
Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution


Abstract
1. Introduction
2. Materials and Methods
2.1. WGS Data Used
2.2. Single Gene Datasets
2.3. Phylogeny Reconstruction
2.4. Network Reconstructions
2.5. Genomic Substitution Hotspots in LSDV WGS Datasets
3. Results and Discussion
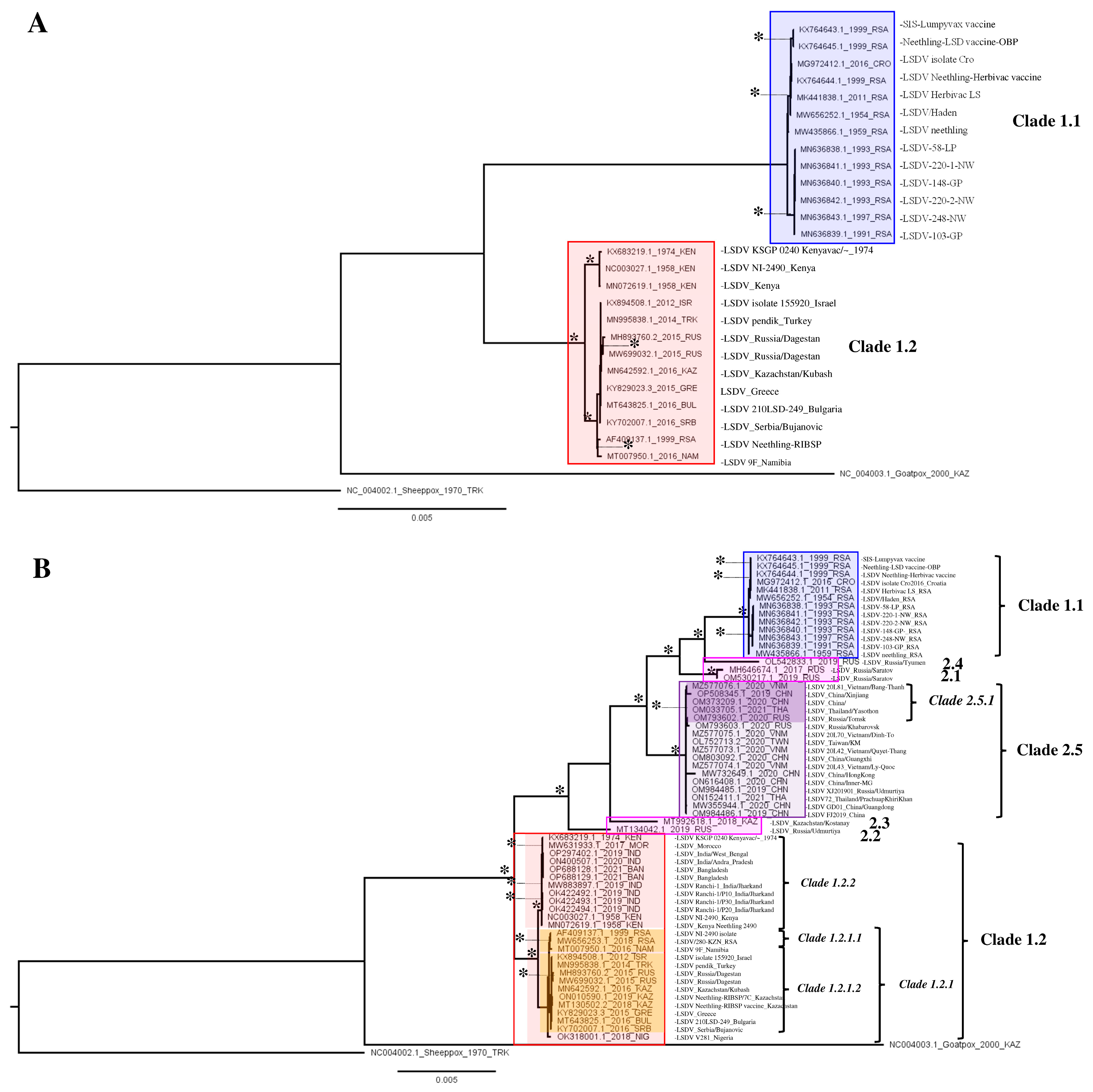
3.1. WGS-Based Phylogeny Reconstruction and Deduced Putative Spatio-Temporal Spread
3.2. Single-Gene Analysis Results in Recombinant Field Strain Phylogenetic Instability
3.3. Genomic Substitution Hotspots in LSDV WGS Datasets
4. Recommendations and Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical Lumpy Skin Disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Coetzer, J.A.W.; Tuppurainen, E.; Babiuk, S.; Wallace, D. Lumpy Skin Disease. In Infectious Diseases of Livestock, Part II; Anipedia: Pretoria, South Africa, 2018. [Google Scholar]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of Lumpy Skin Disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef]
- Kononov, A.; Byadovskaya, O.; Wallace, D.; Prutnikov, P.; Pestova, Y.; Kononova, S.; Nesterov, A.; Rusaleev, V.; Lozovoy, D.; Sprygin, A. Non-vector-borne transmission of Lumpy Skin Disease virus. Sci. Rep. 2020, 10, 7436. [Google Scholar]
- Shumilova, I.; Krotova, A.; Nesterov, A.; Byadovskaya, O.; van Schalkwyk, A.; Sprygin, A. Overwintering of recombinant Lumpy Skin Disease virus in northern latitudes, Russia. Transbound. Emerg. Dis. 2022, 69, e3239–e3243. [Google Scholar] [CrossRef]
- Shumilova, I.; Nesterov, A.; Byadovskaya, O.; Prutnikov, P.; Wallace, D.B.; Mokeeva, M.; Pronin, V.; Kononov, A.; Chvala, I.; Sprygin, A. A recombinant vaccine-like strain of Lumpy Skin Disease virus causes low-level infection of cattle through virus-inoculated feed. Pathogens 2022, 11, 920. [Google Scholar] [CrossRef]
- Hamdi, J.; Munyanduki, H.; Omari Tadlaoui, K.; El Harrak, M.; Fassi Fihri, O. Capripoxvirus infections in ruminants: A review. Microorganisms 2021, 9, 902. [Google Scholar] [CrossRef]
- Kononov, A.; Prutnikov, P.; Shumilova, I.; Kononova, S.; Nesterov, A.; Byadovskaya, O.; Pestova, Y.; Diev, V.; Sprygin, A. Determination of Lumpy Skin Disease virus in bovine meat and offal products following experimental infection. Transbound. Emerg. Dis. 2019, 66, 1332–1340. [Google Scholar] [CrossRef]
- Haegeman, A.; Sohier, C.; Mostin, L.; De Leeuw, I.; Van Campe, W.; Philips, W.; De Regge, N.; De Clercq, K. Evidence of Lumpy Skin Disease virus transmission from subclinically infected cattle by Stomoxys calcitrans. Viruses 2023, 15, 1285. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Oura, C.A. Review: Lumpy Skin Disease: An emerging threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus diseases: Current status and opportunities for control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Venter, E.; Tuppurainen, E.; Saegerman, C.; De Clercq, K. Comparative evaluation of Lumpy Skin Disease virus-based live attenuated vaccines. Vaccines 2021, 9, 473. