
Exploring the Impact of Environmental Conditions and Bioreactors on Microalgae Growth and Applications



Abstract
1. Introduction

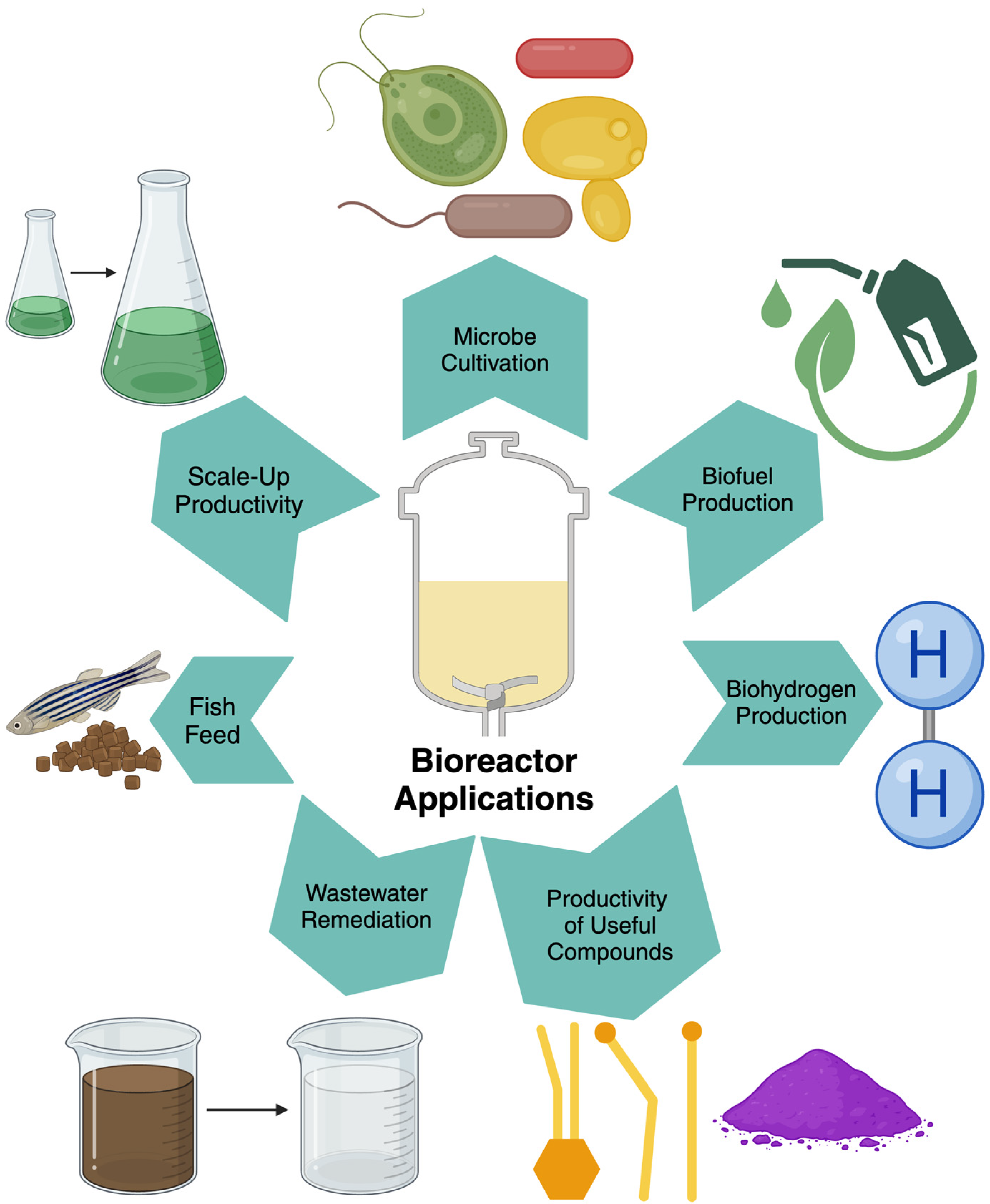
2. Types and Uses of Bioreactors
3. Photobioreactors for Energy
4. Incubation Factors for Photobioreactors
5. Challenges and Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kwon, M.H.; Yeom, S.H. Evaluation of closed photobioreactor types and operation variables for enhancing lipid productivity of Nannochloropsis sp. KMMCC 290 for biodiesel production. Biotechnol. Bioprocess Eng. 2017, 22, 604–611. [Google Scholar] [CrossRef]
- Regueiras, A.; Huguet, Á.; Conde, T.; Couto, D.; Domingues, P.; Domingues, M.R.; Costa, A.M.; da Silva, J.L.; Vasconcelos, V.; Urbatzka, R. Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions. Mar. Drugs 2022, 20, 9. [Google Scholar] [CrossRef]
- Arunkumar, K.; Nalluri, M.; Anjana, K.; Mohan, G.; Raja, R. Fucoxanthin as antioxidant, anti-hyaluronidase and cytotoxic agent: Potential of brown seaweeds decoction for tea supplement. J. Food Meas. Charact. 2023, 17, 3980–3989. [Google Scholar] [CrossRef]
- Truong, T.Q.; Park, Y.J.; Winarto, J.; Huynh, P.K.; Moon, J.; Choi, Y.B.; Song, D.-G.; Koo, S.Y.; Kim, S.M. Understanding the Impact of Nitrogen Availability: A Limiting Factor for Enhancing Fucoxanthin Productivity in Microalgae Cultivation. Mar. Drugs 2024, 22, 93. [Google Scholar] [CrossRef]
- D’Imporzano, G.; Veronesi, D.; Salati, S.; Adani, F. Carbon and nutrient recovery in the cultivation of Chlorella vulgaris: A life cycle assessment approach to comparing environmental performance. J. Clean. Prod. 2018, 194, 685–694. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, Y.; Liu, Y.; Esakkimuthu, S.; Chen, H.; Wang, S. Impact of constant magnetic field on enhancing the microalgal biomass and biomolecules accumulations and life cycle assessment of the approach. Algal Res. 2024, 80, 103563. [Google Scholar] [CrossRef]
- Arashiro, L.T.; Josa, I.; Ferrer, I.; Van Hulle, S.W.H.; Rousseau, D.P.L.; Garfí, M. Life cycle assessment of microalgae systems for wastewater treatment and bioproducts recovery: Natural pigments, biofertilizer and biogas. Sci. Total Environ. 2022, 847, 157615. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, R.; Wang, N.; Yang, T.; Xu, F.; Li, J.; Wu, J.; Huang, Z.; Pan, M.; Lyu, T. Cultivation of microalgae in adjusted wastewater to enhance biofuel production and reduce environmental impact: Pyrolysis performances and life cycle assessment. J. Clean. Prod. 2022, 355, 131768. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, J.; Zhang, Y.; He, S.; Esakkimuthu, S.; Zhu, K.; Kumar, S.; Lv, G.; Hu, X. Biochar assisted cultivation of Chlorella protothecoides for adsorption of tetracycline and electrochemical study on self-cultured Chlorella protothecoides. Bioresour. Technol. 2023, 389, 129810. [Google Scholar] [CrossRef]
- Scaria, J.; Anupama, K.V.; Nidheesh, P.V. Tetracyclines in the environment: An overview on the occurrence, fate, toxicity, detection, removal methods, and sludge management. Sci. Total Environ. 2021, 771, 145291. [Google Scholar] [CrossRef]
- Amangelsin, Y.; Semenova, Y.; Dadar, M.; Aljofan, M.; Bjørklund, G. The Impact of Tetracycline Pollution on the Aquatic Environment and Removal Strategies. Antibiotics 2023, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Prigiobbe, V.; Abimbola, T.; Christodoulatos, C. Integrating biological and chemical CO2 sequestration using green microalgae for bioproducts generation. Front. Clim. 2023, 4, 949411. [Google Scholar] [CrossRef]
- Lan, X.; Tans, P.; Thoning, K.W. Trends in Globally-Averaged CO2 Determined from NOAA Global Monitoring Laboratory Measurements, Version 2024-10; NOAA Global Monitoring Laboratory: Boulder, CO, USA, 2024. [CrossRef]
- Wu, Z.; Vermeulen, A.; Sawa, Y.; Karstens, U.; Peters, W.; de Kok, R.; Lan, X.; Nagai, Y.; Ogi, A.; Tarasova, O. Investigating the differences in calculating global mean surface CO2 abundance: The impact of analysis methodologies and site selection. Atmos. Chem. Phys. 2024, 24, 1249–1264. [Google Scholar] [CrossRef]
- Xin, Y.; Shen, C.; She, Y.; Chen, H.; Wang, C.; Wei, L.; Yoon, K.; Han, D.; Hu, Q.; Xu, J. Biosynthesis of Triacylglycerol Molecules with a Tailored PUFA Profile in Industrial Microalgae. Mol. Plant 2019, 12, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.L.; Falé, P.; Afonso, C.M.; Bandarra, N.M. Investigation of nutraceutical potential of the microalgae Chlorella vulgaris and Arthrospira platensis. Int. J. Food Sci. Technol. 2020, 55, 303–312. [Google Scholar] [CrossRef]
