

Functional Traits Associated with Drought Tolerance Exhibit Low Variability in 21 Provenances of a Montane Tree Species—Eucalyptus delegatensis

 , ,
, ,  and
and 
Abstract
1. Introduction
- Is there variability in the expression of functional traits among provenances of alpine ash sourced across a climatic gradient?
- Do the morphological and physiological traits correlate with each other?
- Is there any relationship between the expression of functional traits and climate of origin?
2. Materials and Methods
2.1. Study Site
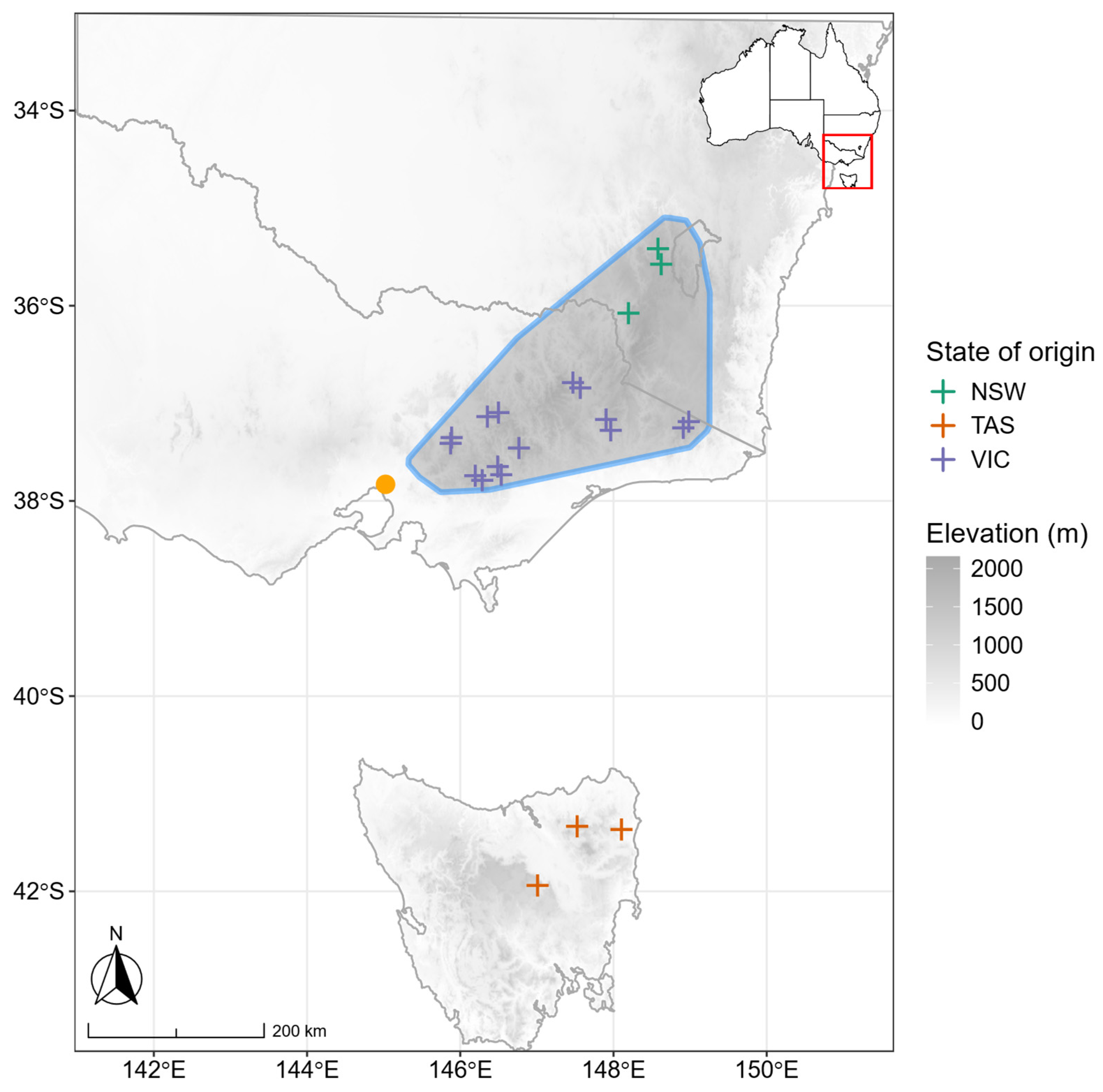
2.2. Plant Material
2.3. Climatic and Topographic Variables Investigated
2.4. Planting Design
2.5. Morphological Trait Measurements
2.5.1. Specific Leaf Area (SLA)
2.5.2. Stomatal Density (SD)
2.6. Physiological Trait Measurements
2.6.1. Osmotic Potential at Full Turgor (π(100))
2.6.2. Minimum Stomatal Conductance (gmin)
2.7. Statistical Analyses
3. Results
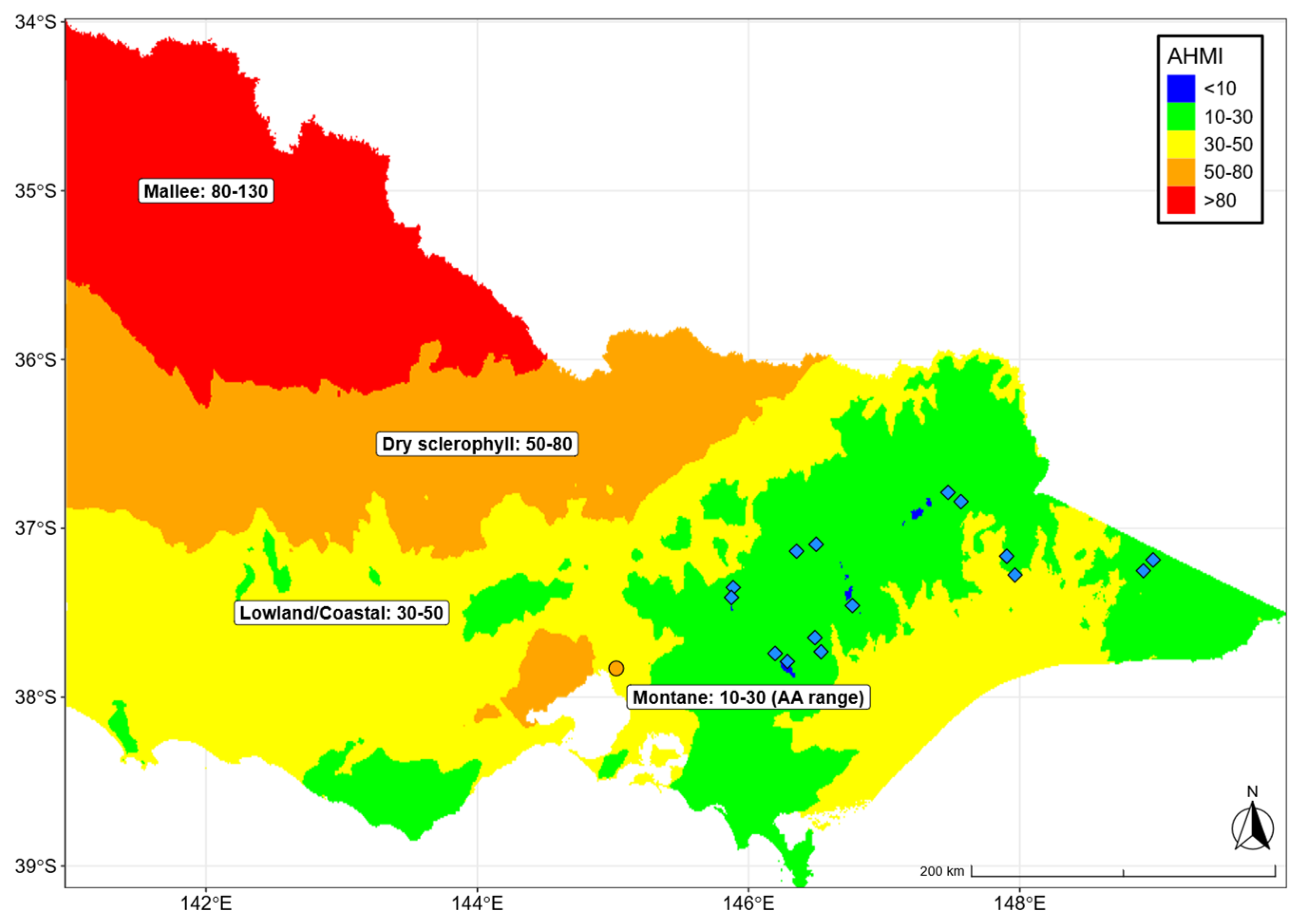
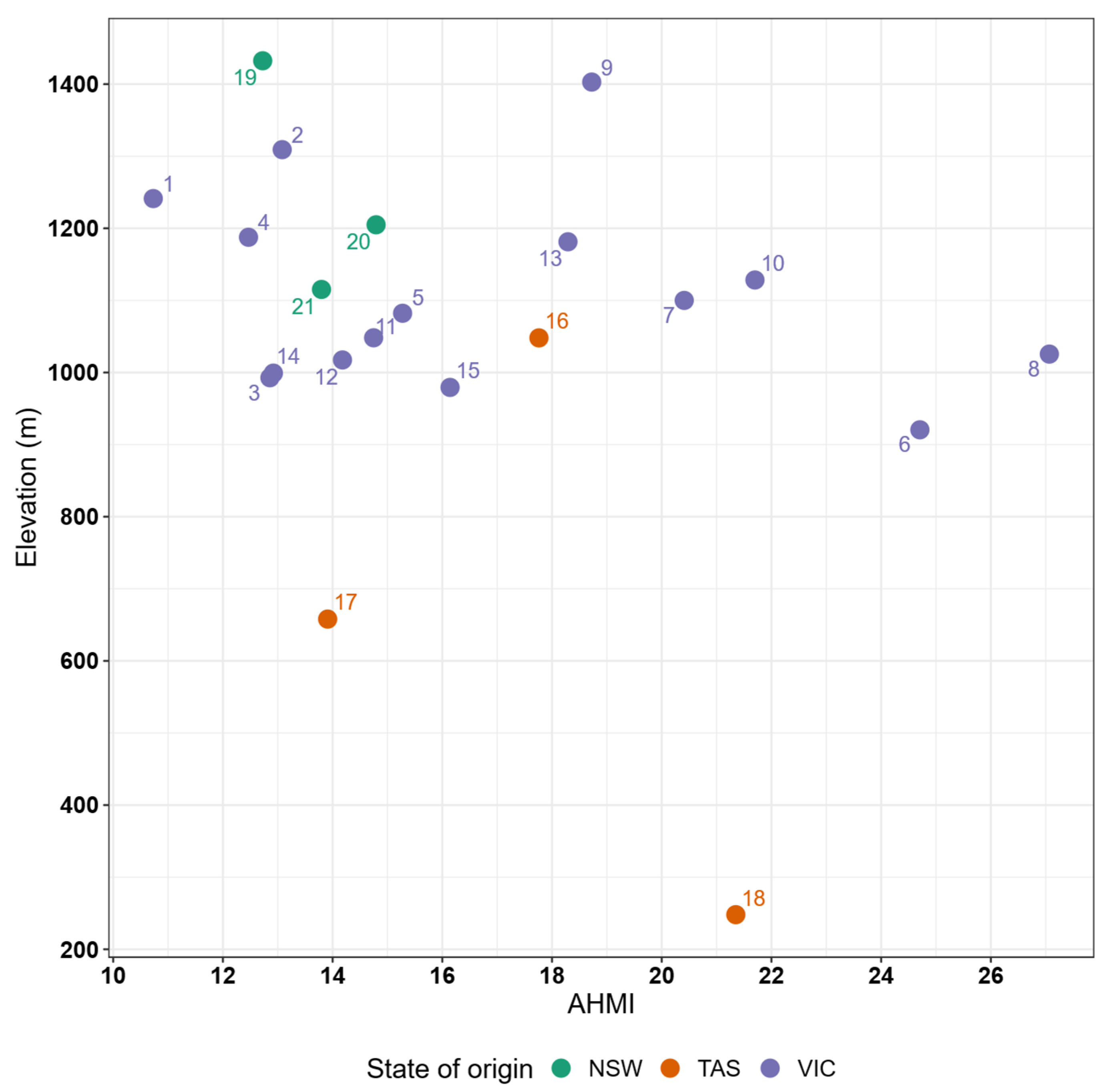
3.1. Climate of Origin
3.2. Variation in Morpho-Physiological Traits
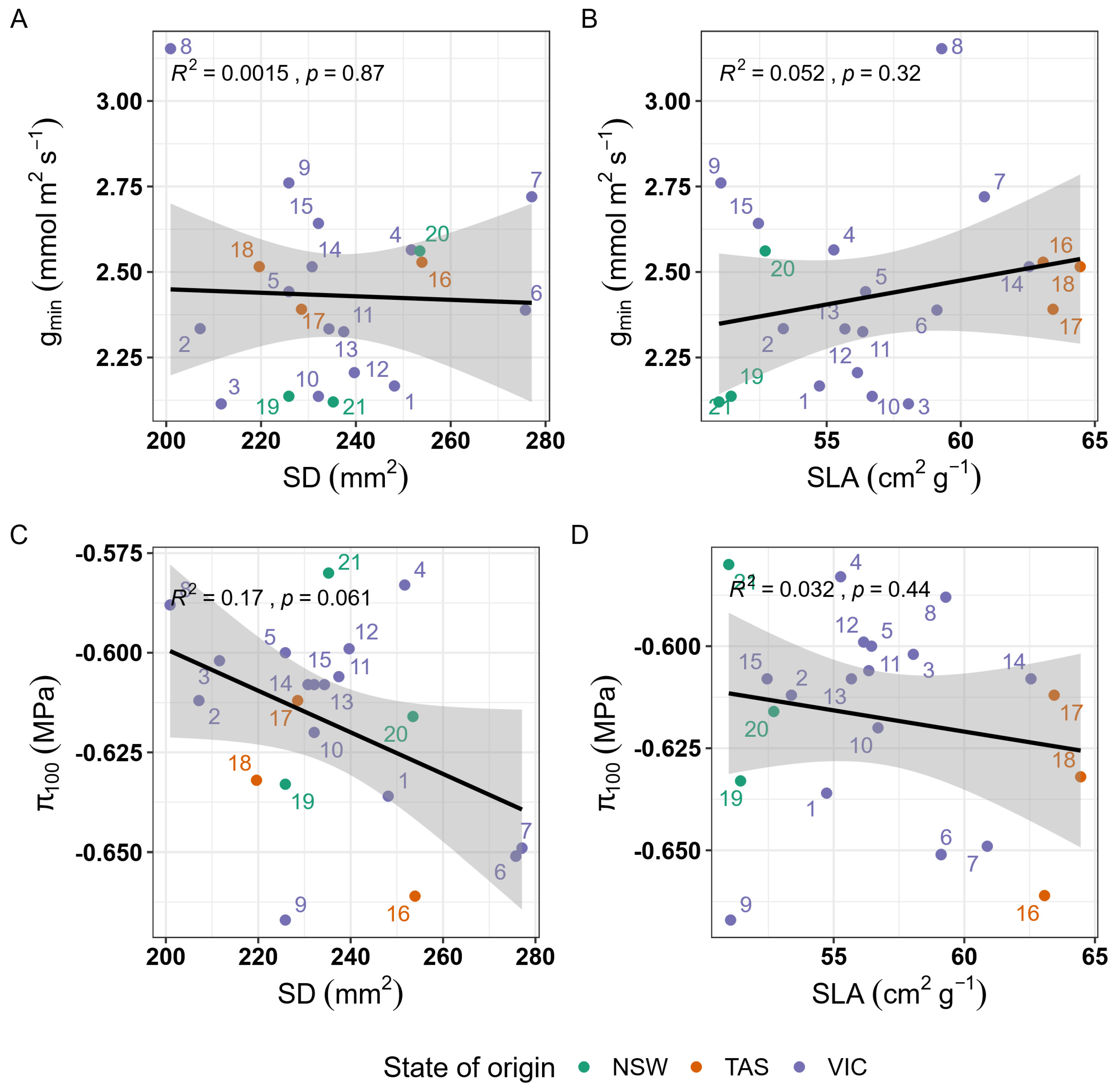
3.3. Correlation Among Morpho-Physiological Traits
3.4. Relationship Between Traits and Climate of Origin
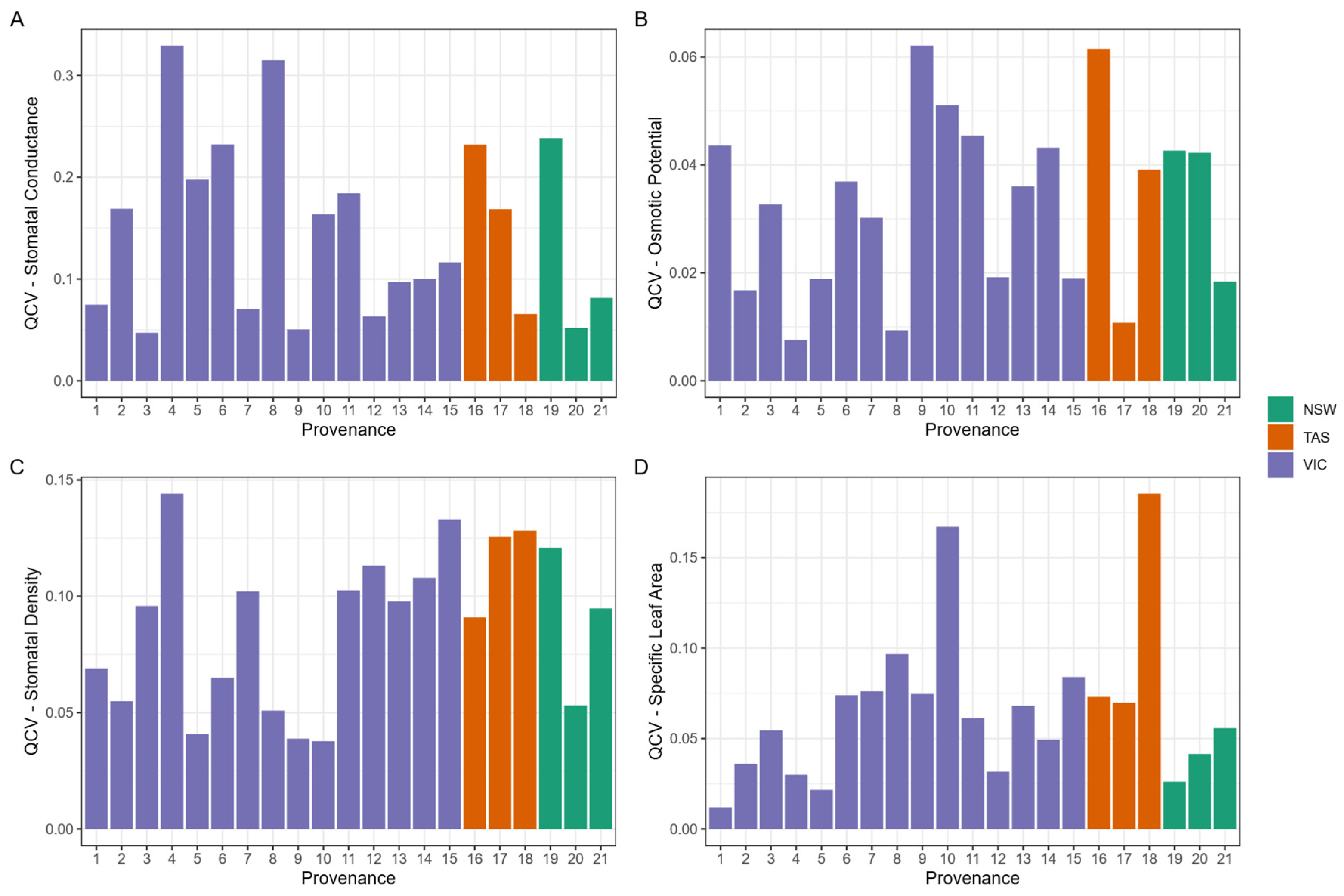
3.5. Intraspecific Trait Variation
4. Discussion
4.1. Variability of Functional Traits Across the Climate Gradient
4.2. Why Was the Variation of Functional Traits So Small Among Provenances?
4.3. Genetic vs. Environmental Influences of Trait Expression
4.4. Intraspecific Trait Variation
4.5. Management Implications
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Figures

Appendix A.2. Tables

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | State | x-Coordinate | y-Coordinate | AHMI | Elevation (m) | MAT (°C) | MAP (mm) |
|---|---|---|---|---|---|---|---|
| 1 | VIC | 146.287 | −37.789 | 10.731 | 1241.250 | 8.083 | 1847 |
| 2 | VIC | 146.766 | −37.458 | 13.081 | 1309.080 | 7.042 | 1573 |
| 3 | VIC | 146.196 | −37.741 | 12.857 | 992.747 | 9.217 | 1796 |
| 4 | VIC | 146.489 | −37.648 | 12.467 | 1187.594 | 8.025 | 1690 |
| 5 | VIC | 146.535 | −37.731 | 15.276 | 1082.156 | 8.750 | 1618 |
| 6 | VIC | 148.983 | −37.187 | 24.706 | 920.500 | 10.100 | 813 |
| 7 | VIC | 148.911 | −37.252 | 20.410 | 1099.987 | 8.817 | 982 |
| 8 | VIC | 147.964 | −37.277 | 27.066 | 1025.535 | 9.275 | 974 |
| 9 | VIC | 147.903 | −37.165 | 18.724 | 1402.855 | 7.933 | 1168 |
| 10 | VIC | 147.566 | −36.842 | 21.700 | 1128.332 | 8.633 | 1218 |
| 11 | VIC | 145.887 | −37.351 | 14.748 | 1048.137 | 9.692 | 1735 |
| 12 | VIC | 146.498 | −37.095 | 14.180 | 1017.419 | 8.033 | 1505 |
| 13 | VIC | 147.446 | −36.793 | 18.291 | 1181.303 | 8.892 | 1245 |
