
Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance

 , , ,
, , ,  ,
,  , , ,
, , , 
Abstract
1. Introduction
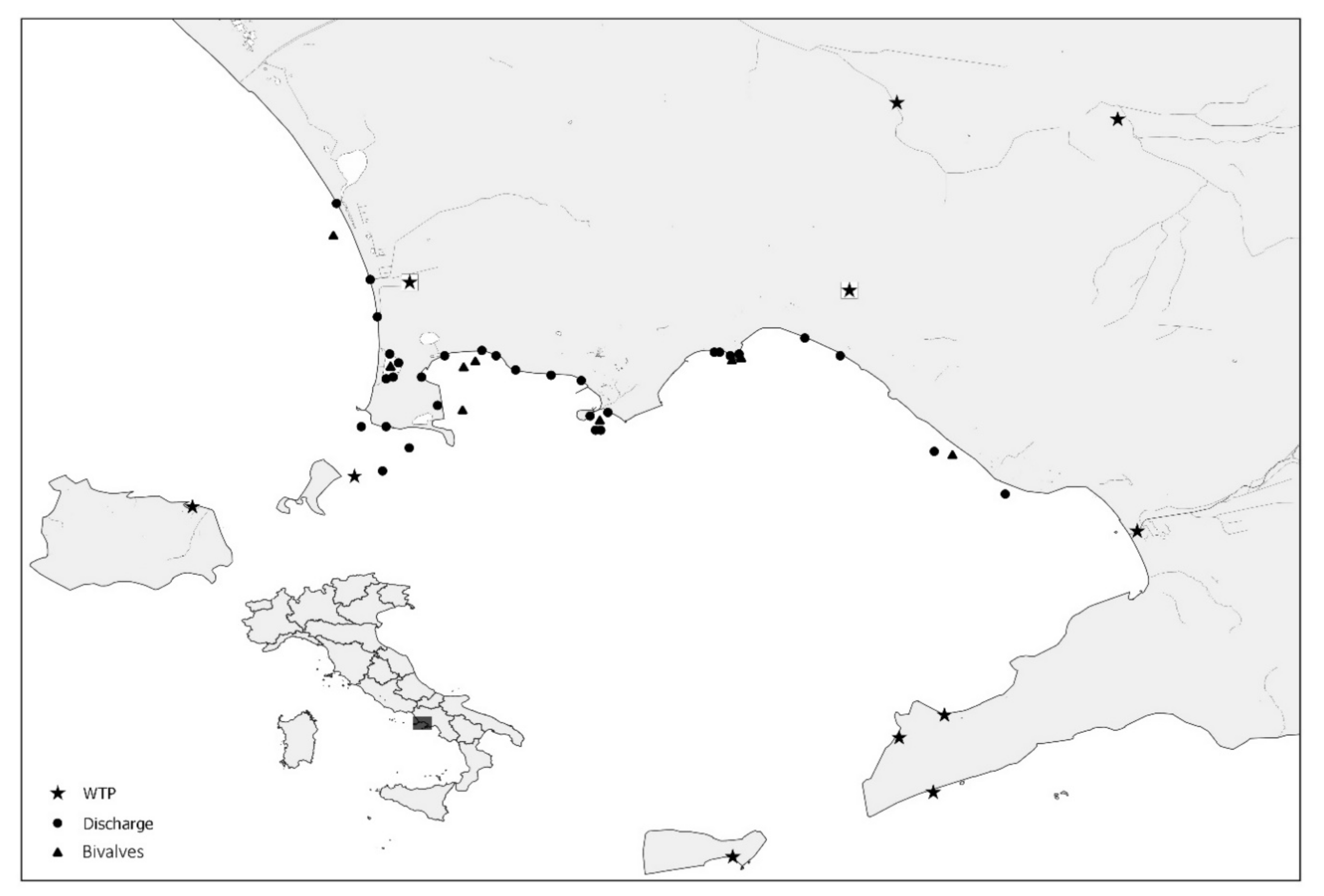
2. Materials and Methods
2.1. Water Samples
2.2. Shellfish Samples
2.3. Real Time PCR and Nested RT-PCR
3. Results
4. Discussion
- Other members of the NRL for Human Viral Hepatitis:
- Michele Equestre—Department of Infectious Diseases, Istituto Superiore di Sanità, Rome, Italy
- Stefania Taffon—Department of Infectious Diseases, Istituto Superiore di Sanità, Rome, Italy
- Angela Costantino—Department of Infectious Diseases, Istituto Superiore di Sanità, Rome, Italy
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koff, R.S. Hepatitis A. Lancet 1998, 351, 1643–1649. [Google Scholar] [CrossRef]
- Cuthbert, J.A. Hepatitis A: Old and New. Clin. Microbiol. Rev. 2001, 14, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, K.H.; Wiersma, S.T. Hepatitis A virus seroprevalence by age and world region, 1990 and 2005. Vaccine 2010, 28, 6653–6657. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Hepatitis A. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-a (accessed on 23 December 2020).
