
Spectroscopic Analysis of the Binding of Paraquat and Diquat Herbicides to Biosubstrates

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
3. Results and Discussion
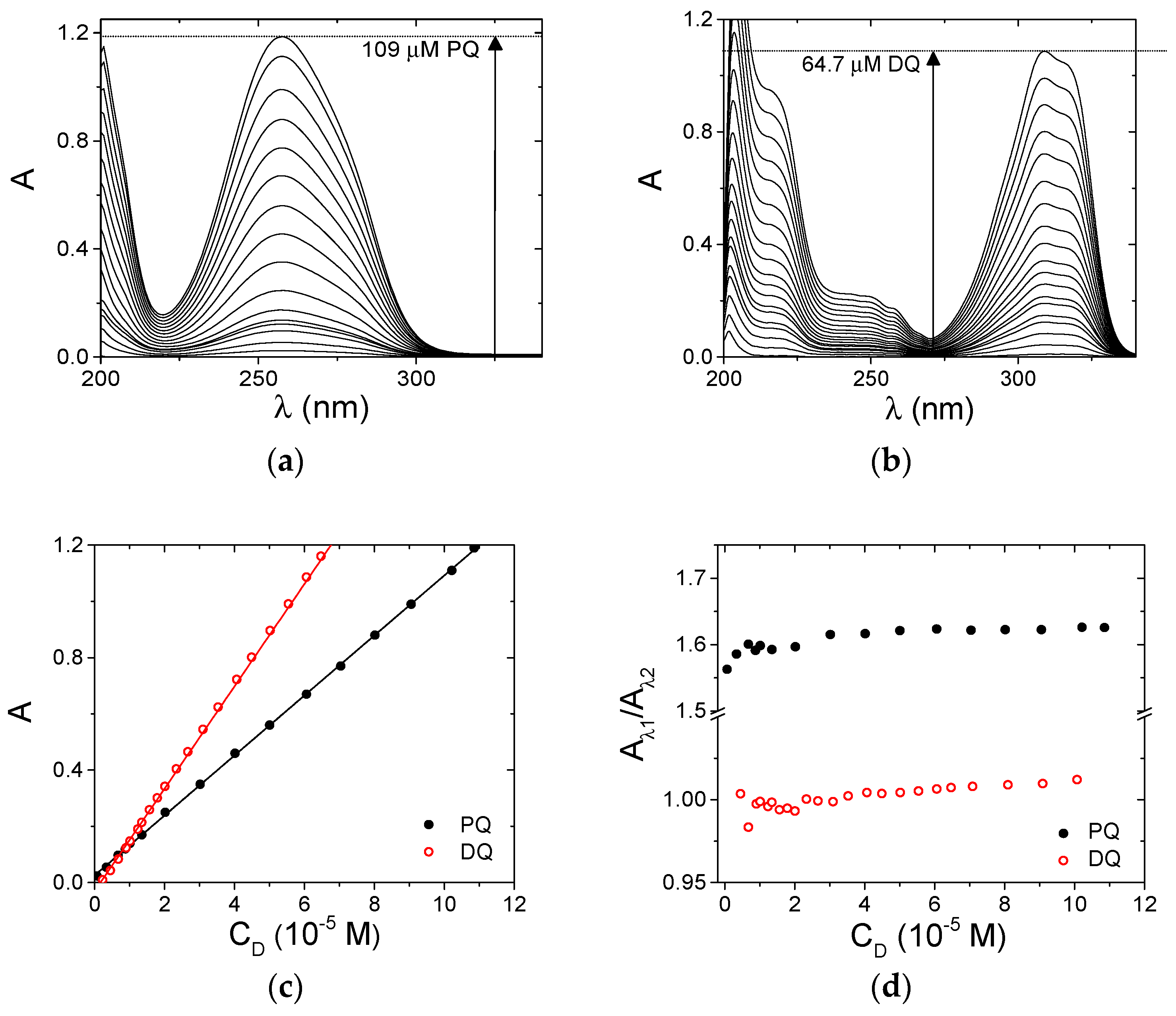
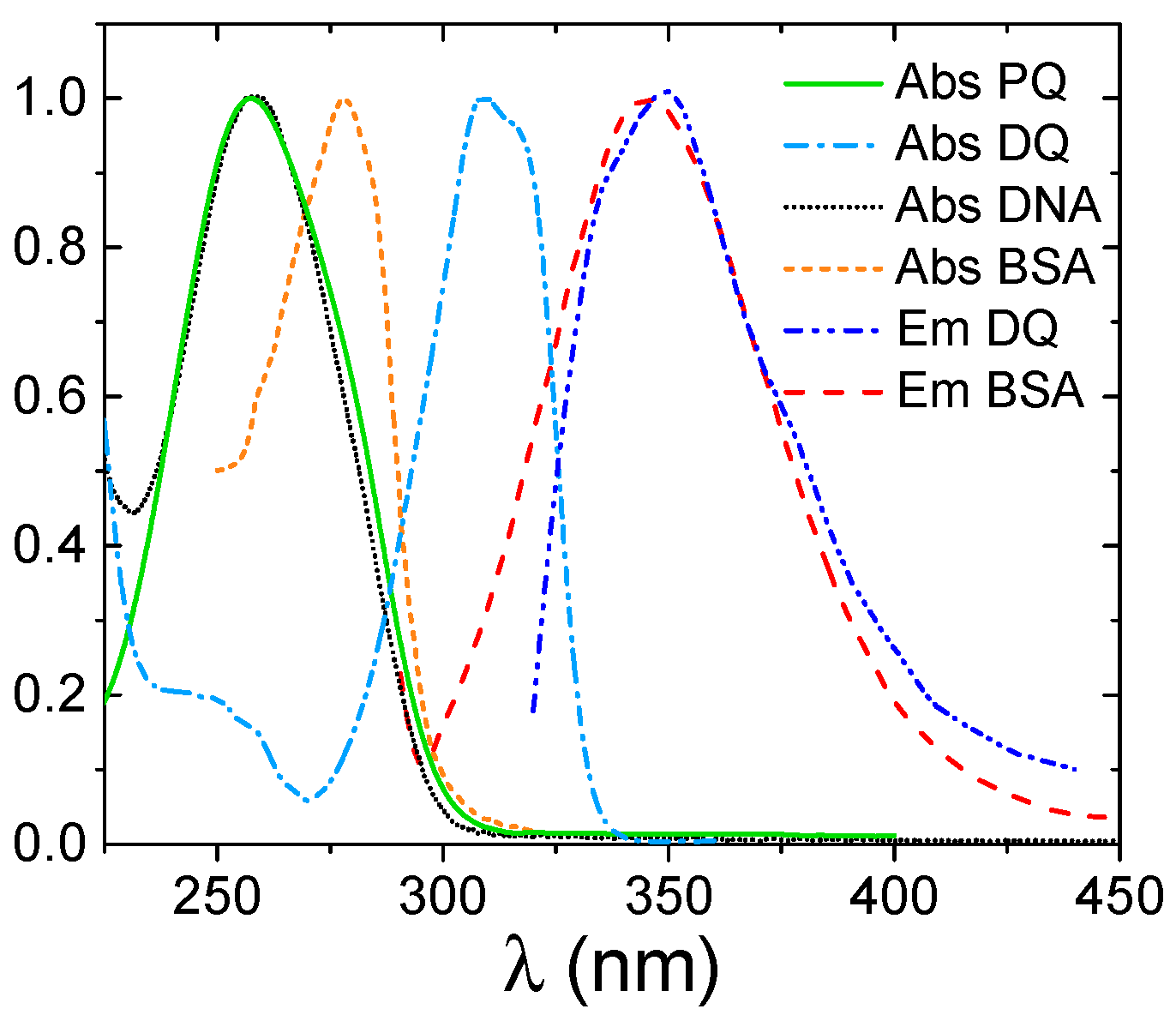
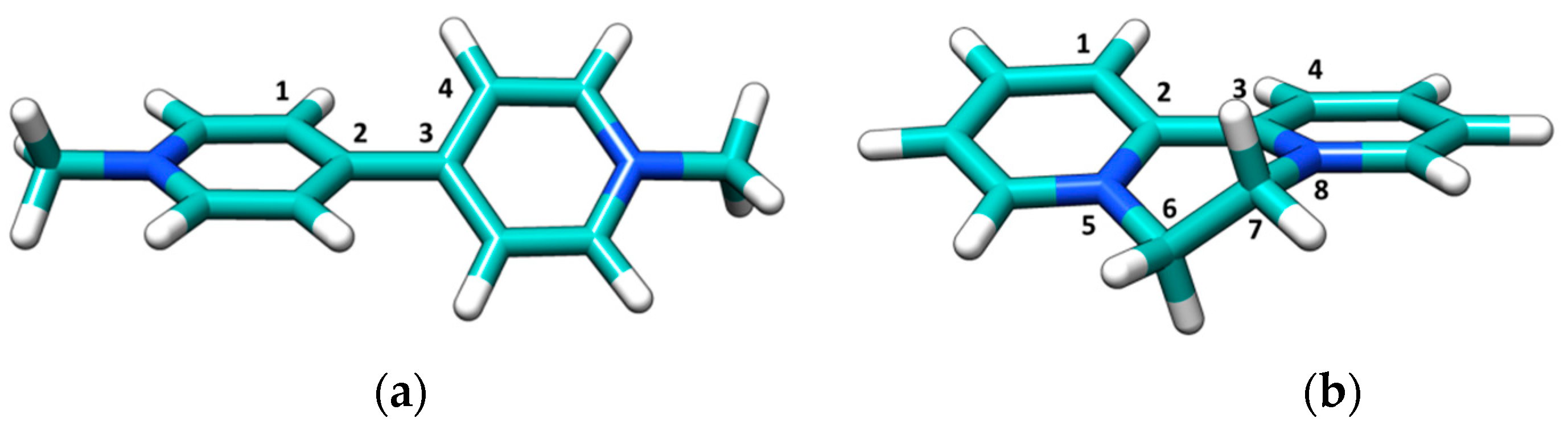
3.1. Spectroscopic Characterization
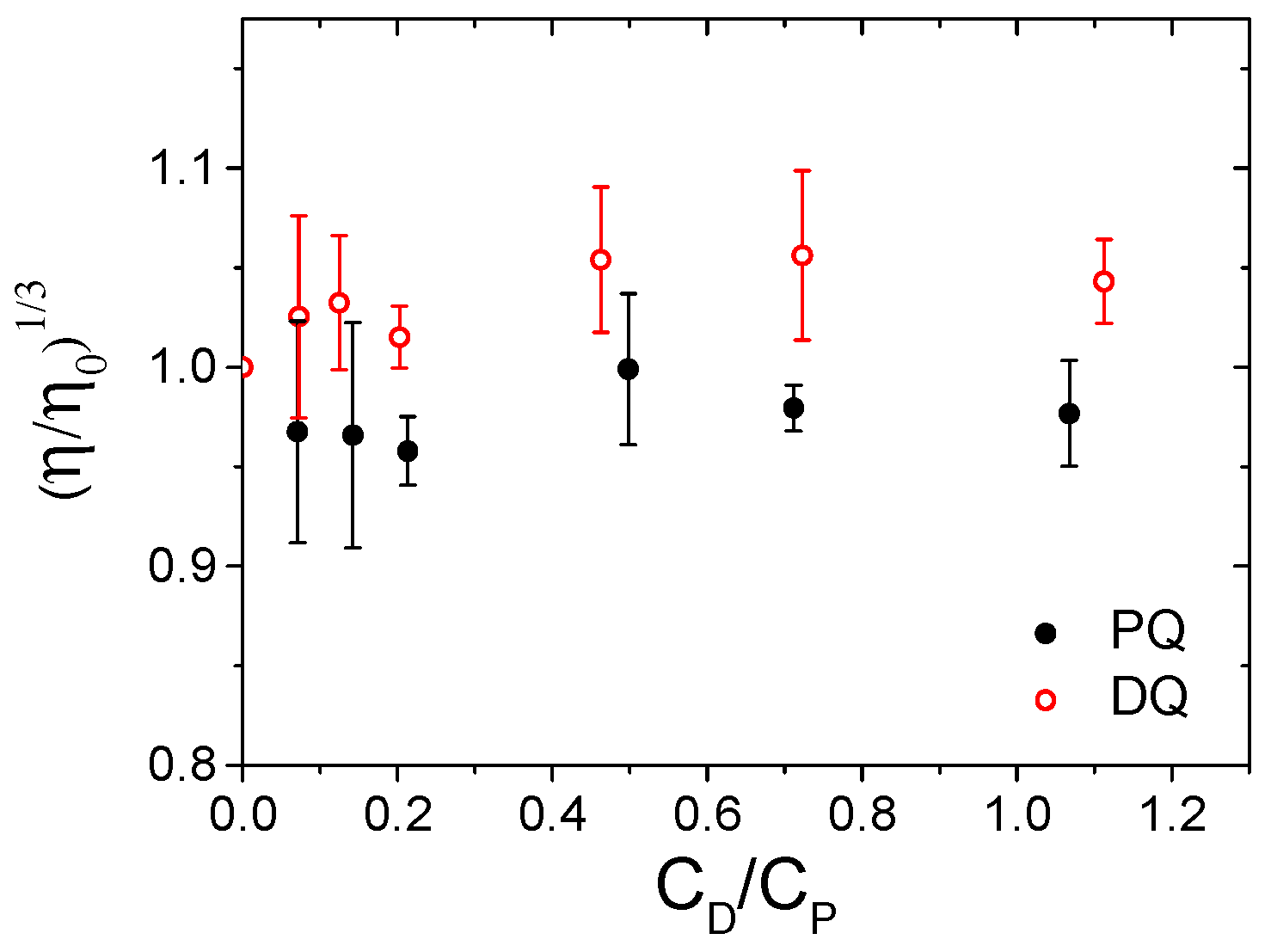
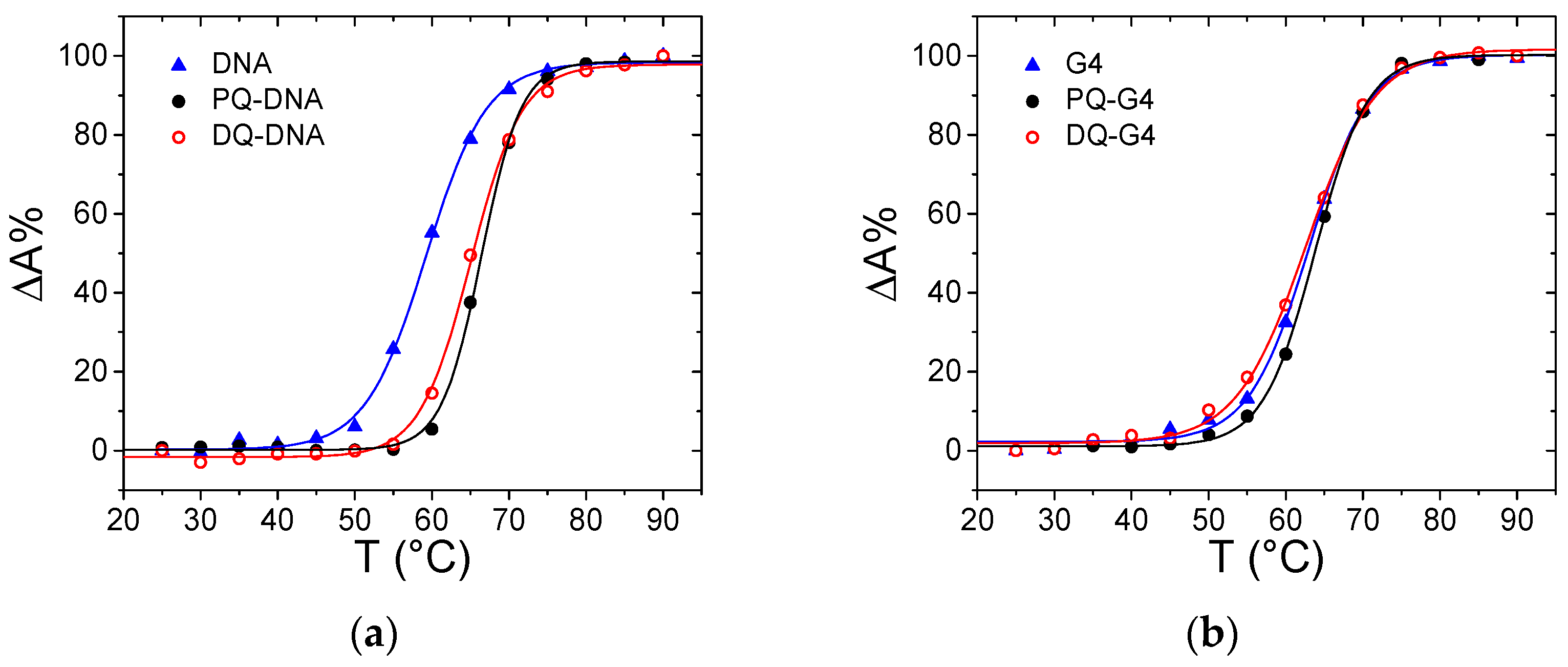
3.2. ct-DNA Binding
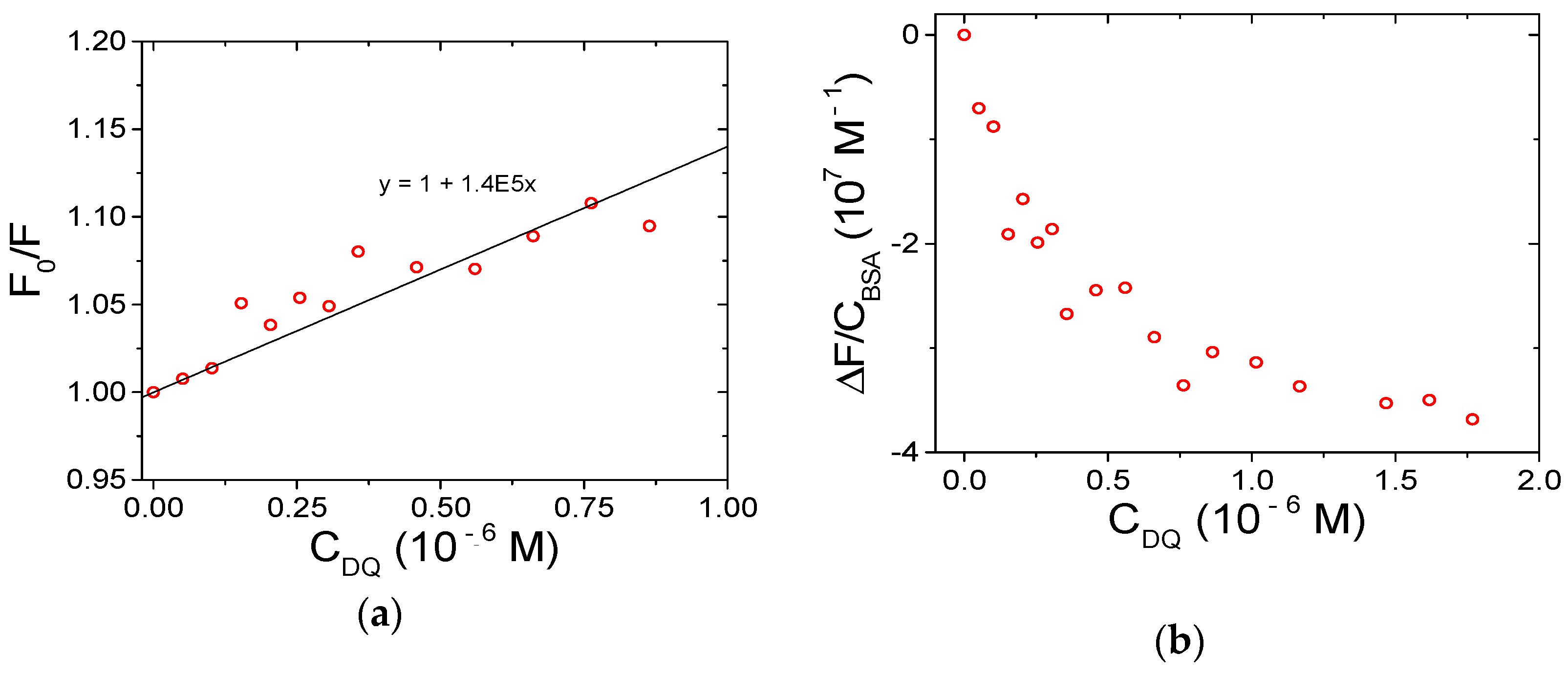
3.3. BSA Binding
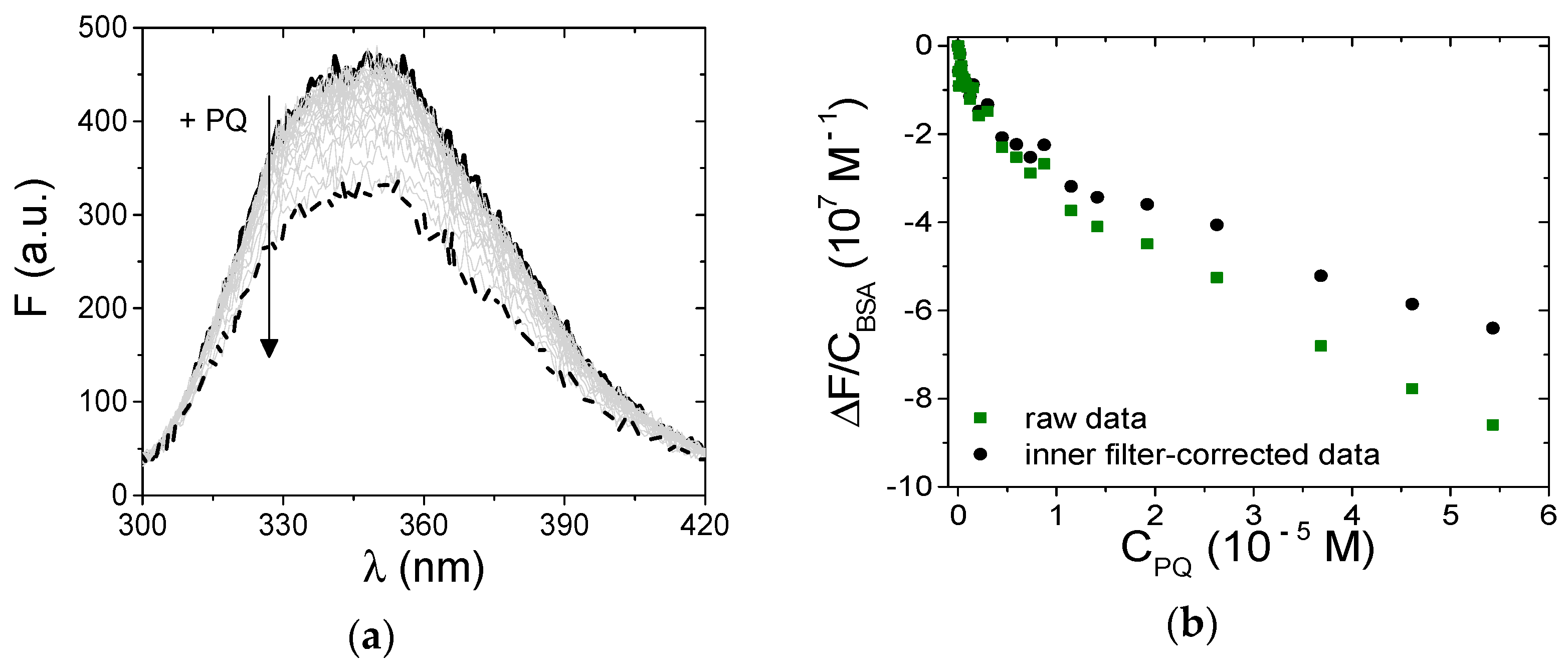
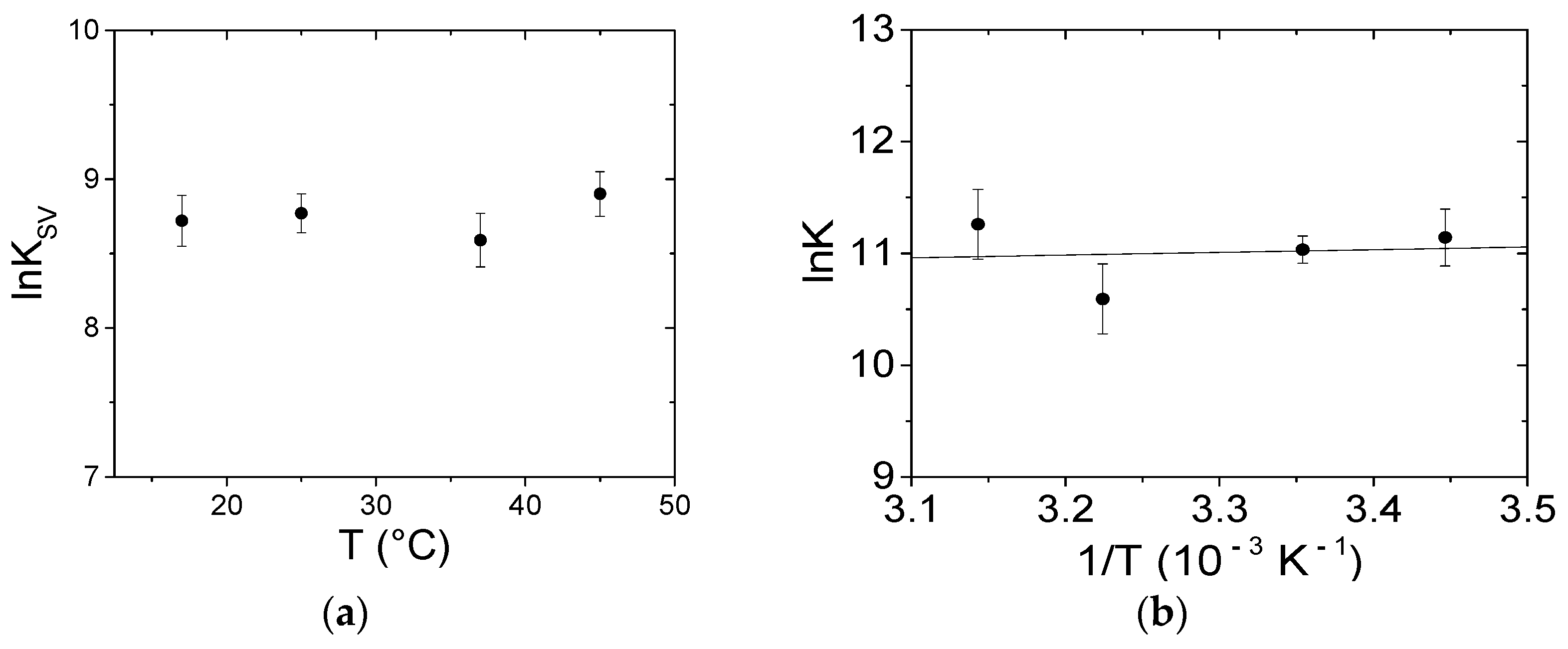
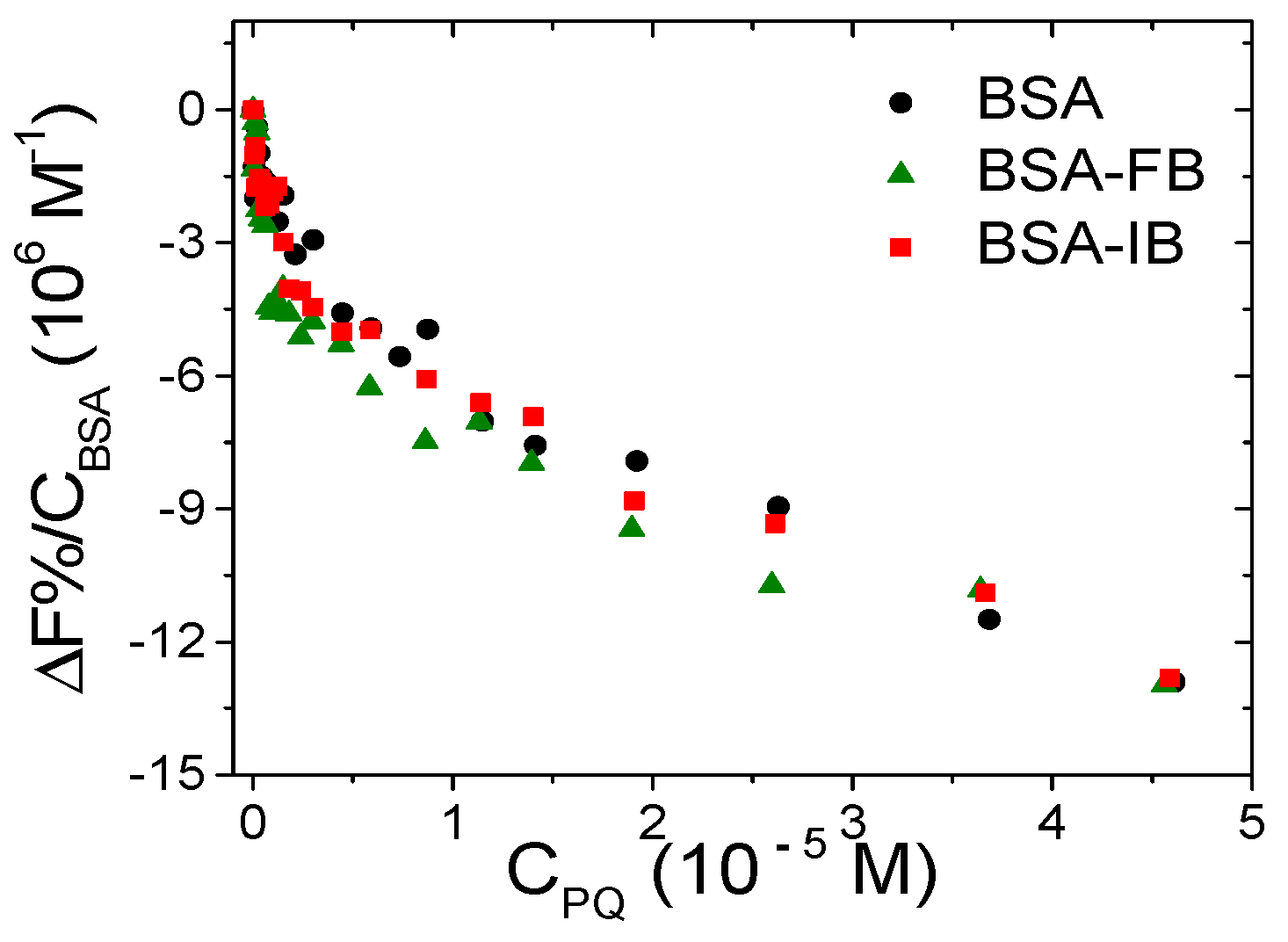
3.3.1. Spectrofluorometric Titrations
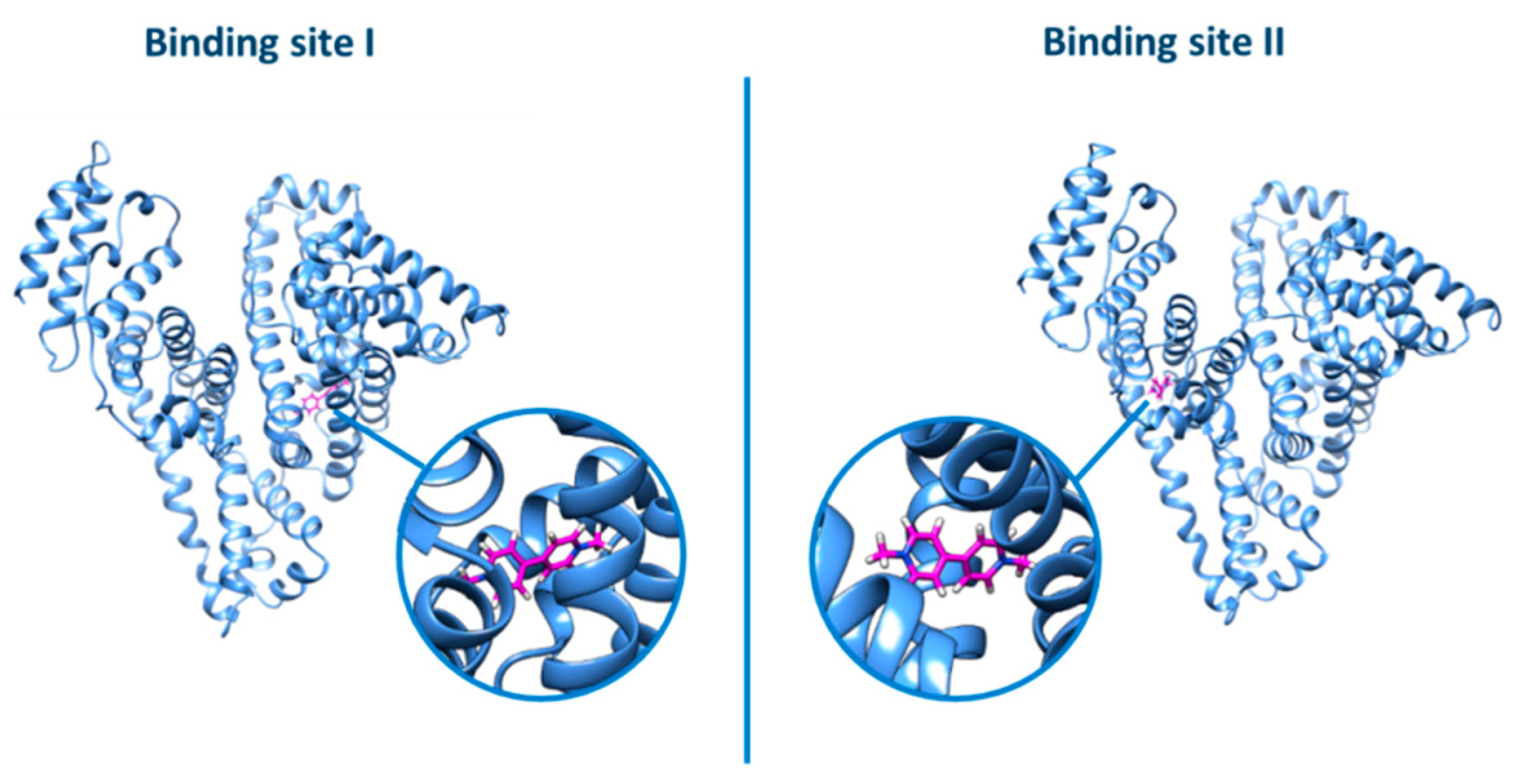
3.3.2. BSA Binding Site
3.4. Micellar Enhanced Ultra-Filtration (MEUF) Tests on Surfactants and Liposomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eddleston, M. Bipyridyl Herbicides. In Critical Care Toxicology, 2nd ed.; Springer International Publishing: Berlin, Germany, 2016; pp. 1–20. [Google Scholar]
- Lohitnavy, M.; Chitsakhon, A.; Jomprasert, K.; Lohitnavy, O.; Reisfeld, B. Development of a physiologically based pharmacokinetic model of paraquat. Proc. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. EMBS 2017, 2732–2735. [Google Scholar] [CrossRef]
- Moravčík, R.; Okuliarová, M.; Kováčová, E.; Zeman, M. Diquat-induced cytotoxicity on Vero and HeLa cell lines: Effect of melatonin and dihydromelatonin. Interdiscip. Toxicol. 2014, 7, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Mehmandost, N.; García-Valverde, M.T.; Laura Soriano, M.; Goudarzi, N.; Lucena, R.; Chamjangali, M.A.; Cardenas, S. Heracleum persicum based biosorbent for the removal of paraquat and diquat from waters. J. Environ. Chem. Eng. 2020, 8, 104481. [Google Scholar] [CrossRef]
- Conde-Cid, M.; Paradelo, R.; Fernández-Calviño, D.; Pérez-Novo, C.; Nóvoa-Múñoz, J.C.; Arias-Estévez, M. Retention of quaternary ammonium herbicides by acid vineyard soils with different organic matter and Cu contents. Geoderma 2017, 293, 26–33. [Google Scholar] [CrossRef]
