Abstract
Hepatitis A virus (HAV) infection is one of the major causes of acute hepatitis, and this infection occasionally causes acute liver failure. HAV infection is associated with HAV-contaminated food and water as well as sexual transmission among men who have sex with men. Although an HAV vaccine has been developed, outbreaks of hepatitis A and life-threatening severe HAV infections are still observed worldwide. Therefore, an improved HAV vaccine and anti-HAV drugs for severe hepatitis A should be developed. Here, we reviewed cell culture systems for HAV infection, and other issues. This review may help with improving the HAV vaccine and developing anti-HAV drugs.
Keywords:
Anti-HAV; drug screening; HAV; Huh7; IRES; PLC/PRF/5; subgenomic replicon; 5′ UTR; vaccine 1. Introduction
Hepatitis A virus (HAV) infection is a health problem in developing and developed countries [1,2]. HAV is transmitted mainly through the fecal–oral route or close physical contact [3]. HAV infection is associated with HAV-contaminated food and water [4,5]. An increase in HAV infection has also occurred among men who have sex with men (MSM) [6,7]. HAV infection occasionally causes acute liver failure and leads to death in HAV-infected patients [8,9].
HAV is a single-stranded positive-stranded RNA virus that circulates in blood as a membrane-cloaked quasi-enveloped virus (eHAV), but is shed in the feces as a naked, non-enveloped virion during acute hepatitis A virus infection [10]. The HAV genome consists of almost 7500 nt [11] and has a single open reading frame (ORF) encoding a polyprotein: structural P1 (viral protein (VP) 4, VP2, VP3, and VP1) and nonstructural proteins P2 (2A, 2B, and 2C) and P3 (3A, 3B, 3C, and 3D). The ORF is preceded by the 5′ untranslated region (UTR) and followed by the 3′ UTR with a poly(A) tail [12,13].
The HAV internal ribosomal entry site (IRES) exists in the 5′ UTR, and HAV initiates the translation by a cap-independent mechanism through the HAV IRES [14,15,16,17]. A single HAV polyprotein is proteolytically processed by the HAV 3C protease and cellular protease(s) into mature proteins. HAV 3D is an RNA-dependent polymerase that is important for viral replication. Highly permissive cell lines are needed to screen antivirals [18,19]. The potential antiviral targets are also important to develop effective antivirals. In this review article, we first discuss the cell culture systems. We next review the drug targets for HAV infections.
2. Cell Culture Systems for HAV Replication
2.1. Cell Lines Permissive for HAV Replication
HAV is a fastidious virus that grows slowly due to a combination of several factors [20]. HAV is difficult to grow, and only a few combinations of HAV strains and cell lines are available. Thus, it has been difficult to develop effective and cheap vaccines and to study potential antivirals.
Efficient infectious cell culture systems of HAV are shown in Table 1. Provost et al. successfully propagated HAV in primary marmoset hepatocytes and the normal fetal rhesus kidney cell line (FRhK6) [21]. They established methods of immunofluorescence, immunofluorescence blockade, serum neutralization, immune adherence, radioimmunoassay, immune electron microscopy, and marmoset inoculation tests for HAV detection [21]. Frösner et al. isolated HAV directly from human feces and propagated the virus serially in the human hepatoma cell line Alexander (PLC/PRF/5) [22]. They confirmed by radioimmunoassay that hepatitis A antigen (HAAg) increased in the cell extracts obtained by the freezing and thawing of cells [22].

Table 1.
Cell lines supporting hepatitis A virus (HAV) replication.
Gauss-Müller et al. demonstrated that the antigen was located within the cytoplasm of HAV-infected human embryo fibroblasts by an immunofluorescence study [23]. Kojima et al. also propagated HAV in the conventional cell lines, FL and Vero cells [24]. They confirmed HAAg in cytoplasm by radioimmunoassay (RIA) and immune electron microscopy although they did not observe any cytopathic effects in the cell culture [24]. In primary African green monkey (Cercopithecus aethiops) kidney (AGMK) cell cultures, HAV strains were recovered from the stool specimen of a patient with HAV and confirmed by direct immunofluorescence [25].
Lemon et al. developed a new radioimmunofocus assay method, which retained many of the advantages of conventional plaque assays, for the quantitation of HAV using African green monkey kidney BS-C-1 cells [26]. Wheeler et al. reported that the HAS-15 strain, which was recovered from wild HAV, was adapted to rapid growth in FRhK-4 cells by more than 20 7-day passages, and confirmed HAV using a radioimmunoassay and virus-specific immunofluorescence [27].
Crance et al. reported that PLC/PRF/5 cells supported continuous production of the HAV CF53 strain, which was isolated from the stools of a patient with HAV infection 3 days after the onset of jaundice and was adapted to grow in PLC/PRF/5 cells [28]. Robertson et al. reported the growth and recovery of purified HAV from FRhK4 cells persistently infected with HAV isolate HAS-15 over a 2- to 3-month period [29]. Simian HAV strain AGM-27 and cell culture-adapted HM-175 grew in CV-1, FRhK-1, and primary AGMK cells [30].
