
Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish, Bacteria and Virus
2.2. Screening of Surface Fibrin VP56 Epitope of GCRV
2.3. Construction of Recombinant Plasmid and Recombinant B. subtilis
2.4. Screening Recombinant Strains and Obtaining Recombinant Spores
2.5. SDS-PAGE and Western Blot (WB)
2.6. Immunofluorescence
2.7. Flow Cytometry
2.8. Preparation of Feed and Oral Immunization
2.9. Sample Collection and Antibody Neutralization Test
2.10. Intestinal Colonization Test of Spores
2.11. Virus Challenge and Viral Load Determination
2.12. Determination of Immune-Related Genes Expression by qRT-PCR
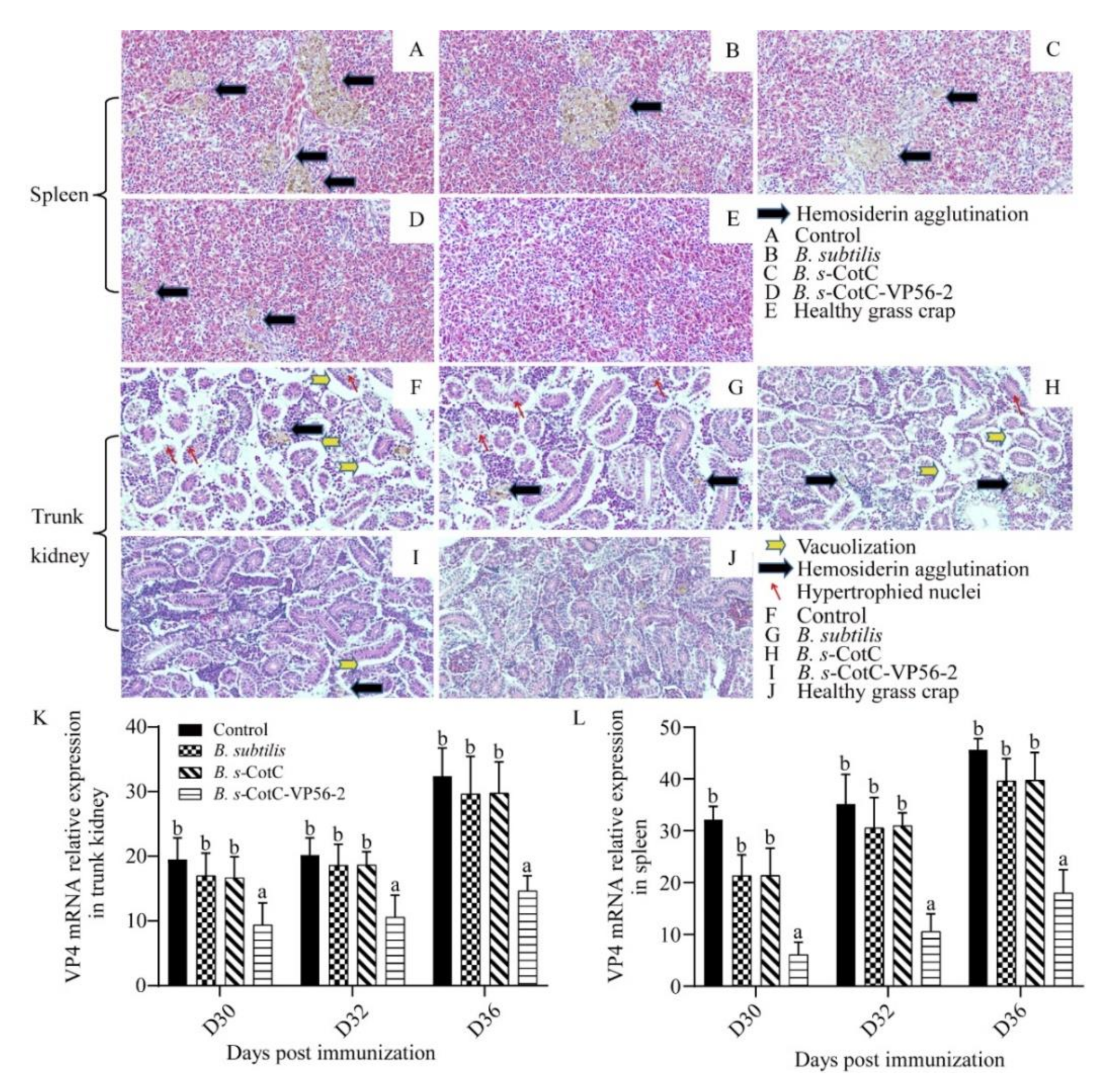
2.13. Histopathological Examination
2.14. Statistical Analysis
3. Results
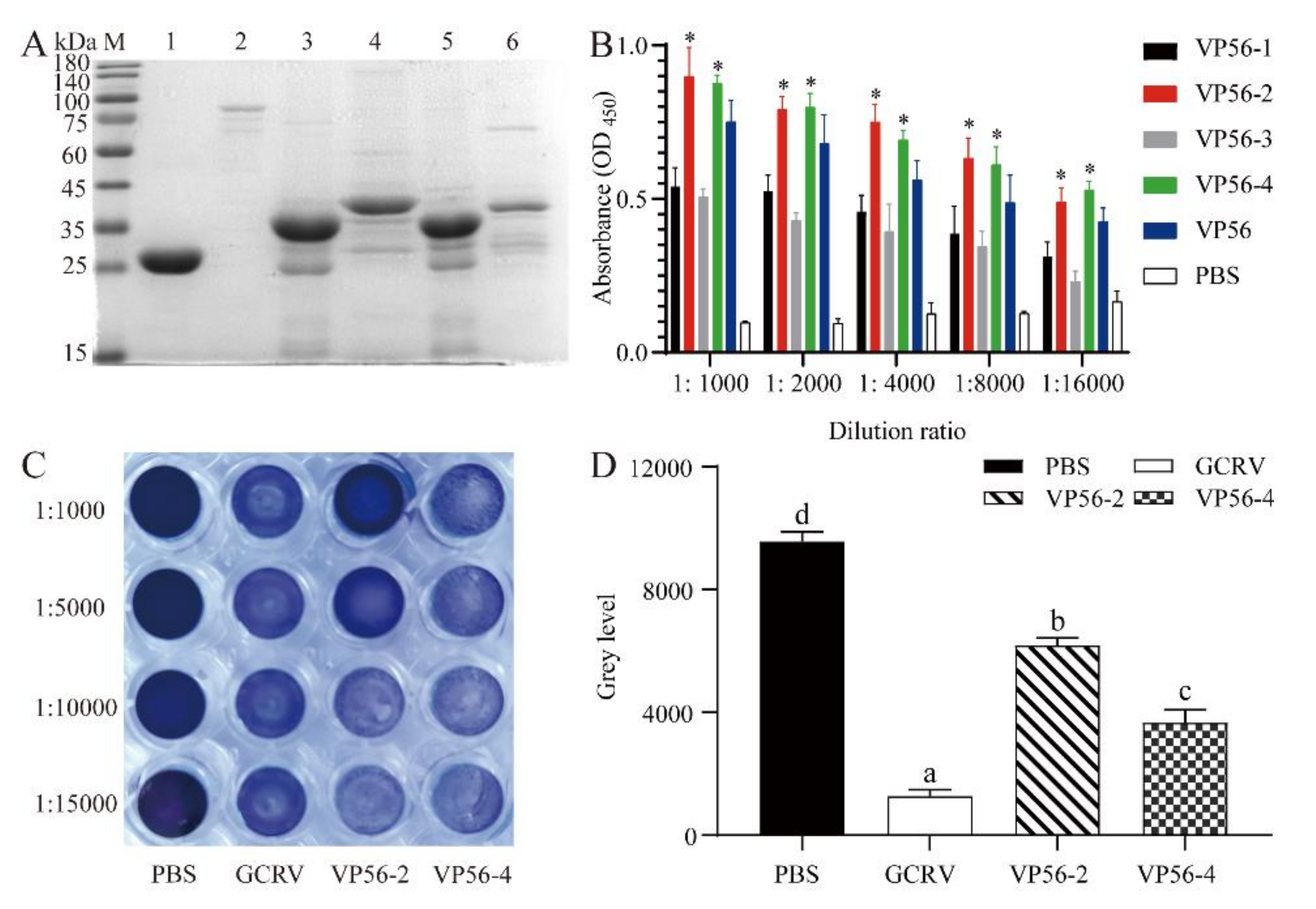
3.1. Expression of VP56 Protein and Screening of Epitopes
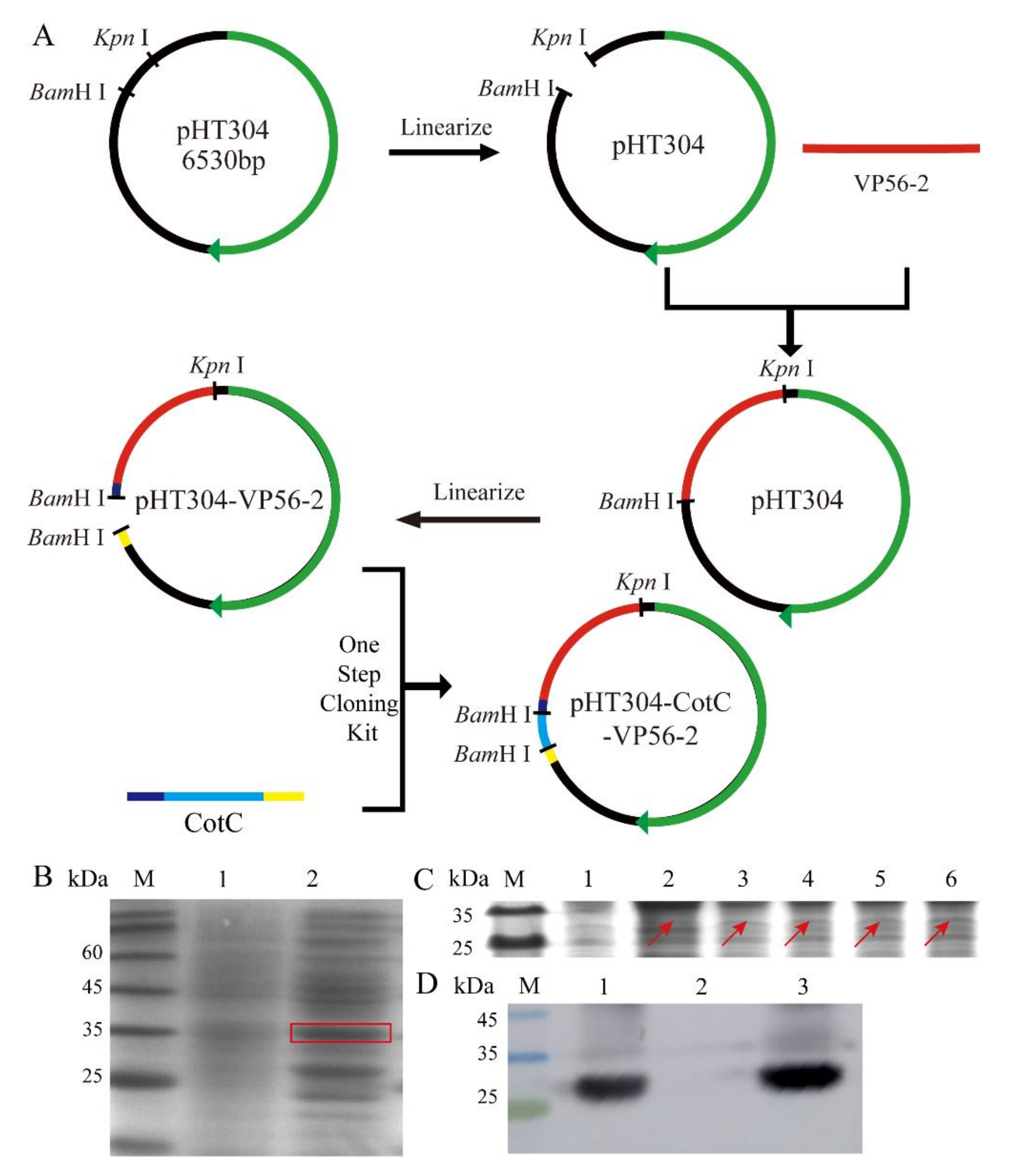
3.2. Successful Construction of pHT304-CotC-VP56-2 Recombinant Plasmid and B. s-CotC-VP56-2 Recombinant Spores
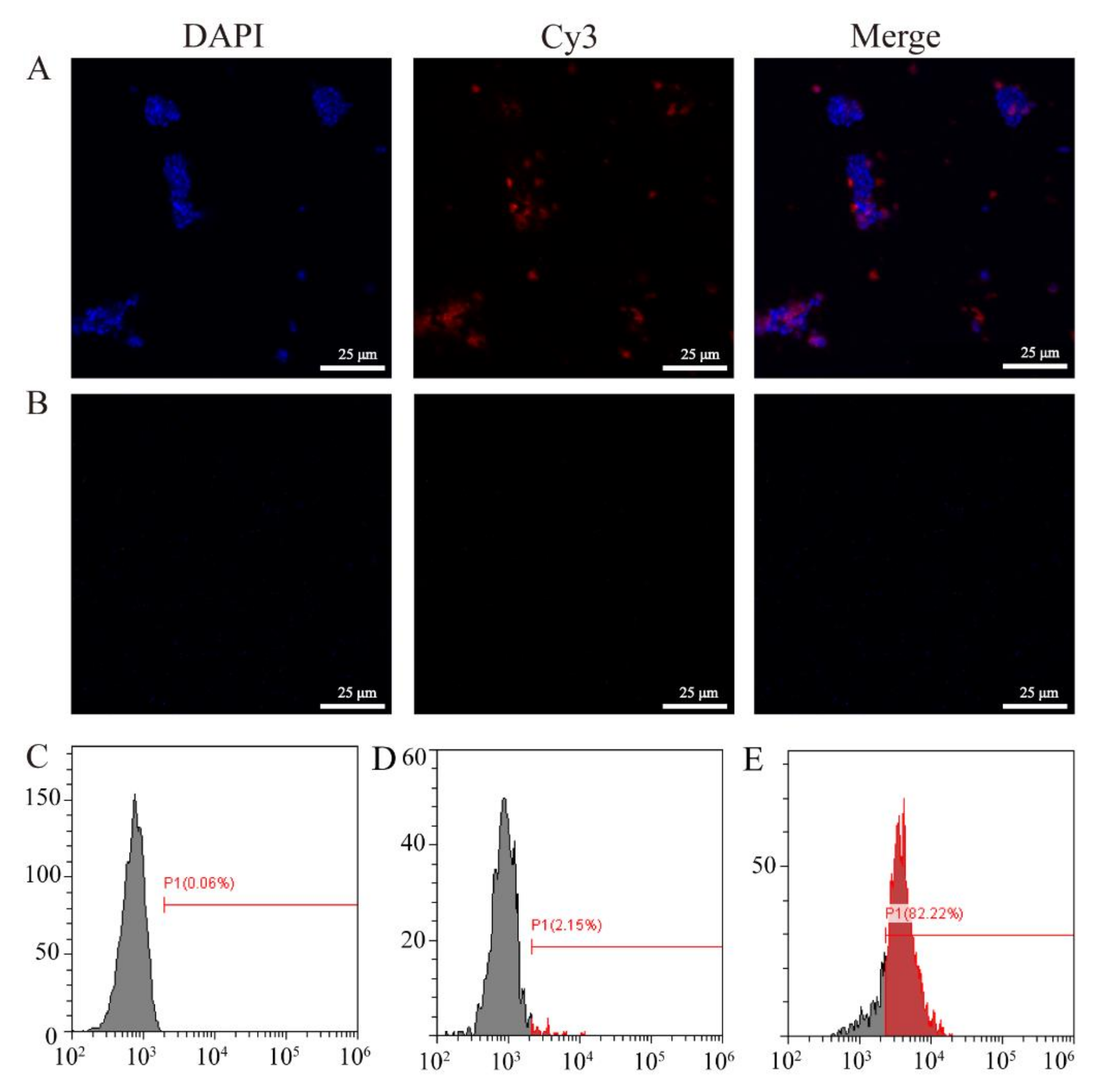
3.3. Immunofluorescence and Flow Cytometry Analysis of VP56-2 Expressed on the Surface of Spores
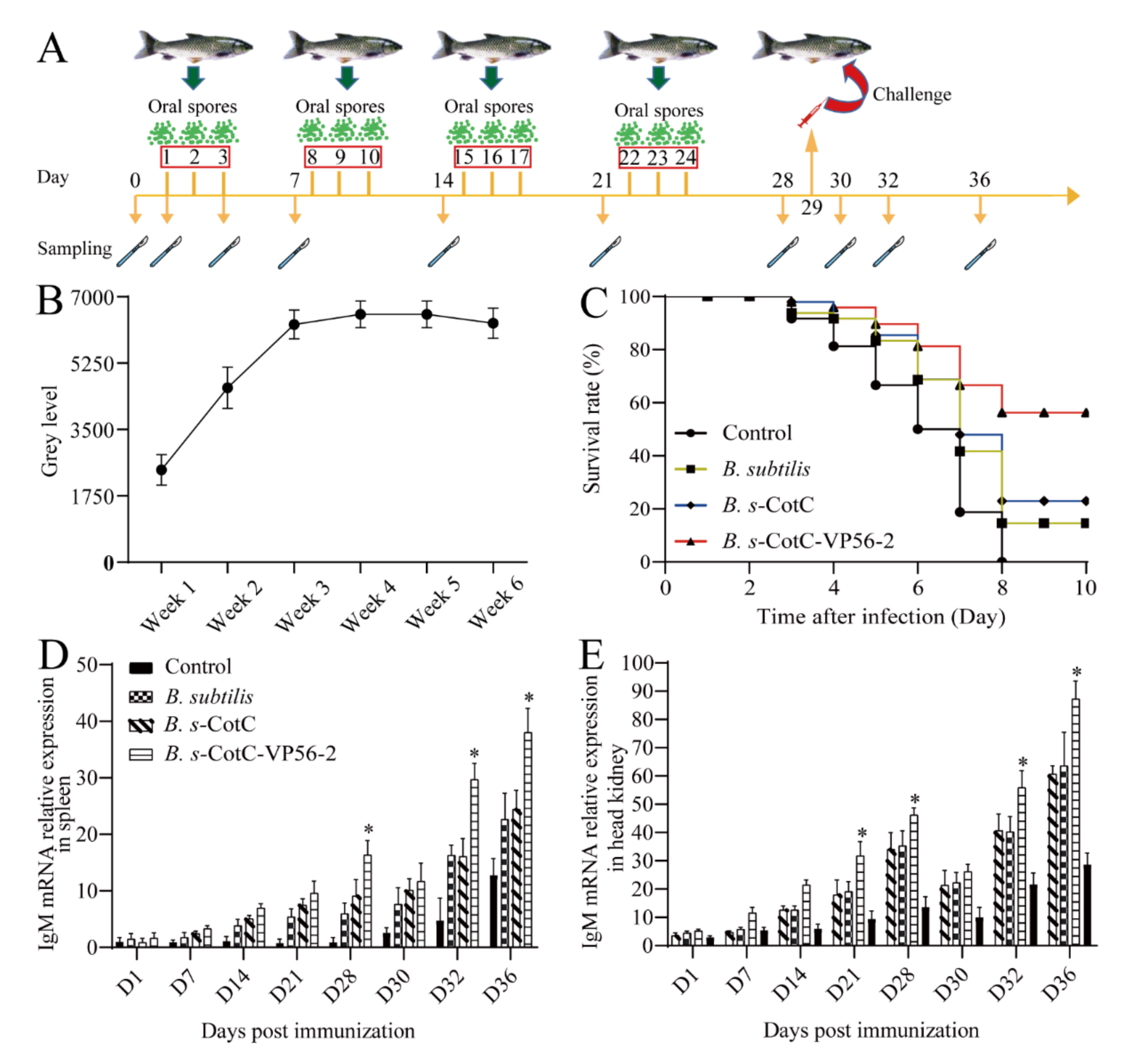
3.4. Colonization of Oral Spores in the Intestine and Evaluation of the Protective Effect of CotC-VP56-2 Spores
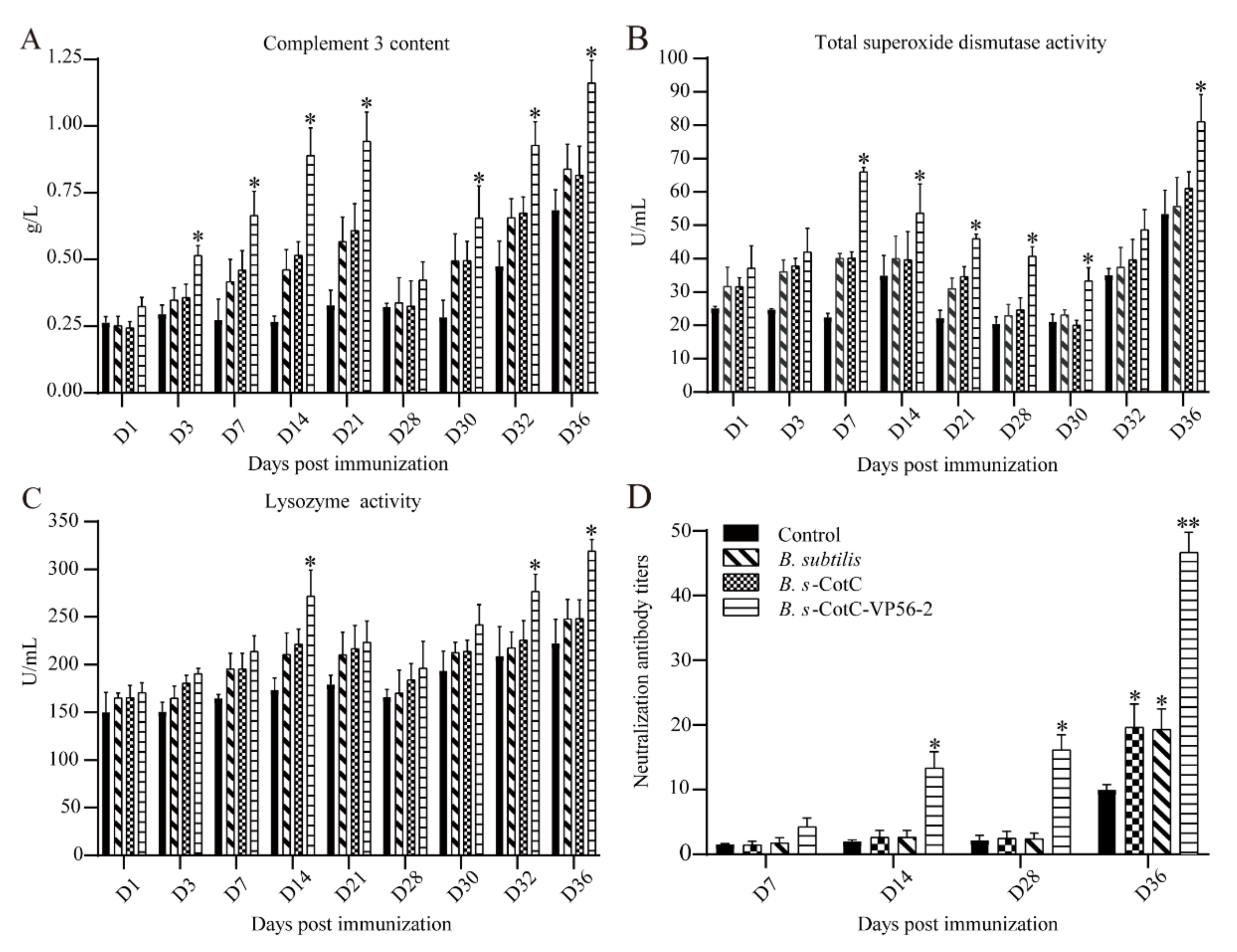
3.5. Detection of Serum Innate Immunity Level
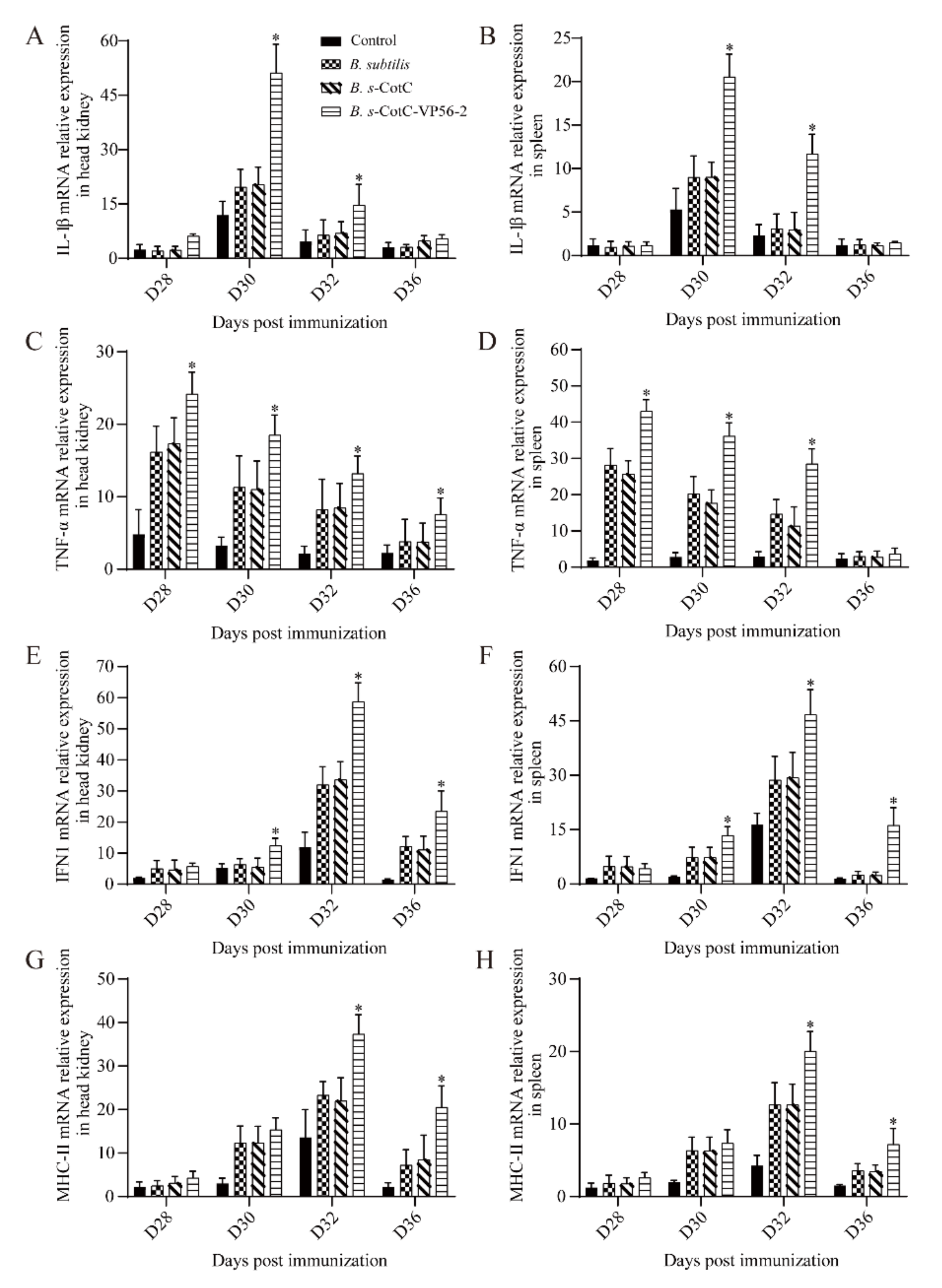
3.6. Expression of Immune-Related Genes in Different Tissues
3.7. B. s-CotC-VP56-2 Oral Vaccine Can Effectively Alleviate Viral Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.-E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef]
- Attoui, H.; Fang, Q.; Jaafar, F.M.; Cantaloube, J.-F.; Biagini, P.; De Micco, P.; De Lamballerie, X. Common evolutionary origin of aquareoviruses and orthoreoviruses revealed by genome characterization of Golden shiner reovirus, Grass carp reovirus, Striped bass reovirus and golden ide reovirus (genus Aquareovirus, family Reoviridae). J. Gen. Virol. 2002, 83, 1941–1951. [Google Scholar] [CrossRef]
- Fan, Y.; Rao, S.; Zeng, L.; Ma, J.; Zhou, Y.; Xu, J.; Zhang, H. Identification and genomic characterization of a novel fish reovirus, Hubei grass carp disease reovirus, isolated in 2009 in China. J. Gen. Virol. 2013, 94, 2266–2277. [Google Scholar] [CrossRef] [Green Version]