- López-Blanco, R.; Moreno-González, D.; Nortes-Méndez, R.; García-Reyes, J.F.; Molina-Díaz, A.; Gilbert-López, B. Experimental and theoretical determination of pesticide processing factors to model their behavior during virgin olive oil production. Food Chem. 2018, 239, 9–16. [Google Scholar] [CrossRef]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Mireles-Ramírez, M.A.; Flores-Alvarado, L.J.; González-Usigli, H.; Sánchez-López, A.L.; Sánchez-Romero, L.; Velázquez-Brizuela, I.E.; González-Renovato, E.D.; Torres-Sánchez, E.D. Oxidative Stress and Parkinson’s Disease: Effects on Environmental Toxicology. In Free Radicals and Diseases; IntechOpen Limited: London, UK, 2016; pp. 183–209. [Google Scholar] [CrossRef]
- Fortenberry, G.Z.; Beckman, J.; Schwartz, A.; Prado, J.B.; Graham, L.S.; Higgins, S.; Lackovic, M.; Mulay, P.; Bojes, H.; Waltz, J.; et al. Magnitude and characteristics of acute paraquat- and diquat-related illnesses in the US: 1998–2013. Environ. Res. 2016, 146, 191–199. [Google Scholar] [CrossRef]
- Sha, O.; Cui, B.; Chen, X.; Liu, H.; Yao, J.; Zhu, Y. Separation and determination of paraquat and diquat in human plasma and urine by magnetic dispersive solid phase extraction coupled with high-performance liquid chromatography. J. Anal. Methods Chem. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Pan, M.; Xiang, P.; Yu, Z.; Zhao, Y.; Yan, H. Development of a high-throughput screening analysis for 288 drugs and poisons in human blood using Orbitrap technology with gas chromatography-high resolution accurate mass spectrometry. J. Chromatogr. A 2019, 1587, 209–226. [Google Scholar] [CrossRef]
- Yastrub, T.O.; Omelchuk, S.T.; Yastrub, A.M. Dermal absorption of diquat and potential occupational risk. Wiad. Lek. 2020, 73, 1459–1464. [Google Scholar] [CrossRef]
- McCuaig, L.M.; Martyniuk, C.J.; Marlatt, V.L. Morphometric and proteomic responses of early-life stage rainbow trout (Oncorhynchus mykiss) to the aquatic herbicide diquat dibromide. Aquat. Toxicol. 2020, 222, 105446. [Google Scholar] [CrossRef]
- Tajai, P.; Suriyo, T.; Rangkadilok, N.; Fedeles, B.I.; Essigmann, J.M.; Satayavivad, J. Andrographolide, an antioxidant, counteracts paraquatinduced mutagenesis in mammalian cells. Asian Pac. J. Cancer Prev. 2020, 21, 3–8. [Google Scholar] [CrossRef]