- World Health Organization. The Global Prevalence of Hepatitis A Virus Infection and Susceptibility: A Systematic Review. Available online: Whqlibdoc.who.int/hq/2010/WHO_IVB_10.01_eng.pdf (accessed on 23 December 2020).
- Daniels, D.; Grytdal, S.; Wasley, A. Surveillance for acute viral hepatitis—United States, 2007. MMWR Surveill. Summ. 2009, 58, 1–27. [Google Scholar] [PubMed]
- Romanò, L.; Paladini, S.; Tagliacarne, C.; Zappa, A.; Zanetti, A.R. The changing face of the epidemiology of type A, B, and D viral hepatitis in Italy, following the implementation of vaccination. Vaccine 2009, 27, 3439–3442. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, C.; Alfonsi, V.; Bruni, R.; Busani, L.; Ciccaglione, A.R.; De Medici, D.; Di Pasquale, S.; Equestre, M.; Escher, M.; Montaño-Remacha, M.C.; et al. Ongoing outbreak of hepatitis A in Italy: Preliminary report as of 31 May 2013. Eurosurveillance 2013, 18. [Google Scholar] [CrossRef]
- La Rosa, G.; Della Libera, S.; Iaconelli, M.; Ciccaglione, A.R.; Bruni, R.; Taffon, S.; Equestre, M.; Alfonsi, V.; Rizzo, C.; Tosti, M.E.; et al. Surveillance of hepatitis A virus in urban sewages and comparison with cases notified in the course of an outbreak, Italy 2013. BMC Infect. Dis. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Scavia, G.; Alfonsi, V.; Taffon, S.; Escher, M.; Bruni, R.; De Medici, D.; Di Pasquale, S.; Guizzardi, S.; Cappelletti, B.; Iannazzo, S.; et al. A large prolonged outbreak of hepatitis A associated with consumption of frozen berries, Italy, 2013–14. J. Med. Microbiol. 2017, 66, 342–349. [Google Scholar] [CrossRef]
- Costantino, A.; Coppola, N.; Spada, E.; Bruni, R.; Taffon, S.; Equestre, M.; Marcantonio, C.; Sagnelli, C.; Dell’Isola, C.; Tosone, G.; et al. Hepatitis A virus strains circulating during 1997-2015 in Campania, a Southern Italy region with periodic outbreaks. J. Med. Virol. 2017, 89, 1931–1936. [Google Scholar] [CrossRef]
- Chironna, M.; Prato, R.; Sallustio, A.; Martinelli, D.; Tafuri, S.; Quarto, M.; Germinario, C. Hepatitis A in Puglia (South Italy) after 10 years of universal vaccination: Need for strict monitoring and catch-up vaccination. BMC Infect. Dis. 2012, 12. [Google Scholar] [CrossRef]
- Rossati, A.; Bargiacchi, O.; Kroumova, V.; Nebbiolo, C.; Tacca, M.G.; Pratillo, S.; Garavelli, P.L. Outbreak of severe Hepatitis A in Eastern Piedmont, Italy. Infez. Med. Riv. Period. Eziolog. Epidemiol. Diagn. Clin. Ter. Patol. Infett. 2017, 25, 344–346. [Google Scholar]
- Lanini, S.; Minosse, C.; Vairo, F.; Garbuglia, A.; Di Bari, V.; Agresta, A.; Rezza, G.; Puro, V.; Pendenza, A.; Loffredo, M.R.; et al. A large ongoing outbreak of hepatitis A predominantly affecting young males in Lazio, Italy; August 2016–March 2017. PLoS ONE 2017, 12, e0185428. [Google Scholar] [CrossRef] [PubMed]