| 14 | VIC | 145.874 | −37.410 | 12.921 | 999.127 | 9.833 | 1761 |
| 15 | VIC | 146.354 | −37.136 | 16.141 | 979.337 | 9.783 | 1379 |
| 16 | TAS | 147.009 | −41.940 | 17.762 | 1048.000 | 6.625 | 936 |
| 17 | TAS | 147.526 | −41.334 | 13.910 | 658.000 | 8.708 | 1345 |
| 18 | TAS | 148.104 | −41.367 | 21.351 | 248.000 | 11.842 | 1023 |
| 19 | NSW | 148.623 | −35.574 | 12.726 | 1432.359 | 7.942 | 1404 |
| 20 | NSW | 148.581 | −35.416 | 14.793 | 1204.887 | 10.058 | 1260 |
| 21 | NSW | 148.195 | −36.076 | 13.798 | 1115.211 | 10.625 | 1170 |
References
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Change 2012, 3, 30–36. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.; Comita, L.S.; Condit, R.; Kursar, T.A.; Tyree, M.T.; Turner, B.L.; Hubbell, S.P. Drought sensitivity shapes species distribution patterns in tropical forests. Nature 2007, 447, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fule, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef]
- Anderson, N.S.; Fontaine, J.B.; Lewandrowski, W.; Walden, L.; Ruthrof, K.X. Drought and Wildfire Legacies Highlight Vulnerability of a Mediterranean Climate-Type Forest. Austral Ecol. 2025, 50, e70011. [Google Scholar] [CrossRef]
- Ahrens, C.W.; Andrew, M.E.; Mazanec, R.A.; Ruthrof, K.X.; Challis, A.; Hardy, G.; Byrne, M.; Tissue, D.T.; Rymer, P.D. Plant functional traits differ in adaptability and are predicted to be differentially affected by climate change. Ecol. Evol. 2020, 10, 232–248. [Google Scholar] [CrossRef]
- Heilmeier, H. Functional traits explaining plant responses to past and future climate changes. Flora 2019, 254, 1–11. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Pritzkow, C.; Williamson, V.; Szota, C.; Trouve, R.; Arndt, S.K. Phenotypic plasticity and genetic adaptation of functional traits influences intra-specific variation in hydraulic efficiency and safety. Tree Physiol. 2020, 40, 215–229. [Google Scholar] [CrossRef]
- Warren, C.R.; Tausz, M.; Adams, M.A. Does rainfall explain variation in leaf morphology and physiology among populations of red ironbark (Eucalyptus sideroxylon subsp. tricarpa) grown in a common garden? Tree Physiol. 2005, 25, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of Stomatal Density and Morphology on Water-Use Efficiency in a Changing World. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef]
- James, A.T.; Lawn, R.J.; Cooper, M. Genotypic variation for drought stress response traits in soybean. I. Variation in soybean and wild Glycine spp. for epidermal conductance, osmotic potential, and relative water content. Aust. J. Agric. Res. 2008, 59, 656–669. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Li, X.; Blackman, C.J.; Choat, B.; Duursma, R.A.; Rymer, P.D.; Medlyn, B.E.; Tissue, D.T. Tree hydraulic traits are coordinated and strongly linked to climate-of-origin across a rainfall gradient. Plant Cell Environ. 2018, 41, 646–660. [Google Scholar] [CrossRef]