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Oura, C. Lumpy Skin Disease: An African cattle disease getting closer to the EU. Vet. Rec. 2014, 175, 300–301. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Mathijs, E.; Haegeman, A.; Al-Majali, A.; Van Borm, S.; De Clercq, K. Complete genome sequence of Capripoxvirus strain KSGP 0240 from a commercial live attenuated vaccine. Genome Announc. 2016, 4, e01114-16. [Google Scholar] [CrossRef]
- Bamouh, Z.; Fellahi, S.; Khayi, S.; Hamdi, J.; Omari Tadlaoui, K.; Fassi-Fihri, O.; Elharrak, M. Draft genome sequence of the Capripoxvirus vaccine strain KSGP 0240, reisolated from cattle. Microbiol. Resour. Announc. 2021, 10, e0044021. [Google Scholar] [CrossRef]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of Lumpy Skin Disease virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef]
- Biswas, S.; Noyce, R.S.; Babiuk, L.A.; Lung, O.; Bulach, D.M.; Bowden, T.R.; Boyle, D.B.; Babiuk, S.; Evans, D.H. Extended sequencing of vaccine and wild-type Capripoxvirus isolates provides insights into genes modulating virulence and host range. Transbound. Emerg. Dis. 2020, 67, 80–97. [Google Scholar] [CrossRef]
- van Schalkwyk, A.; Byadovskaya, O.; Shumilova, I.; Wallace, D.B.; Sprygin, A. Estimating evolutionary changes between highly passaged and original parental Lumpy Skin Disease virus strains. Transbound. Emerg. Dis. 2021, 69, e486–e496. [Google Scholar] [CrossRef]
- Mercier, A.; Arsevska, E.; Bournez, L.; Bronner, A.; Calavas, D.; Cauchard, J.; Falala, S.; Caufour, P.; Tisseuil, C.; Lefrançois, T.; et al. Spread rate of Lumpy Skin Disease in the Balkans, 2015–2016. Transbound. Emerg. Dis. 2018, 65, 240–243. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Mathijs, E.; Philips, W.; Saduakassova, M.; De Leeuw, I.; Sultanov, A.; Haegeman, A.; De Clercq, K. Recombinant LSDV strains in Asia: Vaccine spillover or natural emergence? Viruses 2022, 14, 1429. [Google Scholar] [CrossRef]
- Sprygin, A.; Babin, Y.; Pestova, A.; Kononova, S.; Byadovskaya, A.; Kononov, A. Complete genome sequence of the Lumpy Skin Disease virus recovered from the first outbreak in the northern Caucasus of Russia in 2015. Genome Announc. 2019, 8, e01733-18. [Google Scholar] [CrossRef]
- Sprygin, A.; van Schalkwyk, A.; Shumilova, I.; Nestorov, A.; Kononova, S.; Prutnikov, P.; Byadovskaya, O.; Kononov, A. Full-length genome characterization of a novel recombinant vaccine-like Lumpy Skin Disease virus strain detected during the climatic winter in Russia, 2019. Arch. Virol. 2020, 165, 2675–2677. [Google Scholar] [CrossRef]