- Pei, C.; Ke, F.; Chen, Z.-Y.; Zhang, Q.-Y. Complete genome sequence and comparative analysis of grass carp reovirus strain 109 (GCReV-109) with other grass carp reovirus strains reveals no significant correlation with regional distribution. Arch. Virol. 2014, 159, 2435–2440. [Google Scholar] [CrossRef]
- Ye, X.; Tian, Y.-Y.; Deng, G.-C.; Chi, Y.-Y.; Jiang, X.-Y. Complete genomic sequence of a reovirus isolated from grass carp in China. Virus Res. 2012, 163, 275–283. [Google Scholar] [CrossRef]
- Wang, Q.; Zeng, W.; Liu, C.; Zhang, C.; Wang, Y.; Shi, C.; Wu, S. Complete Genome Sequence of a Reovirus Isolated from Grass Carp, Indicating Different Genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Fang, Q.; Shah, S.; Atanasov, I.C.; Zhou, Z.H. Subnanometer-Resolution Structures of the Grass Carp Reovirus Core and Virion. J. Mol. Biol. 2008, 382, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Guo, H.; Sun, X.; Shao, L.; Fang, Q. Characterization of grass carp reovirus minor core protein VP4. Virol. J. 2012, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Xu, H.; He, Y.; Li, J. Protection of grass carp, Ctenopharyngon idellus (Valenciennes), through oral administration of a subunit vaccine against reovirus. J. Fish Dis. 2011, 34, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Yan, L.; Zhang, X.; Yuan, L.; Fang, Q.; Zhang, Y.-A.; Dai, H. Yeast Surface Display of Capsid Protein VP7 of Grass Carp Reovirus: Fundamental Investigation for the Development of Vaccine Against Hemorrhagic Disease. J. Microbiol. Biotechnol. 2015, 25, 2135–2145. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, G.-L.; Li, D.-L.; Ling, F.; Zhu, B.; Wang, G.-X. The protective immunity against grass carp reovirus in grass carp induced by a DNA vaccination using single-walled carbon nanotubes as delivery vehicles. Fish Shellfish Immunol. 2015, 47, 732–742. [Google Scholar] [CrossRef]
- Su, H.; Su, J. Cyprinid viral diseases and vaccine development. Fish Shellfish Immunol. 2018, 83, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Wang, Q.; Wang, Y.; Zhao, C.; Li, Y.; Shi, C.; Wu, S.; Song, X.; Huang, Q.; Li, S. Immunogenicity of a cell cul-ture-derived inactivated vaccine against a common virulent isolate of grass carp reovirus. Fish Shellfish Immunol. 2016, 54, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sheng, J.; Yu, F.; Wang, H.; Lv, L. Studies on the interaction of Grass Carp Fibulin-4Protein with grass carp reovious outer capsid proteins. Chin. J. Virol. 2018, 34, 557–564. [Google Scholar]