- Pfautsch, S.; Harbusch, M.; Wesolowski, A.; Smith, R.; Macfarlane, C.; Tjoelker, M.G.; Reich, P.B.; Adams, M.A. Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol. Lett. 2016, 19, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Trueba, S.; Pouteau, R.; Lens, F.; Feild, T.S.; Isnard, S.; Olson, M.E.; Delzon, S. Vulnerability to xylem embolism as a major correlate of the environmental distribution of rain forest species on a tropical island. Plant Cell Environ. 2017, 40, 277–289. [Google Scholar] [CrossRef]
- Kibria, M.G.; Tjoelker, M.G.; Marchin, R.M.; Arndt, S.K.; Rymer, P.D. Can species climate niche predict canopy growth, functional traits and phenotypic plasticity in urban trees? Urban For. Urban Green. 2024, 98, 128417. [Google Scholar] [CrossRef]
- Warren, C.R.; Dreyer, E.; Tausz, M.; Adams, M.A. Ecotype adaptation and acclimation of leaf traits to rainfall in 29 species of 16-year-old Eucalyptus at two common gardens. Funct. Ecol. 2006, 20, 929–940. [Google Scholar] [CrossRef]
- Umana, M.N.; Swenson, N.G. Intraspecific variation in traits and tree growth along an elevational gradient in a subtropical forest. Oecologia 2019, 191, 153–164. [Google Scholar] [CrossRef]
- Li, X.; Blackman, C.J.; Choat, B.; Rymer, P.D.; Medlyn, B.E.; Tissue, D.T. Drought tolerance traits do not vary across sites differing in water availability in Banksia serrata (Proteaceae). Funct. Plant Biol. 2019, 46, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Lobo, A.; Torres-Ruiz, J.M.; Burlett, R.; Lemaire, C.; Parise, C.; Francioni, C.; Truffaut, L.; Tomaskova, I.; Hansen, J.K.; Kjaer, E.D.; et al. Assessing inter- and intraspecific variability of xylem vulnerability to embolism in oaks. For. Ecol. Manag. 2018, 424, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Maherali, H.; DeLucia, E.H. Xylem conductivity and vulnerability to cavitation of ponderosa pine growing in contrasting climates. Tree Physiol. 2000, 20, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.D.; Lavorel, S.; Colloff, M.J.; Williams, K.J.; Williams, R.J. Moving from autonomous to planned adaptation in the montane forests of southeastern Australia under changing fire regimes. Austral Ecol. 2017, 42, 309–316. [Google Scholar] [CrossRef]
- McLean, E.H.; Prober, S.M.; Stock, W.D.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E.; Byrne, M. Plasticity of functional traits varies clinally along a rainfall gradient in Eucalyptus tricarpa. Plant Cell Environ. 2014, 37, 1440–1451. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Xu, G.Q.; Farrell, C.; Arndt, S.K. Climate of origin has no influence on drought adaptive traits and the drought responses of a widely distributed polymorphic shrub. Tree Physiol. 2022, 42, 86–98. [Google Scholar] [CrossRef]