- Comelli, A.; Izzo, I.; Casari, S.; Spinetti, A.; Bergamasco, A.; Castelli, F. Hepatitis A outbreak in men who have sex with men (MSM) in Brescia (Northern Italy), July 2016–July 2017. Infez. Med. Riv. Period. Eziolog. Epidemiol. Diagn. Clin. Ter. Patol. Infett. 2018, 26, 46–51. [Google Scholar]
- Mauro, M.V.; Greco, F.; Tenuta, R.; Apuzzo, G.; Costantino, A.; Bruni, R.; Equestre, M.; Ciccaglione, A.R.; Giraldi, C.; Mastroianni, A. Hepatitis A outbreak affecting men who have sex with men (MSM) in South Italy. New Microbiol. 2019, 42, 181–183. [Google Scholar]
- Suffredini, E.; Proroga, Y.T.R.; Di Pasquale, S.; Di Maro, O.; Losardo, M.; Cozzi, L.; Capuano, F.; De Medici, D. Occurrence and Trend of Hepatitis A Virus in Bivalve Molluscs Production Areas Following a Contamination Event. Food Environ. Virol. 2017, 9, 423–433. [Google Scholar] [CrossRef]
- Plunkett, J.; Mandal, S.; Balogun, K.; Beebeejaun, K.; Ngui, S.L.; Ramsay, M.; Edelstein, M. Hepatitis A outbreak among men who have sex with men (MSM) in England, 2016–2018: The contribution of past and current vaccination policy and practice. Vaccine X 2019, 1. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Hepatitis A Outbreaks in the EU/EEA Mostly Affecting Men Who Have Sex with Men (Rapid Risk Assessment; Third Update, 12 Sep 2018). Stockholm: ECDC. 2018. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-hepatitis-outbreak-eueea-mostly-affecting-men-who-have-sex-men-2 (accessed on 23 December 2020).
- World Health Organization. Guidelines for Environmental Surveillance of Poliovirus Circulation; World Health Organization: Geneva, Switzerland, 2003; Available online: https://apps.who.int/iris/handle/10665/67854 (accessed on 23 December 2020).
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, Evaluation, and Standardization of a Real-Time TaqMan Reverse Transcription-PCR Assay for Quantification of Hepatitis A Virus in Clinical and Shellfish Samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef]
- ISO/TS 15216-1:2013: Microbiology of Food and Animal Feed—Horizontal Method for Determination of Hepatitis A Virus and Norovirus in Food Using Real-Time RT-PCR—Part 1: Method for Quantification; International Organization for Standardization: Geneva, Switzerland, 2013.
- ISO 15216-1:2017: Microbiology of the Food Chain—Horizontal Method for Determination of Hepatitis A Virus and Norovirus Using Real-Time RT-PCR—Part 1: Method for Quantification; International Organization for Standardization: Geneva, Switzerland, 2017.