- Shimidzu, N.; Goto, M.; Miki, W. Carotenoids as Singlet Oxygen Quenchers in Marine Organisms. Fish. Sci. 1996, 62, 134–137. [Google Scholar] [CrossRef]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, B.; Hu, Q.; Sommerfeld, M.; Li, Y.; Han, D. A New Paradigm for Producing Astaxanthin from the Unicellular Green Alga Haematococcus pluvialis. Biotechnol. Bioeng. 2016, 113, 2088–2099. [Google Scholar] [CrossRef]
- Chaudhary, A.; Mishra, P.; Amaz, S.A.; Mahato, P.L.; Das, R.; Jha, R.; Mishra, B. Dietary supplementation of microalgae mitigates the negative effects of heat stress in broilers. Poult. Sci. 2023, 102, 102958. [Google Scholar] [CrossRef]
- Sforza, E.; Bertucco, A.; Morosinotto, T.; Giacometti, G.M. Photobioreactors for microalgal growth and oil production with Nannochloropsis salina: From lab-scale experiments to large-scale design. Chem. Eng. Res. Des. 2012, 90, 1151–1158. [Google Scholar] [CrossRef]
- Manhaeghe, D.; Blomme, T.; Van Hulle, S.W.H.; Rousseau, D.P.L. Experimental assessment and mathematical modelling of the growth of Chlorella vulgaris under photoautotrophic, heterotrophic and mixotrophic conditions. Water Res. 2020, 184, 116152. [Google Scholar] [CrossRef] [PubMed]
- Al Jabri, H.; Taleb, A.; Touchard, R.; Saadaoui, I.; Goetz, V.; Pruvost, J. Cultivating Microalgae in Desert Conditions: Evaluation of the Effect of Light-Temperature Summer Conditions on the Growth and Metabolism of Nannochloropsis QU130. Appl. Sci. 2021, 11, 3799. [Google Scholar] [CrossRef]
- Romagnoli, F.; Weerasuriya-Arachchige, A.R.P.P.; Paoli, R.; Feofilovs, M.; Ievina, B. Growth Kinetic Model for Microalgae Cultivation in Open Raceway Ponds: A System Dynamics Tool. Environ. Clim. Technol. 2021, 25, 1317–1336. [Google Scholar] [CrossRef]
- Bartley, M.L.; Boeing, W.J.; Dungan, B.N.; Holguin, F.O.; Schaub, T. pH effects on growth and lipid accumulation of the biofuel microalgae Nannochloropsis salina and invading organisms. J. Appl. Phycol. 2014, 26, 1431–1437. [Google Scholar] [CrossRef]
- Qiu, R.; Gao, S.; Lopez, P.A.; Ogden, K.L. Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Res. 2017, 28, 192–199. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Hrůzová, K.; Rova, U.; Christakopoulos, P. Biosynthesis of Nutraceutical Fatty Acids by the Oleaginous Marine Microalgae Phaeodactylum tricornutum Utilizing Hydrolysates from Organosolv-Pretreated Birch and Spruce Biomass. Mar. Drugs 2019, 17, 119. [Google Scholar] [CrossRef]
- Aussant, J.; Guihéneuf, F.; Stengel, D.B. Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Appl. Microbiol. Biotechnol. 2018, 102, 5279–5297. [Google Scholar] [CrossRef]
- Perin, G.; Cimetta, E.; Monetti, F.; Morosinotto, T.; Bezzo, F. Novel micro-photobioreactor design and monitoring method for assessing microalgae response to light intensity. Algal Res. 2016, 19, 69–76. [Google Scholar] [CrossRef]
- Nzayisenga, J.C.; Farge, X.; Groll, S.L.; Sellstedt, A. Effects of light intensity on growth and lipid production in microalgae grown in wastewater. Biotechnol. Biofuels 2020, 13, 4. [Google Scholar] [CrossRef]
- Fawzy, M.A.; El-Otify, A.M.; Adam, M.S.; Moustafa, S.S.A. The impact of abiotic factors on the growth and lipid accumulation of some green microalgae for sustainable biodiesel production. Environ. Sci. Pollut. Res. 2021, 28, 42547–42561. [Google Scholar] [CrossRef]
- Gao, B.; Hong, J.; Chen, J.; Zhang, H.; Hu, R.; Zhang, C. The growth, lipid accumulation and adaptation mechanism in response to variation of temperature and nitrogen supply in psychrotrophic filamentous microalga Xanthonema hormidioides (Xanthophyceae). Biotechnol. Biofuels Bioprod. 2023, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Josephine, A.; Kumar, T.S.; Surendran, B.; Rajakumar, S.; Kirubagaran, R.; Dharani, G. Evaluating the effect of various environmental factors on the growth of the marine microalgae, Chlorella vulgaris. Front. Mar. Sci. 2022, 9, 954622. [Google Scholar] [CrossRef]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of Microalgae Cultivation in Photobioreactor, Open Raceway Pond, and a Two-Stage Hybrid System. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Prado, L.O.; Bolzani, H.R.; Souza, H.H.S.; Ruas, G.; Silva, G.H.R. Microalgal cultivation in open and closed systems under a tropical climate: A life cycle comparison. J. Clean. Prod. 2023, 422, 138631. [Google Scholar] [CrossRef]
- Naumann, T.; Çebi, Z.; Podola, B.; Melkonian, M. Growing microalgae as aquaculture feeds on twin-layers: A novel solid-state photobioreactor. J. Appl. Phycol. 2013, 25, 1413–1420. [Google Scholar] [CrossRef]