- Ramirez-Valiente, J.A.; Cavender-Bares, J. Evolutionary trade-offs between drought resistance mechanisms across a precipitation gradient in a seasonally dry tropical oak (Quercus oleoides). Tree Physiol. 2017, 37, 889–901. [Google Scholar] [CrossRef]
- Walters, M.B.; Gerlach, J.P. Intraspecific growth and functional leaf trait responses to natural soil resource gradients for conifer species with contrasting leaf habit. Tree Physiol. 2013, 33, 297–310. [Google Scholar] [CrossRef]
- Booth, T.H.; Broadhurst, L.M.; Pinkard, E.; Prober, S.M.; Dillon, S.K.; Bush, D.; Pinyopusarerk, K.; Doran, J.C.; Ivkovich, M.; Young, A.G. Native forests and climate change: Lessons from eucalypts. For. Ecol. Manag. 2015, 347, 18–29. [Google Scholar] [CrossRef]
- Hughes, L.; Cawsey, E.M.; Westoby, M. Climatic Range Sizes of Eucalyptus Species in Relation to Future Climate Change. Glob. Ecol. Biogeogr. Lett. 1996, 5, 23–29. [Google Scholar] [CrossRef]
- Westoby, M.; Andrew, S.C.; Gallagher, R.V.; Schrader, J. Species gain and loss per degree Celsius. Oikos 2024, 2024, e10556. [Google Scholar] [CrossRef]
- Bowman, D.M.; Murphy, B.P.; Neyland, D.L.; Williamson, G.J.; Prior, L.D. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Change Biol. 2014, 20, 1008–1015. [Google Scholar] [CrossRef]
- McColl-Gausden, S.C.; Bennett, L.T.; Ababei, D.A.; Clarke, H.G.; Penman, T.D. Future fire regimes increase risks to obligate-seeder forests. Divers. Distrib. 2021, 17, 542–558. [Google Scholar] [CrossRef]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- Singh, A.; Baker, P.J.; Kasel, S.; Trouvé, R.; Stewart, S.B.; Nitschke, C.R. The role of climatic variability on Eucalyptus regeneration in southeastern Australia. Glob. Ecol. Conserv. 2021, 32, e01929. [Google Scholar] [CrossRef]
- Mok, H.F.; Arndt, S.K.; Nitschke, C.R. Modelling the potential impact of climate variability and change on species regeneration potential in the temperate forests of South-Eastern Australia. Global Change Biol. 2012, 18, 1053–1072. [Google Scholar] [CrossRef]
- Tng, D.Y.P.; Williamson, G.J.; Jordan, G.J.; Bowman, D. Giant eucalypts—Globally unique fire-adapted rain-forest trees? New Phytol. 2012, 196, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.; Clarke, H.; Clarke, M.F.; McColl Gausden, S.C.; Nolan, R.H.; Penman, T.; Bradstock, R.; Varner, M. Warmer and drier conditions have increased the potential for large and severe fire seasons across south-eastern Australia. Global Ecol. Biogeogr. 2022, 31, 1933–1948. [Google Scholar] [CrossRef]