Cohen et al. cloned the cDNA of a cell culture-adapted HAV (HAV HM-175/7 MK-5) full-length genome into pBR322 [31]. They transfected the cells using an infectious transcript from HAV cDNA into AGMK and CV-1 cells and inoculated with transfection-derived virus into marmosets and observed the appearance of anti-HAV antibodies and hepatitis [31]. Emerson et al. performed transfection of HM-175 (wild type and cell-culture-adapted) and an infectivity assay in FRhK-4 and AGMK cells and revealed that the attenuation of virulence may also require multiple mutations [32,33]. Morace et al. revealed that genome mutations of HAV 3A regions were associated with two cytopathic HAV strains [34]. The HAV strains FG and SI0 were isolated from the feces of a young patient from Southern Italy collected four days before the onset of clinical symptoms and from a water sample taken from the Tiber River (central Italy), respectively [34,35]. These sequences were compared to those of three cytopathic clones of HM175 described by Lemon et al. [36]. A study reported that HM175-cytopathogenic strains have mutations in both the 5′ and 3′ UTRs and in the nonstructural proteins 2A, 2B, 2C, 3A, and 3Dpol, which may be associated with the cytopathic phenotype [36]. Sequence analysis revealed cell culture-adapted HAV mutations [37,38,39,40].
The cynomolgus monkey renal cell line JTC-12.P3 could support HAV replication [41]. The pig cell line IB-RS-2 cells also supported HAV replication [42]. Mouse liver cells were coded for functional HAV receptors and other factors required for efficient HAV replication in cell culture [43].
Primary human hepatocyte PXB cells, which were derived from a humanized severe combined immunodeficiency albumin promoter/enhancer-driven urokinase-type plasminogen activator mouse model, could support HAV replication [44]. Cell culture-grown HAV strains and HAV derived from fecal extracts were used for the infection of these cell lines. Similar to the cell culture-grown HCV JFH1 strain [45,46,47], it may be easier to infect human hepatocytes with cell culture-grown HAV strains. Cytopathic effects are not always observed when these HAV strains infect human cell lines. Perez-Rodriguez et al. recently reported that quasispecies genomic selection and molecular breeding using deep sequencing identified high-fitness individuals improving HAV [48].
2.2. Cell Culture for HAV Vaccine Development
Inactivated vaccines are licensed and commonly used to control HAV infections [3]. HAV strain CR326, adapted to grow in Macaca mulatta kidney LLC-MK2 cells, was highly purified, inactivated with formalin, adsorbed to alum, and tested for the capacity to induce anti-HAV antibodies in both mice and marmosets [53]. As LLC-MK2 cells are unacceptable to prepare human vaccines, the HAV strain CR326 can also be prepared in a similar manner in Medical Research Council cell strain 5 (MRC-5) cells, which are diploid human cell lines composed of fibroblasts and acceptable for the manufacturing of human vaccines [53].
Flehming et al. performed HAV propagation and adaptation in human embryo kidney cells (HKC) [54]. They also demonstrated that HAV from the 10th passage through HKC can replicate in a human embryo fibroblast strain (HFS) [54]. They also developed the HAV strain HFS/GBM, which can be propagated in human fibroblast cells in quantities sufficient for producing inactivated vaccines [55]. These fibroblasts were derived from the lungs of a normal 25-week embryo.
As an alternative strain for a vaccine strain, a fast-growing strain of HAV with a great potential for HAV antigen production has been isolated by quasispecies genomic selection and molecular breeding [48]. As the production of vaccines is expensive [12], a fast-growing HAV strain may be useful for making the production costs of HAV vaccines lower.
2.3. Cell Culture for the Development of Anti-HAV Drugs
Despite the use of an effective vaccine, antivirals against HAV would be of great use [12]. The effective anti-HAV drugs and cell culture systems in which they were found are shown in Table 2. We previously reviewed other anti-HAV drugs (see the reference [13]). The human hepatoma cell lines PLC/PRF/5 and Huh7 are often used for the discovery of anti-HAV drugs, although HAV has various strains. HAV had high IRES activities in HLE and Huh7 cells [16]. Although excellent HAV vaccines exist, further development of therapeutic options to prevent severe hepatitis A is needed. In Japan, due to the legal difficulties associated with cadaveric donation that existed ~20 years ago, the number of liver transplantations is still lower than in other countries. Therefore, anti-HAV drugs must be developed. As most of the investigations did not go beyond tests on cell cultures, it would be useful and important to improve HAV cell culture systems.
Table 2.
Effective drugs inhibiting hepatitis A virus (HAV) replication discovered in cell culture systems for HAV.
4. Blocking the Entry Pathway as an Antiviral Strategy
Kaplan et al. reported that HAV cellular receptor 1 (HAVcr-1) is an attachment receptor for HAV as well as a functional receptor for HAV infection in African green monkey kidney (AGMK) GL37 cells [71]. Liver, as well as kidneys and colorectal tissues, have a high expression of HAVcr-1 mRNA [72]. HAVcr-1 is also known as kidney injury molecule-1 (KIM-1), T cell immunoglobulin (Ig), and mucin domain containing 1 (TIM-1). Major sequence variants of TIM-1 were completely co-segregated with T cells and the airway phenotype regulator (Tapr), which is associated with asthma susceptibility genes [73]. HAVcr-1 may also have an inverse relationship between HAV infection and atopy development [73]. Immunotherapies may regulate HAVcr-1 function and downmodulate allergic inflammatory diseases [74].