- Soares, J.J.; Rodrigues, D.T.; Gonçalves, M.B.; Lemos, M.C.; Gallarreta, M.S.; Bianchini, M.C.; Gayer, M.C.; Puntel, R.L.; Roehrs, R.; Denardin, E.L.G. Paraquat exposure-induced Parkinson’s disease-like symptoms and oxidative stress in Drosophila melanogaster: Neuroprotective effect of Bougainvillea glabra Choisy. Biomed. Pharmacother. 2017, 95, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lu, M.; Weng, J.; Wang, Z.; Sun, F.; Geng, P.; Wang, S.; Hu, L.; Gao, Z.; Wang, X.; et al. Serum metabolic changes in rats of acute paraquat poisoning treated by pirfenidone. Int. J. Clin. Exp. Med. 2017, 10, 6656–6663. [Google Scholar]
- Cao, S.; Shen, Z.; Wang, C.; Zhang, Q.; Hong, Q.; He, Y.; Hu, C. Resveratrol improves intestinal barrier function, alleviates mitochondrial dysfunction and induces mitophagy in diquat challenged piglets. Food Funct. 2019, 10, 344–354. [Google Scholar] [CrossRef]
- Llabres-Campaner, P.J.; Guijarro, L.; Giarratano, C.; Ballesteros-Garrido, R.; Zaragozá, R.J.; Aurell, M.J.; García-España, E.; Ballesteros, R.; Abarca, B. Synthesis, optical properties, and DNA interaction of new diquats based on triazolopyridines and triazoloquinolines. Chem.—A Eur. J. 2017, 23, 12825–12832. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Reiter, R.J.; Zúñiga, G.; Melchiorri, D.; Sewerynek, E.; Pablos, M.I.; Oh, C.S.; García, J.J.; Bitzer-Quintero, O.K. Genotoxicity of paraquat: Micronuclei induced in bone marrow and peripheral blood are inhibited by melatonin. Mutat. Res.—Genet. Toxicol. Environ. Mutagen. 2000, 464, 239–245. [Google Scholar] [CrossRef]
- Muangphra, P.; Kwankua, W.; Gooneratne, R. Genotoxic effects of glyphosate or paraquat on earthworm coelomocytes. Environ. Toxicol. 2014, 29, 612–620. [Google Scholar] [CrossRef]
- Kim, S.J.; Gil, H.W.; Yang, J.O.; Lee, E.Y.; Hong, S.Y. The clinical features of acute kidney injury in patients with acute paraquat intoxication. Nephrol. Dial. Transplant. 2009, 24, 1226–1232. [Google Scholar] [CrossRef]
- Mandal, S.; Pathak, M.P.; Sharma Bora, N.; Patowary, P.; Barman, P.K.; Kishor, S.; Goyary, D.; Verma, N.; Chattopadhyay, P. Determination of LCt50 of aerosolized paraquat and its pulmonary toxic implications in non-anesthetized rats. Drug. Chem. Toxicol. 2019, 42, 552–558. [Google Scholar] [CrossRef]
- Adam, A.; Smith, L.L.; Cohen, G.M. An evaluation of the redox cycling potencies of paraquat and nitrofurantoin in microsomal and lung slice systems. Biochem. Pharmacol. 1990, 40, 1533–1539. [Google Scholar] [CrossRef]