- Bassett, O.D.; Prior, L.D.; Slijkerman, C.M.; Jamieson, D.; Bowman, D.M.J.S. Aerial sowing stopped the loss of alpine ash (Eucalyptus delegatensis) forests burnt by three short-interval fires in the Alpine National Park, Victoria, Australia. For. Ecol. Manag. 2015, 342, 39–48. [Google Scholar] [CrossRef]
- Mátyás, C. Climatic adaptation of trees: Rediscovering provenance tests. Euphytica 1996, 92, 45–54. [Google Scholar] [CrossRef]
- de Villemereuil, P.; Gaggiotti, O.E.; Mouterde, M.; Till-Bottraud, I. Common garden experiments in the genomic era: New perspectives and opportunities. Heredity 2016, 116, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.B.; Nitschke, C.R. Improving temperature interpolation using MODIS LST and local topography: A comparison of methods in south east Australia. Int. J. Climatol. 2017, 37, 3098–3110. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Paudel, S.K.; Waeber, P.O.; Simard, S.W.; Innes, J.L.; Nitschke, C.R. Multiple factors influence plant richness and diversity in the cold and dry boreal forest of southwest Yukon, Canada. Plant Ecol. 2016, 217, 505–519. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Yanchuk, A.; O’Neill, G.A.; Aitken, S.N. Use of response functions in selecting lodgepole pine populations for future climates. Global Change Biol. 2006, 12, 2404–2416. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L. The shuttle radar topography mission. Rev. Geophys. 2007, 45, 1–33. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Camargo, M.A.B.; Marenco, R.A. Density, size and distribution of stomata in 35 rainforest tree species in Central Amazonia. Acta Amaz. 2011, 41, 205–212. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Leitch, I.J.; Patel, S.; Pendharkar, A.; Knight, C.A. Genome size is a strong predictor of cell size and stomatal density in angiosperms. New Phytol. 2008, 179, 975–986. [Google Scholar] [CrossRef]
- Callister, A.N.; Arndt, S.K.; Adams, M.A. Comparison of four methods for measuring osmotic potential of tree leaves. Physiol. Plant. 2006, 127, 383–392. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Ehleringer, J.R.; Mooney, H.A.; Rundel, P.W. Plant Physiological Ecology: Field Methods and Instrumentation; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Sack, L.; Cowan, P.D.; Jaikumar, N.; Holbrook, N.M. The ‘hydrology’ of leaves: Co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef]
- Schreiber, L.; Skrabs, M.; Hartmann, K.D.; Diamantopoulos, P.; Simanova, E.; Santrucek, J. Effect of humidity on cuticular water permeability of isolated cuticular membranes and leaf disks. Planta 2001, 214, 274–282. [Google Scholar] [CrossRef]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Botta-Dukat, Z. Quartile coefficient of variation is more robust than CV for traits calculated as a ratio. Sci. Rep. 2023, 13, 4671. [Google Scholar] [CrossRef] [PubMed]
