Physiological and Growth Responses to Increasing Drought of an Endangered Tree Species in Southwest China

 ,
,
Abstract
1. Introduction
2. Data and Methods
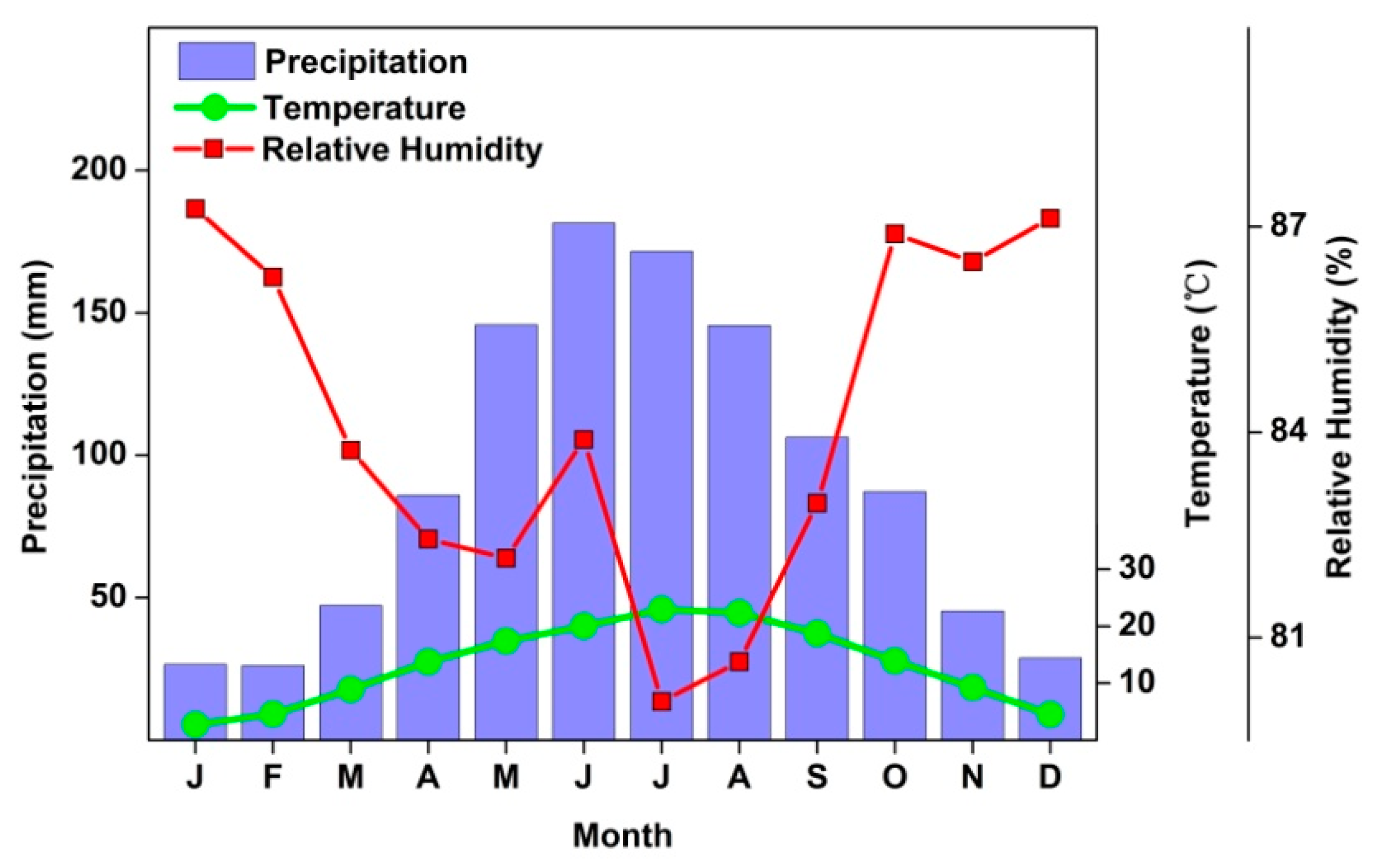
2.1. Research Area
2.2. Climate Data
2.3. Core Sampling and Analysis
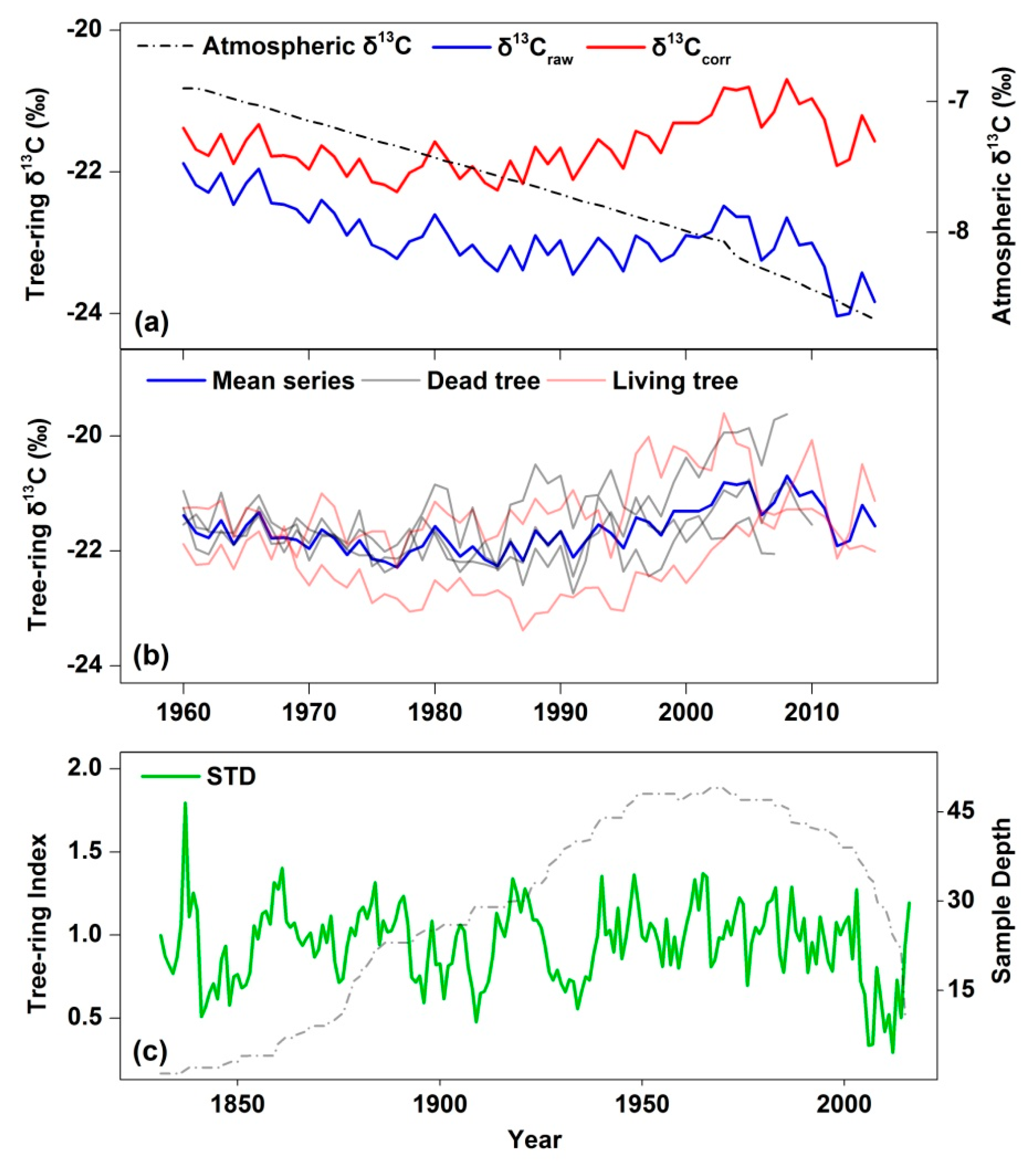
2.4. Carbon Isotope Chronology and Calculations for iWUE
2.5. Climate-Growth Relationships
3. Results
3.1. Climatic Characteristic
3.2. Temporal Characteristics of δ13C and STD Chronologies
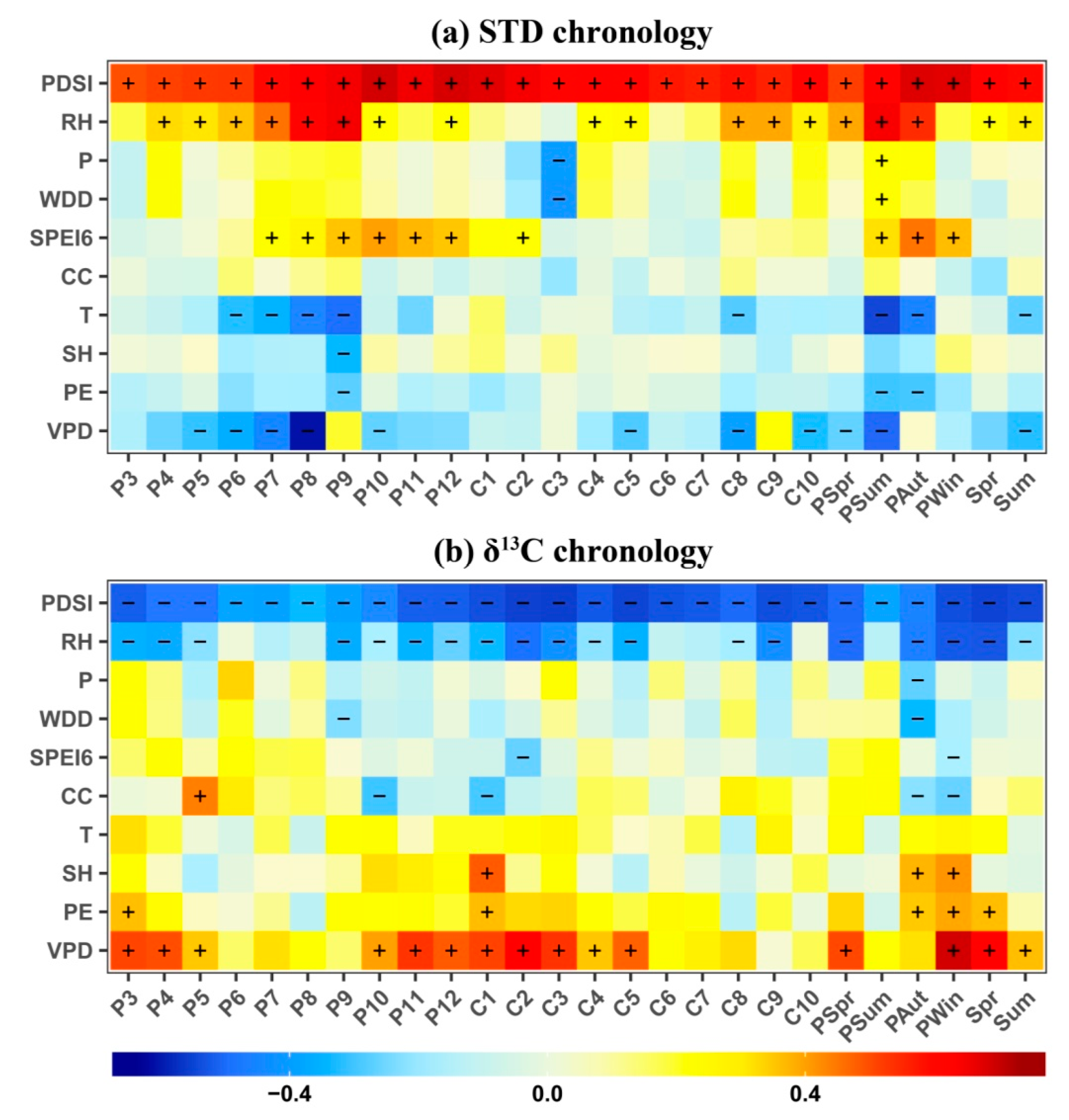
3.3. Climate Response of STD and δ13C Chronologies
3.4. Temporal Variations in Ca, Ci, Ci/Ca, BAI, STD, and iWUE and Their Correlations with Climate.
4. Discussion
4.1. Climatic Information Recorded by the Tree-Ring δ13C Series
4.2. The Physiological Implications of Radial Growth and iWUE
4.3. Possible Mechanisms of Tree Mortality
4.4. Implications for Forest Management and Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers of Climate Change 2013: The Physical Science Basis; Contribution of working group i to the fifth assessment report of the intergovernmental panel on climate change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Stott, P.A.; Mitchell, J.F.B.; Allen, M.R.; Delworth, T.L.; Gregory, J.M.; Meehl, G.A.; Santer, B.D. Observational constraints on past attributable warming and predictions of future global warming. Am. Meteorol. Soc. 2006, 19, 3055–3069. [Google Scholar] [CrossRef]
- Cox, P.M.; Betts, R.A.; Jones, C.D.; Spall, S.A.; Totterdell, I.J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 2000, 6809, 184. [Google Scholar] [CrossRef] [PubMed]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Vilalta, J.; Prat, E.; Oliveras, I.; Pinol, J. Xylem hydraulic properties of roots and stems of nine mediterranean woody species. Oecologia 2002, 133, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Wei, L.; Xu, C.; Jansen, S.; Zhou, H.; Christoffersen, B.O.; Pockman, W.T.; Middleton, R.S.; Marshall, J.D.; McDowell, N.G. A heuristic classification of woody plants based on contrasting shade and drought strategies. Tree Physiol. 2019. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhausser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Su, J.; Gou, X.; Deng, Y.; Zhang, R.; Liu, W.; Zhang, F.; Lu, M.; Chen, Y.; Zheng, W. Tree growth response of fokienia hodginsii to recent climate warming and drought in southwest china. Int. J. Biometeorol. 2017, 61, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annual review of plant biology. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Long, A. Drought indicated in carbon-13lcarbon-12 ratios of southwestern tree rings. J. Am. Water Res. Assoc. 1989, 25, 341–347. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.T.W.; Schweingruber, F.H. Carbon isotope discrimination indicates improving water-use efficiency of trees in northern eurasia over the last 100 years. Glob. Chang. Biol. 2004, 10, 2109–2120. [Google Scholar] [CrossRef]
- Herrero, A.; Castro, J.; Zamora, R.; Delgado-Huertas, A.; Querejeta, J.I. Growth and stable isotope signals associated with drought-related mortality in saplings of two coexisting pine species. Oecologia 2013, 173, 1613–1624. [Google Scholar] [CrossRef]
- Seibt, U.; Rajabi, A.; Griffiths, H.; Berry, J.A. Carbon isotopes and water use efficiency: Sense and sensitivity. Oecologia 2008, 155, 441–454. [Google Scholar] [CrossRef]
- Lévesque, M.; Rigling, A.; Bugmann, H.; Weber, P.; Brang, P. Growth response of five co-occurring conifers to drought across a wide climatic gradient in central europe. Agric. For. Meteorol. 2014, 197, 1–12. [Google Scholar] [CrossRef]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.F.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Levanic, T.; Cater, M.; McDowell, N.G. Associations between growth, wood anatomy, carbon isotope discrimination and mortality in a quercus robur forest. Tree Physiol. 2011, 31, 298–308. [Google Scholar] [CrossRef]
- Timofeeva, G.; Treydte, K.; Bugmann, H.; Rigling, A.; Schaub, M.; Siegwolf, R.; Saurer, M. Long-term effects of drought on tree-ring growth and carbon isotope variability in scots pine in a dry environment. Tree Physiol. 2017, 37, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cerrillo, R.M.; Sarmoum, M.; Gazol, A.; Abdoun, F.; Camarero, J.J. The decline of algerian cedrus atlantica forests is driven by a climate shift towards drier conditions. Dendrochronologia 2019, 55, 60–70. [Google Scholar] [CrossRef]