- Magalhães, I.B.; Ferreira, J.; Castro, J.d.S.; de Assis, L.R.; Calijuri, M.L. Agro-industrial wastewater-grown microalgae: A techno-environmental assessment of open and closed systems. Sci. Total Environ. 2022, 834, 155282. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.D.; Mota, A.; Ferreira, A.; Dragone, G.; Teixeira, J.A.; Vicente, A.A. Characterization of split cylinder air-lift photobioreactors for efficient microalgae cultivation. Chem. Eng. Sci. 2014, 117, 445–454. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chang, Y.-H.; Chang, H.-Y. Outdoor cultivation of Chlorella vulgaris FSP-E in vertical tubular-type photobioreactors for microalgal protein production. Algal Res. 2016, 13, 264–270. [Google Scholar] [CrossRef]
- del Rio-Chanona, E.A.; Liu, J.; Wagner, J.L.; Zhang, D.; Meng, Y.; Xue, S.; Shah, N. Dynamic modeling of green algae cultivation in a photobioreactor for sustainable biodiesel production. Biotechnol. Bioeng. 2018, 115, 359–370. [Google Scholar] [CrossRef]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Sci. Rep. 2019, 9, 13935. [Google Scholar] [CrossRef]
- Ratomski, P.; Hawrot-Paw, M.; Koniuszy, A. Utilisation of CO2 from Sodium Bicarbonate to Produce Chlorella vulgaris Biomass in Tubular Photobioreactors for Biofuel Purposes. Sustainability 2021, 13, 9118. [Google Scholar] [CrossRef]
- Peng, Y.-Y.; Gao, F.; Yang, H.-L.; Wu, H.-W.-J.; Li, C.; Lu, M.-M.; Yang, Z.-Y. Simultaneous removal of nutrient and sulfonamides from marine aquaculture wastewater by concentrated and attached cultivation of Chlorella vulgaris in an algal biofilm membrane photobioreactor (BF-MPBR). Sci. Total Environ. 2020, 725, 138524. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yang, Z.-Y.; Zhao, Q.-L.; Chen, D.-Z.; Li, C.; Liu, M.; Yang, J.-S.; Liu, J.-Z.; Ge, Y.-M.; Chen, J.-M. Mixotrophic cultivation of microalgae coupled with anaerobic hydrolysis for sustainable treatment of municipal wastewater in a hybrid system of anaerobic membrane bioreactor and membrane photobioreactor. Bioresour. Technol. 2021, 337, 125457. [Google Scholar] [CrossRef]
- Huang, K.-X.; Mao, B.-D.; Lu, M.-M.; Chen, D.-Z.; Qiu, J.; Gao, F. Effect of external acetate added in aquaculture wastewater on mixotrophic cultivation of microalgae, nutrient removal, and membrane contamination in a membrane photobioreactor. J. Environ. Manag. 2024, 349, 119391. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Simultaneous production of DHA and squalene from Aurantiochytrium sp. grown on forest biomass hydrolysates. Biotechnol. Biofuels 2019, 12, 255. [Google Scholar] [CrossRef]
- Díaz, J.P.; Inostroza, C.; Acién, F.G. Scale-up of a Fibonacci-Type Photobioreactor for the Production of Dunaliella salina. Appl. Biochem. Biotechnol. 2021, 193, 188–204. [Google Scholar] [CrossRef] [PubMed]
- Oncel, S.; Kose, A. Comparison of tubular and panel type photobioreactors for biohydrogen production utilizing Chlamydomonas reinhardtii considering mixing time and light intensity. Bioresour. Technol. 2014, 151, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Novoveská, L.; Zapata, A.K.M.; Zabolotney, J.B.; Atwood, M.C.; Sundstrom, E.R. Optimizing microalgae cultivation and wastewater treatment in large-scale offshore photobioreactors. Algal Res. 2016, 18, 86–94. [Google Scholar] [CrossRef]
- Fang, F.; Xu, R.-Z.; Huang, Y.-Q.; Wang, S.-N.; Zhang, L.-L.; Dong, J.-Y.; Xie, W.-M.; Chen, X.; Cao, J.-S. Production of polyhydroxyalkanoates and enrichment of associated microbes in bioreactors fed with rice winery wastewater at various organic loading rates. Bioresour. Technol. 2019, 292, 121978. [Google Scholar] [CrossRef]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef]
- Schädler, T.; Thurn, A.-L.; Brück, T.; Weuster-Botz, D. Continuous Production of Lipids with Microchloropsis salina in Open Thin-Layer Cascade Photobioreactors on a Pilot Scale. Energies 2021, 14, 500. [Google Scholar] [CrossRef]
- Liu, H.; Tian, Y.; Zhou, Y.; Kan, Y.; Wu, T.; Xiao, W.; Luo, Y. Multi-modular engineering of Saccharomyces cerevisiae for high-titre production of tyrosol and salidroside. Microb. Biotechnol. 2021, 14, 2605–2616. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Solsvik, J.; Wagner, J.L.; Zhang, D.; Hellgardt, K.; Park, C.W. CFD and kinetic-based modeling to optimize the sparger design of a large-scale photobioreactor for scaling up of biofuel production. Biotechnol. Bioeng. 2019, 116, 2200–2211. [Google Scholar] [CrossRef] [PubMed]
- Bani, A.; Fernandez, F.G.A.; D’Imporzano, G.; Parati, K.; Adani, F. Influence of photobioreactor set-up on the survival of microalgae inoculum. Bioresour. Technol. 2021, 320, 124408. [Google Scholar] [CrossRef]
- Cimini, D.; Iacono, I.D.; Carlino, E.; Finamore, R.; Restaino, O.F.; Diana, P.; Bedini, E.; Schiraldi, C. Engineering S. equi subsp. Zooepidemicus towards concurrent production of hyaluronic acid and chondroitin biopolymers of biomedical interest. AMB Express 2017, 7, 61. [Google Scholar] [CrossRef]