- Duursma, R.A.; Blackman, C.J.; Lopez, R.; Martin-StPaul, N.K.; Cochard, H.; Medlyn, B.E. On the minimum leaf conductance: Its role in models of plant water use, and ecological and environmental controls. New Phytol. 2019, 221, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sack, L.; Li, Y.; Zhang, J.; Yu, K.; Zhang, Q.; He, N.; Yu, G. Relationships of stomatal morphology to the environment across plant communities. Nat. Commun. 2023, 14, 6629. [Google Scholar] [CrossRef]
- Portelli, A.M.; Windecker, S.M.; Pollock, L.J.; Neal, W.C.; Morris, W.K.; Khot, R.; Vesk, P.A.; Williams, D. From mallees to mountain ash, specific leaf area is coordinated with eucalypt tree stature, resprouting, stem construction, and fruit size. Aust. J. Bot. 2023, 71, 506–522. [Google Scholar] [CrossRef]
- Givnish, T.J.; Wong, S.C.; Stuart-Williams, H.; Holloway-Phillips, M.; Farquhar, G.D. Determinants of maximum tree height inEucalyptusspecies along a rainfall gradient in Victoria, Australia. Ecology 2014, 95, 2991–3007. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Ardy, R.; Zhang, Y.; Sun, S.; Cao, K.; Sack, L. Rapid determination of comparative drought tolerance traits: Using an osmometer to predict turgor loss point. Methods Ecol. Evol. 2012, 3, 880–888. [Google Scholar] [CrossRef]
- Salvi, A.M.; Smith, D.D.; Adams, M.A.; McCulloh, K.A.; Givnish, T.J. Mesophyll photosynthetic sensitivity to leaf water potential in Eucalyptus: A new dimension of plant adaptation to native moisture supply. New Phytol. 2021, 230, 1844–1855. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Lusk, C.H.; Bellingham, P.J.; Burslem, D.; Simpson, A.H.; Kramer-Walter, K.R. Intraspecific trait variation can weaken interspecific trait correlations when assessing the whole-plant economic spectrum. Ecol. Evol. 2017, 7, 8936–8949. [Google Scholar] [CrossRef]
- Koerner, C.; Bannister, P.; Mark, A.F. Altitudinal variation in stomatal conductance, nitrogen content and leaf anatomy in different plant life forms in New Zealand. Oecologia 1986, 69, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zheng, L.; Qi, D. Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes. Ecol. Evol. 2020, 10, 8166–8175. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, L.D.L.; Loy, X.; Markham, I.P.; Elmer, C.M.; Hovenden, M.J.; HilleRisLambers, J.; Mayfield, M.M. Aridity drives coordinated trait shifts but not decreased trait variance across the geographic range of eight Australian trees. New Phytol. 2021, 229, 1375–1387. [Google Scholar] [CrossRef]
