmopterus spinax | Linnaeus, 1758 | 0.30 | 1.67 | d | |
| Rajidae | Raja clavata | Linnaeus, 1758 | 0.09 | 2.63 | c | |
| Rajidae | Raja oxyrhynchus | Linnaeus, 1758 | 0.01 | 0.69 | c | |
| Squalidae | Squalus blainvillei | Risso, 1827 | 0.05 | 2.45 | d | |
| Dalatiidae | Dalatias licha | Bonnaterre, 1788 | 0.00 | 0.28 | d | |
| Dasyatidae | Dasyatis pastinaca | Linnaeus, 1758 | 0.00 | 0.29 | d | |
| Pentanchidae | Galeus melastomus | Rafinesque, 1810 | 0.36 | 2.36 | d | |
| Hexanchidae | Heptranchias perlo | Bonnaterre, 1788 | 0.00 | 0.02 | d | |
| Oxynotidae | Oxynotus centrina | Linnaeus, 1758 | 0.00 | 0.06 | d | |
| Scyliorhinidae | Scyliorhinus canicula | Linnaeus, 1758 | 0.34 | 2.34 | d | |
| Actinopterygii | Sparidae | Boops boops | Linnaeus, 1758 | 0.05 | 0.24 | c |
| Citharidae | Citharus linguatula | Linnaeus, 1758 | 0.01 | 0.02 | c | |
| Congridae | Conger conger | Linnaeus, 1758 | 0.02 | 0.47 | c | |
| Dussumieriidae | Etrumeus teres | DeKay, 1842 | 0.00 | 0.06 | c | |
| Sebastidae | Helicolenus dactylopterus | Delaroche, 1809 | 1.11 | 0.10 | c | |
| Lophiidae | Lophius budegassa | Spinola, 1807 | 0.01 | 0.71 | c | |
| Merlucciidae | Merluccius merluccius | Linnaeus, 1758 | 0.13 | 2.44 | c | |
| Mullidae | Mullus barbatus | Linnaeus, 1758 | 0.04 | 0.25 | c | |
| Mullidae | Mullus surmuletus | Linnaeus, 1758 | 0.00 | 0.05 | c | |
| Sparidae | Pagellus acarne | Risso, 1827 | 0.02 | 0.07 | c | |
| Sparidae | Pagellus erythrinus | Linnaeus, 1758 | 0.00 | 0.00 | c | |
| Phycidae | Phycis blennoides | Brünnich, 1768 | 0.03 | 0.54 | c | |
| Scombridae | Scomber japonicus | Houttuyn, 1782 | 0.10 | 1.21 | c | |
| Scorpaenidae | Scorpaena elongata | Cadenat, 1943 | 0.02 | 0.37 | c | |
| Serranidae | Serranus hepatus | Linnaeus, 1758 | 0.02 | 0.00 | c | |
| Carangidae | Trachurus mediterraneus | Steindachner, 1868 | 0.02 | 0.14 | c | |
| Carangidae | Trachurus picturatus | Bowdich, 1825 | 0.01 | 0.10 | c | |
| Carangidae | Trachurus trachurus | Linnaeus, 1758 | 0.09 | 0.39 | c | |
| Triglidae | Trigla lucerna | Linnaeus, 1758 | 0.01 | 0.07 | c | |
| Triglidae | Trigla lyra | Linnaeus, 1758 | 0.09 | 0.32 | c | |
| Mullidae | Upeneus moluccensis | Bleeker, 1855 | 0.01 | 0.03 | c | |
| Zeidae | Zeus faber | Linnaeus, 1758 | 0.02 | 0.13 | c | |
| Argentinidae | Argentina sphyraena | Linnaeus, 1758 | 14.94 | 9.25 | d | |
| Sternoptychidae | Argyropelecus hemigymnus | Cocco, 1829 | 0.00 | 0.00 | d | |
| Bothidae | Arnoglossus imperialis | Rafinesque, 1810 | 0.40 | 0.25 | d | |
| Triglidae | Aspitrigla cuculus | Linnaeus, 1758 | 0.66 | 0.96 | d | |
| Caproidae | Capros aper | Linnaeus, 1758 | 1.24 | 0.75 | d | |
| Myctophidae | Ceratoscopelus maderensis | Lowe, 1839 | 0.00 | 0.00 | d | |