- Bianconi, M.; Ceriotti, L.; Cuzzocrea, S.; Esposito, E.; Pressi, G.; Sgaravatti, E.; Bertaiola, O.; Guarnerio, C.; Barbieri, E.; Semenzato, A.; et al. Red Carrot Cells Cultured in vitro Are Effective, Stable, and Safe Ingredients for Skin Care, Nutraceutical, and Food Applications. Front. Bioeng. Biotechnol. 2020, 8, 575079. [Google Scholar] [CrossRef]
- Abdel-Baset, A.; Matter, I.A.; Ali, M.A. Enhanced Scenedesmus obliquus Cultivation in Plastic-Type Flat Panel Photobioreactor for Biodiesel Production. Sustainability 2024, 16, 3148. [Google Scholar] [CrossRef]
- Mousavi, S.; Najafpour, G.D.; Mohammadi, M. CO2 bio-fixation and biofuel production in an air-lift photobioreactor by an isolated strain of microalgae Coelastrum sp. SM under high CO2 concentrations. Environ. Sci. Pollut. Res. 2018, 25, 30139–30150. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Show, P.L.; Lin, H.C.; Chang, C.K.; Loh, H.-S.; Lan, J.C.-W.; Ling, T.C. Preliminary integrated economic and environmental analysis of polyhydroxyalkanoates (PHAs) biosynthesis. Bioresour. Bioprocess. 2016, 3, 41. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Bernasconi, A.A.; Wiest, M.M.; Lavie, C.J.; Milani, R.V.; Laukkanen, J.A. Effect of Omega-3 Dosage on Cardiovascular Outcomes: An Updated Meta-Analysis and Meta-Regression of Interventional Trials. Mayo Clin. Proc. 2021, 96, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.L.; Lee, S.-M.; Choi, H.-J. A mini review: Photobioreactors for large scale algal cultivation. World J. Microbiol. Biotechnol. 2015, 31, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-J.; Tong, Z.-X.; Zhou, Z.-J.; Huang, D.; Wang, R.-L. A numerical model coupling bubble flow, light transfer, cell motion and growth kinetics for real timescale microalgae cultivation and its applications in flat plate photobioreactors. Algal Res. 2019, 44, 101727. [Google Scholar] [CrossRef]
- Chaisutyakorn, P.; Praiboon, J.; Kaewsuralikhit, C. The effect of temperature on growth and lipid and fatty acid composition on marine microalgae used for biodiesel production. J. Appl. Phycol. 2018, 30, 37–45. [Google Scholar] [CrossRef]
- Feng, P.; Xu, Z.; Qin, L.; Alam, M.A.; Wang, Z.; Zhu, S. Effects of different nitrogen sources and light paths of flat plate photobioreactors on the growth and lipid accumulation of Chlorella sp. GN1 outdoors. Bioresour. Technol. 2020, 301, 122762. [Google Scholar] [CrossRef]
- Putri, D.S.; Sari, D.A.; Marianah; Astuti, S.P.; Wangiyana, I.G.A.S. Effect of medium type, light intensity, and photoperiod on the growth rate of microalgae Chlorococcum sp. local isolate. IOP Conf. Ser. Earth Environ. Sci. 2021, 913, 012071. [Google Scholar] [CrossRef]
- Guzmán-Palomino, A.; Aguilera-Vázquez, L.; Hernández-Escoto, H.; García-Vite, P.M.; Martínez-Salazar, A.L. Dynamical Simulation, Sensitivity, and Productivity Analysis of a Light-Photoacclimation Model for Microalgae-Based Carbohydrate Production in Continuous Photobioreactors. Processes 2023, 11, 1866. [Google Scholar] [CrossRef]
- Gao, X.; Kong, B.; Vigil, R.D. Multiphysics simulation of algal growth in an air-lift photobioreactor: Effects of fluid mixing and shear stress. Bioresour. Technol. 2018, 251, 75–83. [Google Scholar] [CrossRef]
- Cheng, J.; Miao, Y.; Guo, W.; Song, Y.; Tian, J.; Zhou, J. Reduced generation time and size of carbon dioxide bubbles in a volute aerator for improving Spirulina sp. growth. Bioresour. Technol. 2018, 270, 352–358. [Google Scholar] [CrossRef]
- Villaró, S.; Sánchez-Zurano, A.; Ciardi, M.; Alarcón, F.J.; Clagnan, E.; Adani, F.; Morillas-España, A.; Álvarez, C.; Lafarga, T. Production of microalgae using pilot-scale thin-layer cascade photobioreactors: Effect of water type on biomass composition. Biomass Bioenergy 2022, 163, 106534. [Google Scholar] [CrossRef]
- Falinski, K.A.; Timmons, M.B.; Callan, C.; Laidley, C. Response of Tisochrysis lutea [Prymnesiophycidae] to aeration conditions in a bench-scale photobioreactor. J. Appl. Phycol. 2018, 30, 2203–2214. [Google Scholar] [CrossRef]
- Venkata Subhash, G.; Rohit, M.V.; Devi, M.P.; Swamy, Y.V.; Venkata Mohan, S. Temperature induced stress influence on biodiesel productivity during mixotrophic microalgae cultivation with wastewater. Bioresour. Technol. 2014, 169, 789–793. [Google Scholar] [CrossRef]
- Leupold, M.; Hindersin, S.; Gust, G.; Kerner, M.; Hanelt, D. Influence of mixing and shear stress on Chlorella vulgaris, Scenedesmus obliquus, and Chlamydomonas reinhardtii. J. Appl. Phycol. 2013, 25, 485–495. [Google Scholar] [CrossRef]
- Guler, B.A.; Deniz, I.; Demirel, Z.; Oncel, S.S.; Imamoglu, E. Computational fluid dynamics modelling of stirred tank photobioreactor for Haematococcus pluvialis production: Hydrodynamics and mixing conditions. Algal Res. 2020, 47, 101854. [Google Scholar] [CrossRef]