- Taffon, S.; Bidini, G.; Vichi, F.; Corti, G.; Genovese, D.; Kondili, L.A.; Bindi, R.; Armellini, F.; Leoncini, F.; Bartoloni, A.; et al. A unique HAV strain circulated in patients with acute HAV infection with different risk exposures in Tuscany, Italy. J. Clin. Virol. 2011, 50, 142–147. [Google Scholar] [CrossRef]
- Pintó, R.M.; Costafreda, M.I.; Bosch, A. Risk Assessment in Shellfish-Borne Outbreaks of Hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef]
- Spada, E.; Genovese, D.; Tosti, M.E.; Mariano, A.; Cuccuini, M.; Proietti, L.; Di Giuli, C.; Lavagna, A.; Crapa, G.E.; Morace, G.; et al. An outbreak of hepatitis A virus infection with a high case-fatality rate among injecting drug users. J. Hepatol. 2005, 43, 958–964. [Google Scholar] [CrossRef]
- Bosch, A.; Guix, S.; Sano, D.; Pintó, R.M. New tools for the study and direct surveillance of viral pathogens in water. Curr. Opin. Biotechnol. 2008, 19, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Barrett, C.E.; Pape, B.J.; Benedict, K.M.; Foster, M.A.; Roberts, V.A.; Rotert, K.; Mattioli, M.C.; Yoder, J.S. Impact of Public Health Interventions on Drinking Water–Associated Outbreaks of Hepatitis A—United States, 1971–2017. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, G.; Rossi, L.M.G.; Forbi, J.C.; De Paula, V.S.; Purdy, M.A.; Xia, G.; Khudyakov, Y.E. Hepatitis A virus: Host interactions, molecular epidemiology and evolution. Infect. Genet. Evol. 2014, 21, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Franco, E. Hepatitis A: Epidemiology and prevention in developing countries. World J. Hepatol. 2012, 4, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Wasley, A.; Fiore, A.; Bell, B.P. Hepatitis A in the Era of Vaccination. Epidemiol. Rev. 2006, 28, 101–111. [Google Scholar] [CrossRef]
- Van der Poel, W.; Rzezutka, A. Hepatitis A. In Global Water Pathogen Project; Rose, J.B., Jiménez-Cisneros, B., Eds.; 2017. Part 3 Viruses. Available online: http://www.waterpathogens.org/book/hepatitisA (accessed on 23 December 2020).
- Brooks, H.A.; Gersberg, R.M.; Dhar, A.K. Detection and quantification of hepatitis A virus in seawater via real-time RT-PCR. J. Virol. Methods 2005, 127, 109–118. [Google Scholar] [CrossRef] [PubMed]
- De Paula, V.S.; Diniz-Mendes, L.; Villar, L.; Luz, S.; Silva, L.; Jesus, M.; Da Silva, N.; Gaspar, A. Hepatitis A virus in environmental water samples from the Amazon Basin. Water Res. 2007, 41, 1169–1176. [Google Scholar] [CrossRef]
- Pintó, R.M.; Alegre, D.; Domínguez, A.; El-Senousy, W.M.; Sánchez, G.; Villena, C.; Costafreda, M.I.; Aragonés, L.; Bosch, A. Hepatitis A virus in urban sewage from two Mediterranean countries. Epidemiol. Infect. 2006, 135, 270–273. [Google Scholar] [CrossRef]
- Villar, L.M.; De Paula, V.; Diniz-Mendes, L.; Guimarães, F.; Ferreira, F.; Shubo, T.; Miagostovich, M.; Lampe, E.; Gaspar, A. Molecular detection of hepatitis A virus in urban sewage in Rio de Janeiro, Brazil. Lett. Appl. Microbiol. 2007, 45, 168–173. [Google Scholar] [CrossRef]