- Yu, F.; Wang, H.; Liu, W.; Lu, L. Grass carp Ctenopharyngodon idella Fibulin-4 as a potential interacting partner for grass carp reovirus outer capsid proteins. Fish Shellfish Immunol. 2016, 48, 169–174. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, L.-F.; Li, Z.-C.; Zhou, X.-Y.; Zhou, Y.; Chen, D.-D.; Li, S.; Zhang, Y.-A. Grass carp reovirus VP56 represses interferon production by degrading phosphorylated IRF7. Fish Shellfish Immunol. 2020, 99, 99–106. [Google Scholar] [CrossRef]
- Rao, Y.; Su, J. Insights into the Antiviral Immunity against Grass Carp (Ctenopharyngodon idella) Reovirus (GCRV) in Grass Carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-I.; Chiou, P.P.; Gong, H.-Y.; Chou, H.-Y. Cloning of the Major Capsid Protein (MCP) of Grouper Iridovirus of Taiwan (TGIV) and Preliminary Evaluation of a Recombinant MCP Vaccine against TGIV. Int. J. Mol. Sci. 2015, 16, 28647–28656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.-D.; Yao, Y.-Y.; Cui, Z.-W.; Zhang, X.-Y.; Peng, K.-S.; Guo, X.; Wang, B.; Zhou, Y.-Y.; Li, S.; Wu, N.; et al. Comparative study of the immunoprotective effect of two DNA vaccines against grass carp reovirus. Fish Shellfish Immunol. 2018, 75, 66–73. [Google Scholar] [CrossRef]
- Xiao, B.; Chi, X.; Zhang, L.; Qu, H.; Liu, Y.; Wang, X.; Zhou, J. Enhanced Expression of GCRV VP6 in CIK Cells by Relative Sequence Optimization. Appl. Biochem. Biotechnol. 2014, 173, 2129–2139. [Google Scholar] [CrossRef]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Lindborg, M.; Magnusson, C.G.; Zargari, A.; Scheynius, A.; Whitley, P.; Schmidt, M.; Crameri, R. Selective Cloning of Allergens from the Skin Colonizing Yeast Malassezia furfur by Phage Surface Display Technology. J. Investig. Dermatol. 1999, 113, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boder, E.T.; Wittrup, K.D. Yeast surface display for screening combinatorial polypeptide libraries. Nat. Biotechnol. 1997, 15, 553–557. [Google Scholar] [CrossRef]
- Hu, S.; Kong, J.; Sun, Z.; Han, L.; Kong, W.; Yang, P. Heterologous protein display on the cell surface of lactic acid bacteria mediated by the s-layer protein. Microb. Cell Factories 2011, 10, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; An, Y.; Zabed, H.; Guo, Q.; Yang, M.; Yuan, J.; Li, W.; Sun, W.; Qi, X. Bacillus subtilis Spore Surface Display Technology: A Review of Its Development and Applications. J. Microbiol. Biotechnol. 2019, 29, 179–190. [Google Scholar]
