Genome-Wide Identification and Expression Profile Analysis of the NF-Y Transcription Factor Gene Family in Eucalyptus grandis

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Sequence Analysis of NF-Y in E. grandis
2.2. Gene Structure and Cis-Regulatory Elements Analysis of the EgrNF-Y Gene Family
2.3. Chromosome Distribution, Duplication, and Synteny Analysis of the EgrNF-Y Genes
2.4. Multiple Alignments and Evolutionary Analysis of EgrNF-Y Proteins
2.5. miRNA Target Site and Gene Interaction NetworkPrediction of the EgrNF-Y Genes
2.6. Plant Material, Cultivation Environment, and P Treatment
2.7. Total RNA Extraction and Real-Time Quantitative PCR Analysis
2.8. Statistical Analysis
3. Results
3.1. Isolation and Identification of the NF-Y Family Members in E. grandis
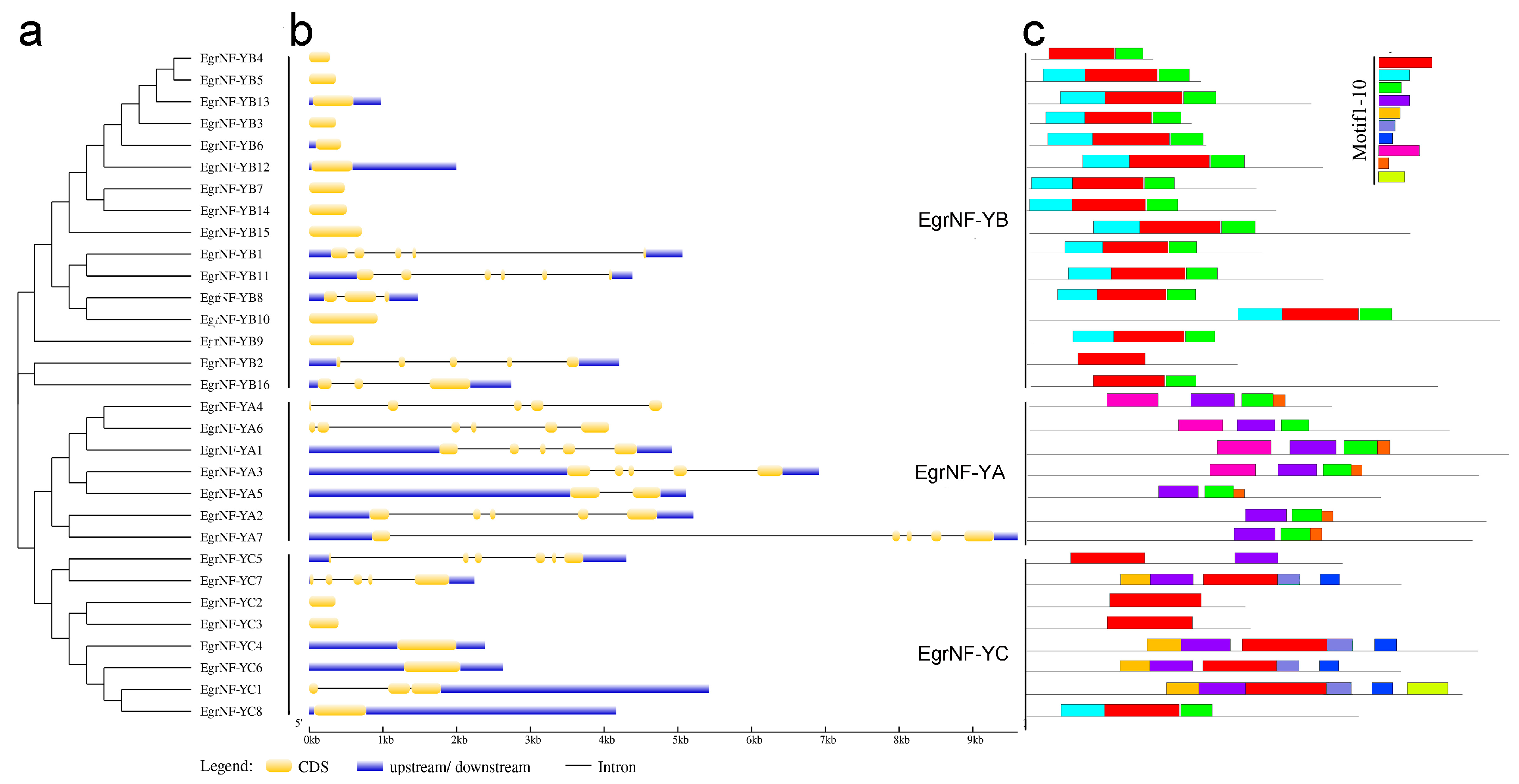
3.2. Analysis of Gene Structure and Conserved Motifs of the EgrNF-Y Genes
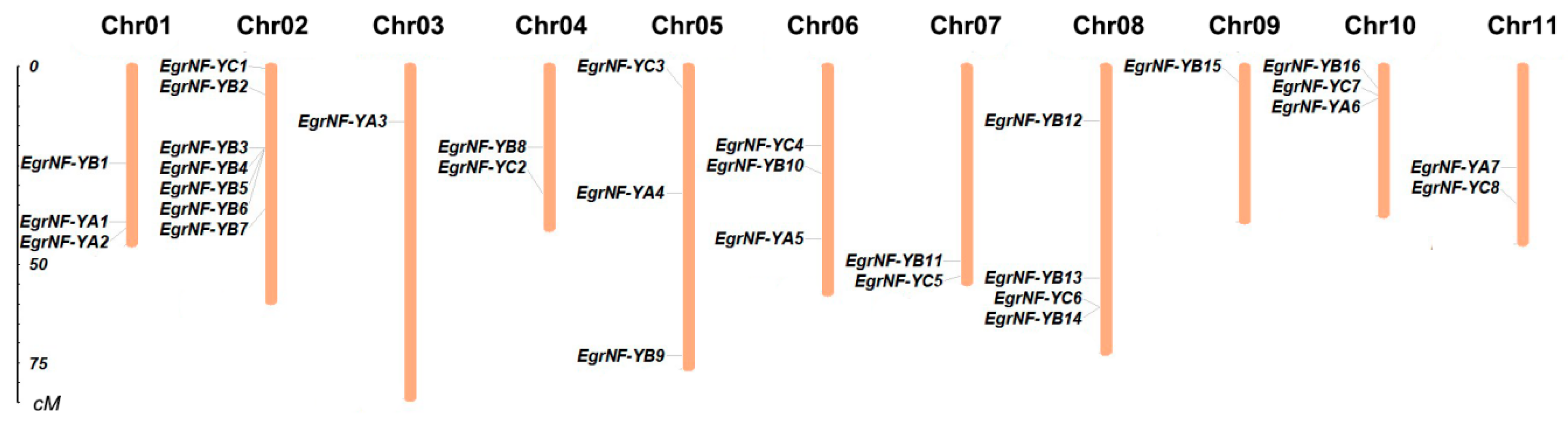
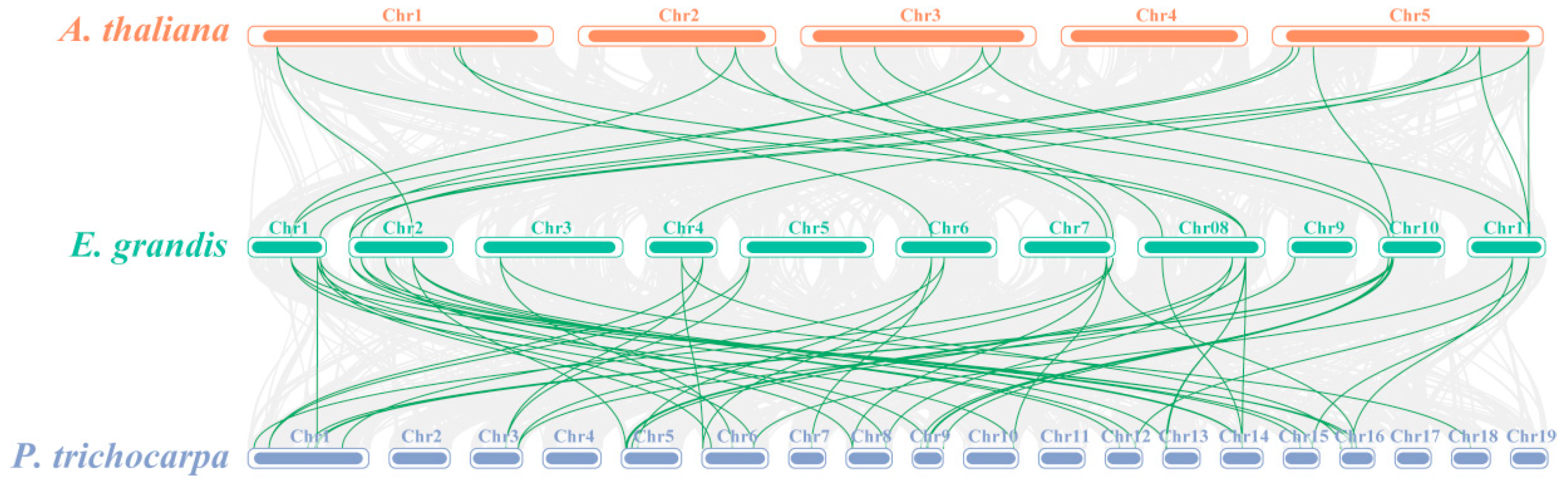
3.3. Chromosomal Distribution and Collinearity Analysis of EgrNF-Y Genes
3.4. Conserved Regions and Phylogenetic Relationships of EgrNF-Ys
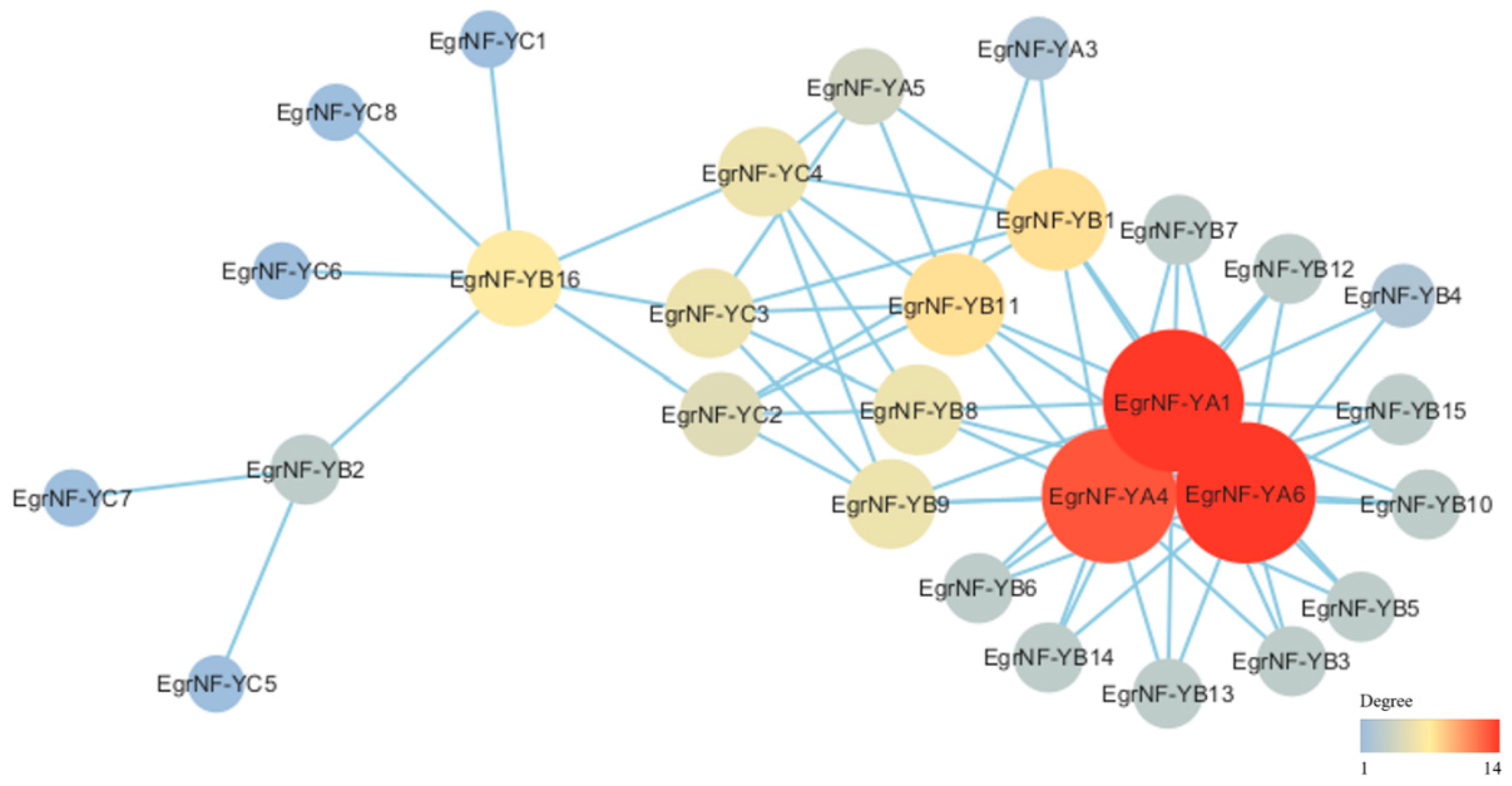
3.5. miRNA Target Site and Protein–Protein Interaction Prediction of the EgrNF-Y Genes
3.6. Identification of Cis-Elements in the Promoter Regions of EgrNF-Y Genes
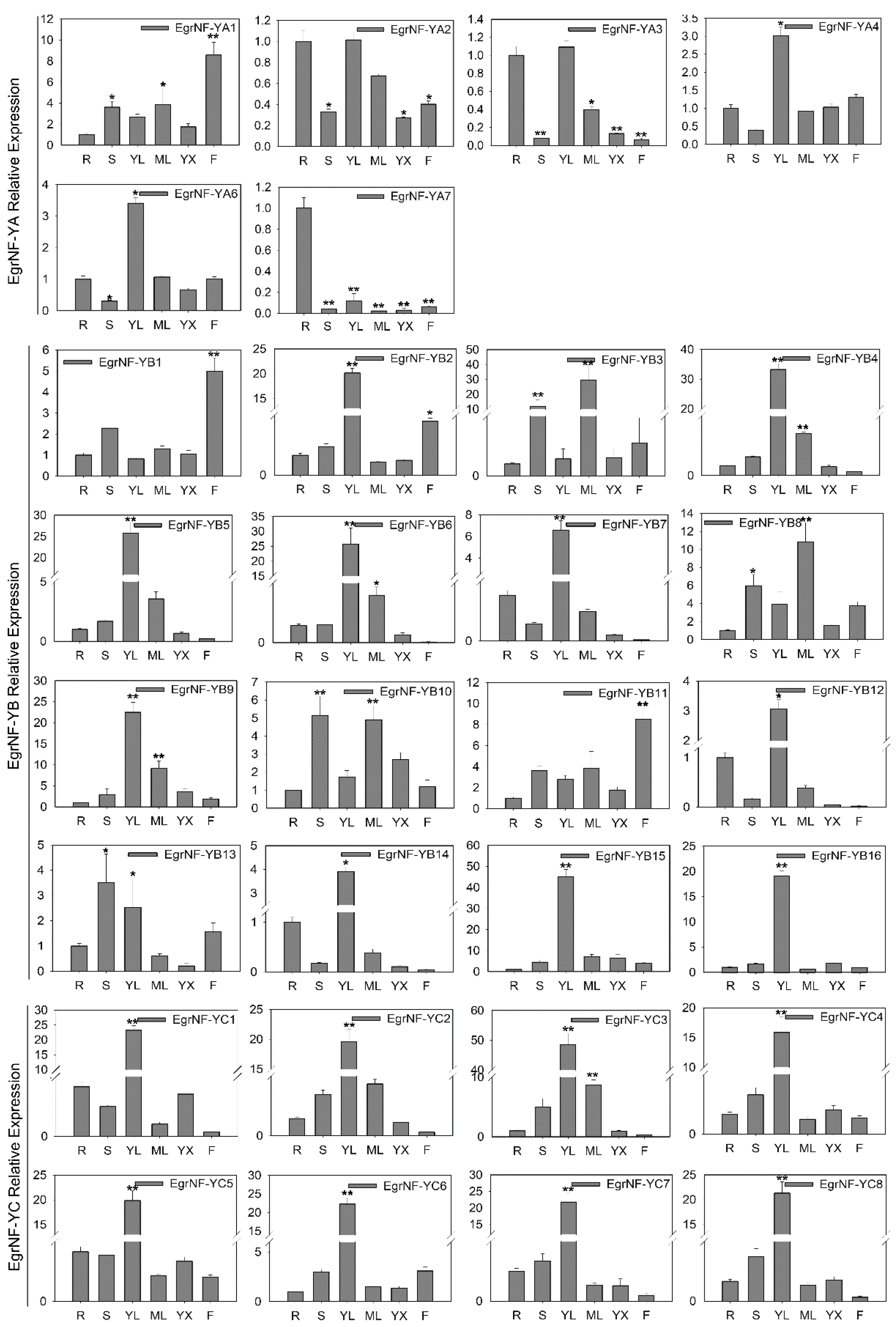
3.7. Expression Patterns of the EgrNF-Y Genes Across Various Tissues
3.8. Expression Profiles of the EgrNF-Y Genes Under Low-Phosphorus Conditions
4. Discussion
4.1. Identification and Characterization of EgrNF-Y Genes in E. grandis
4.2. Core Protein Regions and Phylogenetic Relationships of EgrNF-Ys
4.3. Tissue-Specific Transcription Patterns of EgrNF-Y Genes in E. grandis
