

Review of Marine Cyanobacteria and the Aspects Related to Their Roles: Chemical, Biological Properties, Nitrogen Fixation and Climate Change



 , ,
, , 
 ,
,  ,
,  ,
,
Abstract
1. Introduction
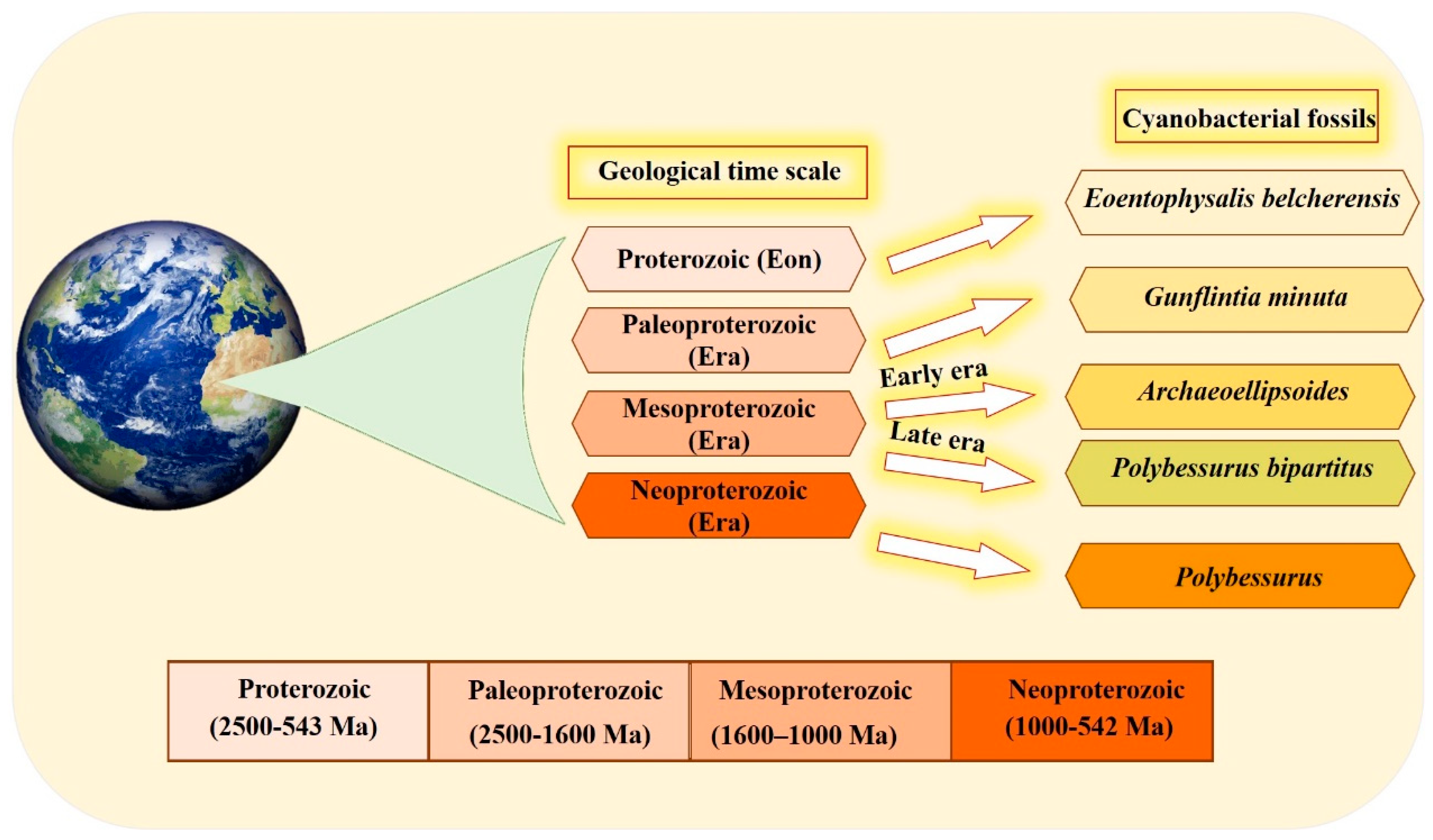
2. History and Existence
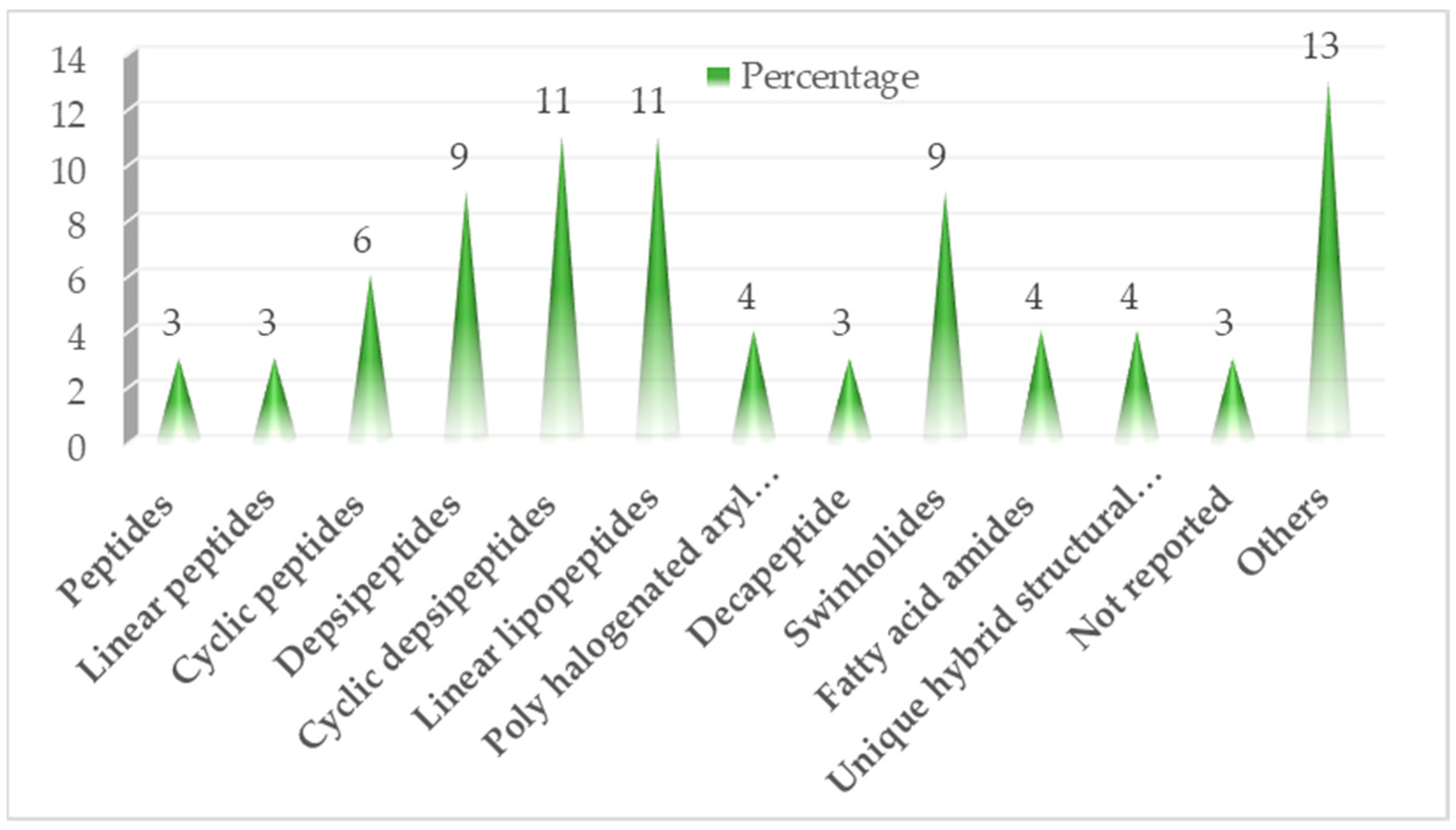
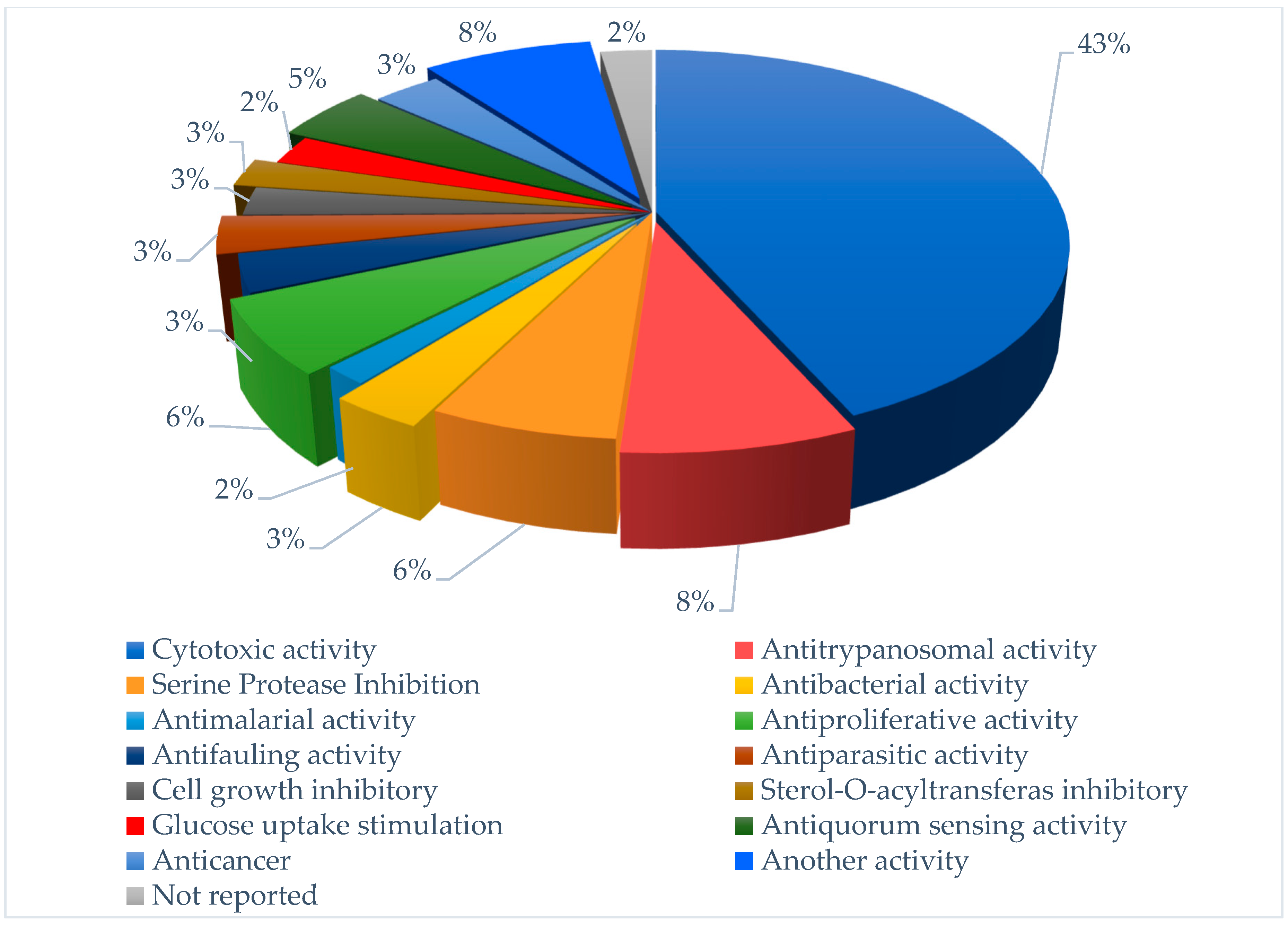
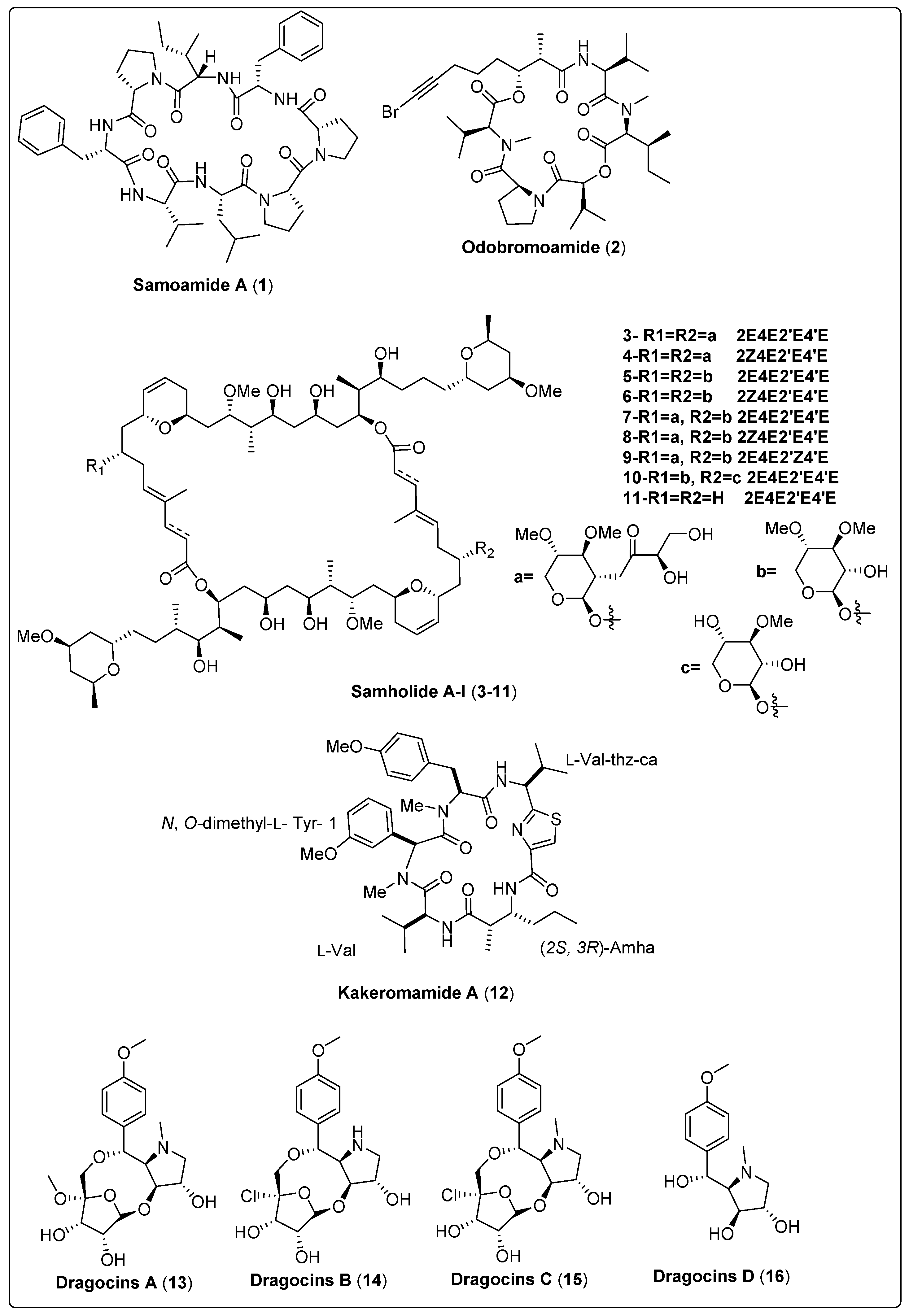
3. Bioactive Compounds Isolated from Marine Cyanobacteria
3.1. Cytotoxic Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name/Class | Source/Place and DATE | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
| Samoamide A, cyclic octapeptide (1) | cf. Symploca sp./Vatia Bay, American Samoa (July 2014) | HPLC, SPE and VLC | Cytotoxic activity Model: H460 cells and H116 cells. Assay: MTT assay IC50 = 1.1 mM for H460 cells IC50 = 4.5 mM for H116 cells PC: Doxorubicin (IC50 = 0.2 µM NC: DMSO | [48] |
| Odobromoamide, cyclo-depsipeptide (2) | Okeania sp./Odo, Okinawa Prefecture, Japan (May 2009) | HPLC and ODS-CC | Cytotoxic activity Model: HeLa S3 cells Assay: MTT assay IC50 = 0.31 µM | [49] |
| Samholide A, swinholide (3) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.17 ± 0.01 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide B, swinholide (4) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.52 ± 0.02 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide C, swinholide (5) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.21 ± 0.08 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide D, swinholide (6) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.17 ± 0.06 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide E, swinholide (7) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.17 ± 0.01 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide F, swinholide (8) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.17 ± 0.00 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide G, swinholide (9) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.21 ± 0.01 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide H, swinholide (10) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.47 ± 0.04 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Samholide I, swinholide (11) | Phormidium sp./Fagaalu Park in American Samoa, U.S. (July 2014) | VLC, SPE, RP-HPLC and HPLC | Cytotoxic activity Model: Human lung carcinoma (H-460) Assay: MTT assay IC50 = 0.91 ± 0.05 µM PC: doxorubicin (IC50 = 0.30 ± 0.02 µM) | [50] |