- Cheon, J.; Kim, S.B.; Park, S.W.; Han, J.K.; Kim, P. Characterization of L-Arabinose Isomerase in Bacillus subtilis, a GRAS Host, for the Production of Edible Tagatose. Food Biotechnol. 2009, 23, 8–16. [Google Scholar] [CrossRef]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface Display of Recombinant Proteins on Bacillus subtilis Spores. J. Bacteriol. 2001, 183, 6294–6301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.-Y.; Chen, D.-D.; Cui, Z.-W.; Zhang, X.-Y.; Zhou, Y.-Y.; Guo, X.; Li, A.-H.; Zhang, Y.-A. Oral vaccination of tilapia against Streptococcus agalactiae using Bacillus subtilis spores expressing Sip. Fish Shellfish Immunol. 2019, 86, 999–1008. [Google Scholar] [CrossRef]
- Lin, P.; Yuan, H.; Du, J.; Liu, K.; Liu, H.; Wang, T. Progress in research and application development of surface display technology using Bacillus subtilis spores. Appl. Microbiol. Biot. 2020, 104, 2319–2331. [Google Scholar] [CrossRef]
- Jiang, H.; Bian, Q.; Zeng, W.; Ren, P.; Sun, H.; Lin, Z.; Tang, Z.; Zhou, X.; Wang, Q.; Wang, Y.; et al. Oral delivery of Bacillus subtilis spores expressing grass carp reovirus VP4 protein produces protection against grass carp reovirus infection. Fish Shellfish Immunol. 2019, 84, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Vakharia, V.N.; Zhou, Y.; Jiang, N.; Liu, W.; Meng, Y.; Li, Y.; Xue, M.; Zhang, J.; Zeng, L.; et al. A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus). Pathogenes 2020, 9, 945. [Google Scholar] [CrossRef]
- Magadan, S.; Sunyer, O.J.; Boudinot, P. Unique Features of Fish Immune Repertoires: Particularities of Adaptive Immunity Within the Largest Group of Vertebrates. In Pathogen-Host Interactions: Antigenic Variation v. Somatic Adaptations; Results and Problems in Cell Differentiation; Springer: Cham, Switzerland, 2015; Volume 57, pp. 235–264. [Google Scholar]
- Salinas, I.; Zhang, Y.-A.; Sunyer, J.O. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef] [Green Version]
- Bengtén, E.; Wilson, M. Antibody Repertoires in Fish. In Pathogen-Host Interactions: Antigenic Variation v. Somatic Adaptations; Results and Problems in Cell Differentiation; Springer: Cham, Switzerland, 2015; Volume 57, pp. 193–234. [Google Scholar]