4.4. Expression Patterns of EgrNF-Y Genes in Response to Pi Starvation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bucher, P.; Trifonov, E.N. CCAAT box revisited: Bidirectionality, location and context. J. Biomol. Struct. Dyn. 1988, 5, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Maity, S.N.; De Crombrugghe, B. Role of the CCAAT-binding protein CBF/NF-Y in transcription. Trends Biochem. Sci. 1998, 23, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Gusmaroli, G.; Tonelli, C.; Mantovani, R. Regulation of novel members of the Arabidopsis thaliana CCAATbinding nuclear factor Y subunits. Genes 2002, 283, 41–48. [Google Scholar] [CrossRef]
- Chaves-Sanjuan, A.; Gnesutta, N.; Gobbini, A.; Martignago, D.; Bernardini, A.; Fornara, F.; Mantovani, R.; Nardini, M. Structural determinants for NF-Y subunit organization and NF-Y/DNA association in plants. Plant J. 2021, 105, 49–61. [Google Scholar] [CrossRef]
- Siefers, N.; Dang, K.K.; Kumimoto, R.W.; Bynum, W.E.; Tayrose, G.; Holt, B.F., 3rd. Tissue-specific expression patterns of Arabidopsis NF-Y transcription factors suggest potential for extensive combinatorial complexity. Plant Physiol. 2009, 149, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Quach, T.N.; Nguyen, H.T.; Valliyodan, B.; Joshi, T.; Xu, D.; Nguyen, H.T. Genome-wide expression analysis of soybean NF-Y genes reveals potential function in development and drought response. Mol. Genet Genom. 2015, 290, 1095–1115. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jin, Z.; Zhou, M.; Yu, Y.; Liang, M. Characterization of NF-Y transcription factor families in industrial rapeseed (Brassica napus L.) and identification of BnNF-YA3, which functions in the abiotic stress response. Ind. Crops Prod. 2020, 148, 112253. [Google Scholar] [CrossRef]