| Chlorophthalmidae | Chlorophthalmus agassizii | Bonaparte, 1840 | 53.13 | 37.11 | d | |
| Macrouridae | Coelorhynchus coelorhynchus | Risso, 1810 | 2.75 | 3.40 | d | |
| Trachichthyidae | Hoplostethus mediterraneus | Cuvier, 1829 | 0.48 | 0.75 | d | |
| Myctophidae | Hygophum benoiti | Cocco, 1838 | 0.00 | 0.00 | d | |
| Macrouridae | Hymenocephalus italicus | Giglioli, 1884 | 6.64 | 2.08 | d | |
| Tetraodontidae | Lagocephalus suezensi | Clark and Gohar, 1953 | 0.00 | 0.01 | d | |
| Myctophidae | Lampanyctus crocodilus | Risso, 1810 | 0.00 | 0.00 | d | |
| Trichiuridae | Lepidopus caudatus | Euphrasen, 1788 | 0.04 | 0.26 | d | |
| Scophthalmidae | Lepidorhombus whiffiagonis | Walbaum, 1792 | 0.09 | 1.54 | d | |
| Triglidae | Lepidotrigla cavillone | Lacepède, 1801 | 0.08 | 0.12 | d | |
| Centriscidae | Macrorhamphosus scolopax | Linnaeus, 1758 | 0.06 | 0.04 | d | |
| Myctophidae | Myctophum punctatum | Rafinesque, 1810 | 0.00 | 0.00 | d | |
| Nettastomatidae | Nettastoma melanurum | Rafinesque, 1810 | 0.05 | 0.58 | d | |
| Peristediidae | Peristedion cataphractum | Linnaeus, 1758 | 0.03 | 0.03 | d | |
| Sparidae | Spicara maena | Linnaeus, 1758 | 0.04 | 0.07 | d | |
| Stomiidae | Stomias boa | Risso, 1810 | 0.00 | 0.00 | d | |
| Callionymidae | Synchiropus phaeton | Günther, 1861 | 0.86 | 0.49 | d | |
| Tetraodontidae | Torquigener flavimaculosus | Hardy and Randall, 1983 | 0.00 | 0.21 | d | |
| Trichiurida | Trichiurus lepturus | Linnaeus, 1758 | 0.00 | 0.03 | d | |
| Malacostraca | Aristeidae | Aristaeomorpha foliacea | Risso, 1827 | 1.66 | 5.37 | t |
| Aristeidae | Aristeus antennatus | Risso, 1816 | 0.23 | 0.73 | t | |
| Penaeidae | Parapenaeus longirostris | H. Lucas, 1846 | 8.66 | 4.00 | t | |
| Polybiidae | Bathynectes maravigna | Prestandrea, 1839 | 0.00 | 0.00 | d | |
| Calappidae | Calappa granulosa | Linnaeus, 1758 | 0.01 | 0.04 | d | |
| Majidae | Maia goltziana | d’Oliveira, 1888 | 0.00 | 0.01 | d | |
| Pandalidae | Plesionika edwardsii | JF Brandt in von Middendorf, 1851 | 2.28 | 1.06 | d | |
| Pandalidae | Plesionika heterocarpus | A. Costa, 1871 | 1.11 | 0.17 | d | |
| Pandalidae | Plesionika martia | A. Milne-Edwards, 1883 | 1.13 | 0.42 | d | |
| Polychelidae | Polycheles typhlops | Heller, 1862 | 0.05 | 0.04 | d | |
| Cephalopoda | Ommastrephidae | Illex coindetii | Vérany, 1839 | 0.01 | 0.07 | d |
| Loliginidae | Loligo vulgaris | Lamarck, 1798 | 0.00 | 0.00 | c | |
| Sepiidae | Sepia officinalis | Linnaeus, 1758 | 0.02 | 3.80 | c | |
| Ommastrephidae | Todarodes sagittatus | Lamarck, 1798 | 0.02 | 0.00 | d | |
| Rhynchonellata | Terebratulidae | Gryphus vitreus | Born, 1778 | 0.25 | 0.05 | d |