- Mutoloki, S.; Munang’Andu, H.M.; Evensen, Ø. Oral Vaccination of Fish—Antigen Preparations, Uptake, and Immune Induction. Front. Immunol. 2015, 6, 519. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; La Patra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.-H.; Wu, Y.-C.; Chi, S.-C. Immune gene expressions in grouper larvae (Epinephelus coioides) induced by bath and oral vaccinations with inactivated betanodavirus. Fish Shellfish Immunol. 2014, 40, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, N.A.; Castro, R.; Abos, B.; Rodriguez Saint-Jean, S.S.; Perez-Prieto, S.I.; Tafalla, C. The pyloric caeca area is a major site for IgM+ and IgT+ B cell recruitment in response to oral vaccination in rainbow trout. PLoS ONE 2013, 8, e66118. [Google Scholar]
- Huo, X.; Fan, C.; Ai, T.; Su, J. The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection. Vaccines 2020, 8, 567. [Google Scholar] [CrossRef]
- Sun, H.; Lin, Z.; Zhao, L.; Chen, T.; Shang, M.; Jiang, H.; Tang, Z.; Zhou, X.; Shi, M.; Zhou, L.; et al. Bacillus subtilis spore with surface display of paramyosin from Clonorchis sinensis potentializes a promising oral vaccine candidate. Parasites Vectors 2018, 11, 156. [Google Scholar] [CrossRef]
- Tang, Z.; Sun, H.; Chen, T.; Lin, Z.; Jiang, H.; Zhou, X.; Shi, C.; Pan, H.; Chang, O.; Ren, P.; et al. Oral delivery of Bacillus subtilis spores expressing cysteine protease of Clonorchis sinensis to grass carp (Ctenopharyngodon idellus): Induces immune responses and has no damage on liver and intestine function. Fish Shellfish Immunol. 2017, 64, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, W.; Tian, Y.; Mao, Q.; Lv, X.; Shang, M.; Li, X.; Yu, X.; Huang, Y. Surface display of Clonorchis sinensis enolase on Bacillus subtilis spores potentializes an oral vaccine candidate. Vaccine 2014, 32, 1338–1345. [Google Scholar] [CrossRef]
- Zhou, Z.; Xia, H.; Hu, X.; Huang, Y.; Li, Y.; Li, L.; Ma, C.; Chen, X.; Hu, F.; Xu, J.; et al. Oral administration of a Bacillus subtilis spore-based vaccine expressing Clonorchis sinensis tegumental protein 22.3kDa confers protection against Clonorchis sinensis. Vaccine 2008, 26, 1817–1825. [Google Scholar] [CrossRef]
- Qu, H.; Xu, Y.; Sun, H.; Lin, J.; Yu, J.; Tang, Z.; Shen, J.; Liang, C.; Li, S.; Chen, W.; et al. Systemic and local mucosal immune responses induced by orally delivered Bacillus subtilis spore expressing leucine aminopeptidase 2 of Clonorchis sinensis. Parasitol. Res. 2014, 113, 3095–3103. [Google Scholar] [CrossRef] [PubMed]