- Wang, A.; Liu, Y.; Liu, S.; Yang, T. A preliminary report on the geographic distribution of nature forest fokienia hodginsii in china. Chin. J. Ecol. 1984, 4, 19–23. (In Chinese) [Google Scholar]
- Li, M.; Deng, L.; Pan, D.; Mu, J.; Li, C.; Yang, C. Study on growth process of fokienia hodginsii community in xishui nature reserve in guizhou. J. Anhui Agric. 2015, 43, 199–202. (In Chinese) [Google Scholar]
- Li, M.; Deng, L.; Jiang, Y.; Yang, C.; Mu, J. Community composition and structure of fokienia hodginsii community in xishui nature reserve in guizhou. J. Northwest For. Univ. 2013, 28, 46–50. (In Chinese) [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Cook, E.R. A Time Series Approach to Tree-Ring Standardization; University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: Cambridge, MA, USA, 1976. [Google Scholar]
- Phipps, R.; Whiton, J. Decline in long-term growth trends of white oak. Can. J. For. Res. 1988, 18, 24–32. [Google Scholar] [CrossRef]
- Brendel, O.; Iannetta, P.P.M.; Stewart, D. A rapid and simple method to isolate pure alpha-cellulose. Phytochem. Anal. 2000, 11, 7–10. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W.; Zhou, W.; Huang, G. Teleconnected influence of tropical northwest pacific sea surface temperature on interannual variability of autumn precipitation in southwest china. Clim. Dyn. 2015, 45, 2527–2539. [Google Scholar] [CrossRef]
- Zhang, M.; He, J.; Wang, B.; Wang, S.; Li, S.; Liu, W.; Ma, X. Extreme drought changes in southwest china from 1960 to 2009. J. Geogr. Sci. 2013, 23, 3–16. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.; Comita, L.S.; Condit, R.; Kursar, T.A.; Tyree, M.T.; Turner, B.L.; Hubbell, S.P. Drought sensitivity shapes species distribution patterns in tropical forests. Nature 2007, 447, 80–82. [Google Scholar] [CrossRef] [PubMed]
- D’Orangeville, L.; Duchesne, L.; Houle, D.; Kneeshaw, D.; Cote, B.; Pederson, N. Northeastern north america as a potential refugium for boreal forests in a warming climate. Science 2016, 352, 1452–1455. [Google Scholar] [CrossRef] [PubMed]
- Phillips, O.L.; Aragao, L.E.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; Lopez-Gonzalez, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef] [PubMed]