| Holothuroidea | Stichopodidae | Stichopus regalis | Cuvier, 1817 | 0.06 | 0.89 | d |
| Catch | N | N % | W | W % |
|---|---|---|---|---|
| Commercial | 22,455 | 12.87 | 669.97 | 32.46 |
| Discard | 151,983 | 87.13 | 1393.91 | 67.54 |
| Commercial/total | 0.13 | 0.32 | ||
| Commercial/discard | 0.15 | 0.48 |
| CPUE | CPUA | W | N | |
|---|---|---|---|---|
| Target | 72.26 | 0.79 | 188.67 | 18,407 |
| Bycatch | 145.12 | 1.59 | 481.30 | 4048 |
| Discard | 385.52 | 2.92 | 1393.91 | 151,983 |
| Depth | Season | Target | Bycatch | Discard | Total |
|---|---|---|---|---|---|
| 200–299 | Winter | 9.44 | 11.22 | 33.72 | 54.38 |
| 300–399 | Autumn | 1.09 | 3.07 | 16.99 | 21.15 |
| Winter | 3.45 | 30.47 | 93.92 | 127.84 | |
| Spring | 4.35 | 39.09 | 56.17 | 99.61 | |
| 400–499 | Autumn | 2.15 | 16.33 | 114.35 | 132.83 |
| 500–599 | Autumn | 3.29 | 5.81 | 19.86 | 28.96 |
| Winter | 22.10 | 9.21 | 18.48 | 49.79 | |
| Spring | 15.18 | 16.29 | 9.21 | 40.68 | |
| 600–700 | Autumn | 11.21 | 13.63 | 22.82 | 47.66 |
| DEPTH | S | N | D | J’ | H′ | 1-Lambda’ |
|---|---|---|---|---|---|---|
| 200–299 | 35 | 33269 | 6.82 | 0.517 | 2.21 | 0.152 |
| 300–399 | 57 | 87026 | 15.6 | 0.438 | 2.27 | 0.209 |
| 400–499 | 33 | 40277 | 6.98 | 0.489 | 2.11 | 0.212 |
| 500–599 | 35 | 8045 | 12.1 | 0.660 | 3.10 | 0.0685 |
| 600–699 | 28 | 5834 | 6.69 | 0.668 | 2.72 | 0.0938 |
| Indices’ rating range and values | Range 0–>5 Integrated > 5 Semi-disturbed > 2.05–5 Disturbed ≤ 2.05 | Range 0–1 Balanced 0. 8–1 Semi-balanced 0.5–0.8 Unbalanced ≤ 0.5 | Range 0–5 High status 4–5 Good status 3–4 Moderate status 2–3 Poor status 1–2 Bad status 0–1 | Range 0–1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cömert, N.; Deniz, T.; Göktürk, D. Sustainable Fisheries and Non-Target Species Management: A Seasonal and Depth-Based Study in the Deep-Sea Fisheries of Antalya Bay. Sustainability 2025, 17, 5040. https://doi.org/10.3390/su17115040
Cömert N, Deniz T, Göktürk D. Sustainable Fisheries and Non-Target Species Management: A Seasonal and Depth-Based Study in the Deep-Sea Fisheries of Antalya Bay. Sustainability. 2025; 17(11):5040. https://doi.org/10.3390/su17115040
Chicago/Turabian StyleCömert, Nurdan, Tomris Deniz, and Didem Göktürk. 2025. "Sustainable Fisheries and Non-Target Species Management: A Seasonal and Depth-Based Study in the Deep-Sea Fisheries of Antalya Bay" Sustainability 17, no. 11: 5040. https://doi.org/10.3390/su17115040
APA StyleCömert, N., Deniz, T., & Göktürk, D. (2025). Sustainable Fisheries and Non-Target Species Management: A Seasonal and Depth-Based Study in the Deep-Sea Fisheries of Antalya Bay. Sustainability, 17(11), 5040. https://doi.org/10.3390/su17115040