- Belohlav, V.; Zakova, T.; Jirout, T.; Kratky, L. Effect of hydrodynamics on the formation and removal of microalgal biofilm in photobioreactors. Biosyst. Eng. 2020, 200, 315–327. [Google Scholar] [CrossRef]
- Hussin, A.A.; To, S.W.; Sani, M.H.; Amin, M.F.M.; Kamaroddin, M.F. Optimisation and growth kinetic analysis of Microalgae, Arthrospira platensis in 2-L Photobioreactors. IOP Conf. Ser. Earth Environ. Sci. 2021, 842, 012036. [Google Scholar] [CrossRef]
- Wang, Y.; Castillo-Keller, M.; Eustance, E.; Sommerfeld, M. Early detection and quantification of zooplankton grazers in algal cultures by FlowCAM. Algal Res. 2017, 21, 98–102. [Google Scholar] [CrossRef]
- Deruyck, B.; Nguyen, K.H.T.; Decaestecker, E.; Muylaert, K. Modeling the impact of rotifer contamination on microalgal production in open pond, photobioreactor and thin layer cultivation systems. Algal Res. 2019, 38, 101398. [Google Scholar] [CrossRef]
- Martínez, C.; Pessi, B.A.; Bernard, O. Dynamics and productivity of microalgae in presence of predators. IFAC-PapersOnLine 2021, 54, 673–678. [Google Scholar] [CrossRef]
- Tan, K.M.; Kassim, M.A.; Ng, Z.J.; Lalung, J. Isolation and characterization of novel acidophilic microalgae from abandoned mining site area for carbohydrate biosynthesis and its kinetic growth study in photobioreactor. IOP Conf. Ser. Mater. Sci. Eng. 2020, 716, 012011. [Google Scholar] [CrossRef]
- Yuarrina, W.P.; Pradana, Y.S.; Budiman, A.; Majid, A.I.; Indarto; Suyono, E.A. Study of cultivation and growth rate kinetic for mixed cultures of local microalgae as third generation (G-3) bioethanol feedstock in thin layer photobioreactor. J. Phys. Conf. Ser. 2018, 1022, 012051. [Google Scholar] [CrossRef]
- Pruvost, J.; Le Gouic, B.; Cornet, J.-F. Kinetic Modeling of CO2 Biofixation by Microalgae and Optimization of Carbon Supply in Various Photobioreactor Technologies. ACS Sustain. Chem. Eng. 2022, 10, 12826–12842. [Google Scholar] [CrossRef]
- Wang, S.; Mukhambet, Y.; Esakkimuthu, S.; Abomohra, A.E.-F. Integrated microalgal biorefinery—Routes, energy, economic and environmental perspectives. J. Clean. Prod. 2022, 348, 131245. [Google Scholar] [CrossRef]
- Bicer, Y.; Dincer, I.; Vezina, G.; Raso, F. Impact Assessment and Environmental Evaluation of Various Ammonia Production Processes. Environ. Manag. 2017, 59, 842–855. [Google Scholar] [CrossRef]
- Ye, L.; Li, H.; Xie, K. Sustainable ammonia production enabled by membrane reactor. Nat. Sustain. 2022, 5, 787–794. [Google Scholar] [CrossRef]
- Deprá, M.C.; Severo, I.A.; dos Santos, A.M.; Zepka, L.Q.; Jacob-Lopes, E. Environmental impacts on commercial microalgae-based products: Sustainability metrics and indicators. Algal Res. 2020, 51, 102056. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category of Use | Type of Bioreactor | Species Cultivated | Volume | Reference |
|---|---|---|---|---|
| Biohydrogen production | Tubular PBR | Chlamydomonas reinhardtii strain CC124 | 0.004 m3 | [48] |
| Panel PBR | 0.004 m3 | |||
| Fishmeal alternative production | Vertical tubular-type PBR | Chlorella vulgaris FSP-E | 50 L | [39] |
| Biofuel feedstock cultivation and wastewater remediation | Floating offshore PBR | Scenedesmus spp., Chlorella spp., Cryptomonas spp., Micractinium spp., Desmodesmus spp., Chlamydomonas spp., Euglena spp., Pandorina spp., Coelastrum spp., and Geitlerinema spp. | 4.18–20.91 m3 | [49] |
| Wastewater remediation | Membrane PBR (MPBR) | Chlorella pyrenoidosa | 4.71 L | [45] |
| Anaerobic membrane bioreactor (AnMBR)-MPBR system | Chlorella pyrenoidosa | AnMBR: 1.0 L MPBR: 4.0 L | [44] | |
| Biofilm MPBR (BF-MPBR) | Chlorella vulgaris | 1.0 L | [43] | |
| Polyhydroxyalkanoate productivity in rice winery wastewater | Sequencing batch reactors | Zoogloea | 3 L | [50] |
| Scale-up productivity | Fibonacci-type photobioreactor (PBR) | Dunaliella salina | 1250 L | [47] |
| Tubular PBR | Tetraselmis sp. CTP4 | 35 m3 | [51] | |
| 100 m3 | ||||
| 8 m2 thin-layer cascade PBR | Microchloropsis salina | 55 L | [52] | |
| 50 m2 thin-layer cascade PBR | 330 L | |||
| Bioreactor | Saccharomyces cerevisiae | 5 L | [53] | |
| Photobioreactor | Chlorella vulgaris | 100 m3 | [41] | |
| Pilot-scale flat-plate PBR | Chlamydomonas reinhardtii | 120 L | [54] | |
| Cultivation | Tubular reactor | Scenedesmus almeriensis | 3 m3 | [55] |
| Raceway reactor | 20 m3 | |||
| 4 m3 | ||||
| Thin-layer reactor | 1.5 m3 | |||
| Recombinant bacteria cultivation | Bioreactor | Streptococcus equi subsp. zooepidemicus | 3 L | [56] |
| Plant cell line cultivation | Bioreactor | Red carrot R4G cell line | 50 L | [57] |