| Kakeromamide A, cyclic pentapeptide (12) | Moorea bouillonii/Kakeroma Island in Kagoshima, Japan | RP-HPLC | Cytotoxic activity Model: HeLa cells IC50 = 10 µM PC: Not reported | [51] |
| Dragocin A, uniquehybrid structural class (13) | Symploca-like morphology/Boca del Drago, Panama | VLC and RP-HPLC | Cytotoxic activity Model: Lung cancer cells (H460) Survival: At 3 µg/mL concentration = 23% At 30 µg/mL concentration = 10% PC: doxorubicin (EC50 = 0.0236–0.0078 µM) | [53] |
| Dragocin B, uniquehybrid structural class (14) | Symploca-like morphology/Boca del Drago, Panama | VLC and RP-HPLC | Cytotoxic activity Model: Lung cancer cells (H460) Survival: At 3 µg/mL concentration > 140% At 30 µg/mL concentration ≃ 60% PC: doxorubicin (EC50 = 0.0236–0.0078 µM) | [53] |
| Dragocin C, uniquehybrid structural class (15) | Symploca-like morphology/Boca del Drago, Panama | VLC and RP-HPLC | Cytotoxic activity Model: Lung cancer cells (H460) Survival: At 3 µg/mL concentration ≃ 136% At 30 µg/mL concentration ≃ 100% PC: doxorubicin (EC50 = 0.0236–0.0078 µM) | [53] |
| Dragocin D, uniquehybrid structural class (16) | Symploca-like morphology/Boca del Drago, Panama | VLC and RP-HPLC | Cytotoxic activity Model: Lung cancer cells (H460) Survival: At 3 µg/mL concentration ≃ 77% At 30 µg/mL concentration ≃ 15% PC: doxorubicin (EC50 = 0.0236–0.0078 mM) | [53] |
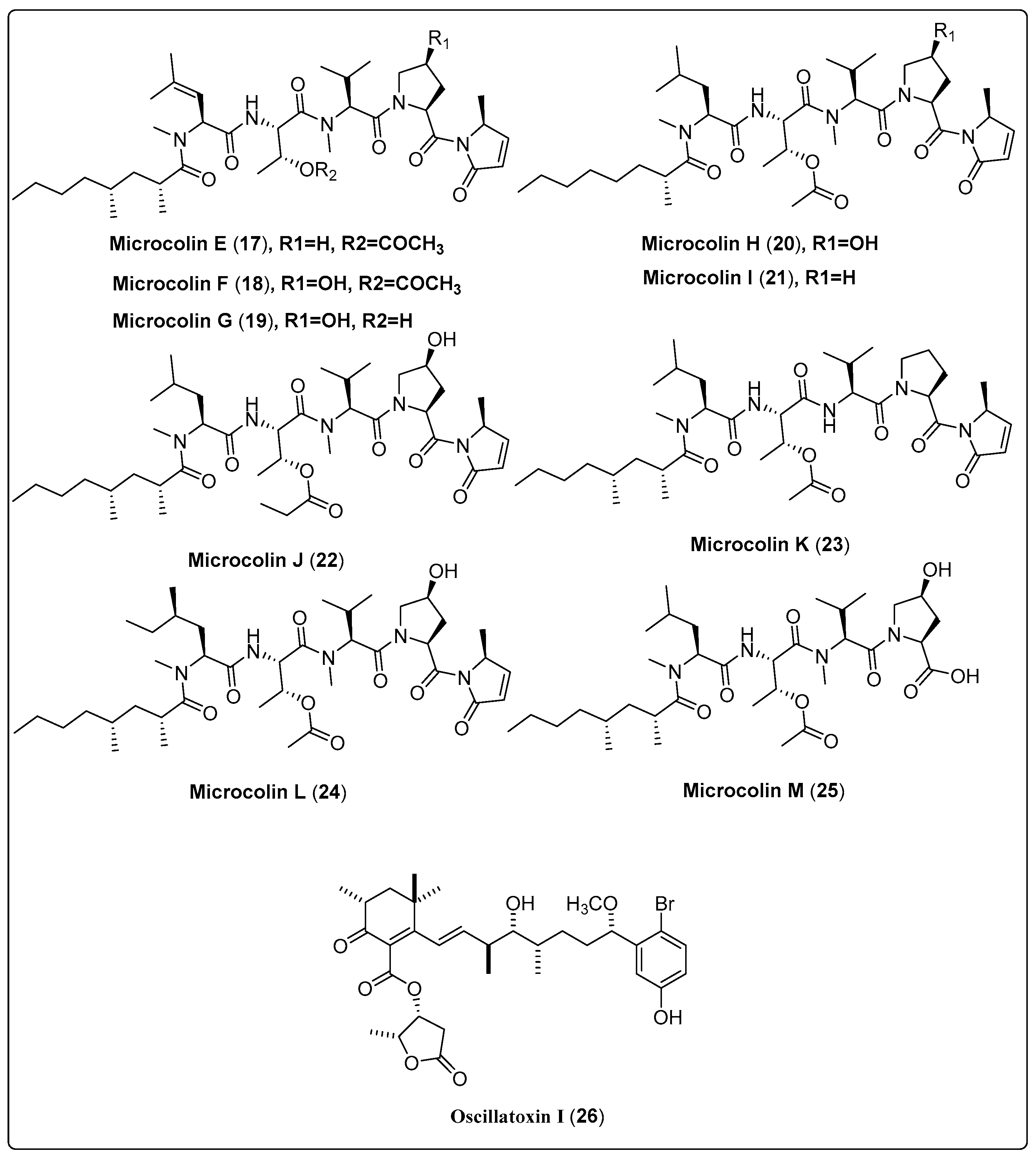
| Microcolin E, linear lipopeptide (17) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 1000 ± 20 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin F, linear lipopeptide (18) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 37 ± 4 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin G, linear lipopeptide (19) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 160 ± 20 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin H, linear lipopeptide (20) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 47 ± 5 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin I, linear lipopeptide (21) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 550 ± 50 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin J, linear lipopeptide (22) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 69 ± 10 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin K, linear lipopeptide (23) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 200 ± 50 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin L, linear lipopeptide (24) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = not tested PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Microcolin M, linear lipopeptide (25) | Moorea producens/Playa Kalki, Curaçao (September 1997) | VLC and HPLC | Cytotoxic activity Model: lung carcinoma (H460) Assay: MTT IC50 = 510 ± 60 nM PC: doxorubicin (IC50 = 180 ± 10 nM) | [52] |
| Oscillatoxin I, aplysiatoxin (26) | Moorea producens/Kuba Beach, Nakagusuku, Okinawa, Japan (July 2010) | RP-HPLC | Cytotoxic activity Model: Mouse leukemia cells (L1210) IC50 = 4.6 µg/mL Assay: XTT Concentration: 10 µg/mL | [54] |
3.2. Antiparasitic Activity
| Compound Name/Class | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
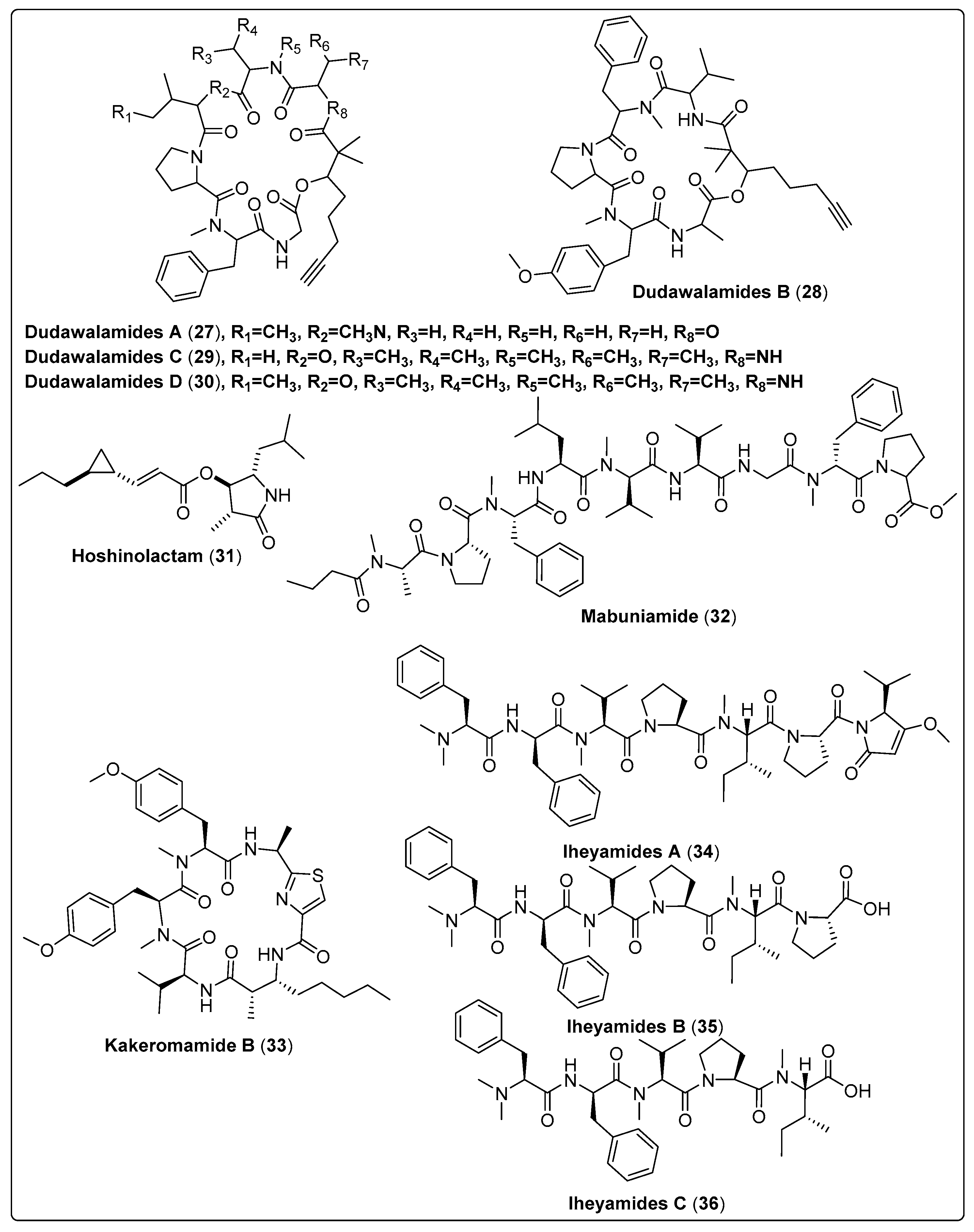
| Dudawalamide A, cyclic depsipeptide (27) | Moorea producens/Dudawali Bay in Papua New Guinea (in April 2006) | SPE, VLC and RP-HPLC | Antiparasitic activity Models: Malaria (Plasmodium falciparum) IC50: 3.6 µM PC: chloroquine leishmaniasis (Leishmania donovani) IC50: >10 µM PC: amphotericin B Chagas disease (Trypanosoma cruzi) GI: 12% µg/mL PC: benznidazole Concentrations: 10, 2, 0.4, and 0.08 µg/mL | [28] |
| Dudawalamide B, cyclic depsipeptide (28) | Moorea producens/Dudawali Bay in Papua New Guinea (in April 2006) | SPE, VLC and RP-HPLC | Antiparasitic activity Models: Malaria (Plasmodium falciparum) IC50: 8.0 µM PC: chloroquine leishmaniasis (Leishmania donovani) IC50: >10 µM PC: amphotericin B Chagas disease (Trypanosoma cruzi) GI: 7% µg/mL PC: benznidazole Concentrations: 10, 2, 0.4, and 0.08 µg/mL | [28] |