- An, Y.; Suo, X.; Niu, Q.; Yin, S.; Chen, L. Genome-wide identification and analysis of the NF-Y transcription factor family reveal its potential roles in salt stress in alfalfa (Medicago sativa L.). Int. J. Mol. Sci. 2022, 23, 6426. [Google Scholar] [CrossRef]
- Wei, Q.; Wen, S.; Lan, C.; Yu, Y.; Chen, G. Genome-wide identification and expression profile analysis of the NF-Y transcription factor gene family in Petunia hybrida. Plants 2020, 9, 336. [Google Scholar] [CrossRef]
- Li, J.; Gao, K.; Khan, W.U.; Yang, X.; Yang, X.; Zhao, T.; Chen, Z.; An, X. Genome-wide analysis of the poplar NF-Y gene family and its expression in floral bud development of Populus tomentosa. Trees 2019, 34, 285–296. [Google Scholar] [CrossRef]
- Guo, Y.; Niu, S.; El-Kassaby, Y.A.; Li, W. Transcriptome-wide isolation and expression of NF-Y gene family in male cone development and hormonal treatment of Pinus tabuliformis. Physiol. Plant. 2021, 171, 34–47. [Google Scholar] [CrossRef]
- Pereira, S.L.; Martins, C.P.; Sousa, A.O.; Camillo, L.R.; Araújo, C.P.; Alcantara, G.M.; Camargo, D.S.; Cidade, L.C.; de Almeida, A.A.; Costa, M.G. Genome-wide characterization and expression analysis of citrus NUCLEAR FACTOR-Y (NF-Y) transcription factors identified a novel NF-YA gene involved in drought-stress response and tolerance. PLoS ONE 2018, 13, e0199187. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Hu, A.; Zhang, J.; Liao, W.; Ma, H.; Wu, J. NF-YB-mediated active responses of plant growth under salt and temperature stress in Eucalyptus grandis. Plants 2021, 10, 1107. [Google Scholar] [CrossRef]
- Braybrook, S.A.; Harada, J.J. LECs go crazy in embryo development. Trends Plant Sci. 2008, 13, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zhou, J.; Liu, C.; Liu, L.; Shen, L.; Yu, H. Nuclear factor Y-mediated H3K27me3 demethylation of the SOC1 locus orchestrates flowering responses of Arabidopsis. Nat. Commun. 