- Chowdhury, A.R.; Zielonka, J.; Kalyanaraman, B.; Hartley, R.C.; Murphy, M.P.; Avadhani, N.G. Mitochondria-targeted paraquat and metformin mediate ROS production to induce multiple pathways of retrograde signaling: A dose-dependent phenomenon. Redox Biol. 2020, 36, 101606. [Google Scholar] [CrossRef]
- Zhan, Y.; Guo, Z.; Zheng, F.; Zhang, Z.; Li, K.; Wang, Q.; Wang, L.; Cai, Z.; Chen, N.; Wu, S.; et al. Reactive oxygen species regulate miR-17-5p expression via DNA methylation in paraquat-induced nerve cell damage. Environ. Toxicol. 2020, 35, 1364–1373. [Google Scholar] [CrossRef]
- Jalil, A.S.; Reddy, S.B.; Plautz, C.Z. Cellular effects of diquat dibromide exposure: Interference with Wnt signaling and cytoskeletal development. Toxicol. Res. Appl. 2019, 3, 1–13. [Google Scholar] [CrossRef]
- Magalhães, N.; Carvalho, F.; Dinis-Oliveira, R.J. Human and experimental toxicology of diquat poisoning: Toxicokinetics, mechanisms of toxicity, clinical features, and treatment. Hum. Exp. Toxicol. 2018, 37, 1131–1160. [Google Scholar] [CrossRef]
- Babalola, O.O.; Truter, J.C.; Van Wyk, J.H. Lethal and Teratogenic Impacts of imazapyr, diquat dibromide, and glufosinate ammonium herbicide formulations using frog embryo teratogenesis assay-xenopus (FETAX). Arch. Environ. Contam. Toxicol. 2020. [Google Scholar] [CrossRef]
- Vanholder, R.; Colardyn, F.; De Reuck, J.; Praet, M.; Lameire, N.; Ringoir, S. Diquat intoxication: Report of two cases and review of the literature. Am. J. Med. 1981, 70, 1267–1271. [Google Scholar] [CrossRef]
- Niu, L.M.; Liu, Y.; Lian, K.Q.; Ma, L.; Kang, W.J. Characterization of a sensitive biosensor based on an unmodified DNA and gold nanoparticle composite and its application in diquat determination. Arab. J. Chem. 2018, 11, 655–661. [Google Scholar] [CrossRef]
- Stobiecka, M. Novel DNA-Biosensors for Studies of GMO, Pesticides and Herbicides. In State of the Art in Biosensors—Environmental and Medical Applications; IntechOpen Limited: London, UK, 2013; pp. 210–222. [Google Scholar] [CrossRef]