However, there are important discussions on the actual role of HAVcr-1 as a cell surface receptor for HAV [75,76,77]. Costafreda and Kaplan (2018) reported that AGMK HAVcr-1 knockout cells lost susceptibility to HAV infection, including HAV-free viral particles and exosomes purified from HAV-infected cells [76]. Naked HAV virions were responsible for fecal–oral transmission of HAV between individuals and eHAV mediated the spread within the newly infected host [10,75].
In contrast, in other studies, the HAVcr-1 was identified as a non-essential entry factor for either naked HAV or eHAV [77,78]. Integrin β1 binds elsewhere on the HAV capsid. Trafficking of eHAV to the lysosome is essential for the entry and uncoating of HAV genome and requires the endosomal sorting complexes required for transport (ESCRT) and programmed cell death 6-interacting protein/apoptosis-linked gene 2 (ALG2)-interacting protein X (PDCD6IP/ALIX) in addition to RAB5A, the member RAS oncogene family (Rab5), and RAB7, the member RAS oncogene family (Rab7) GTPase [75]. The present proposed life cycle model of HAV is shown in Figure 1. Although the mechanism of endocytosis has been developed in detail, the involvement of HAV and cell surface receptors for HAV is not clearly established yet. Drugs inhibiting the steps of entry process could be efficient antivirals.
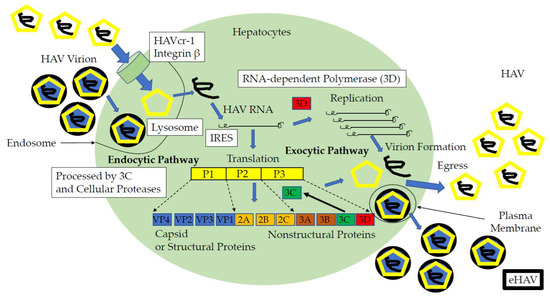
Figure 1.
Life cycle of the hepatitis A virus and candidates of anti-HAV drug targets (open squares). HAVcr-1, hepatitis A virus cellular receptor 1; IRES, internal ribosomal entry site; HAV, naked HAV; eHAV, quasi-enveloped HAV; bold black circle, envelope; yellow pentagon, viral capsid [10,13,71,72,73,74,75,76,77,78,79,80,81,82].
5. Inhibiting the HAV IRES-Mediated Translation in Human Hepatoma Cell Lines
HAV translation is initiated in a cap-independent style by a mechanism involving the binding of the 40S RNA subunit at a portion located hundreds of bases downstream of the 5′ end of the HAV RNA genome, which has been termed “HAV IRES”. HAV IRES spans a region from ~nt.161 to the first initiator codon AUG, located at ~nt. 734, and encompasses most of the 5′ UTR of HAV RNA [79]. We previously reported that some siRNAs targeting the HAV IRES suppressed the genome translation and replication [14].
As the nucleotide sequence homology of the 5′ end of the HAV RNA genome is well-preserved among different HAV strains, HAV IRES may be used as a universal and effective target for anti-HAV drugs. HAV IRES is highly ineffective [83,84]; however, mutants with higher activity have been described [20,48], which may be useful for the study of antivirals. Additionally, HAV IRES is much more active in the FRhK-4 cells than in the Huh7 cells. However, the Huh7 cells represent a better model of the natural infection and a closer model for testing antivirals.
As HAV had higher IRES activities in Huh7 cells [16], researchers typically used Huh7 to examine HAV IRES activity with transient transfection of a bicistronic reporter construct containing HAV IRES as an intragenic spacer (pSV40 HAV IRES). After 48–72 h of transfection and treatment with or without specific drugs, the activities of HAV IRES were measured using a luciferase assay. Studies demonstrated that amantadine inhibited HAV IRES-mediated translation in human hepatoma cells [15,16,17]. Researchers also used the stable cell line, COS-7 HAV IRES, and found that the JAK2 inhibitor AZD1480 inhibited HAV replication [63]. HAV IRES was an attractive target for anti-HAV drugs and could be useful for screening effective anti-HAV drugs (Figure 1).
6. HAV 3C Protease and 3D Polymerase May Be other Candidates for Anti-HAV Drug Targets
In the treatment of chronic hepatitis C infection, HCV NS3/4A protease inhibitors and HCV NS5B nucleotide inhibitors now play major roles in achieving a sustained virological response (SVR) [85,86]. HAV 3C is a cysteine protease inhibitor and processes HAV proteins [13]. HAV 3C protease can impair the induction of β interferon through the cleavage of the NF-κB essential modulator (NEMO) and plays an immune role in the evasion mechanism of HAV [87]. HAV 3C also induced caspase-independent cell death with the vacuolization of lysosomal and endosomal organelles [88]. There are several reports on the development of HAV 3C inhibitors [13,81].
HAV 3D is an RNA-dependent RNA polymerase and plays a role in the progression of acute hepatitis A. Amino acid changes in HAV 3D could modulate the growth rate of HAV [82]. HAV 3D was also involved in the disruption of TLR3 signaling, an important pathway for HAV eradication [89]. Although there are a few reports regarding HAV 3D polymerase inhibitors, HAV 3D could be a potential target for the treatment of hepatitis A.
7. Conclusions
In the present article, we described the relationship between anti-HAV drugs and cell culture models for HAV infection, HAV subgenomic replicons, and HAV IRES reporter assays. Creating better and cheaper HAV vaccines and developing anti-HAV drugs against severe hepatitis A remains important and further exploration is needed.