- Schlindwein, A.D.; Rigotto, C.; Simões, C.M.O.; Barardi, C.R.M. Detection of enteric viruses in sewage sludge and treated wastewater effluent. Water Sci. Technol. 2010, 61, 537–544. [Google Scholar] [CrossRef]
- Fongaro, G.; Padilha, J.; Schissi, C.D.; Nascimento, M.A.; Bampi, G.B.; Viancelli, A.; Barardi, C.R.M. Human and animal enteric virus in groundwater from deep wells, and recreational and network water. Environ. Sci. Pollut. Res. 2015, 22, 20060–20066. [Google Scholar] [CrossRef] [PubMed]
- Elmahdy, E.M.; Fongaro, G.; Schissi, C.D.; Petrucio, M.M.; Barardi, C. Enteric viruses in surface water and sediment samples from the catchment area of Peri Lagoon, Santa Catarina State, Brazil. J. Water Health 2015, 14, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Reid, T.M.S.; Robinson, H.G. Frozen raspberries and hepatitis A. Epidemiol. Infect. 1987, 98, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, L.S.; Mirkin, I.R.; Allen, D.T.; Safford, S.; Hadler, S.C. A multifocal outbreak of hepatitis A traced to commercially distributed lettuce. Am. J. Public Health 1990, 80, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.T.; Polish, L.B.; Robertson, B.H.; Khanna, B.K.; Woodruff, B.A.; Shapiro, C.N.; Miller, M.A.; Smith, J.D.; Gedrose, J.K.; Alter, M.J.; et al. Multistate Outbreak of Hepatitis A Associated with Frozen Strawberries. J. Infect. Dis. 1992, 166, 518–524. [Google Scholar] [CrossRef]
- Hutin, Y.J.; Pool, V.; Cramer, E.H.; Nainan, O.V.; Weth, J.; Williams, I.T.; Goldstein, S.T.; Gensheimer, K.F.; Bell, B.P.; Shapiro, C.N.; et al. A Multistate, Foodborne Outbreak of Hepatitis A. N. Engl. J. Med. 1999, 340, 595–602. [Google Scholar] [CrossRef]
- Dentinger, C.M.; Bower, W.A.; Nainan, O.V.; Cotter, S.M.; Myers, G.; Dubusky, L.M.; Fowler, S.; Salehi, E.D.P.; Bell, B.P. An Outbreak of Hepatitis A Associated with Green Onions. J. Infect. Dis. 2001, 183, 1273–1276. [Google Scholar] [CrossRef]
- Wheeler, C.; Vogt, T.M.; Armstrong, G.L.; Vaughan, G.; Weltman, A.; Nainan, O.V.; Dato, V.; Xia, G.; Waller, K.; Amon, J.; et al. An Outbreak of Hepatitis A Associated with Green Onions. N. Engl. J. Med. 2005, 353, 890–897. [Google Scholar] [CrossRef]
- Petrignani, M.W.F.; Harms, M.; Verhoef, L.; Van Hunen, R.; Swaan, C.; Van Steenbergen, J.; Boxman, I.; Sala, R.P.I.; Ober, H.; Vennema, H.; et al. Update: A food-borne outbreak of hepatitis A in the Netherlands related to semi-dried tomatoes in oil, January-February 2010. Eurosurveillance 2010, 15. [Google Scholar] [CrossRef]
- Severi, E.; Verhoef, L.; Thornton, L.; Herrador, B.R.G.; Faber, M.; Sundqvist, L.; Rimhanen-Finne, R.; Roque-Afonso, A.M.; Ngui, S.L.; Allerberger, F.; et al. Large and prolonged food-borne multistate hepatitis A outbreak in Europe associated with consumption of frozen berries, 2013 to 2014. Eurosurveillance 2015, 20. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Verhaelen, K.; Rzeżutka, A.; Kozyra, I.; Maunula, L.; Von Bonsdorff, C.-H.; Vantarakis, A.; Kokkinos, P.; Petrović, T.; Lazić, S.; et al. Quantitative farm-to-fork risk assessment model for norovirus and hepatitis A virus in European leafy green vegetable and berry fruit supply chains. Int. J. Food Microbiol. 