| Dudawalamide C, cyclic depsipeptide (29) | Moorea producens/Dudawali Bay in Papua New Guinea (in April 2006) | SPE, VLC and RP-HPLC | Antiparasitic activity Models: Malaria (Plasmodium falciparum) IC50: 10 µM PC: chloroquine leishmaniasis (Leishmania donovani) IC50 = Not tested PC: amphotericin B Chagas disease (Trypanosoma cruzi) GI = Not tested PC: benznidazole Concentrations: 10, 2, 0.4, and 0.08 µg/mL | [28] |
| Dudawalamide D, cyclic depsipeptide (30) | Moorea producens/Dudawali Bay in Papua New Guinea (in April 2006) | SPE, VLC and RP-HPLC | Antiparasitic activity Models: Malaria (Plasmodium falciparum) IC50: 3.5 µM PC: chloroquine leishmaniasis (Leishmania donovani) IC50: 2.6 µM PC: amphotericin B Chagas disease (Trypanosoma cruzi) GI: 60 µg/mL PC: benznidazole Concentrations: 10, 2, 0.4, and 0.08 µg/mL | [28] |
| Hoshinolactam, contain cyclopropane ring and a γ-lactam ring system (31) | Not reported/Hoshino, Okinawa | RP-HPLC | Anti-trypanosomal activity Model: Trypanosoma brucei IC50: 6.1 nM PC: pentamidine (IC50: 4.7 nM) | [55] |
| Mabuniamide, lipopeptide (32) | Okeania sp./Odo, Okinawa, Japan (May 2018) | HPLC | Antimalarial activity Model: P. falciparium (3D7 clone) IC50: 1.4 ± 0.2 μM PC: chloroquine (IC50: 7.6 ± 0.5 nM) | [56] |
| Kakeromamide B, cyclic peptide (33) | Moorea producens/Tuvuca Island in Fiji (September 2007) | VLC, SPE and HPLC | Antimalarial activity Model: Asexual blood-stage Plasmodium falciparum (EC50 = 0.89 µM) PC: Atovaquone (EC50 = 0.0061 µM) Liver-stage P. berghei (EC50 = 1.1 µM) PC: Atovaquone (EC50: <0.00028 µM) | [57] |
| Iheyamide A, linear peptide (34) | Dapis sp./Noho Island, Iheya Village, Okinawa, Japan (August 2019) | RP-HPLC | Anti-trypansomal Model: Trypanosoma brucei rhodesiense IC50:1.5 μM PC: Pentamide (IC50: 0.005 μM) Trypanosoma brucei brucei IC50: 1.5 μM PC: Pentamide (IC50: 0.001 μM) | [58] |
| Iheyamide B, linear peptide (35) | Dapis sp./Noho Island, Iheya Village, Okinawa, Japan (August 2019) | RP-HPLC | Anti-trypansomal Model: T. brucei rhodesiense IC50: >20 μM PC: Pentamide (IC50: 0.005) T. b. brucei IC50: >20 μM PC: Pentamide (IC50: 0.001) | [58] |