Author Contributions
Data curation, T.K.; writing—original draft preparation, T.K.; writing—review and editing, R.S., R.M., N.M., and M.O.; supervision, M.M.; funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Japan Agency for Medical Research and Development (AMED) under Grant number JP20fk0210075.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gupta, E.; Ballani, N.; Kumar, M.; Sarin, S.K. Role of non-hepatotropic viruses in acute sporadic viral hepatitis and acute-on-chronic liver failure in adults. Indian J. Gastroenterol. 2015, 34, 448–452. [Google Scholar] [CrossRef]
- Tominaga, A.; Kanda, T.; Akiike, T.; Komoda, H.; Ito, K.; Abe, A.; Aruga, A.; Kaneda, S.; Saito, M.; Kiyohara, T.; et al. Hepatitis A outbreak associated with a revolving sushi bar in Chiba, Japan: Application of molecular epidemiology. Hepatol. Res. 2012, 42, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, P.; Brett, C.; Batool, A.; Sapra, A. Hepatitis A Vaccine; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554604/ (accessed on 11 May 2020).
- Tsukada, R.; Ono, S.; Kobayashi, H.; Wada, Y.; Nishizawa, K.; Fujii, M.; Takeuchi, M.; Kuroiwa, K.; Kobayashi, Y.; Ishii, K.; et al. A Cluster of Hepatitis A Infections Presumed to be Related to Asari Clams and Investigation of the Spread of Viral Contamination from Asari Clams. Jpn. J. Infect. Dis. 2019, 72, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Shiota, T.; Yoshizaki, S.; Saito-Obata, M.; Malbas, F.F., Jr.; Lupisan, S.P.; Oshitani, H.; Takeda, N.; Muramatsu, M.; Wakita, T.; et al. Detection of Subgenotype IA and IIIA Hepatitis A Viruses in Rivers Flowing through Metro Manila, the Philippines. Jpn. J. Infect. Dis. 2019, 72, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, N.; Owada, T.; Tanaka, A.; Matsubayashi, K.; Nagai, T.; Satake, M. Hepatitis A virus and hepatitis E virus prevalence relates to Human Immunodeficiency Virus infection in Japanese male blood donors. Microbiol. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.A.; Hofmeister, M.G.; Kupronis, B.A.; Lin, Y.; Xia, G.L.; Yin, S.; Teshale, E. Increase in Hepatitis A Virus Infections-United States, 2013–2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Miyamura, T.; Ishii, K.; Kanda, T.; Tawada, A.; Sekimoto, T.; Wu, S.; Nakamoto, S.; Arai, M.; Fujiwara, K.; Imazeki, F.; et al. Possible widespread presence of hepatitis A virus subgenotype IIIA in Japan: Recent trend of hepatitis A causing acute liver failure. Hepatol. Res. 2012, 42, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Maki, Y.; Kimizuka, Y.; Sasaki, H.; Yamamoto, T.; Hamakawa, Y.; Tagami, Y.; Miyata, J.; Hayashi, N.; Fujikura, Y.; Kawana, A. Hepatitis A virus-associated fulminant hepatitis with human immunodeficiency virus coinfection. J. Infect. Chemother. 2020, 26, 282–285. [Google Scholar] [CrossRef]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef]
- Cohen, J.I.; Rosenblum, B.; Ticehurst, J.R.; Daemer, R.J.; Feinstone, S.M.; Purcell, R.H. Complete nucleotide sequence of an attenuated hepatitis A virus: Comparison with wild-type virus. Proc. Natl. Acad. Sci. USA 1987, 84, 2497–2501. [Google Scholar] [CrossRef]
- Debing, Y.; Neyts, J.; Thibaut, H.J. Molecular biology and inhibitors of hepatitis A virus. Med. Res. Rev. 2014, 34, 895–917. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Nakamoto, S.; Wu, S.; Nakamura, M.; Jiang, X.; Haga, Y.; Sasaki, R.; Yokosuka, O. Direct-acting Antivirals and Host-targeting Agents against the Hepatitis A Virus. J. Clin. Transl. Hepatol. 2015, 3, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Zhang, B.; Kusov, Y.; Yokosuka, O.; Gauss-Müller, V. Suppression of hepatitis A virus genome translation and replication by siRNAs targeting the internal ribosomal entry site. Biochem. Biophys. Res. Commun. 2005, 330, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Yokosuka, O.; Imazeki, F.; Fujiwara, K.; Nagao, K.; Saisho, H. Amantadine inhibits hepatitis A virus internal ribosomal entry site-mediated translation in human hepatoma cells. Biochem. Biophys. Res. Commun. 2005, 331, 621–629. [Google Scholar] [CrossRef]
- Kanda, T.; Imazeki, F.; Nakamoto, S.; Okitsu, K.; Fujiwara, K.; Yokosuka, O. Internal ribosomal entry-site activities of clinical isolate-derived hepatitis A virus and inhibitory effects of amantadine. Hepatol. Res. 2010, 40, 415–423. [Google Scholar] [CrossRef]