2014, 5, 4601. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, C.; Xu, Y.; Wang, T.; Chen, Y.; Lü, J.; Zhang, L.; Jiang, C.Z.; Hong, B.; Gao, J. Control of chrysanthemum flowering through integration with an aging pathway. Nat. Commun. 2017, 8, 829. [Google Scholar] [CrossRef]
- Bai, A.N.; Lu, X.D.; Li, D.Q.; Liu, J.X.; Liu, C.M. NF-YB1-regulated expression of sucrose transporters in aleurone facilitates sugar loading to rice endosperm. Cell Res. 2015, 26, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Wang, X.; Han, X.; An, Y.; Lin, S.; Shen, C.; Wen, J.; Liu, C.; Yin, W.; et al. Root-specific NF-Y family transcription factor, PdNF-YB21, positively regulates root growth and drought resistance by abscisic acid-mediated indoylacetic acid transport in Populus. New Phytol. 2020, 227, 407–426. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, K.; Ju, Z.; Cao, D.; Fu, D.; Zhu, H.; Zhu, B.; Luo, Y. Genome-wide analysis of tomato NF-Y factors and their role in fruit ripening. BMC Genom. 2016, 17, 36. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, W.; Chen, Z.; Han, B.; Haque, M.E.; Liu, A. Gene structure, expression pattern and interaction of Nuclear Factor-Y family in castor bean (Ricinus communis). Planta 2017, 247, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Su, H.; Sun, S.; Sun, J.; Zhang, X.; Yu, J. Genome-wide identification and expression profiles of Nuclear Factor Y a transcription factors in blueberry under abiotic stress. Int. J. Mol. Sci. 2024, 25, 12832. [Google Scholar] [CrossRef]
- Ma, X.J.; Yu, T.F.; Li, X.H.; Cao, X.Y.; Xu, Z.S. Overexpression of GmNF-YA5 confers drought tolerance to transgenic Arabidopsis and soybean plants. BioMed Central. 2020, 20, 123. [Google Scholar]