- Mathijs, E.; Vandenbussche, F.; Nguyen, L.; Aerts, L.; Nguyen, T.; De Leeuw, I.; Quang, M.; Nguyen, H.D.; Philips, W.; Dam, T.V.; et al. Coding-complete sequences of recombinant Lumpy Skin Disease viruses collected in 2020 from four outbreaks in northern Vietnam. Microbiol. Resour. Announc. 2021, 10, e89721. [Google Scholar] [CrossRef]
- Sanjuán, R.; Nebot, M.R.; Chirico, N.; Mansky, L.M.; Belshaw, R. Viral mutation rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef]
- Babkin, I.V.; Shchelkunov, S.N. The time scale in poxvirus evolution. Mol. Biol. 2006, 40, 20–24. [Google Scholar] [CrossRef]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of Myxoma virus on two continents. PLoS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef]
- McLysaght, A.; Pierre, F.; Baldi, P.F.; Gaut, B.S. Extensive gene gain associated with adaptive evolution of poxviruses. Proc. Natl. Acad. Sci. USA 2003, 100, 15655–15660. [Google Scholar] [CrossRef]
- Elde, N.C.; Child, S.J.; Eickbush, M.T.; Kitzman, J.O.; Rogers, K.S.; Shendure, J.; Geballe, A.P.; Malik, H.S. Poxviruses deploy genomic accordions to adapt rapidly against host antiviral defenses. Cell 2012, 150, 831–841. [Google Scholar] [CrossRef]
- Brennan, G.; Stoian, A.M.M.; Yu, H.; Rahman, M.J.; Banerjee, S.; Stroup, J.N.; Park, C.; Tazi, L.; Rothenburg, S. Molecular mechanisms of poxvirus evolution. mBio 2023, 14, e0152622. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum -likelihood phylogenies Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of nucleotide substitution. Bioinform. Appl. Note 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Dress, A.M.W.; Huson, D.H. Constructing splits graphs. IEEE/ACM Trans. Comput. Biol. Bioinform. 2004, 1, 109–115. [Google Scholar] [CrossRef]
- Holland, B.; Moulton, V. Consensus networks: A method for visualizing incompatibilities in collections of trees. In Proceedings of the Algorithms in Bioinformatics: Third International Workshop, WABI 2003, Budapest, Hungary, 15–20 September 2003; Benson, G., Page, R., Eds.; Springer: Chem, Switzerland, 2003; Volume 2812 of LNBI, pp. 165–176. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Dress, A.W. Split decomposition: A new and useful approach to phylogenetic analysis of distance data. Mol. Phylogenet Evol. 1992, 1, 242–252. [Google Scholar] [CrossRef]
- Hamming, R.W. Error detecting and error correcting codes. Bell Syst. Tech. J. 1950, 29, 147–160. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- van Schalkwyk, A.; Kara, P.; Heath, L. Phylogenomic characterization of historic Lumpy Skin Disease virus isolates from South Africa. Arch. Virol. 2022, 167, 2063–2070. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D.M.; Holmes, E.C. Molecular Evolution: A Phylogenetic Approach; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 1998; ISBN 0-86542-889-1. [Google Scholar]