- Yang, L.; Kiyohara, T.; Kanda, T.; Imazeki, F.; Fujiwara, K.; Gauss-Müller, V.; Ishii, K.; Wakita, T.; Yokosuka, O. Inhibitory effects on HAV IRES-mediated translation and replication by a combination of amantadine and interferon-alpha. Virol. J. 2010, 7, 212. [Google Scholar] [CrossRef]
- Cao, J.; Isaacson, J.; Patick, A.K.; Blair, W.S. High-throughput human immunodeficiency virus type 1 (HIV-1) full replication assay that includes HIV-1 Vif as an antiviral target. Antimicrob. Agents Chemother. 2005, 49, 3833–3841. [Google Scholar] [CrossRef]
- Verrier, E.R.; Colpitts, C.C.; Schuster, C.; Zeisel, M.B.; Baumert, T.F. Cell Culture Models for the Investigation of Hepatitis B and D Virus Infection. Viruses 2016, 8, 261. [Google Scholar] [CrossRef]
- Pintó, R.M.; Pérez-Rodríguez, F.J.; D’Andrea, L.; de Castellarnau, M.; Guix, S.; Bosch, A. Hepatitis A Virus Codon Usage: Implications for Translation Kinetics and Capsid Folding. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef]
- Provost, P.J.; Hilleman, M.R. Propagation of human hepatitis A virus in cell culture in vitro. Proc. Soc. Exp. Biol. Med. 1979, 160, 213–221. [Google Scholar] [CrossRef]
- Frösner, G.G.; Deinhardt, F.; Scheid, R.; Gauss-Müller, V.; Holmes, N.; Messelberger, V.; Siegl, G.; Alexander, J.J. Propagation of human hepatitis A virus in a hepatoma cell line. Infection 1979, 7, 303–305. [Google Scholar] [CrossRef]
- Gauss-Müller, V.; Frösner, G.G.; Deinhardt, F. Propagation of hepatitis A virus in human embryo fibroblasts. J. Med. Virol. 1981, 7, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Shibayama, T.; Sato, A.; Suzuki, S.; Ichida, F.; Hamada, C. Propagation of human hepatitis A virus in conventional cell lines. J. Med. Virol. 1981, 7, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Daemer, R.J.; Feinstone, S.M.; Gust, I.D.; Purcell, R.H. Propagation of human hepatitis A virus in African green monkey kidney cell culture: Primary isolation and serial passage. Infect. Immun. 1981, 32, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Lemon, S.M.; Binn, L.N.; Marchwicki, R.H. Radioimmunofocus assay for quantitation of hepatitis A virus in cell cultures. J. Clin. Microbiol. 1983, 17, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, C.M.; Fields, H.A.; Schable, C.A.; Meinke, W.J.; Maynard, J.E. Adsorption, purification, and growth characteristics of hepatitis A virus strain HAS-15 propagated in fetal rhesus monkey kidney cells. J. Clin. Microbiol. 1986, 23, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Crance, J.M.; Passagot, J.; Biziagos, E.; Deloince, R. Continuous production of hepatitis A virus in PLC/PRF/5 cell cultures: Use of antigen for serology. J. Virol. Methods 1987, 18, 193–203. [Google Scholar] [CrossRef]
- Robertson, B.H.; Khanna, B.; Brown, V.K.; Margolis, H.S. Large scale production of hepatitis A virus in cell culture: Effect of type of infection on virus yield and cell integrity. J. Gen. Virol. 1988, 69, 2129–2134. [Google Scholar] [CrossRef]
- Tsarev, S.A.; Emerson, S.U.; Balayan, M.S.; Ticehurst, J.; Purcell, R.H. Simian hepatitis A virus (HAV) strain AGM-27: Comparison of genome structure and growth in cell culture with other HAV strains. J. Gen. Virol. 1991, 72, 1677–1683. [Google Scholar] [CrossRef]
- Cohen, J.I.; Ticehurst, J.R.; Feinstone, S.M.; Rosenblum, B.; Purcell, R.H. Hepatitis A virus cDNA and its RNA transcripts are infectious in cell culture. J. Virol. 1987, 61, 3035–3039. [Google Scholar] [CrossRef]
- Emerson, S.U.; Huang, Y.K.; McRill, C.; Lewis, M.; Shapiro, M.; London, W.T.; Purcell, R.H. Molecular basis of virulence and growth of hepatitis A virus in cell culture. Vaccine 1992, 10 (Suppl. 1), S36–S39. [Google Scholar] [CrossRef]
- Emerson, S.U.; Huang, Y.K.; McRill, C.; Lewis, M.; Purcell, R.H. Mutations in both the 2B and 2C genes of hepatitis A virus are involved in adaptation to growth in cell culture. J. Virol. 1992, 66, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Morace, G.; Pisani, G.; Beneduce, F.; Divizia, M.; Panà, A. Mutations in the 3A genomic region of two cytopathic strains of hepatitis A virus isolated in Italy. Virus Res. 1993, 28187–28194. [Google Scholar] [CrossRef]