- Bourne, A.E.; Creek, D.; Peters, J.M.R.; Ellsworth, D.S.; Choat, B. Species climate range influences hydraulic and stomatal traits in Eucalyptus species. Ann. Bot. 2017, 120, 123–133. [Google Scholar] [CrossRef]
- Peters, J.M.R.; Lopez, R.; Nolf, M.; Hutley, L.B.; Wardlaw, T.; Cernusak, L.A.; Choat, B. Living on the edge: A continental-scale assessment of forest vulnerability to drought. Glob. Change Biol. 2021, 27, 3620–3641. [Google Scholar] [CrossRef]
- Sanders, G.J.; Arndt, S.K. Osmotic Adjustment Under Drought Conditions. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 199–229. [Google Scholar]
- Matheny, A.M.; Bohrer, G.; Garrity, S.R.; Morin, T.H.; Howard, C.J.; Vogel, C.S. Observations of stem water storage in trees of opposing hydraulic strategies. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Larter, M.; Pfautsch, S.; Domec, J.C.; Trueba, S.; Nagalingum, N.; Delzon, S. Aridity drove the evolution of extreme embolism resistance and the radiation of conifer genus Callitris. New Phytol. 2017, 215, 97–112. [Google Scholar] [CrossRef]
- Ellis, T.W.; Hatton, T.J. Relating leaf area index of natural eucalypt vegetation to climate variables in southern Australia. Agric. Water Manag. 2008, 95, 743–747. [Google Scholar] [CrossRef]
- Blackman, C.J.; Halliwell, B.; Brodribb, T.J. All together now: A mixed-planting experiment reveals adaptive drought tolerance in seedlings of 10 Eucalyptus species. Plant Physiol. 2024, 197, kiae632. [Google Scholar] [CrossRef]
- Smith, D.D.; Adams, M.A.; Salvi, A.M.; Krieg, C.P.; Ane, C.; McCulloh, K.A.; Givnish, T.J. Ecophysiological adaptations shape distributions of closely related trees along a climatic moisture gradient. Nat. Commun. 2023, 14, 7173. [Google Scholar] [CrossRef] [PubMed]
- Fontes, C.G.; Pinto-Ledezma, J.; Jacobsen, A.L.; Pratt, R.B.; Cavender-Bares, J. Adaptive variation among oaks in wood anatomical properties is shaped by climate of origin and shows limited plasticity across environments. Funct. Ecol. 2021, 36, 326–340. [Google Scholar] [CrossRef]
- Russo, S.E.; Kitajima, K. The Ecophysiology of Leaf Lifespan in Tropical Forests: Adaptive and Plastic Responses to Environmental Heterogeneity. In Tropical Tree Physiology; Tree Physiology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 357–383. [Google Scholar]
- Aspinwall, M.J.; Blackman, C.J.; Maier, C.; Tjoelker, M.G.; Rymer, P.D.; Creek, D.; Chieppa, J.; Griffin-Nolan, R.J.; Tissue, D.T. Aridity drives clinal patterns in leaf traits and responsiveness to precipitation in a broadly distributed Australian tree species. Plant Environ. Interact. 2023, 4, 70–85. [Google Scholar] [CrossRef]
- Breed, M.F.; Stead, M.G.; Ottewell, K.M.; Gardner, M.G.; Lowe, A.J. Which provenance and where? Seed sourcing strategies for revegetation in a changing environment. Conserv. Genet. 2012, 14, 1–10. [Google Scholar] [CrossRef]