- Wortley, A.H.; Rudall, P.J.; Harris, D.J.; Scotland, R.W. How much data are needed to resolve a difficult phylogeny? Case study in Lamiales. Syst. Biol. 2005, 54, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Schierup, M.H.; Jotun Hein, J. Consequences of recombination on traditional phylogenetic analysis. Genetics 2000, 156, 879–891. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. The effect of recombination on the accuracy of phylogeny estimation. J. Mol. Evol. 2002, 54, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Nir, O.; Braverman, Y.; Davidson, M.; Grinstein, H.; Haymovitch, M.; Zamir, O. Spread of Lumpy Skin Disease in Israeli dairy herds. Vet. Rec. 1995, 137, 91–93. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Van der Waal, K. Spatial and temporal epidemiology of Lumpy Skin Disease in the Middle East, 2012–2015. Front. Vet. Sci. 2016, 3, 19. [Google Scholar] [CrossRef]
- Agianniotaki, E.I.; Mathijs, E.; Vandenbussche, F.; Tasioudi, K.E.; Haegeman, A.; Iliadou, P.; Chaintoutis, S.C.; Dovas, C.I.; Van Borm, S.; Chondrokouki, E.D.; et al. Complete genome sequence of the Lumpy Skin Disease virus isolated from the first reported case in Greece in 2015. Genome Announc. 2017, 5, e00550-17. [Google Scholar] [CrossRef]
- Toplak, I.; Petrović, T.; Vidanović, D.; Lazić, S.; Šekler, M.; Manić, M.; Petrović, M.; Kuhar, U. Complete genome sequence of Lumpy Skin Disease virus isolate SERBIA/Bujanovac/2016, detected during an outbreak in the Balkan area. Genome Announc. 2017, 5, e00882-17. [Google Scholar] [CrossRef]
- Klement, E.; Broglia, A.; Antoniou, S.E.; Tsiamadis, V.; Plevraki, E.; Petrovi’c, T.; Polaček, V.; Debeljak, Z.; Miteva, A.; Alexandrov, T.; et al. Neethling vaccine proved highly effective in controlling Lumpy Skin Disease epidemics in the Balkans. Prev. Vet. Med. 2020, 181, 104595. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Dietze, K.; Wolff, J.; Bergmann, H.; Beltran-Alcrudo, D.; Fahrion, A.; Lamien, C.E.; Busch, F.; Sauter-Louis, C.; Conraths, F.J.; et al. Review: Vaccines and vaccination against Lumpy Skin Disease. Vaccines 2021, 9, 1136. [Google Scholar] [CrossRef]
- Krotova, A.; Mazloum, A.; van Schalkwyk, A.; Prokhvatilova, L.; Gubenko, O.; Byadovskaya, O.; Chvala, I.; Sprygin, A. The characterization and differentiation of recombinant Lumpy Skin Disease isolates using a region within ORF134. Appl. Microbiol. 2023, 3, 35–44. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Y.; Sun, Y.; Bu, Z. Direct Submission by: Important Zoonoses and Severe Exotic Diseases Innovation Teams, Harbin Veterinary Research Institute, No. 678 Haping Road, Xiangfang District, Harbin 150069, P.R. China. 2022. Available online: https://ncbi.nlm.nih.gov/nuccore/OP508345, (accessed on 22 June 2023).