- Su, J.G.; Zhang, R.F.; Dong, Z.J.; Yang, C.R. Evaluation of internal control genes for qRT-PCR normalization in tissues and cell culture for antiviral studies of grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2011, 30, 830–835. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Kong, X.; Zhao, X.; Pei, C.; Li, L. A C-type lectin, Nattectin-like protein (CaNTC) in Qihe crucian carp Carassius auratus: Binding ability with LPS, PGN and various bacteria, and agglutinating activity against bacteria. Fish Shellfish Immunol. 2017, 67, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Reiter, D.M.; Frierson, J.M.; Halvorson, E.E.; Kobayashi, T.; Dermody, T.S.; Stehle, T. Crystal Structure of Reovirus Attachment Protein σ1 in Complex with Sialylated Oligosaccharides. PLOS Pathog. 2011, 7, e1002166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Xie, H.; Zeng, W.; Wang, L.; Liu, C.; Wu, J.; Wang, Y.; Li, Y.; Bergmann, S.M. Development of indirect im-munofluorescence assay for TCID50 measurement of grass carp reovirus genotype II without cytopathic effect onto cells. Microb. Pathog. 2018, 114, 68–74. [Google Scholar] [CrossRef]
- Barton, E.S.; Forrest, J.C.; Connolly, J.L.; Chappell, J.D.; Liu, Y.; Schnell, F.J.; Nusrat, A.; Parkos, C.A.; Dermody, T.S. Junction Adhesion Molecule Is a Receptor for Reovirus. Cell 2001, 104, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Jiao, Z.; Dong, J.; Sun, C.; Jiang, X.; Ye, X. Grass carp reovirus-GD108 fiber protein is involved in cell attachment. Virus Genes 2017, 53, 613–622. [Google Scholar] [CrossRef]
- Lee, S.-M.; Wu, C.-K.; Plieskatt, J.L.; Miura, K.; Hickey, J.M.; King, C.R. N-Terminal Pfs230 Domain Produced in Baculovirus as a Biological Active Transmission-Blocking Vaccine Candidate. Clin. Vaccine Immunol. 2017, 24, 00140-17. [Google Scholar] [CrossRef] [Green Version]
- Mauriello, E.M.; Duc, L.H.; Isticato, R.; Cangiano, G.; Hong, H.A.; De Felice, M.; Ricca, E.; Cutting, S.M. Display of heterologous antigens on the Bacillus subtilis spore coat using CotC as a fusion partner. Vaccine 2004, 22, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Zwollo, P.; Cole, S.; Bromage, E.; Kaattari, S. B Cell Heterogeneity in the Teleost Kidney: Evidence for a Maturation Gradient from Anterior to Posterior Kidney. J. Immunol. 2005, 174, 6608–6616. [Google Scholar] [CrossRef] [Green Version]