- Dahiya, V.; Chaubey, B.; Dhaharwal, A.K.; Pal, S. Solvent-dependent binding interactions of the organophosphate pesticide, chlorpyrifos (CPF), and its metabolite, 3,5,6-trichloro-2-pyridinol (TCPy), with Bovine Serum Albumin (BSA): A comparative fluorescence quenching analysis. Pestic. Biochem. Physiol. 2017, 139, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Cortez, C.M.; Cunha-Bastos, J.; Louro, S.R.W. Methyl parathion interaction with human and bovine serum albumin. Toxicol. Lett. 2004, 147, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Liu, S.; Cao, S.; Yin, S.; Zhou, C.; Gao, S.; Jia, C.; Ji, Y.; Liu, Y. Self-assembled bovine serum albumin nanoparticles as pesticide delivery vectors for controlling trunk-boring pests. J. Nanobiotechnol. 2020, 18, 165. [Google Scholar] [CrossRef] [PubMed]
- Jafari, F.; Moradi, S.; Nowroozi, A.; Sadrjavadi, K.; Hosseinzadeh, L.; Shahlaei, M. Exploring the binding mechanism of paraquat to DNA by a combination of spectroscopic, cellular uptake, molecular docking and molecular dynamics simulation methods. New J. Chem. 2017, 41, 14188–14198. [Google Scholar] [CrossRef]
- Gupta, S.; Kleiner, H.E.; Rogers, L.K.; Lau, S.S.; Smith, C.V. Redox stress and hepatic DNA fragmentation induced by diquat in vivo are not accompanied by increased 8-hydroxydeoxyguanosine contents. Redox Rep. 1997, 3, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, C.; Liu, W.; Zhang, X.; Zhuang, S. Evidence for DNA-diquat interaction and cytotoxicity in in vitro rat cells. Environ. Chem. Lett. 2012, 10, 35–39. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, Y.; Zhang, H.; Tang, S.; Tao, W. Human serum albumin interaction with paraquat studied using spectroscopic methods. Pestic. Biochem. Physiol. 2007, 87, 23–29. [Google Scholar] [CrossRef]
- Tunç, S.; Duman, O.; Soylu, I.; Kanci Bozoǧlan, B. Study on the bindings of dichlorprop and diquat dibromide herbicides to human serum albumin by spectroscopic methods. J. Hazard. Mater. 2014, 273, 36–43. [Google Scholar] [CrossRef]
- Moriwaki, H.; Yamada, K.; Nakanishi, H. Evaluation of the Interaction between Pesticides and a Cell Membrane Model by Surface Plasmon Resonance Spectroscopy Analysis. J. Agric. Food Chem. 2017, 65, 5390–5396. [Google Scholar] [CrossRef] [PubMed]
- Balleza, D.; Mescola, A.; Marín–Medina, N.; Ragazzini, G.; Pieruccini, M.; Facci, P.; Alessandrini, A. Complex Phase Behavior of GUVs Containing Different Sphingomyelins. Biophys. J. 2019, 116, 503–517. [Google Scholar] [CrossRef]