| Iheyamide C, linear peptide (36) | Dapis sp./Noho Island, Iheya Village, Okinawa, Japan (August 2019) | RP-HPLC | Anti-trypansomal Model: T. b. rhodesiense IC50: >20 μM PC: Pentamide (IC50: 0.005) T. b. brucei IC50: >20 μM PC: Pentamide (IC50: 0.001) | [58] |
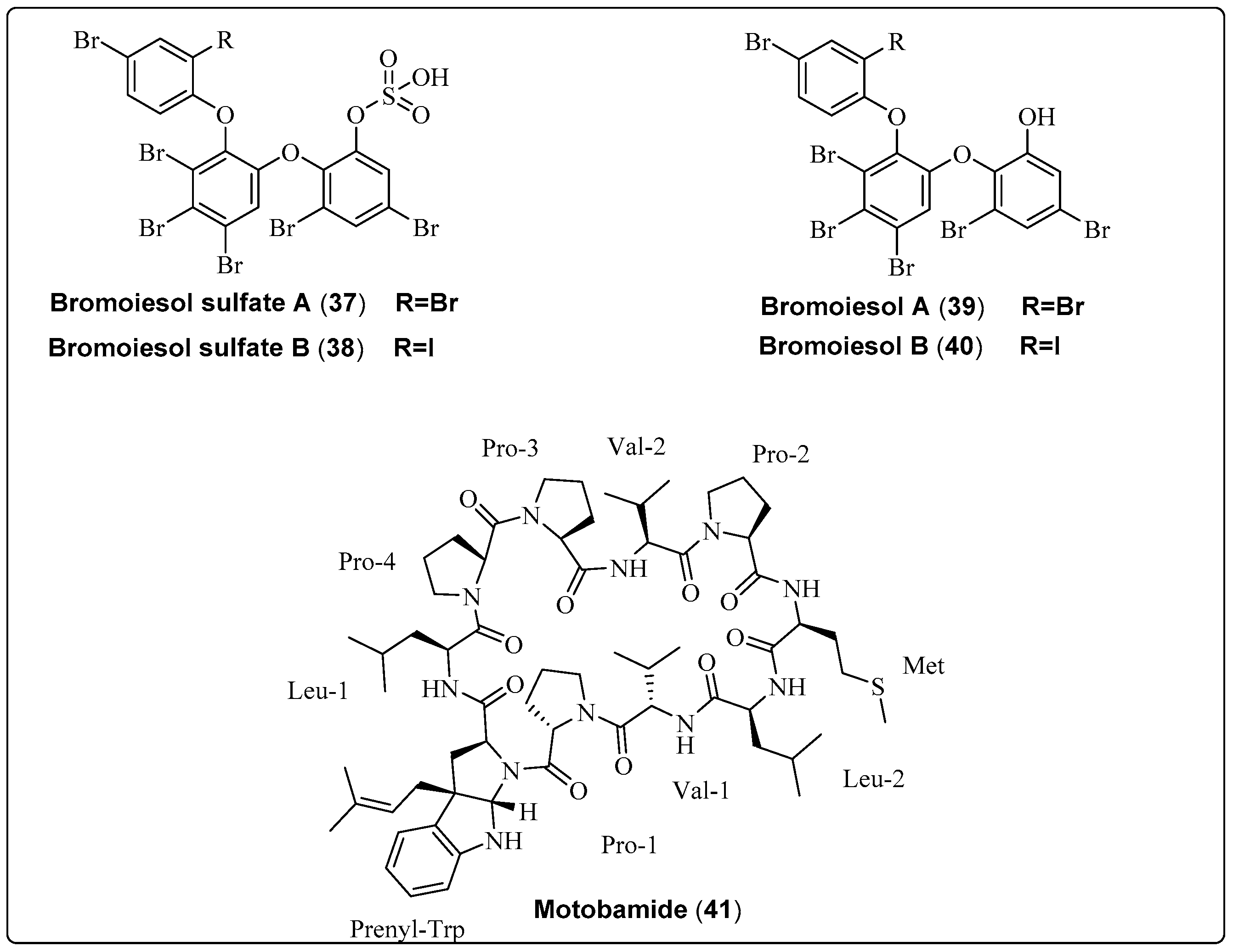
| Bromoiesol sulfate A, polyhalogenated aryl sulfate (37) | Salileptolyngbya sp./Ie-Island, Okinawa, Japan (September 2020) | HPLC | Anti-trypansomal activity Model: bloodstream Trypanosoma brucei Rhodesiense Assay: Anti-trypansomal assay IC50: 8.8 ± 1.3 μM PC: NR NC: NR | [59] |
| Bromoiesol sulfate B, polyhalogenated aryl sulfate (38) | Salileptolyngbya sp./Ie-Island, Okinawa, Japan (September 2020) | HPLC | Anti-trypansomal activity Model: bloodstream Trypanosoma brucei Rhodesiense Assay: Anti-trypansomal assay IC50: 7.9 ± 1.8 μM PC: NR NC: NR | [59] |
| Bromoiesol A, polyhalogenated aryl (39) | Salileptolyngbya sp./Ie-Island, Okinawa, Japan (September 2020) | HPLC | Anti-trypansomal activity Model: bloodstream Trypanosoma brucei Rhodesiense Assay: Anti-trypansomal assay IC50: 1.2 ± 0.1 μM PC: NR NC: NR | [59] |
| Bromoiesol B, polyhalogenated aryl (40) | Salileptolyngbya sp./Ie-Island, Okinawa, Japan (July 2020) | HPLC | Anti-trypansomal activity Model: bloodstream Trypanosoma brucei Rhodesiense Assay: Anti-trypansomal assay IC50: 0.70 ± 0.23 μM PC: NR NC: NR | [59] |
| Motobamide, cyclic peptide (41) | Leptolyngbya sp./Bise, Okinawa Island, Okinawa Prefecture, Japan (April 2018) | HPLC | Anti-trypansomal activity Trypanosoma brucei rhodesiense strains IL-1501 Assay: Anti-trypansomal Model: bloodstream IC50: 2.3 μM | [60] |
3.3. Serine Proteases Inhibitory Effect
| Compound Name/Class | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
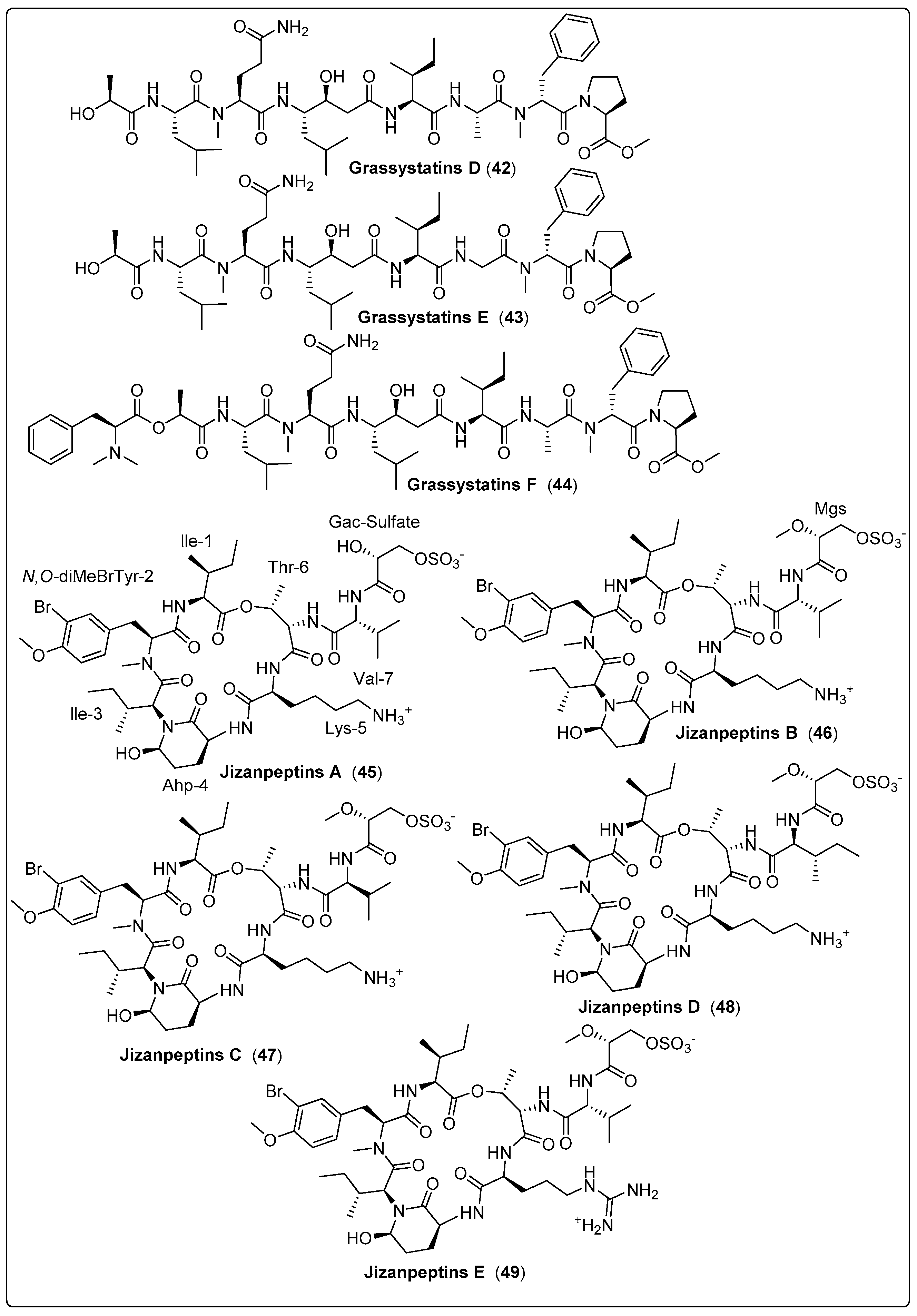
| Grassystatin D, peptide (42) | VPG 14-61/Cetti Bay, Guam (June 2014) | SPE and RP-HPLC | Protease inhibitory activity Assays: Cell viability and inhibition assay Model: Breast cancer cell line (MDA-MB-231) Cathepsin D IC50: 2000 nM Cathepsin E IC50: 30 nM NC: DMSO | [61] |