- De Wit, J.; Souwer, Y.; Jorritsma, T.; Bos, H.K.; Brinke, A.T.; Neefjes, J.; Van Ham, S.M. Antigen-Specific B Cells Reactivate an Effective Cytotoxic T Cell Response against Phagocytosed Salmonella through Cross-Presentation. PLoS ONE 2010, 5, e13016. [Google Scholar] [CrossRef]
- Chiu, S.-T.; Tsai, R.-T.; Hsu, J.-P.; Liu, C.-H.; Cheng, W. Dietary sodium alginate administration to enhance the non-specific immune responses, and disease resistance of the juvenile grouper Epinephelus fuscoguttatus. Aquaculture 2008, 277, 66–72. [Google Scholar] [CrossRef]
- Osman, A.G.; Koutb, M.; Sayed, A.E.-D.H. Use of hematological parameters to assess the efficiency of quince (Cydonia oblonga Miller) leaf extract in alleviation of the effect of ultraviolet—A radiation on African catfish Clarias gariepinus (Burchell, 1822). J. Photochem. Photobiol. B Biol. 2010, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boshra, H.; Li, J.; Sunyer, J. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, Y.; Zhang, J.; Yuan, G.; Liu, X.; Ai, T.; Su, J. Astragalus polysaccharides, chitosan and poly(I:C) obviously enhance inactivated Edwardsiella ictaluri vaccine potency in yellow catfish Pelteobagrus fulvidraco. Fish Shellfish Immunol. 2019, 87, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The Interleukin-1 Family: Back to the Future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef] [Green Version]
- Baud, V.; Karin, M. Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol. 2001, 11, 372–377. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Zhang, Y.-B.; Jiang, J.; Wang, B.; Chen, C.; Zhang, J.; Gui, J.-F. Gig1, a novel antiviral effector involved in fish interferon response. Virology 2014, 448, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Zhu, W.; Zhang, Y.; Liao, Z.; Wu, C.; Yang, C.; Zhang, Y.; Xiao, S.; Su, J. Broad-Spectrum Robust Direct Bactericidal Activity of Fish IFN Ф1 Reveals an Antimicrobial Peptide-like Function for Type I IFNs in Vertebrates. J. Immunol. 2021, 206, 1337–1347. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Innate immune recognition of viral infection. Nat. Immunol. 2006, 7, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Akira, S. Recognition of viruses by innate immunity. Immunol. Rev. 2007, 220, 214–224. [Google Scholar] [CrossRef]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 2005, 437, 1167–1172. [Google Scholar] [CrossRef]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-κB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wang, Y.