- Eugene, A.V.; Hughes, M.K.; Alexander, V.S. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments; Springer Science & Business Media: Berlin, Germany, 2006; p. 351. [Google Scholar]
- Rolland, C. Tree-ring and climate relationships for abies alba in the internal alps. Tree-Ring Bull. 1993, 53, 1–11. [Google Scholar]
- Shi, J.; Lu, H.; Li, J.; Shi, S.; Wu, S.; Hou, X.; Li, L. Tree-ring based february–april precipitation reconstruction for the lower reaches of the yangtze river, southeastern china. Glob. Planet. Chang. 2015, 131, 82–88. [Google Scholar] [CrossRef]
- Evans, M.N.; Reichert, B.K.; Kaplan, A.; Anchukaitis, K.J.; Vaganov, E.A.; Hughes, M.K.; Cane, M.A. A forward modeling approach to paleoclimatic interpretation of tree-ring data. J. Geophys. Res. Biogeosci. 2006, 111, G03008. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Buckley, B.M.; Anchukaitis, K.J.; Penny, D.; Fletcher, R.; Cook, E.R.; Sano, M.; Nam le, C.; Wichienkeeo, A.; Minh, T.T.; Hong, T.M. Climate as a contributing factor in the demise of angkor, cambodia. Proc. Natl. Acad. Sci. USA 2010, 107, 6748–6752. [Google Scholar] [CrossRef]
- Sano, M.; Buckley, B.M.; Sweda, T. Tree-ring based hydroclimate reconstruction over northern vietnam from fokienia hodginsii: Eighteenth century mega-drought and tropical pacific influence. Clim. Dyn. 2008, 33, 331–340. [Google Scholar] [CrossRef]
- Fan, Z.X.; Bräuning, A.; Cao, K.F. Tree-ring based drought reconstruction in the central hengduan mountains region (China) since a.D. 1655. Int. J. Climatol. 2008, 28, 1879–1887. [Google Scholar] [CrossRef]
- Soule, P.T.; Knapp, P.A. Radial growth rate increases in naturally occurring ponderosa pine trees: A late-20th century CO2 fertilization effect? N. Phytol. 2006, 171, 379–390. [Google Scholar] [CrossRef]
- Silva, L.C.R.; Anand, M.; Shipley, B. Probing for the influence of atmospheric co2 and climate change on forest ecosystems across biomes. Glob. Ecol. Biogeogr. 2013, 22, 83–92. [Google Scholar] [CrossRef]
- Andreu-Hayles, L.; Planells, O.; GutiÉRrez, E.; Muntan, E.; Helle, G.; Anchukaitis, K.J.; Schleser, G.H. Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five iberian pine forests. Glob. Chang. Biol. 2011, 17, 2095–2112. [Google Scholar] [CrossRef]