| Grassystatin E, peptide (43) | VPG 14-61/Cetti Bay, Guam (June 2014) | SPE and RP-HPLC | Protease inhibitory activity Assays: Cell viability and inhibition assay Model: Breast cancer cell line (MDA-MB-231) Cathepsin D IC50: 900 nM Cathepsin E IC50: 5 nM NC: DMSO | [61] |
| Grassystatin F, peptide (44) | VPG 14-61/Cetti Bay, Guam (June 2014) | SPE and RP-HPLC | Protease inhibitory activity Assays: Cell viability and inhibition assay Model:Breast cancer cell line (MDA-MB-231) Cathepsin D IC50: 50 nM Ccathepsin E IC50: 0.5 nM NC: DMSO | [61] |
| Jizanpeptin A, depsipeptide (45) | Symploca sp./Red Sea, coast of Jizan, Saudi Arabia (2013) | HPLC, NP-VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Trypsin IC50: 160 ± 30 nM Chymotrypsin IC50: >10,000 nM | [62] |
| Jizanpeptin B, depsipeptide (46) | Symploca sp./Red Sea, coast of Jizan, Saudi Arabia (2013) | HPLC, NP-VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Trypsin IC50: 190 ± 20 nM Chymotrypsin IC50: >10,000 nM | [62] |
| Jizanpeptin C, depsipeptide (47) | Symploca sp./Red Sea, coast of Jizan, Saudi Arabia (2013) | HPLC, NP-VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Trypsin IC50: 72 ± 17 nM Chymotrypsin IC50: 1400 ± 700 nM | [62] |
| Jizanpeptin D, depsipeptide (48) | Symploca sp./Red Sea, coast of Jizan, Saudi Arabia (2013) | HPLC, NP-VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Trypsin IC50: 1000 ± 250 nM Chymotrypsin IC50: >10,000 nM | [62] |
| Jizanpeptin E, depsipeptide (49) | Symploca sp./Red Sea, coast of Jizan, Saudi Arabia (2013) | HPLC, NP-VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Trypsin IC50: 150 ± 20 nM Chymotrypsin IC50: >10,000 nM | [62] |
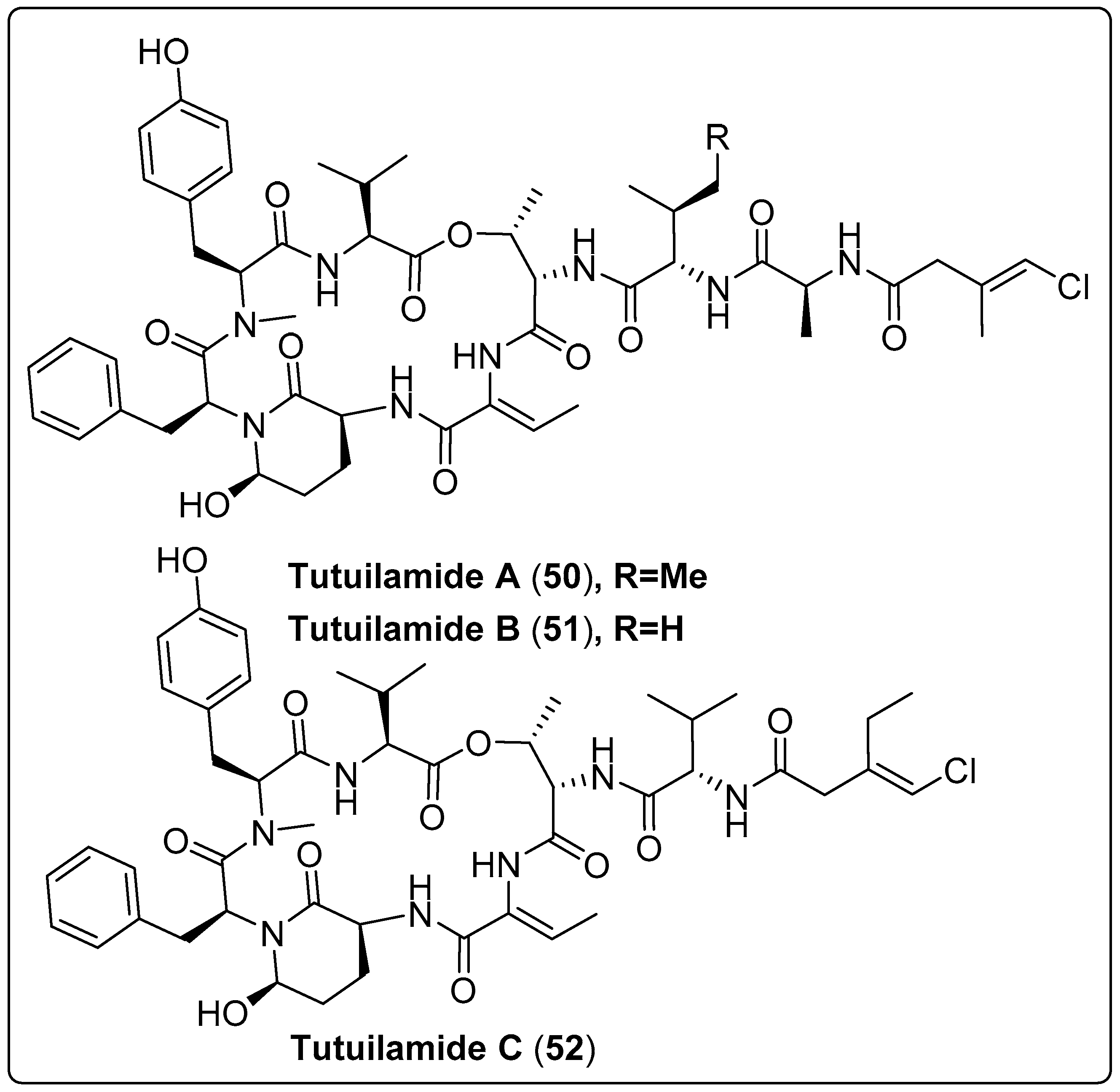
| Tutuilamide A, cyclic peptide (50) | Schizothrix sp./Island of Tutuila in American Samoa (2016) | HPLC, VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Elastase IC50: 1.18 nM Chymotrypsin IC50: 1014 nM Trypsin IC50: >20,000 nM Proteinase K IC50: 103.7 nM | [63] |
| Tutuilamides B, cyclic peptide (51) | Schizothrix sp./Island of Tutuila in American Samoa (2016) | HPLC, VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Elastase IC50: 2.05 nM Chymotrypsin IC50: 576.6 nM Trypsin IC50: >20,000 nM Proteinase K IC50 = 87.6 nM | [63] |