2015, 198, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Terio, V.; Bottaro, M.; Di Pinto, A.; Catella, C.; Chironna, M.; Bozzo, G.; Kingsley, D.H.; Bonerba, E.; Morea, A.; Martella, V. Outbreak of Hepatitis A in Italy Associated with Frozen Redcurrants Imported from Poland: A Case Study. Food Environ. Virol. 2015, 7, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Sewlikar, S.; D’Souza, D.H. Survival of hepatitis A virus and Aichi virus in cranberry-based juices at refrigeration (4 °C). Food Microbiol. 2017, 62, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Croci, L.; Ciccozzi, M.; De Medici, D.; Di Pasquale, S.; Fiore, A.; Mele, A.; Toti, L. Inactivation of Hepatitis A virus in heat-treated mussels. J. Appl. Microbiol. 1999, 87, 884–888. [Google Scholar] [CrossRef]
- Croci, L.; De Medici, D.; Scalfaro, C.; Fiore, A.; Divizia, M.; Donia, D.; Cosentino, A.M.; Moretti, P.; Costantini, G. Determination of enterovirus, Hepatitis A virus; Bacteriophages and E.coli in Adriatic Sea mussels. J. Food Protect. 2000, 88, 293–296. [Google Scholar]
- Conaty, S.; Bird, P.; Bell, G.; Kraa, E.; Grohmann, G.; McAnulty, J.M. Hepatitis A in New South Wales, Australia, from consumption of oysters: The first reported outbreak. Epidemiol. Infect. 2000, 124, 121–130. [Google Scholar] [CrossRef]
- De Medici, D.; Croci, L.; Di Pasquale, S.; Fiore, A.; Toti, L. Detecting the presence of infectious hepatitis A virus in molluscs positive to RT-nested-PCR. Lett. Appl. Microbiol. 2001, 33, 362–366. [Google Scholar] [CrossRef]
- Koopmans, M.; von Bonsdorff, C.H.; Vinje, J.; de Medici, D.; Monroe, S. Foodborne viruses. FEMS Microbiol. Rev. 2002, 26, 187–205. [Google Scholar] [CrossRef]
- Sánchez, G.; Pintó, R.M.; Vanaclocha, H.; Bosch, A. Molecular Characterization of Hepatitis A Virus Isolates from a Transcontinental Shellfish-Borne Outbreak. J. Clin. Microbiol. 2002, 40, 4148–4155. [Google Scholar] [CrossRef]
- Di Pasquale, S.; Paniconi, M.; De Medici, D.; Suffredini, E.; Croci, L. Duplex Real Time PCR for the detection of hepatitis A virus in shellfish using Feline Calicivirus as a process control. J. Virol. Methods 2010, 163, 96–100. [Google Scholar] [CrossRef]
- Sincero, T.; Levin, D.; Simões, C.; Barardi, C.R.M. Detection of hepatitis A virus (HAV) in oysters (Crassostrea gigas). Water Res. 2006, 40, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Bazzardi, R.; Fattaccio, M.C.; Salza, S.; Canu, A.; Marongiu, E.; Pisanu, M. Preliminary study on Norovirus, hepatitis A virus, Escherichia coli and their potential seasonality in shellfish from different growing and harvesting areas in Sardinia region. Ital. J. Food Saf. 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Suffredini, E.; Lanni, L.; Arcangeli, G.; Pepe, T.; Mazzette, R.; Ciccaglioni, G.; Croci, L. Qualitative and quantitative assessment of viral contamination in bivalve molluscs harvested in Italy. Int. J. Food Microbiol. 2014, 184, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Di Bartolo, I.; Cioffi, B.; Ianiro, G.; Palermo, P.; Monini, M.; Amoroso, M.G. Prevalence of Foodborne Viruses in Mussels in Southern Italy. Food Environ. Virol. 2017, 9, 187–194. [Google Scholar] [CrossRef]