- Linares, J.C.; Delgado-Huertas, A.; Julio Camarero, J.; Merino, J.; Carreira, J.A. Competition and drought limit the response of water-use efficiency to rising atmospheric carbon dioxide in the mediterranean fir abies pinsapo. Oecologia 2009, 161, 611–624. [Google Scholar] [CrossRef]
- Manzanedo, R.D.; Ballesteros-Canovas, J.; Schenk, F.; Stoffel, M.; Fischer, M.; Allan, E. Increase in CO2 concentration could alter the response of hedera helix to climate change. Ecol. Evol. 2018, 8, 8598–8606. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faeia, T.; Pinheiro, C. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef]
- Silva, L.C.; Horwath, W.R. Explaining global increases in water use efficiency: Why have we overestimated responses to rising atmospheric CO2 in natural forest ecosystems? PLoS ONE 2013, 8, e53089. [Google Scholar] [CrossRef]
- Nock, C.A.; Baker, P.J.; Wanek, W.; Leis, A.; Grabner, M.; Bunyavejchewin, S.; Hietz, P. Long-term increases in intrinsic water-use efficiency do not lead to increased stem growth in a tropical monsoon forest in western thailand. Glob. Chang. Biol. 2011, 17, 1049–1063. [Google Scholar] [CrossRef]
- Peñuelas, J.; Hunt, J.M.; Ogaya, R.; Jump, A.S. Twentieth century changes of tree-ring δ13c at the southern range-edge of fagus sylvatica: Increasing water-use efficiency does not avoid the growth decline induced by warming at low altitudes. Glob. Chang. Biol. 2008, 14, 1076–1088. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Sala, A. Lack of direct evidence for the carbon-starvation hypothesis to explain drought-induced mortality in trees. Proc. Natl. Acad. Sci. USA 2009, 106, E68. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Meinzer, F.C.; McCulloh, K.A.; Lachenbruch, B.; Woodruff, D.R.; Johnson, D.M. The blind men and the elephant: The impact of context and scale in evaluating conflicts between plant hydraulic safety and efficiency. Oecologia 2010, 164, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Meinzer, F.C.; Johnson, D.M.; Lachenbruch, B.; McCulloh, K.A.; Woodruff, D.R. Xylem hydraulic safety margins in woody plants: Coordination of stomatal control of xylem tension with hydraulic capacitance. Funct. Ecol. 2009, 23, 922–930. [Google Scholar] [CrossRef]