- Venuti, A.; Di Russo, C.; del Grosso, N.; Patti, A.M.; Ruggeri, F.; De Stasio, P.R.; Martiniello, M.G.; Pagnotti, P.; Degener, A.M.; Midulla, M.; et al. Isolation and molecular cloning of a fast-growing strain of human hepatitis A virus from its double-stranded replicative form. J. Virol. 1985, 56, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Lemon, S.M.; Murphy, P.C.; Shields, P.A.; Ping, L.H.; Feinstone, S.M.; Cromeans, T.; Jansen, R.W. Antigenic and genetic variation in cytopathic hepatitis A virus variants arising during persistent infection: Evidence for genetic recombination. J. Virol. 1991, 65, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.; Normann, A.; Feinstone, S.M.; Flehmig, B. Nucleotide sequence of wild-type hepatitis A virus GBM in comparison with two cell culture-adapted variants. J. Virol. 1994, 68, 548–554. [Google Scholar] [CrossRef]
- Graff, J.; Kasang, C.; Normann, A.; Pfisterer-Hunt, M.; Feinstone, S.M.; Flehmig, B. Mutational events in consecutive passages of hepatitis A virus strain GBM during cell culture adaptation. Virology 1994, 204, 60–68. [Google Scholar] [CrossRef]
- Zhang, H.; Chao, S.F.; Ping, L.H.; Grace, K.; Clarke, B.; Lemon, S.M. An infectious cDNA clone of a cytopathic hepatitis A virus: Genomic regions associated with rapid replication and cytopathic effect. Virology 1995, 212, 686–697. [Google Scholar] [CrossRef][Green Version]
- Funkhouser, A.W.; Raychaudhuri, G.; Purcell, R.H.; Govindarajan, S.; Elkins, R.; Emerson, S.U. Progress toward the development of a genetically engineered attenuated hepatitis A virus vaccine. J. Virol. 1996, 70, 7948–7957. [Google Scholar] [CrossRef]
- Baba, M.; Takegawa, M.; Kaito, M.; Miyamoto, K.; Suzuki, S. Propagation of hepatitis A virus in a renal cell line JTC-12.P3 of cynomolgus monkey origin. Acta Virol. 1993, 37, 209–222. [Google Scholar]
- Dotzauer, A.; Feinstone, S.M.; Kaplan, G. Susceptibility of nonprimate cell lines to hepatitis A virus infection. J. Virol. 1994, 68, 6064–6068. [Google Scholar] [CrossRef] [PubMed]
- Feigelstock, D.A.; Thompson, P.; Kaplan, G.G. Growth of hepatitis A virus in a mouse liver cell line. J. Virol. 2005, 79, 2950–2955. [Google Scholar] [CrossRef][Green Version]
- Win, N.N.; Kanda, T.; Nakamoto, S.; Moriyama, M.; Jiang, X.; Suganami, A.; Tamura, Y.; Okamoto, H.; Shirasawa, H. Inhibitory effect of Japanese rice-koji miso extracts on hepatitis A virus replication in association with the elevation of glucose-regulated protein 78 expression. Int. J. Med. Sci. 2018, 15, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Wakita, T.; Pietschmann, T.; Kato, T.; Date, T.; Miyamoto, M.; Zhao, Z.; Murthy, K.; Habermann, A.; Kräusslich, H.G.; Mizokami, M.; et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat. Med. 2005, 11, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Gastaminza, P.; Cheng, G.; Kapadia, S.; Kato, T.; Burton, D.R.; Wieland, S.F.; Uprichard, S.L.; Wakita, T.; Chisari, F.V. Robust hepatitis C virus infection in vitro. Proc. Natl. Acad. Sci. USA 2005, 102, 9294–9299. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Basu, A.; Steele, R.; Wakita, T.; Ryerse, J.S.; Ray, R.; Ray, R.B. Generation of infectious hepatitis C virus in immortalized human hepatocytes. J. Virol. 2006, 80, 4633–4639. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, F.J.; D’Andrea, L.; de Castellarnau, M.; Costafreda, M.I.; Guix, S.; Ribes, E.; Quer, J.; Gregori, J.; Bosch, A.; Pintó, R.M. Improving virus production through quasispecies genomic selection and molecular breeding. Sci. Rep. 2016, 6, 35962. [Google Scholar] [CrossRef]
- Konduru, K.; Kaplan, G.G. Stable growth of wild-type hepatitis A virus in cell culture. J. Virol. 2006, 80, 1352–1360. [Google Scholar] [CrossRef][Green Version]
- Kusov, Y.; Kanda, T.; Palmenberg, A.; Sgro, J.Y.; Gauss-Müller, V. Silencing of hepatitis A virus infection by small interfering RNAs. J. Virol. 2006, 80, 5599–5610. [Google Scholar] [CrossRef]
- Jiang, X.; Kanda, T.; Wu, S.; Nakamoto, S.; Saito, K.; Shirasawa, H.; Kiyohara, T.; Ishii, K.; Wakita, T.; Okamoto, H.; et al. Suppression of La antigen exerts potential antiviral effects against hepatitis A virus. PLoS ONE 2014, 9, e101993. [Google Scholar] [CrossRef]
- Hirai-Yuki, A.; Hensley, L.; Whitmire, J.K.; Lemon, S.M. Biliary Secretion of Quasi-Enveloped Human Hepatitis A Virus. mBio 2016, 7, e01998-16. [Google Scholar] [CrossRef] [PubMed]