- Li, L.; Wang, Z.; Qi, C.; Liu, S.; Gong, M.; Li, J.; Wu, X.; Wang, Z. Genetic analysis of genome sequence characteristics of two Lumpy Skin Disease viruses isolated from China. BMC Vet. Res. 2022, 18, 426. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yuan, Y.; Shao, J.; Sun, M.; He, W.; Chen, J.; Liu, Q. Genomic characterization of Lumpy Skin Disease virus in southern China. Transbound. Emerg. Dis. 2022, 69, 2788–2799. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.J.; Tu, Y.C.; Wu, C.H.; Huang, C.W.; Ting, L.J.; Huang, Y.L.; Pan, C.H.; Chang, C.Y.; Deng, M.C.; Lee, F. First detection and phylogenetic analysis of Lumpy Skin Disease virus from Kinmen island, Taiwan in 2020. J. Vet. Med. Sci. 2022, 84, 1093–1100. [Google Scholar] [CrossRef]
- Paungpin, W.; Sariya, L.; Chaiwattanarungruengpaisan, S.; Thongdee, M.; Kornmatitsuk, B.; Jitwongwai, A.; Taksinoros, S.; Sutummaporn, K.; Boonmasawai, S.; Nakthong, C. Coding-complete genome sequence of a Lumpy Skin Disease virus isolated during the 2021 Thailand outbreak. Microbiol. Resour. Announc. 2022, 11, e0037522. [Google Scholar] [CrossRef]
- Trinh, T.B.N.; Nguyen, V.T.; Nguyen, T.T.H.; Mai, N.T.A.; Le, P.N.; Lai, T.N.H.; Phan, T.H.; Tran, D.H.; Pham, N.T.; Dam, V.P.; et al. Molecular and histopathological characterization of Lumpy Skin Disease in cattle in northern Vietnam during the 2020-2021 outbreaks. Arch. Virol. 2022, 167, 2143–2149. [Google Scholar] [CrossRef]
- Zan, X.; Huang, H.; Guo, Y.; Di, D.; Fu, C.; Wang, S.; Wu, Y.; Wang, J.; Wang, Y.; Ma, Y.; et al. Molecular characterization of a novel subgenotype of Lumpy Skin Disease virus strain isolated in Inner Mongolia of China. BMC Vet. Res. 2022, 18, 295. [Google Scholar] [CrossRef]
- Sudhakar, S.B.; Mishra, N.; Kalaiyarasu, S.; Jhade, S.K.; Singh, V.P. Genetic and phylogenetic analysis of Lumpy Skin Disease viruses (LSDV) isolated from the first and subsequent field outbreaks in India during 2019 reveals close proximity with unique signatures of historical Kenyan NI-2490/Kenya/KSGP-like field strains. Transbound. Emerg. Dis. 2022, 69, e451–e462. [Google Scholar] [CrossRef]
- Badhy, S.C.; Chowdhury, M.G.A.; Settypalli, T.B.K.; Cattoli, G.; Lamien, C.E.; Fakir, M.A.U.; Akter, S.; Osmani, M.G.; Talukdar, F.; Begum, N.; et al. Molecular characterization of Lumpy Skin Disease virus (LSDV) emerged in Bangladesh reveals unique genetic features compared to contemporary field strains. BMC Vet. Res. 2021, 17, 61. [Google Scholar] [CrossRef]
- Kumar, N.; Chander, Y.; Kumar, R.; Khandelwal, N.; Riyesh, T.; Chaudhary, K.; Shanmugasundaram, K.; Kumar, S.; Kumar, A.; Gupta, M.K.; et al. Isolation and characterization of Lumpy Skin Disease virus from cattle in India. PLoS ONE 2021, 16, e0241022. [Google Scholar] [CrossRef]
- Koirala, P.; Meki, I.K.; Maharjan, M.; Settypalli, B.K.; Manandhar, S.; Yadav, S.K.; Cattoli, G.; Lamien, C.E. Molecular characterization of the 2020 outbreak of Lumpy Skin Disease in Nepal. Microorganisms 2022, 10, 539. [Google Scholar] [CrossRef]
- Parvin, R.; Chowdhury, E.H.; Islam, M.T.; Begum, J.A.; Nooruzzaman, M.; Globig, A.; Dietze, K.; Hoffmann, B.; Tuppurainen, E. Clinical epidemiology, pathology, and molecular investigation of Lumpy Skin Disease outbreaks in Bangladesh during 2020–2021 indicate the re-emergence of an old African strain. Viruses 2022, 14, 2529. [Google Scholar] [CrossRef] [PubMed]