- Sato, H.; Suzuki, T.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NF-YB2 and NF-YB3 have functionally diverged and differentially induce drought and heat stress-specific genes. Plant Physiol. 2019, 180, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Soyano, T.; Kouchi, H.; Hirota, A.; Hayashi, M. NODULE INCEPTION Directly targets NF-Y subunit genes to regulate essential processes of root nodule development in Lotus japonicus. PLoS Genet. 2013, 9, e1003352. [Google Scholar] [CrossRef] [PubMed]
- Leyva-González, M.A.; Ibarra-Laclette, E.; Cruz-Ramírez, A.; Herrera-Estrella, L. Functional and transcriptome analysis reveals an acclimatization strategy for abiotic stress tolerance mediated by Arabidopsis NF-YA family members. PLoS ONE 2012, 7, e48138. [Google Scholar] [CrossRef]
- Qu, B.; He, X.; Wang, J.; Zhao, Y.; Teng, W.; Shao, A.; Zhao, X.; Ma, W.; Wang, J.; Li, B.; et al. A wheat CCAAT box-binding transcription factor increases the grain yield of wheat with less fertilizer input. Plant Physiol. 2015, 167, 411–423. [Google Scholar] [CrossRef]
- Liu, X.; Cai, Y.; Yao, W.; Chen, L.; Hou, W. The soybean NUCLEAR FACTOR-YC4 and α-EXPANSIN 7 module influences phosphorus uptake by regulating root morphology. Plant Physiol. 2025, 197, kiae478. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Kumimoto, R.W.; Gnesutta, N.; Calvenzani, V.; Fornari, M.; Tonelli, C.; Holt, I.I.I.B.F.; Mantovani, R. The promiscuous life of plant NUCLEAR FACTOR Y transcription factors. Plant Cell 2012, 24, 4777–4792. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 6: Recent updates and new developments. Nucleic Acids Res. 2009, 37, D229–D232. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Magali, L.; Patrice, D.; Gert, T.; Kathleen, M.; Yves, M.; Yves, V.D.P. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tong, Y.; Li, Y.; Cheng, Z.M.; Zhong, Y. Genome-wide identification of the HKT genes in five Rosaceae species and expression analysis of HKT genes in response to salt-stress in Fragaria vesca. Genes Genom. 2019, 41, 325–336. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam HClustal, W.; Clustal, X. version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Thirumurugan, T.; Ito, Y.; Kubo, T.; Serizawa, A.; Kurata, N. Identification, characterization and interaction of HAP family genes in rice. Mol. Genet. Genom. 2008, 279, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Böhlenius, H.; Huang, T.; Charbonnel-Campaa, L.; Brunner, A.M.; Jansson, S.; Strauss, S.H.; Nilsson, O. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 2006, 312, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef]
- Xu, M.Y.; Zhang, L.; Li, W.W.; Hu, X.L.; Wang, M.B.; Fan, Y.L.; Zhang, C.Y.; Wanfg, L. Stress-induced early flowering is mediated by miR169 in Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 89–101. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Transcript Name | DNA Length | CDS Length | Peptide Residue | WM (Kd) | pI | Locus | Chromosomes Position | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| NF-YA | |||||||||
| EgrNF-YA1 | Eucgr.A02381.1 | 4923 | 912 | 304 | 33.43 | 9.48 | Chr01 | 39266249–39271172 | Nucleus |
| EgrNF-YA2 | Eucgr.A02570.1 | 5211 | 978 | 326 | 35.34 | 9.13 | Chr01 | 40871807–40877018 | Nucleus |
| EgrNF-YA3 | Eucgr.C00842.1 | 6915 | 1017 | 339 | 37.21 | 9.43 | Chr03 | 13879073–13885988 | Nucleus |
| EgrNF-YA4 | Eucgr.K01943.1 | 4782 | 606 | 202 | 22.24 | 7.90 | Chr05 | 31968551–31973333 | Nucleus |
| EgrNF-YA5 | Eucgr.F03169.1 | 5111 | 777 | 259 | 29.57 | 9.29 | Chr06 | 43547878–43552989 | Nucleus |
| EgrNF-YA6 | Eucgr.J00764.1 | 4067 | 966 | 322 | 34.72 | 6.76 | Chr10 | 8053229–8057296 | Nucleus |