- Provost, P.J.; Hughes, J.V.; Miller, W.J.; Giesa, P.A.; Banker, F.S.; Emini, E.A. An inactivated hepatitis A viral vaccine of cell culture origin. J. Med. Virol. 1986, 19, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Flehmig, B.; Vallbracht, A.; Wurster, G. Hepatitis A virus in cell culture. III. Propagation of hepatitis A virus in human embryo kidney cells and human embryo fibroblast strains. Med. Microbiol. Immunol. 1981, 170, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Flehmig, B.; Heinricy, U.; Pfisterer, M. Immunogenicity of a killed hepatitis A vaccine in seronegative volunteers. Lancet 1989, 1, 1039–1041. [Google Scholar] [CrossRef]
- Widell, A.; Hansson, B.G.; Oberg, B.; Nordenfelt, E. Influence of twenty potentially antiviral substances on in vitro multiplication of hepatitis A virus. Antivir. Res. 1986, 6, 103–112. [Google Scholar] [CrossRef]
- Biziagos, E.; Crance, J.M.; Passagot, J.; Deloince, R. Effect of antiviral substances on hepatitis A virus replication in vitro. J. Med. Virol. 1987, 22, 57–66. [Google Scholar] [CrossRef]
- Biziagos, E.; Crance, J.M.; Passagot, J.; Deloince, R. Inhibitory effects of atropine, protamine, and their combination on hepatitis A virus replication in PLC/PRF/5 cells. Antimicrob. Agents Chemother. 1990, 34, 1112–1117. [Google Scholar] [CrossRef]
- Crance, J.M.; Biziagos, E.; Passagot, J.; van Cuyck-Gandré, H.; Deloince, R. Inhibition of hepatitis A virus replication in vitro by antiviral compounds. J. Med. Virol. 1990, 31, 155–160. [Google Scholar] [CrossRef]
- Girond, S.; Crance, J.M.; Van Cuyck-Gandre, H.; Renaudet, J.; Deloince, R. Antiviral activity of carrageenan on hepatitis A virus replication in cell culture. Res. Virol. 1991, 142, 261–270. [Google Scholar] [CrossRef]
- Crance, J.M.; Lévêque, F.; Chousterman, S.; Jouan, A.; Trépo, C.; Deloince, R. Antiviral activity of recombinant interferon-alpha on hepatitis A virus replication in human liver cells. Antivir. Res. 1995, 28, 69–80. [Google Scholar] [CrossRef]
- Kanda, T.; Wu, S.; Kiyohara, T.; Nakamoto, S.; Jiang, X.; Miyamura, T.; Imazeki, F.; Ishii, K.; Wakita, T.; Yokosuka, O. Interleukin-29 suppresses hepatitis A and C viral internal ribosomal entry site-mediated translation. Viral Immunol. 2012, 25, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Kanda, T.; Nakamoto, S.; Saito, K.; Nakamura, M.; Wu, S.; Haga, Y.; Sasaki, R.; Sakamoto, N.; Shirasawa, H.; et al. The JAK2 inhibitor AZD1480 inhibits hepatitis A virus replication in Huh7 cells. Biochem. Biophys. Res. Commun. 2015, 458, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Sasaki, R.; Nakamoto, S.; Haga, Y.; Nakamura, M.; Shirasawa, H.; Okamoto, H.; Yokosuka, O. The sirtuin inhibitor sirtinol inhibits hepatitis A virus (HAV) replication by inhibiting HAV internal ribosomal entry site activity. Biochem. Biophys. Res. Commun. 2015, 466, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kanda, T.; Suganami, A.; Nakamoto, S.; Win, N.N.; Tamura, Y.; Nakamura, M.; Matsuoka, S.; Yokosuka, O.; Kato, N.; et al. Antiviral activity of zinc sulfate against hepatitis A virus replication. Future Virol. 2019, 14, 399–406. [Google Scholar] [CrossRef]
- Yi, M.; Lemon, S.M. Replication of subgenomic hepatitis A virus RNAs expressing firefly luciferase is enhanced by mutations associated with adaptation of virus to growth in cultured cells. J. Virol. 2002, 76, 1171–1180. [Google Scholar] [CrossRef]
- Gauss-Müller, V.; Kusov, Y.Y. Replication of a hepatitis A virus replicon detected by genetic recombination in vivo. J. Gen. Virol. 2002, 83, 2183–2192. [Google Scholar] [CrossRef]
- Kusov, Y.Y.; Shatirishvili, G.; Klinger, M.; Gauss-Müller, V. A vaccinia virus MVA-T7-mediated recovery of infectious hepatitis A virus from full-size cDNA or from two cDNAs, both by themselves unable to complete the virus life cycle. Virus Res. 2002, 89, 75–88. [Google Scholar] [CrossRef]
- Yang, Y.; Yi, M.; Simonds, P.; Lemon, S.M. Identification of a conserved RNA replication element (cre) within the 3Dpol-coding sequence of hepatoviruses. J. Virol. 2008, 82, 10118–10128. [Google Scholar] [CrossRef]
- Kanda, T.; Kusov, Y.; Yokosuka, O.; Gauss-Müller, V. Interference of hepatitis A virus replication by small interfering RNAs. Biochem. Biophys. Res. Commun. 2004, 318, 341–345. [Google Scholar] [CrossRef]
- Kaplan, G.; Totsuka, A.; Thompson, P.; Akatsuka, T.; Moritsugu, Y.; Feinstone, S.M. Identification of a surface glycoprotein on African green monkey kidney cells as a receptor for hepatitis A virus. EMBO J. 1996, 15, 4282–4296. [Google Scholar] [CrossRef]
- BioGPS. HAVCR1 (Hepatitis A Virus Cellular Receptor 1), Gene Expression/Activity Chart. Available online: http://biogps.org/#goto=genereport&id=26762 (accessed on 26 March 2020).