- Mescola, A.; Marín-Medina, N.; Ragazzini, G.; Accolla, M.; Alessandrini, A. Magainin-H2 effects on the permeabilization and mechanical properties of giant unilamellar vesicles. J. Colloid Interface Sci. 2019, 553, 247–258. [Google Scholar] [CrossRef]
- Kreutzberger, M.A.; Pokorny, A.; Almeida, P.F. Daptomycin-phosphatidylglycerol domains in lipid membranes. Langmuir 2017, 33, 13669–13679. [Google Scholar] [CrossRef]
- Macii, F.; Salvadori, G.; Bonini, R.; Giannarelli, S.; Mennucci, B.; Biver, T. Binding of model polycyclic aromatic hydrocarbons and carbamate-pesticides to DNA, BSA, micelles and liposomes. Spectrochim. Acta—Part A Mol. Biomol. Spectrosc. 2019, 223, 117313. [Google Scholar] [CrossRef]
- Biver, T.; Secco, F.; Tinè, M.R.; Venturini, M.; Bencini, A.; Bianchi, A.; Giorgi, C. Intercalation of Zn(II) and Cu(II) complexes of the cyclic polyamine Neotrien into DNA: Equilibria and kinetics. J. Inorg. Biochem. 2004, 98, 1531–1538. [Google Scholar] [CrossRef]
- Agonigi, G.; Biancalana, L.; Lupo, M.G.; Montopoli, M.; Ferri, N.; Zacchini, S.; Binacchi, F.; Biver, T.; Campanella, B.; Pampaloni, G.; et al. Exploring the anticancer potential of diiron bis-cyclopentadienyl complexes with bridging hydrocarbyl ligands: Behavior in aqueous media and in vitro cytotoxicity. Organometallics 2020, 39, 645–657. [Google Scholar] [CrossRef]
- Aydinoglu, S.; Biver, T.; Secco, F.; Venturini, M. Effects of micelle nature and concentration on the acid dissociation constants of the metal extractor PADA. Colloids Surfaces A Physicochem. Eng. Asp. 2014, 461, 303. [Google Scholar] [CrossRef][Green Version]
- Biancardi, A.; Biver, T.; Burgalassi, A.; Mattonai, M.; Secco, F.; Venturini, M. Mechanistic aspects of thioflavin-T self-aggregation and DNA binding: Evidence for dimer attack on DNA grooves. Phys. Chem. Chem. Phys. 2014, 16, 20061–20072. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.E.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16. Gaussian Inc. Wallingford CT 2016. [Google Scholar]
- Cancès, E.; Mennucci, B.; Tomasi, J. A new integral equation formalism for the polarizable continuum model: Theoretical background and applications to Isotropic and anisotropic dielectrics. J. Chem. Phys. 1997, 107, 3032–3041. [Google Scholar] [CrossRef]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum mechanical continuum solvation models. Chem. Rev. 2005, 105, 2999–3093. [Google Scholar] [CrossRef]