| Cultivation and productivity | Bioreactor | Aurantiochytrium sp. T66 | 1 L | [46] |
| Productivity | Tubular PBR | Chlorella vulgaris | 100 L | [42] |
| Fatty acid productivity | Plastic-type flat-panel PBR | Scenedesmus obliquus | 5 L | [58] |
| CO2 biofixation and biofuel productivity | Air-lift PBR | Coelastrum sp. SM | 3.26 L | [59] |
| Lipid productivity | Flat-plate PBR | Nannochloropsis sp. KMMCC 290 | 5 L | [1] |
| Bubble column PBR | ||||
| Air-lift PBR |
| Purpose | PBR Type | Challenge(s) | Reference(s) |
|---|---|---|---|
| Research | Fibonacci-type PBR | Smaller scaled versions of this design decreases illuminated surface area and increases the ratio of space to culture volume. | [47] |
| Industry | Fibonacci-type PBR | Productivity varies with strains’ light requirements as large scale outside designs’ light source is solar. | [47] |
| Floating offshore PBR | In listed reference, this design was utilized to cultivate polyculture, thus replicability of data is uncertain. Additionally, relayed low lipid productivity rates. | [49] | |
| Tubular PBR | For outdoor models, growth of culture is dependent on season. | [51] | |
| High lag phase/time compared to panel PBR. Higher pressure accumulation may result in lower productivity compared to panel PBR. | [48] | ||
| Vertical tubular-type PBR | This design can generate high shear stress. High aeration rate is not viable for large-scale growth. | [39] | |
| Pilot-scale flat-plate PBR | This design can generate high shear stress. | [54] | |
| Research and industry | Stir tank PBR | This design can generate high shear stress. | [1,63] |
| Horizontal tubular PBR | This design requires more space. Challenges also include gas transfer and heat transfer. | [1,63] |
| Microalgae Species | PBR Type | Growth Factor Type | Growth Requirements | Productivity | Reference |
|---|---|---|---|---|---|
| Ankistrodesmus braunii | - | Salinity | 50 mM NaCl | After six days of cultivation, lipid content reached 34.4% dry weight, an approximate 15% increase in comparison to control conditions | [31] |
| Ankistrodesmus falcatus | - | Salinity | 100 mM NaCl | After 10 days of cultivation, lipid content reached 53% dry weight, an approximate 20% increase in comparison to control conditions | |
| Chaetoceros sp. FIKU035 | - | Temperature | 25 °C | Biomass was about 600 mg/L and biomass productivity was approximately 350 mg/L·d | [65] |
| Lipid productivity reached approximately 66.73 mg/L·d, 8.77% increase compared to cultivation at 30 °C | |||||
| 30 °C | Biomass was about 777.93 mg/L and biomass productivity was approximately 388.97 mg/L·d, approximately a 30% and 11% increase, respectively, compared to 25 °C cultivation | ||||
| Lipid productivity reached approximately 61.35 mg/L·d | |||||
| Chlamydomonas reinhardtii mutant | Flat-plate PBR | Gas exchange | Airflow rate of 5.0–7.5 L/min | Increases biomass concentration by 18% | [54] |
| Chlamydomonas reinhardtii strain CC124 | Tubular PBR | pH | 7.65 | Biomass productivity was about 31.8 mg/L·h, approximately an 11% increase compared to cultivation in a panel PBR | [48] |
| Light intensity | At tube: 150 µE/m2s At tank: 400 µE/m2s | ||||
| Panel PBR | pH | 7.8 | Biohydrogen productivity was about 1.3 mL/L·h, 24% increase compared to cultivation in a tubular PBR | ||
| Light intensity | 150 µE/m2s | ||||
| Chlorella sorokiniana DOE1412 | Flat-panel air-lift PBR | pH | 6.5 | Biomass productivity was 6.51 g/d, the highest productivity value compared to higher pH conditions | [26] |
| 8 | CO2 addition was 2.01 g CO2/g biomass, the lowest out of other pH values tested | ||||
| Chlorella sp. GN1 | Tubular bubble column PBR | Light intensity | 5 cm light path, supplying higher light intensity than larger light paths | Lipid productivity reached 92.3 mg/L/d, a 13% increase compared to 10 cm light path and the largest productivity rates out of the light paths observed | [66] |
| Nitrogen supply | 0.8 g/L urea, nitrogen concentration of approximately 3 mM | Biomass productivity rate was about 345 mg/L·d, compared to the same concentration of sodium nitration and ammonium chloride a 60% and 103% increase, respectively, occurred | |||
| Nitrogen supply | Nitrogen deprived conditions: 0.01 g/L urea in growth medium | Lipid content comprised of 48.65% cells’ dry weight, a 61.6% increase compared to nitrogen sufficient conditions | |||
| Phosphorous supply | Phosphorous deprived conditions: 0.001 g/L K2HPO4·3H2O in growth medium | Lipid content comprised of 36.28% cells’ dry weight, a 20.5% increase compared to phosphorous sufficient conditions | |||
| Chlorella vulgaris | MC 1000 multi-cultivator (Photon System Instruments) | Light intensity | 150 µE/m2s | After 8 days of cultivation, biomass productivity was 0.6 g/L, a 50% increase compared to measurements at 50 µE/m2s | [30] |