- Rice, K.J.; Emery, N.C. Managing microevolution: Restoration in the face of global change. Front. Ecol. Environ. 2003, 1, 469–478. [Google Scholar] [CrossRef]
- Lopez, R.; Cano, F.J.; Choat, B.; Cochard, H.; Gil, L. Plasticity in Vulnerability to Cavitation of Pinus canariensis Occurs Only at the Driest End of an Aridity Gradient. Front. Plant Sci. 2016, 7, 769. [Google Scholar] [CrossRef]
- Buras, A.; Sass-Klaassen, U.; Verbeek, I.; Copini, P. Provenance selection and site conditions determine growth performance of pedunculate oak. Dendrochronologia 2020, 61, 125705. [Google Scholar] [CrossRef]

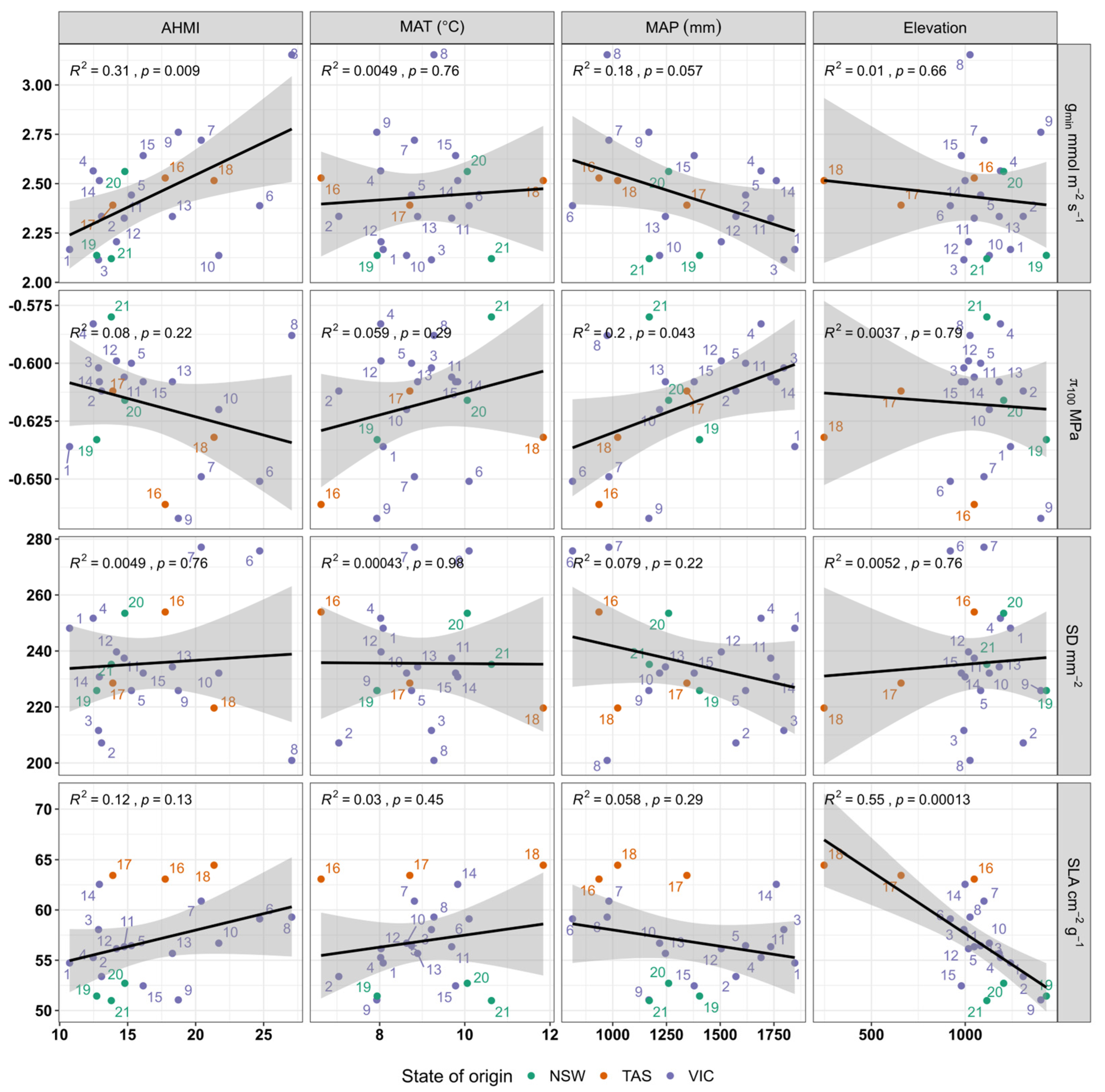

| Climate Variables (Predictor) | Traits | Values | ||
|---|---|---|---|---|
| p-Value | Estimate ± S.E. | Adjusted R-Squared | ||
| AHMI | gmin | 0.011 | 0.67 ± 0.08 | 0.27 |
| π100 | 0.22 | 0.59 ± 0.02 | 0.03 | |
| SD | 0.76 | 230.41 ± 17.49 | −0.05 | |
| SLA | 0.07 | 51.44 ± 3.56 | 0.07 | |
| MAT | gmin | 0.76 | 2.30 ± 0.44 | −0.05 |
| π100 | 0.29 | 0.66 ± 0.04 | 0.01 | |
| SD | 0.98 | 236.52 ± 33.25 | −0.05 | |
| SLA | 0.45 | 51.50 ± 7.07 | −0.02 | |
| MAP | gmin | 0.056 | 2.90 ± 0.24 | 0.13 |
| π100 | 0.043 | 0.67 ± 0.02 | 0.16 | |
| SD | 0.22 | 259.27 ± 19.04 | 0.03 | |
| SLA | 0.30 | 61.26 ± 4.15 | 0.01 | |
| Elevation (DEM) | gmin | 0.66 | 2.54 ± 0.26 | −0.04 |
| π100 | 0.80 | 0.61 ± 0.02 | −0.05 | |
| SD | 0.76 | 229.70 ± 19.37 | −0.05 | |
| SLA | 0.0001 | 70.01 ± 2.83 | 0.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gurung, A.; Wagner, B.; Pryde, E.C.; Nitschke, C.R.; Arndt, S.K. Functional Traits Associated with Drought Tolerance Exhibit Low Variability in 21 Provenances of a Montane Tree Species—Eucalyptus delegatensis. Forests 2025, 16, 898. https://doi.org/10.3390/f16060898
Gurung A, Wagner B, Pryde EC, Nitschke CR, Arndt SK. Functional Traits Associated with Drought Tolerance Exhibit Low Variability in 21 Provenances of a Montane Tree Species—Eucalyptus delegatensis. Forests. 2025; 16(6):898. https://doi.org/10.3390/f16060898
Chicago/Turabian StyleGurung, Anita, Benjamin Wagner, Elizabeth C. Pryde, Craig R. Nitschke, and Stefan K. Arndt. 2025. "Functional Traits Associated with Drought Tolerance Exhibit Low Variability in 21 Provenances of a Montane Tree Species—Eucalyptus delegatensis" Forests 16, no. 6: 898. https://doi.org/10.3390/f16060898
APA StyleGurung, A., Wagner, B., Pryde, E. C., Nitschke, C. R., & Arndt, S. K. (2025). Functional Traits Associated with Drought Tolerance Exhibit Low Variability in 21 Provenances of a Montane Tree Species—Eucalyptus delegatensis. Forests, 16(6), 898. https://doi.org/10.3390/f16060898