-Y.; Han, K.-J.; Li, L.-Y.; Zhai, Z.; Shu, H.-B. VISA Is an Adapter Protein Required for Virus-Triggered IFN-β Signaling. Mol. Cell 2005, 19, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Du, H.; Liu, L.; You, X.; Wu, M.; Liao, Z. MHC class II alpha, beta and MHC class II-associated invariant chains from Chinese sturgeon (Acipenser sinensis) and their response to immune stimulation. Fish Shellfish Immunol. 2017, 70, 1–12. [Google Scholar] [CrossRef]
- Somamoto, T.; Yoshiura, Y.; Nakanishi, T.; Ototake, M. Molecular cloning and characterization of two types of CD8α from ginbuna crucian carp, Carassius auratus langsdorfii. Dev. Comp. Immunol. 2005, 29, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Wang, X.; Chen, C.; Zhou, H. Characterization of grass carp (Ctenopharyngodon idellus) Foxp1a/1b/2: Evidence for their involvement in the activation of peripheral blood lymphocyte subpopulations. Fish Shellfish Immunol. 2010, 28, 289–295. [Google Scholar] [CrossRef]
- Wu, M.; Li, H.; Chen, X.; Jiang, Y.; Jiang, W. Studies on the clinical symptoms, virus distribution, and mRNA expression of several antiviral immunity-related genes in grass carp after infection with genotype II grass carp reovirus. Arch. Virol. 2020, 165, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Su, J.G. Inducible Nitric Oxide Synthase (iNOS) mediates vascular rndothelial cell apoptosis in grass carp reovirus (GCRV)-induced hemorrhage. Int. J. Mol. Sci. 2019, 20, 6335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Huo, X.; Wang, Z.; Yuan, G.; Liu, X.; Ai, T.; Su, J. Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection. Viruses 2022, 14, 30. https://doi.org/10.3390/v14010030
Gao Y, Huo X, Wang Z, Yuan G, Liu X, Ai T, Su J. Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection. Viruses. 2022; 14(1):30. https://doi.org/10.3390/v14010030
Chicago/Turabian StyleGao, Yang, Xingchen Huo, Zhensheng Wang, Gailing Yuan, Xiaoling Liu, Taoshan Ai, and Jianguo Su. 2022. "Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection" Viruses 14, no. 1: 30. https://doi.org/10.3390/v14010030
APA StyleGao, Y., Huo, X., Wang, Z., Yuan, G., Liu, X., Ai, T., & Su, J. (2022). Oral Administration of Bacillus subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection. Viruses, 14(1), 30. https://doi.org/10.3390/v14010030