| EgrNF-YA7 | Eucgr.K01921.1 | 9611 | 948 | 316 | 35.25 | 9.53 | Chr11 | 25456393–25466004 | Nucleus |
| NF-YB | |||||||||
| EgrNF-YB1 | Eucgr.A01179.1 | 5062 | 534 | 178 | 19.11 | 6.44 | Chr01 | 24459334–24464396 | Nucleus |
| EgrNF-YB2 | Eucgr.B00669.1 | 4206 | 471 | 157 | 17.36 | 4.70 | Chr02 | 6999487–7003693 | Nucleus |
| EgrNF-YB3 | Eucgr.B01293.1 | 363 | 363 | 121 | 13.65 | 5.90 | Chr02 | 20499626–20499989 | Nucleus |
| EgrNF-YB4 | Eucgr.B01299.1 | 282 | 282 | 94 | 10.64 | 8.37 | Chr02 | 20570932–20571214 | Nucleus |
| EgrNF-YB5 | Eucgr.B01300.1 | 363 | 363 | 121 | 13.59 | 5.89 | Chr02 | 20589022–20589385 | Nucleus |
| EgrNF-YB6 | Eucgr.B01303.1 | 433 | 345 | 115 | 13.05 | 5.89 | Chr02 | 20605624–20606057 | Nucleus |
| EgrNF-YB7 | Eucgr.B01940.1 | 483 | 483 | 161 | 18.18 | 5.17 | Chr02 | 36132078–36132561 | Nucleus |
| EgrNF-YB8 | Eucgr.L00153.1 | 1477 | 666 | 222 | 23.08 | 5.91 | Chr04 | 20329570–20331047 | Nucleus |
| EgrNF-YB9 | Eucgr.E03857.1 | 606 | 606 | 202 | 22.31 | 5.56 | Chr05 | 73093680–73094286 | Nucleus |
| EgrNF-YB10 | Eucgr.F02051.1 | 927 | 927 | 309 | 32.85 | 7.69 | Chr06 | 27056531–27057458 | Nucleus |
| EgrNF-YB11 | Eucgr.G02796.1 | 4385 | 600 | 200 | 22.16 | 8.59 | Chr07 | 49153405–49157790 | Nucleus |
| EgrNF-YB12 | Eucgr.H01189.1 | 1997 | 555 | 185 | 20.37 | 5.84 | Chr08 | 13701266–13703263 | Nucleus |
| EgrNF-YB13 | Eucgr.H03892.1 | 978 | 552 | 184 | 20.55 | 5.89 | Chr08 | 53309416–53310394 | Nucleus |
| EgrNF-YB14 | Eucgr.H04502.1 | 510 | 570 | 170 | 19.10 | 5.26 | Chr08 | 60897292–60897802 | Nucleus |
| EgrNF-YB15 | Eucgr.I00185.1 | 714 | 714 | 238 | 26.87 | 6.30 | Chr09 | 3806528–3807242 | Nucleus |
| EgrNF-YB16 | Eucgr.J00499.1 | 2741 | 858 | 286 | 30.98 | 8.86 | Chr10 | 5349209–5351950 | Nucleus |
| NF-YC | |||||||||
| EgrNF-YC1 | Eucgr.B00106.1 | 5425 | 810 | 270 | 30.02 | 5.59 | Chr02 | 459824–465249 | Nucleus |