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global warming-related tree growth decline and mortality on the north-eastern tibetan plateau. Clim. Chang. 2015, 134, 163–176. [Google Scholar] [CrossRef]
- Zhao, S.; Jiang, Y.; Dong, M.; Xu, H.; Manzanedo, R.D.; Pederson, N. Early monsoon failure and mid-summer dryness induces growth cessation of lower range margin picea crassifolia. Trees 2018, 32, 1401–1413. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? N. Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Fensham, R.J.; Fairfax, R.J. Drought-related tree death of savanna eucalypts: Species susceptibility, soil conditions and root architecture. J. Veg. Sci. 2007, 18, 71–80. [Google Scholar] [CrossRef]
- Phillips, O.L.; van der Heijden, G.; Lewis, S.L.; Lopez-Gonzalez, G.; Aragao, L.E.; Lloyd, J.; Malhi, Y.; Monteagudo, A.; Almeida, S.; Davila, E.A.; et al. Drought-mortality relationships for tropical forests. N. Phytol. 2010, 187, 631–646. [Google Scholar] [CrossRef]
- Niklas, K.J.; Spatz, H.C. Growth and hydraulic (not mechanical) constraints govern the scaling of tree height and mass. Proc. Natl. Acad. Sci. USA 2004, 101, 15661–15663. [Google Scholar] [CrossRef]
- Wu, G.; Liu, X.; Chen, T.; Xu, G.; Wang, W.; Zeng, X.; Wang, B.; Zhang, X. Long-term variation of tree growth and intrinsic water-use efficiency in schrenk spruce with increasing CO2 concentration and climate warming in the western tianshan mountains, China. Acta Physiol. Plant. 2015, 37, 150. [Google Scholar] [CrossRef]
- Candel-Pérez, D.; Lo, Y.-H.; Blanco, J.; Chiu, C.-M.; Camarero, J.; González de Andrés, E.; Imbert, J.; Castillo, F. Drought-induced changes in wood density are not prevented by thinning in scots pine stands. Forests 2018, 9, 4. [Google Scholar] [CrossRef]
- Grossiord, C. Having the right neighbors: How tree species diversity modulates drought impacts on forests. N. Phytol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Konings, A.G.; Trugman, A.T.; Yu, K.; Bowling, D.R.; Gabbitas, R.; Karp, D.S.; Pacala, S.; Sperry, J.S.; Sulman, B.N.; et al. Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature 2018, 561, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Peng, C.; Li, Y.; Liu, S.; Zhang, Q.; Tang, X.; Liu, J.; Yan, J.; Zhang, D.; Chu, G. A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in southern china. Glob. Chang. Biol. 2013, 19, 1197–1210. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Dead or Living | Latitude (N) | Longitude (E) | Elevation (m a.s.l.) | Cores/Trees | Slope (°) | Slope Aspect | Time Span (a) |
|---|---|---|---|---|---|---|---|---|
| SCH01 | Dead | 28°33′44″ | 106°24′05″ | 1200–1270 | 30/17 | 40 | Southwest | 125.3 |
| SCH02 | Living | 28°32′14″ | 106°23′54″ | 1200–1240 | 38/18 | 40 | Southwest | 89.4 |
| Cores ID | Dead or Alive | Time Span (years) | Mean Ring Width (mm) | Diameter at Breast Height (cm) | Height (m) | Canopy Position |
|---|---|---|---|---|---|---|
| WXT01B | dead | 1877–2007 | 1.53 | 45 | 19.0 | dominant |
| WXT02A | dead | 1847–2007 | 1.42 | 49 | 23.0 | dominant |
| WXT27A | dead | 1875–2010 | 1.07 | 32 | 20.5 | dominant |
| WXT34C | alive | 1889–2016 | 1.31 | 37 | 16.5 | codominant |
| WXT38A | alive | 1895–2016 | 1.50 | 39 | 15.0 | codominant |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, W.; Gou, X.; Su, J.; Fan, H.; Yu, A.; Liu, W.; Deng, Y.; Manzanedo, R.D.; Fonti, P. Physiological and Growth Responses to Increasing Drought of an Endangered Tree Species in Southwest China. Forests 2019, 10, 514. https://doi.org/10.3390/f10060514
Zheng W, Gou X, Su J, Fan H, Yu A, Liu W, Deng Y, Manzanedo RD, Fonti P. Physiological and Growth Responses to Increasing Drought of an Endangered Tree Species in Southwest China. Forests. 2019; 10(6):514. https://doi.org/10.3390/f10060514
Chicago/Turabian StyleZheng, Wuji, Xiaohua Gou, Jiajia Su, Haowen Fan, Ailing Yu, Wenhuo Liu, Yang Deng, Rubén D. Manzanedo, and Patrick Fonti. 2019. "Physiological and Growth Responses to Increasing Drought of an Endangered Tree Species in Southwest China" Forests 10, no. 6: 514. https://doi.org/10.3390/f10060514
APA StyleZheng, W., Gou, X., Su, J., Fan, H., Yu, A., Liu, W., Deng, Y., Manzanedo, R. D., & Fonti, P. (2019). Physiological and Growth Responses to Increasing Drought of an Endangered Tree Species in Southwest China. Forests, 10(6), 514. https://doi.org/10.3390/f10060514