- Maw, M.T.; Khin, M.M.; Hadrill, D.; Meki, I.K.; Settypalli, T.B.K.; Kyin, M.M.; Myint, W.W.; Thein, W.Z.; Aye, O.; Palamara, E.; et al. First report of Lumpy Skin Disease in Myanmar and molecular analysis of the field virus isolates. Microorganisms 2022, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, L.; Bhoyar, R.C.; Jolly, B.; Israni, R.; Vignesh, H.; Scaria, V.; Sivasubbu, S. The genome sequence of Lumpy Skin Disease virus from an outbreak in India suggests a distinct lineage of the virus. Arch. Virol. 2023, 168, 81. [Google Scholar] [CrossRef]
- Massung, R.F.; Esposito, J.J.; Liu, L.I.; Qi, J.; Utterback, T.R.; Knight, J.C.; Aubin, L.; Yuran, T.E.; Parsons, J.M.; Loparev, V.N.; et al. Potential virulence determinants in terminal regions of Variola smallpox virus genome. Nature 1993, 366, 748–751. [Google Scholar] [CrossRef]
- Broyles, S.S. Vaccinia virus transcription. J. Gen. Virol. 2003, 84 Pt 9, 2293–2303. [Google Scholar] [CrossRef]
- Cresawn, S.G.; Condit, R.C. A targeted approach to identification of Vaccinia virus postreplicative transcription elongation factors: Genetic evidence for a role of the H5R gene in Vaccinia transcription. Virology 2007, 363, 333–341. [Google Scholar] [CrossRef]
- D’Costa, S.M.; Bainbridge, T.W.; Kato, S.E.; Prins, C.; Kelley, K.; Condit, R.C. Vaccinia H5 is a multifunctional protein involved in viral DNA replication, postreplicative gene transcription, and virion morphogenesis. Virology 2010, 401, 49–60. [Google Scholar] [CrossRef]
- Sutter, G. A vital gene for modified Vaccinia virus Ankara replication in human cells. Proc. Natl. Acad. Sci. USA 2020, 117, 6289–6291. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Devasurmutt, Y.; Tatu, U. Phylogenomic and structural analysis of the Monkeypox virus shows evolution towards increased stability. Viruses 2023, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- David, M.; Petit, D.; Bertoglio, J. Cell cycle regulation of Rho signaling pathways. Cell Cycle 2012, 11, 3003–3010. [Google Scholar] [CrossRef]
- Liu, R.; Olano, L.R.; Mirzakhanyan, Y.; Gershon, P.D.; Moss, B. Vaccinia virus ankyrin-repeat/F-box protein targets interferon-induced IFITs for proteasomal degradation. Cell. Rep. 2019, 29, 816–828.e6. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.H.; Squire, C.J.; Mercer, A.A. Poxviral ankyrin proteins. Viruses 2015, 7, 709–738. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.; Van Buuren, N.; Burles, K.; Mottet, K.; Wang, Q.; Teale, A. Poxvirus exploitation of the ubiquitin-proteasome system. Viruses 2010, 2, 2356–2380. [Google Scholar] [CrossRef] [PubMed]
- Sonnberg, S.; Fleming, S.B.; Mercer, A.A. Phylogenetic analysis of the large family of poxvirus ankyrin-repeat proteins reveals orthologue groups within and across Chordopoxvirus genera. J. Gen. Virol. 2011, 92, 2596–2607. [Google Scholar] [CrossRef]
- Flannery, J.; Shih, B.; Haga, I.R.; Ashby, M.; Corla, A.; King, S.; Freimanis, G.; Polo, N.; Tse, A.C.; Brackman, C.J.; et al. A novel strain of Lumpy Skin Disease virus causes clinical disease in cattle in Hong Kong. Transbound. Emerg. Dis. 2022, 69, e336–e343. [Google Scholar] [CrossRef]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect. Genet. Evol. 2014, 21, 15–40. [Google Scholar] [CrossRef] [PubMed]