- Lepecq, J.B.; Paoletti, C. A fluorescent complex between ethidium bromide and nucleic acids. Physical-Chemical characterization. J. Mol. Biol. 1967, 27, 87–106. [Google Scholar] [CrossRef]
- Rocco, D.; Batchelor, L.K.; Agonigi, G.; Braccini, S.; Chiellini, F.; Schoch, S.; Biver, T.; Funaioli, T.; Zacchini, S.; Biancalana, L.; et al. Anticancer potential of diiron vinyliminium complexes. Chem.—A Eur. J. 2019, 1–17. [Google Scholar] [CrossRef]
- Franklin, R.E.; Gosling, R.G. The structure of sodium thymonucleate fibres. I. The influence of water content. Acta Crystallogr. 1953, 6, 673–677. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer International Publishing: Berlin, Germany, 2006. [Google Scholar]
- Macii, F.; Biver, T. Spectrofluorimetric analysis of the binding of a target molecule to serum albumin: Tricky aspects and tips. J. Inorg. Biochem. 2021, 216, 111305. [Google Scholar] [CrossRef] [PubMed]
- Tayeh, N.; Rungassamy, T.; Albani, J.R. Fluorescence spectral resolution of tryptophan residues in bovine and human serum albumins. J. Pharm. Biomed. Anal. 2009, 50, 107–116. [Google Scholar] [CrossRef]
- Khrapunov, S. The enthalpy-entropy compensation phenomenon. limitations for the use of some basic thermodynamic equations. Curr. Protein Pept. Sci. 2018, 19, 1088–1091. [Google Scholar] [CrossRef]
- Ryde, U. A fundamental view of enthalpy-entropy compensation. Medchemcomm 2014, 5, 1324–1336. [Google Scholar] [CrossRef]
- Dragan, A.I.; Read, C.M.; Crane-Robinson, C. Enthalpy–entropy compensation: The role of solvation. Eur. Biophys. J. 2017, 46, 301–308. [Google Scholar] [CrossRef]
- Kenji, W.; Mizutani, T.; Hideki, M.; Susumu, K. A new strategy for the design of water-soluble synthetic receptors: Specific recognition of DNA intercalators and diamines. Chem.—A Eur. J. 2003, 9, 2368–2380. [Google Scholar] [CrossRef]
- Sudlow, G.; Birkett, D.J.; Wade, D.N. The characterization of two specific drug binding sites on human serum albumin. Mol. Pharmacol. 1975, 11, 824–832. [Google Scholar] [PubMed]
- Platford, R.F. The octanol-water partitioning of some hydrophobic and hydrophilic compounds. Chemosphere 1983, 12, 1107–1111. [Google Scholar] [CrossRef]