| EgrNF-YC2 | Eucgr.D01871.1 | 357 | 357 | 119 | 12.83 | 6.73 | Chr04 | 32254203–32254560 | Nucleus, Cytoplasm |
| EgrNF-YC3 | Eucgr.E00570.1 | 399 | 399 | 133 | 14.67 | 8.82 | Chr05 | 5429231–5429630 | Nucleus |
| EgrNF-YC4 | Eucgr.F01474.1 | 2382 | 798 | 266 | 29.66 | 5.62 | Chr06 | 19881307–19883689 | Nucleus |
| EgrNF-YC5 | Eucgr.G03198.1 | 4300 | 645 | 215 | 24.23 | 9.30 | Chr07 | 52859040–52863340 | Nucleus |
| EgrNF-YC6 | Eucgr.H04476.1 | 2631 | 762 | 254 | 28.14 | 5.30 | Chr08 | 60666762–60669393 | Nucleus |
| EgrNF-YC7 | Eucgr.J00664.1 | 2241 | 795 | 265 | 28.73 | 4.49 | Chr10 | 7199557–7201798 | Nucleus |
| EgrNF-YC8 | Eucgr.K02714.1 | 4166 | 705 | 235 | 25.44 | 5.05 | Chr11 | 34667163–34671329 | Nucleus, Cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Gong, C.; Zhuang, L.; Li, G.; Xu, J.; Lu, Z. Genome-Wide Identification and Expression Profile Analysis of the NF-Y Transcription Factor Gene Family in Eucalyptus grandis. Forests 2025, 16, 361. https://doi.org/10.3390/f16020361
Li J, Gong C, Zhuang L, Li G, Xu J, Lu Z. Genome-Wide Identification and Expression Profile Analysis of the NF-Y Transcription Factor Gene Family in Eucalyptus grandis. Forests. 2025; 16(2):361. https://doi.org/10.3390/f16020361
Chicago/Turabian StyleLi, Juan, Chaoyan Gong, Li Zhuang, Guangyou Li, Jianmin Xu, and Zhaohua Lu. 2025. "Genome-Wide Identification and Expression Profile Analysis of the NF-Y Transcription Factor Gene Family in Eucalyptus grandis" Forests 16, no. 2: 361. https://doi.org/10.3390/f16020361
APA StyleLi, J., Gong, C., Zhuang, L., Li, G., Xu, J., & Lu, Z. (2025). Genome-Wide Identification and Expression Profile Analysis of the NF-Y Transcription Factor Gene Family in Eucalyptus grandis. Forests, 16(2), 361. https://doi.org/10.3390/f16020361