| Tutuilamide C, cyclic peptide (52) | Coleofasciculus sp./Island of Tutuila in American Samoa (2016) | HPLC, VLC, and SPE | Inhibitory activity of Serine Proteases Such as: Elastase IC50: 4.93 nM Chymotrypsin IC50: 542.0 nM Trypsin IC50: >20,000 nM Proteinase K IC50: 5000 nM | [63] |
3.4. Antiproliferative and Anticancer Activity
| Compound Name/Class | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
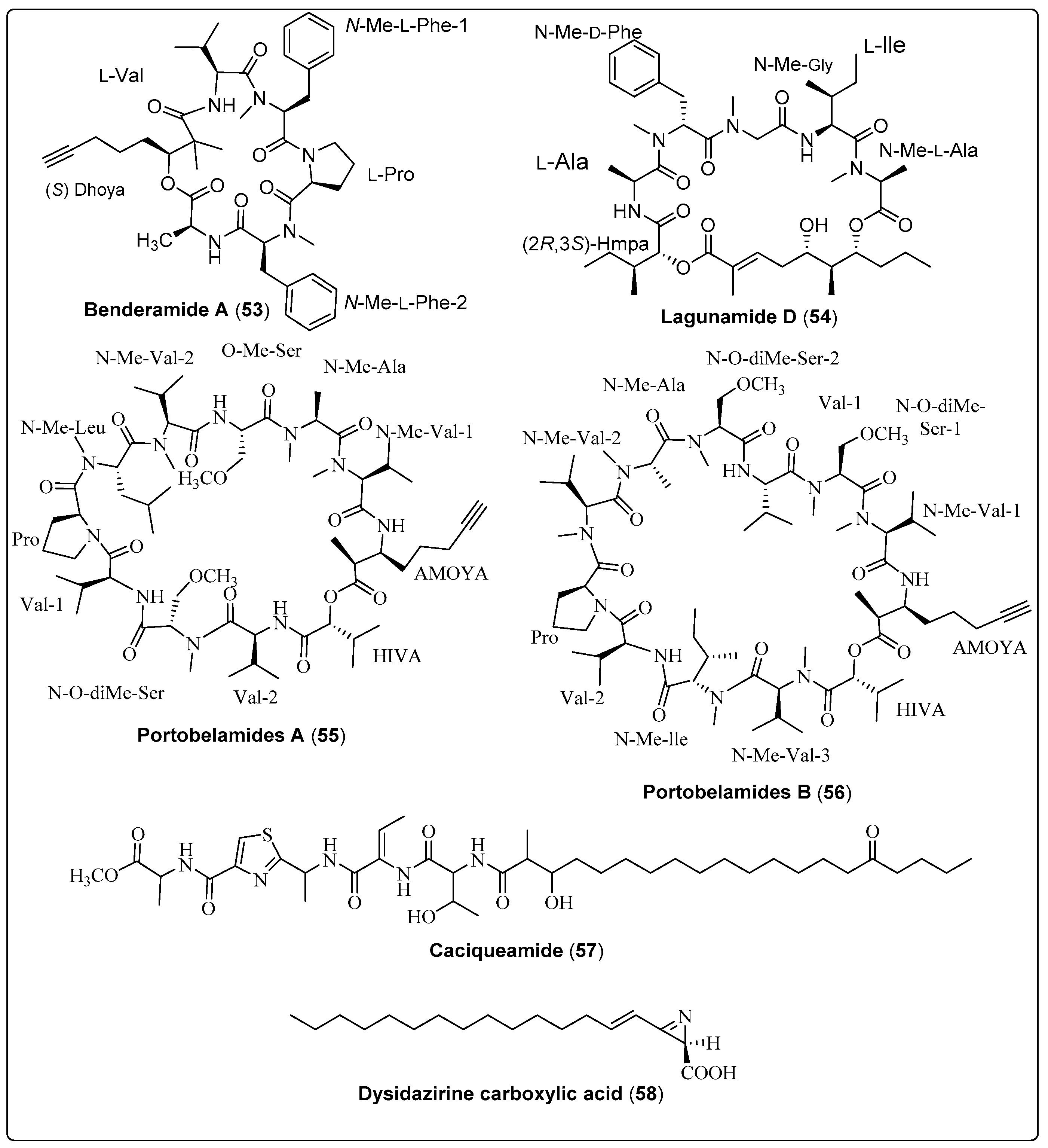
| Benderamide A, cyclic depsipeptide (53) | Lyngbya sp./John’s Island, Singapore (June 2016) | HPLC and VFC | Antiproliferative activity Model: MCF-7 breast and PA1 ovarian cancer cell lines NC: DMSO PC: Nutlin Concentration of PC: 10 µM | [65] |
| Lagunamide D, macrocyclic depsipeptide (54) | Collection of marine cyanobacteria (Dichothrix sp., Lyngbya sp., Ceramium sp. and Rivularia sp.)/Loggerhead Key in the Dry Tortugas, Florida (May 2015) | HPLC, FCL and SPE | Antiproliferative activity Model: Human lung adenocarcinoma cells (A549) Assay: MTT assay IC50: 7.1 ± 1.7 nM NC: cells + medium + solvent control | [64] |
| Portobelamide A, cyclic depsipeptide (55) | Caldora sp./Portobelo, Panama: (June 2012) | VLC and HPLC | Anticancer activity Model: Human lung Cancer Cell (H-460 cells) Cytotoxicity: (33% survival at 0.9 μM) PC: (doxorubicin) NC: (DMSO in RPMI 1640 medium) | [23] |
| Portobelamide B, cyclic depsipeptide (56) | Caldora sp./Portobelo, Panama: (June 2012) | VLC and HPLC | Anticancer activity Model: Human lung Cancer Cell (H-460 cells) Cytotoxicity: Not cytotoxic PC: (doxorubicin) NC: (DMSO in RPMI 1640 medium) | [23] |
| Caciqueamide, long chain lipopeptide (57) | Caldora sp./Portobelo, Panama (June 2012) | VLC and HPLC | Anticancer activity Model: Human lung Cancer Cell (H-460 cells) Cytotoxicity: Not cytotoxic PC: (doxorubicin) NC: (DMSO in RPMI 1640 medium) | [23] |
| Dysidazirine carboxylic acid, carboxylic acid (58) | Caldora sp./Fort Lauderdale, Florida (July 2017) | RP-HPLC | Anticancer activity Assay: Cell Viability Model: Human colon cancer cells (HCT116) IC50: 79.7 µM Positive control: Gatorbulin-1 IC50 for PC: 0.80 µM NC: 0.5% DMSO | [25] |