| Vertical tubular PBR | CO2 source | 2.0 g/L sodium bicarbonate | Increased lipid concentrations by 8% compared to cultures without bicarbonate | [42] | |
| CO2 fixation rate was about 0.925 g/L·d, about a 4.8-fold increase compared to cultures without bicarbonate | |||||
| - | Temperature | 25 °C | Optimal conditions, wherein biomass reached 1.52 g/L | [33] | |
| pH | 8.0 | ||||
| Salinity | 30 PSU | ||||
| Light | Blue light at 499–465 nm | ||||
| Chlorella vulgaris FSP-E | Vertical tubular PBR | Nitrogen supply | 18.6 mM urea concentration | Biomass productivity reached 268.1 mg/L/d, 34% increase compared to lower urea concentrations, and protein was produced at a rate of 155.4 mg/L/d, 41% increase compared to lower urea concentrations | [39] |
| Aeration rate | 0.05 vvm | ||||
| Chlorococcum sp. | - | Light intensity | 2500–3500 lux | After five days of cultivation, optimal growth rate was achieved | [67] |
| Growth medium | Saline water as water source for growth medium | After five days of cultivation, optimal growth rate was achieved at 323 × 104 cells/mL, which was about a three-fold increase compared to seawater and aquadest | |||
| Light cycle | 24 h light period | After nine days of cultivation, optimal growth rate was achieved | |||
| Initial cell density | - | Ideal cell density is dependent on growth conditions | |||
| Coelastrum sp. SM | Air-lift PBR | CO2 supply | 12% | The highest productivity values were achieved. Biomass productivity: 0.267 g/L·d CO2 bio-fixation rate: 0.302 g/L·h Lipid content: 37.91% of cell dry weight Carbohydrate content: 58.45% of cell dry weight | [59] |
| Airflow | Approximately 0.06 vvm | ||||
| Light intensity | 6900 lux | ||||
| Light cycle | 12 h light period, 12 h dark period | ||||
| Desmodesmus sp. | MC 1000 multi-cultivator (Photon System Instruments) | Light intensity | 300 µE/m2s | After 15 days of cultivation, biomass productivity was 1.4 g/L, about a three-fold increase compared to a lower light intensity of 50 µE/m2s | [30] |
| After 8 days of cultivation, fatty acid content increased to 6.2%, about a four-fold increase compared to a lower light intensity of 50 µE/m2s | |||||
| Dunaliella salina | Fibonacci-type PBR | Light intensity | 600–995 μE/m2s | Biomass concentration was 0.96 g/L, a three-fold increase compared to cultivation in a raceway pond reactor | [47] |
| Temperature | 18.2–22.5 °C | ||||
| pH | 7.5–8.5 | ||||
| Isochrysis galbana | PBR | Light intensity | 350 μmol/m2s | Highest carbohydrate production at 48.11 gC/m3d | [68] |
| - | Temperature | 14 °C | After 10 days of cultivation, the highest docosahexaenoic acid (DHA) content was 19.55 mg/g of ash-free dry weight | [28] | |
| After five days of cultivation, the highest DHA productivity was 1.08 mg/L·d | |||||
| Nannochloropsis gaditana | Micro-PBR | Light intensity | 360 µmol photons/(m2s) | Three-fold increase in growth compared to low light conditions | [29] |
| Nannochloropsis oculata | - | Temperature | 20 °C | The maximum eicosapentaenoic acid (EPA) productivity was achieved after five days of cultivation, at 2.52 mg/L·d | [28] |
| Nannochloropsis QU130 | Air-lift flat-panel PBR | Light cycle | 24 h light period at 500 µmolhν/m2s | Biomass productivity increased by 13.6%, to 33 g/m2·d | [23] |
| Temperature | Fluctuating temperatures from 32–41 °C | ||||
| Larger cell size than continuous temperature conditions | |||||
| Nannochloropsis salina | - | Light cycle | 24 h light period | Growth rate reached 0.42 1/d, a higher value than a dark-light (12 h–12 h) cycle by 62% | [21] |
| Biomass concentration was about 0.77 g/L, a higher value than a dark–light (12 h–12 h) cycle by 43% | |||||
| Gas exchange | 1L/h of 5% CO2 supplemented air | Stationary phase occurred after 10 days of cultivation | |||
| Nitrogen supply | Nitrogen-deprived medium with 0.075 g/L NaNO3 | Lipid concentration was 63% of cells’ dry weight | |||
| 1.5 g/L NaNO3 | Growth rate increased approximately 3.5-fold | ||||
| Gas exchange | 5% CO2 supplemented air | ||||
| - | pH | 8 | Largest cell density after 21 days of cultivation at about 95.6 × 106 cells/mL compared to other tested pH conditions | [25] | |
| 9 | Second largest cell density after 21 days of cultivation at about 92.8 × 106 cells/mL compared to other tested pH conditions | ||||
| Nannochloropsis sp. FIKU036 | - | Temperature | 25 °C | Growth rate reached approximately 0.331 1/d. This value decreased while the strain was cultured at 30 °C and 35 °C by about 10% and 13%, respectively. | [65] |
| The highest biomass and its productivity rates reached about 885.35 mg/L and 293.05 mg/L·d, respectively. These values decrease in higher temperatures, decreasing by almost 50% at 35 °C | |||||