- Lamien, C.E.; Le Goff, C.; Silber, R.; Wallace, D.B.; Gulyaz, V.; Tuppurainen, E.; Madani, H.; Caufour, P.; Adam, T.; El Harrak, M.; et al. Use of the Capripoxvirus homologue of Vaccinia virus 30 kDa RNA polymerase subunit (RPO30) gene as a novel diagnostic and genotyping target: Development of a classical PCR method to differentiate Goat poxvirus from Sheep poxvirus. Vet. Microbiol. 2011, 149, 30–39. [Google Scholar] [CrossRef]
- Dao, T.D.; Tran, L.H.; Nguyen, H.D.; Hoang, T.T.; Nguyen, G.H.; Tran, K.V.D.; Nguyen, H.X.; Van Dong, H.; Bui, A.N.; Bui, V.N. Characterization of Lumpy Skin Disease virus isolated from a giraffe in Vietnam. Transbound. Emerg. Dis. 2022, e3268–e3272. [Google Scholar] [CrossRef]
- Kumar, R.; Godara, B.; Chander, Y.; Kachhawa, J.P.; Dedar, R.K.; Verma, A.; Riyesh, T.; Pal, Y.; Barua, S.; Tripathi, B.N.; et al. Evidence of Lumpy Skin Disease virus infection in camels. Acta Trop. 2023, 242, 106922. [Google Scholar] [CrossRef]
- Oliveira, G.P.; Rodrigues, R.A.L.; Lima, M.T.; Drumond, B.P.; Abrahão, J.S. Poxvirus host range genes and virus-host spectrum: A critical review. Viruses 2017, 9, 331. [Google Scholar] [CrossRef] [PubMed]
- Maluquer de Motes, C. Poxvirus cGAMP nucleases: Clues and mysteries from a stolen gene. PLoS Pathog. 2021, 17, e1009372. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Putative Function | References |
|---|---|---|
| LSDV008 | A putative soluble interferon gamma receptor. | [16] |
| LSDV009 | A putative alpha aminitin-sensitive protein. | [16] |
| LSDV026 | Thought to be unique to LSDV in Capripox, acts as potential rho signaling inhibitor which may explain the formation of nodules as the ‘rho GTPases. | [75] |
| LSDV045 | Involved in replication and recombination. | [16] |
| LSDV046 | Involved in membrane formation. | [16] |
| LSDV067 | Is a putative host range protein, similar to VACCP-C7L, which is suggested to interact with SAMD9. SAMD9 is involved in cell death signaling and is an innate antiviral host factor. | [16,18,76] |
| LSDV061 | Encodes a membrane precursor of immature virions and virus factories. | [16] |
| LSDV062 | Involved in early transcription, otherwise unknown function. | [69,70] |
| LSDV098 | Is a protein that also encodes for the early transcription factor 82 kDa subunit. This is part of the early transcription machinery. | [69,70] |
| LSDV128 | Membrane regulation. | [16] |
| LSDV132 | Unknown. | [16] |
| LSDV133 | DNA ligase. | [16] |
| LSDV134 | Is homologous to the Variola virus B22R-like protein. | [73,74] |
| LSDV076 | Late transcription factor whose Vaccinia homologue H5R is thought to be required both for inclusion of virosomes into crescents and for maturation of immature virions into mature virions. | [71,72] |
| LSDV143-146 | Are kelch-like and ankyrin repeat proteins. | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breman, F.C.; Haegeman, A.; Krešić, N.; Philips, W.; De Regge, N. Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution. Viruses 2023, 15, 1471. https://doi.org/10.3390/v15071471
Breman FC, Haegeman A, Krešić N, Philips W, De Regge N. Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution. Viruses. 2023; 15(7):1471. https://doi.org/10.3390/v15071471
Chicago/Turabian StyleBreman, Floris C., Andy Haegeman, Nina Krešić, Wannes Philips, and Nick De Regge. 2023. "Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution" Viruses 15, no. 7: 1471. https://doi.org/10.3390/v15071471
APA StyleBreman, F. C., Haegeman, A., Krešić, N., Philips, W., & De Regge, N. (2023). Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution. Viruses, 15(7), 1471. https://doi.org/10.3390/v15071471