- Fusco, G.; Anastasio, A.; Kingsley, D.; Amoroso, M.G.; Pepe, T.; Fratamico, P.M.; Cioffi, B.; Rossi, R.; La Rosa, G.; Boccia, F. Detection of Hepatitis A Virus and Other Enteric Viruses in Shellfish Collected in the Gulf of Naples, Italy. Int. J. Environ. Res. Public Health 2019, 16, 2588. [Google Scholar] [CrossRef]
- Lees, D.N. Viruses and bivalve shellfish. Int. J. Food Microbiol. 2000, 59, 81–116. [Google Scholar] [CrossRef]
- De Donno, A.; Grassi, T.; Bagordo, F.; Idolo, A.; Serio, F.; Gabutti, G. Detection of Viruses in Coastal Seawater Using Mytilus Galloprovincialis as an Accumulation Matrix. Food Environ. Virol. 2012, 4, 81–88. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Conn, D. Molecular markers and sentinel organisms for environmental monitoring. Parasite 2008, 15, 458–462. [Google Scholar] [CrossRef]
- Lucy, F.E.; Graczyk, T.K.; Tamang, L.; Miraflor, A.; Minchin, D. Biomonitoring of surface and coastal water for Cryptosporidium, Giardia, and human-virulent microsporidia using molluscan shellfish. Parasitol. Res. 2008, 103, 1369–1375. [Google Scholar] [CrossRef]
- Lopalco, P.; Malfait, P.; Salmaso, S.; Germinario, C.A.; Quarto, M.; Barbuti, S.; Cipriani, R.; Mundo, A.; Pesole, G. A persisting outbreak of hepatitis A in Puglia, Italy, 1996: Epidemiological follow-up. Eurosurveillance 1997, 2, 31–32. [Google Scholar] [CrossRef]
- Pontrelli, G.; Boccia, D.; Di Renzi, M.; Massari, M.; Giugliano, F.; Celentano, L.P.; Taffon, S.; Genovese, D.; Di Pasquale, S.; Scalise, F.; et al. Epidemiological and virological characterization of a large community-wide outbreak of hepatitis A in southern Italy. Epidemiol. Infect. 2007, 136, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Ndumbi, P.; Freidl, G.; Williams, C.J.S.; Mårdh, O.; Varela, C.; Avellón, A.; Friesema, I.; Vannema, H.; Beebeejaun, K.; Ngui, S.L. Hepatitis A outbreak disproportionately afecting men who have sex with men (MSM) in the European Union and European Economic Area, June 2016 to May 2017. Eur. Surveill. 2018, 23, 1700641. [Google Scholar] [CrossRef] [PubMed]
- Aulicino, G.; Faccini, M.; Lamberti, A.; Senatori, S.; Ciconali, G.; Gandolfi, C.E.; Galli, C.; Tagliacarne, C.; Castaldi, S.; Romanò, L. Hepatitis A epidemic in men who have sex with men (MSM) in Milan, Italy. Acta Biomed. 2020, 91, 106–110. [Google Scholar]
- Bazzardi, R.; Dore, E.; Ciccozzi, M.; Lai, A.; Pisanu, M.; Babudieri, S.; Maida, I. Outbreak of acute hepatitis A associated with men who have sex with men (MSM) in North Sardinia 2017–2018. J. Infect. Dev. Ctries. 2020, 14, 1065–1070. [Google Scholar] [CrossRef]
- Minosse, C.; Messina, F.; Garbuglia, A.R.; Meschi, S.; Scognamiglio, P.; Capobianchi, M.R.; Ippolito, G.; Lanini, S. Origin of HAV strains responsible for 2016–2017 outbreak among MSM: Viral phylodynamics in Lazio region. PLoS ONE 2020, 15, e0234010. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.; Kasprzyk-Hordern, B. Future perspectives of wastewater-based epidemiology: Monitoring infectious disease spread and resistance to the community level. Environ. Int. 2020, 139. [Google Scholar] [CrossRef]