| Nannochloropsis sp. KMMCC 290 | Air-lift photobioreactor | Light intensity | 11,600 lux | Cell concentration increased by 50%, with a final concentration of 0.51 g/L | [1] |
| Lipid productivity increased by 47.7%, to 13.4 × 10−3 g/L/d | |||||
| Flat-plate photobioreactor | Lipid productivity increased by 45.7%, to 18.8 × 10−3 g/L/d | ||||
| Aeration rate | 1.0 vvm or 5.0 L/min | ||||
| Air-lift photobioreactor | Cell concentration increased by 44.1%, with a final concentration of 0.49 g/L | ||||
| Flat-plate photobioreactor | CO2 feeding | 10% CO2 at 0.5 L/min for 2 h everyday with 12 h intervals in between | Final cell concentration was 0.65 g/L | ||
| Lipid productivity reached 19.8 × 10−3 g/L/d | |||||
| Air-lift photobioreactor | |||||
| Lipid content was 31.5% | |||||
| Phaeodactylum tricornutum | - | Nutrient supply/growth medium | 8.82 mM nitrogen concentration in f/2 growth medium | After 10 days of cultivation, the highest biomass concentration reached about 2.76 g/L | [4] |
| Highest fucoxanthin content was around 2.18 mg/g of fresh weight | |||||
| Highest fucoxanthin productivity reached about 5.07 mg/L/d | |||||
| Increased fucoxanthin production to approximately 9.82 mg/L/d | |||||
| Light intensity | 20 µmol/m2/s | ||||
| Porphyridium sp. | Air-lift PBR | Airflow | 0.16 cm/s | Dry biomass concentration reached approximately 5 g/L | [69] |
| Scenedesmus incrassatulus | - | Salinity | 100 mM NaCl | After six days of cultivation, lipid content reached 37.7% dry weight, an approximate 15% increase in comparison to control conditions | [31] |
| Scenedesmus obliquus | MC 1000 multi-cultivator (Photon System Instruments) | Light intensity | 150 µE/m2s | After 8 days of cultivation, biomass productivity was 0.8 g/L, about a two-fold increase compared to productivity at 50 µE/m2s | [30] |
| 300 µE/m2s | After 15 days of cultivation, biomass productivity was 1.2 g/L, about a two-fold increase compared to productivity at 50 µE/m2s | ||||
| After 15 days of cultivation, fatty acid content increased to 11.6%, about a three-fold increase compared to productivity at 50 µE/m2s | |||||
| Plastic-type flat-panel PBR | Nitrogen supply | Nitrogen source was urea | Cells’ dry biomass was composed of 40% lipids, about a 10% increase compared to the control design | [58] | |
| Light intensity | 3000 lux | ||||
| Spirulina sp. | Photobioreactor | Shear force | Decreased bubble size (1.8 mm) and formation time (3.3 ms) in volute aerator | Average growth rate increased by 26.6% compared to a strip aerator | [70] |
| Biomass productivity increased by 50.7% compared to a strip aerator | |||||
| Tetradesmus almeriensis | Thin-layer cascade PBR | Nutrient supply/growth medium | Freshwater with fertilizer | Highest biomass productivity was 30.3 g/m2·day | [71] |
| Tetraselmis suecica FIKU032 | - | Temperature | 30 °C | Highest growth rate reached approximately 0.378 1/d | [65] |
| Highest biomass was about 978.43 mg/L and maximum biomass productivity was achieved at approximately 369.84 mg/L·d | |||||
| Tisochrysis lutea | Air-lift PBR | Airflow | 6.25 vvm | Reached the highest specific net growth rate at 3.8 L/min | [72] |
| Xanthonema hormidioides | Glass column PBR | Temperature | 20 °C | After three days of cultivation, the highest biomass productivity was achieved at 11.73 g/L | [32] |
| Nitrogen supply | 18 mM nitrogen concentration | ||||
| Temperature | 25 °C | After 18 days of cultivation, maximum lipid content occurred taking up 57.49% of cells’ dry weight | |||
| Nitrogen supply | 3 mM nitrogen concentration | ||||
| Mixed microalgae culture sourced from the Nacharam Cheruvu in India | - | Temperature | 30 °C | Increase in total lipid productivity to 24.5%, an approximate four-fold increase compared to growth phase (non-stress phase) | [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, S.; Du, Z.-Y. Exploring the Impact of Environmental Conditions and Bioreactors on Microalgae Growth and Applications. Energies 2024, 17, 5218. https://doi.org/10.3390/en17205218
Do S, Du Z-Y. Exploring the Impact of Environmental Conditions and Bioreactors on Microalgae Growth and Applications. Energies. 2024; 17(20):5218. https://doi.org/10.3390/en17205218
Chicago/Turabian StyleDo, Sally, and Zhi-Yan Du. 2024. "Exploring the Impact of Environmental Conditions and Bioreactors on Microalgae Growth and Applications" Energies 17, no. 20: 5218. https://doi.org/10.3390/en17205218
APA StyleDo, S., & Du, Z.-Y. (2024). Exploring the Impact of Environmental Conditions and Bioreactors on Microalgae Growth and Applications. Energies, 17(20), 5218. https://doi.org/10.3390/en17205218