- McIntire, J.J.; Umetsu, S.E.; Akbari, O.; Potter, M.; Kuchroo, V.K.; Barsh, G.S.; Freeman, G.J.; Umetsu, D.T.; DeKruyff, R.H. Identification of Tapr (an airway hyperreactivity regulatory locus) and the linked Tim gene family. Nat. Immunol. 2001, 2, 1109–1116. [Google Scholar] [CrossRef]
- Umetsu, S.E.; Lee, W.L.; McIntire, J.J.; Downey, L.; Sanjanwala, B.; Akbari, O.; Berry, G.J.; Nagumo, H.; Freeman, G.J.; Umetsu, D.T.; et al. TIM-1 induces T cell activation and inhibits the development of peripheral tolerance. Nat. Immunol. 2005, 6, 447–454. [Google Scholar] [CrossRef]
- Rivera-Serrano, E.E.; González-López, O.; Das, A.; Lemon, S.M. Cellular entry and uncoating of naked and quasi-enveloped human hepatoviruses. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Costafreda, M.I.; Kaplan, G. HAVCR1 (CD365) and Its Mouse Ortholog Are Functional Hepatitis A Virus (HAV) Cellular Receptors That Mediate HAV Infection. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Maury, W.; Lemon, S.M. TIM1 (HAVCR1): An Essential “Receptor” or an “Accessory Attachment Factor” for Hepatitis A Virus? J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Das, A.; Hirai-Yuki, A.; González-López, O.; Rhein, B.; Moller-Tank, S.; Brouillette, R.; Hensley, L.; Misumi, I.; Lovell, W.; Cullen, J.M.; et al. TIM1 (HAVCR1) Is Not Essential for Cellular Entry of Either Quasi-enveloped or Naked Hepatitis A Virions. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.A.; Zajac, A.J.; Lemon, S.M. In vitro characterization of an internal ribosomal entry site (IRES) present within the 5’ nontranslated region of hepatitis A virus RNA: Comparison with the IRES of encephalomyocarditis virus. J. Virol. 1994, 68, 1066–1074. [Google Scholar] [CrossRef]
- Kassem, A.F.; Batran, R.Z.; Abbas, E.M.H.; Elseginy, S.A.; Shaheen, M.N.F.; Elmahdy, E.M. New 4-phenylcoumarin derivatives as potent 3C protease inhibitors: Design, synthesis, anti-HAV effect and molecular modeling. Eur. J. Med. Chem. 2019, 168, 447–460. [Google Scholar] [CrossRef]
- Banerjee, K.; Bhat, R.; Rao, V.U.B.; Nain, A.; Rallapalli, K.L.; Gangopadhyay, S.; Singh, R.P.; Banerjee, M.; Jayaram, B. Toward development of generic inhibitors against the 3C proteases of picornaviruses. FEBS J. 2019, 286, 765–787. [Google Scholar] [CrossRef]
- Konduru, K.; Kaplan, G.G. Determinants in 3Dpol modulate the rate of growth of hepatitis A virus. J. Virol. 2010, 84, 8342–8347. [Google Scholar] [CrossRef]
- Brown, E.A.; Day, S.P.; Jansen, R.W.; Lemon, S.M. The 5′ nontranslated region of hepatitis A virus RNA: Secondary structure and elements required for translation in vitro. J. Virol. 1991, 65, 5828–5838. [Google Scholar] [CrossRef] [PubMed]
- Whetter, L.E.; Day, S.P.; Elroy-Stein, O.; Brown, E.A.; Lemon, S.M. Low efficiency of the 5′ nontranslated region of hepatitis A virus RNA in directing cap-independent translation in permissive monkey kidney cells. J. Virol. 1994, 68, 5253–5263. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Lau, G.K.K.; Wei, L.; Moriyama, M.; Yu, M.L.; Chuang, W.L.; Ibrahim, A.; Lesmana, C.R.A.; Sollano, J.; Kumar, M.; et al. APASL clinical practice recommendation: How to treat HCV-infected patients with renal impairment? Hepatol. Int. 2019, 13, 103–109. [Google Scholar] [CrossRef]
- AASLD-IDSA HCV Guidance Panel. Hepatitis C Guidance 2018 Update: AASLD-IDSA Recommendations for Testing, Managing, and Treating Hepatitis C Virus Infection. Clin. Infect. Dis. 2018, 67, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fang, L.; Wei, D.; Zhang, H.; Luo, R.; Chen, H.; Li, K.; Xiao, S. Hepatitis A virus 3C protease cleaves NEMO to impair induction of beta interferon. J. Virol. 2014, 88, 10252–10258. [Google Scholar] [CrossRef]
- Shubin, A.V.; Demidyuk, I.V.; Lunina, N.A.; Komissarov, A.A.; Roschina, M.P.; Leonova, O.G.; Kostrov, S.V. Protease 3C of hepatitis A virus induces vacuolization of lysosomal/endosomal organelles and caspase-independent cell death. BMC Cell Biol. 2015, 16, 4. [Google Scholar] [CrossRef]
- Qu, L.; Feng, Z.; Yamane, D.; Liang, Y.; Lanford, R.E.; Li, K.; Lemon, S.M. Disruption of TLR3 signaling due to cleavage of TRIF by the hepatitis A virus protease-polymerase processing intermediate, 3CD. PLoS Pathog. 2011, 7, e1002169. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).