- Burge, C.A.; Closek, C.J.; Friedman, C.S.; Groner, M.L.; Jenkins, C.M.; Shore-Maggio, A.; Welsh, J.E. The Use of Filter-feeders to Manage Disease in a Changing World. Integr. Comp. Biol. 2016, 56, 573–587. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| PCR | Primer Name | Primer Sequence (5′–3′) | Length (bp) | PCR Round | Reference |
|---|---|---|---|---|---|
| Nested-PCR | 2897 | TATTCAGATTGCAAATTAYAAT | 393 | 1st round | [24] |
| 3288 | AAYTTCATYATTTCATGCTCCT | ||||
| 2949 | TATTTGTCTGTYACAGAACAATCAG | 267 | 2nd round | ||
| 3192 | AGGRGGTGGAAGYACTTCATTTGA | ||||
| RT-qPCR | HAV68 | TCACCGCCGTTTGCCTAG | - | - | [21] |
| HAV240 | GGAGAGCCCTGGAAGAAAG | ||||
| HAV150 | FAM-CCTGAACCTGCAGGAATTAA-MGBNFQ | ||||
| Mengo110 | GCGGGTCCTGCCGAAAGT | - | - | [25] | |
| Mengo209 | GAAGTAACATATAGACAGACGCACAC | ||||
| Mengo147 | FAM-ATCACATTACTGGCCGAAGC-MGBNFQ |
| Period | Type of Sample (Suffix) | N° of Samples | N° of Positive Samples | Genotype (N° of Sequences and Clusters) |
|---|---|---|---|---|
| 2015 | Sewage (WTP) | 30 | 4 | IA (3 cluster D)—IB (1) |
| Discharge Water (DW) | 29 | 4 | IA (3 cluster D)—IB (1) | |
| Seawater (SW) | 39 | 5 | IA (4 cluster D)—IB (1) | |
| Bivalve shellfish (BV) | 485 | 79 * | IA (2 cluster A; 7 cluster D)—IB (2) | |
| 2016–2018 | Sewage (WTP) | 41 | 16 | IA (7 cluster A; 8 cluster B)—IB (1) |
| Discharge Water (DW) | 324 | 10 | IA (3 cluster A; 3 cluster B)—IB (4) | |
| Bivalve shellfish (BV) | 261 | 23 * | IA (10 cluster A)—IB (1) | |
| Total | 1209 | 141 | IA (50) **—IB (11) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Rosa, G.; Mancini, P.; Bonanno Ferraro, G.; Iaconelli, M.; Veneri, C.; Paradiso, R.; De Medici, D.; Vicenza, T.; Proroga, Y.T.R.; Di Maro, O.; et al. Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance. Viruses 2021, 13, 16. https://doi.org/10.3390/v13010016
La Rosa G, Mancini P, Bonanno Ferraro G, Iaconelli M, Veneri C, Paradiso R, De Medici D, Vicenza T, Proroga YTR, Di Maro O, et al. Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance. Viruses. 2021; 13(1):16. https://doi.org/10.3390/v13010016
Chicago/Turabian StyleLa Rosa, Giuseppina, Pamela Mancini, Giusy Bonanno Ferraro, Marcello Iaconelli, Carolina Veneri, Rosa Paradiso, Dario De Medici, Teresa Vicenza, Yolande Therese Rose Proroga, Orlandina Di Maro, and et al. 2021. "Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance" Viruses 13, no. 1: 16. https://doi.org/10.3390/v13010016
APA StyleLa Rosa, G., Mancini, P., Bonanno Ferraro, G., Iaconelli, M., Veneri, C., Paradiso, R., De Medici, D., Vicenza, T., Proroga, Y. T. R., Di Maro, O., Ciccaglione, A. R., Bruni, R., Equestre, M., Taffon, S., Costantino, A., Della Rotonda, M., & Suffredini, E. (2021). Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance. Viruses, 13(1), 16. https://doi.org/10.3390/v13010016