- Tomlin, C.D.S. The Pesticide Manual, 13th ed.; British Crop Protection Council: Surrey, UK; Royal Society of Chemistry: Cambridge, UK, 2003. [Google Scholar]
- Olson, R.E.; Christ, D.D. Chapter 33. Plasma protein binding of drugs. Annu. Rep. Med. Chem. 1996, 31, 327–336. [Google Scholar] [CrossRef]
- Gou, Y.; Yang, F.; Liang, H. Designing prodrugs based on special residues of human serum albumin. Curr. Top. Med. Chem. 2016, 9, 996–1008. [Google Scholar] [CrossRef]
- Topală, T.; Bodoki, A.; Oprean, L.; Oprean, R. Bovine serum albumin interactions with metal complexes. Clujul Med. 2014, 87, 5. [Google Scholar] [CrossRef] [PubMed]












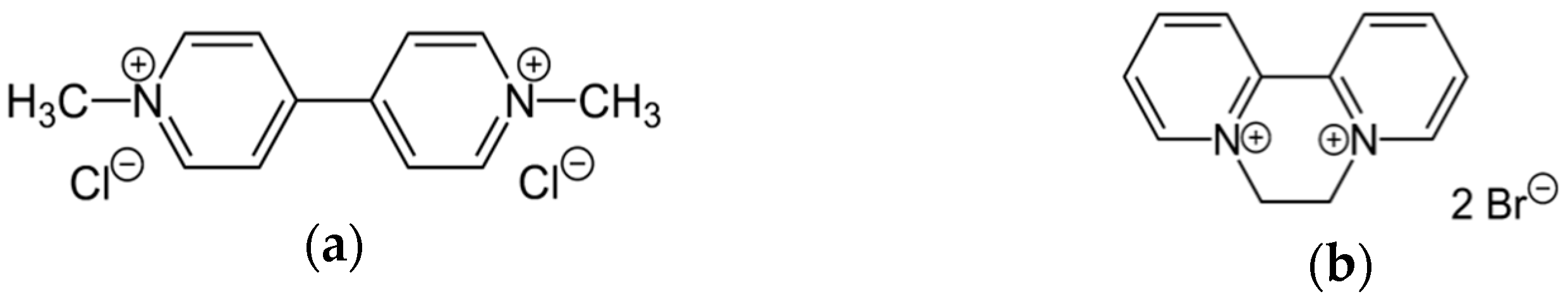{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Value (°) | |
|---|---|---|
| PQ | 1-2-3-4 | 39.4 |
| DQ | 1-2-3-4 | −22.8 |
| DQ | 3-8-7-6 | 39.9 |
| DQ | 2-5-6-7 | 39.5 |
| DQ | 5-6-7-8 | −58.3 |
| PQ | DQ | |||
|---|---|---|---|---|
| NaCl | 0.1 M | 0.5 M | 0.1 M | 0.5 M |
| SDS | 90 ± 1 | 41 ± 5 | 91 ± 1 | 34 ± 5 |
| DTAC | 8 ± 1 | 6 ± 1 | 3 ± 1 | 6 ± 1 |
| TritonX | 11 ± 2 | 18 ± 2 | 15 ± 1 | 15 ± 1 |
| POPC | 5 ± 1 | 4 ± 2 | 5 ± 2 | 6 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macii, F.; Detti, R.; Bloise, F.R.; Giannarelli, S.; Biver, T. Spectroscopic Analysis of the Binding of Paraquat and Diquat Herbicides to Biosubstrates. Int. J. Environ. Res. Public Health 2021, 18, 2412. https://doi.org/10.3390/ijerph18052412
Macii F, Detti R, Bloise FR, Giannarelli S, Biver T. Spectroscopic Analysis of the Binding of Paraquat and Diquat Herbicides to Biosubstrates. International Journal of Environmental Research and Public Health. 2021; 18(5):2412. https://doi.org/10.3390/ijerph18052412
Chicago/Turabian StyleMacii, Francesca, Rebecca Detti, Francesca Rita Bloise, Stefania Giannarelli, and Tarita Biver. 2021. "Spectroscopic Analysis of the Binding of Paraquat and Diquat Herbicides to Biosubstrates" International Journal of Environmental Research and Public Health 18, no. 5: 2412. https://doi.org/10.3390/ijerph18052412
APA StyleMacii, F., Detti, R., Bloise, F. R., Giannarelli, S., & Biver, T. (2021). Spectroscopic Analysis of the Binding of Paraquat and Diquat Herbicides to Biosubstrates. International Journal of Environmental Research and Public Health, 18(5), 2412. https://doi.org/10.3390/ijerph18052412