3.5. Anti-Quorum Sensing Activity
| Compound Name/Class | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
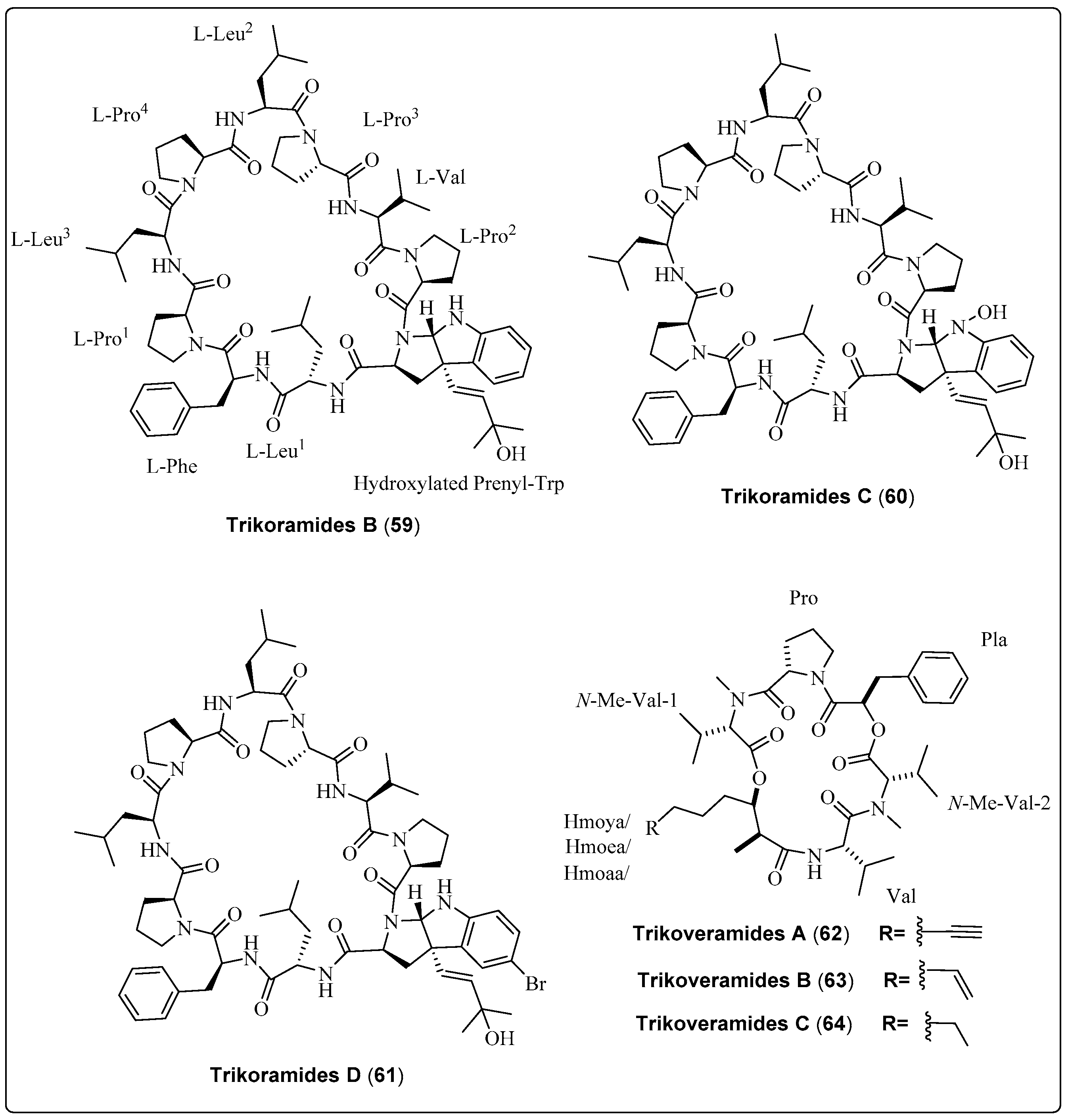
| Trikoramide B, decapeptide (59) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | RP-HPLC, VLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa PAO1 lasB-gfp and rhlA-gfp strains Assay: Anti-quorum-sensing assay IC50 for (lasB-gfp): No dose-dependent response observed IC50 for (rhlA-gfp): No dose-dependent response observed | [66] |
| Trikoramide C, decapeptide (60) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | RP-HPLC, VLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa PAO1 lasB-gfp and rhlA-gfp strains Assay: Anti-quorum-sensing assay IC50 for (lasB-gfp): No dose-dependent response observed IC50 for (rhlA-gfp): No dose-dependent response observed | [66] |
| Trikoramide D, decapeptide (61) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | RP-HPLC, VLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa PAO1 lasB-gfp and rhlA-gfp strains Assay: Anti-quorum-sensing assay IC50 for (lasB-gfp): 19.6 µM IC50 for (rhlA-gfp): 7.3 µM | [66] |
| Trikoveramide A, cyclic depsipeptide (62) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | NP-VLC, RP-HPLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa strain PAO1-lasB-gfp Assay: Anti-quorum-sensing assay Inhibition: 8% | [67] |
| Trikoveramide B, cyclic depsipeptide (63) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | NP-VLC, RP-HPLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa strain PAO1-lasB-gfp Assay: Anti-quorum-sensing assay Inhibition: 26% | [67] |
| Trikoveramide C, cyclic depsipeptide (64) | Symploca hydnoides/Trikora beach, Bintan Island (April 2018) | NP-VLC, RP-HPLC | Anti-quorum-sensing activity Model: Pseudomonas aeruginosa strain PAO1-lasB-gfp Assay: Anti-quorum-sensing assay Inhibition: 45% | [67] |
3.6. Antibacterial Activity
| Compound Name/Compound Description | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
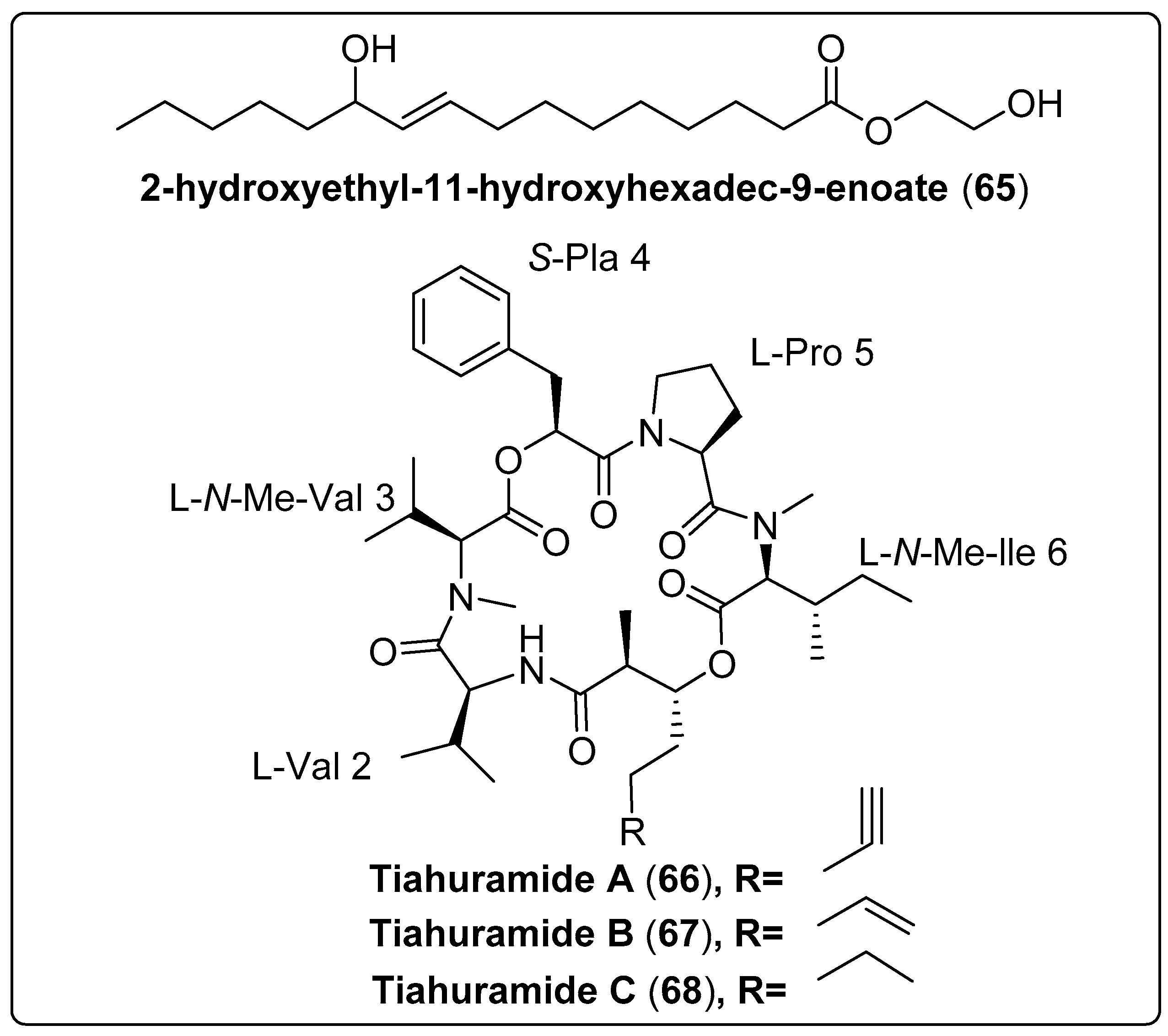
| 2-hydroxyethyl-11-hydroxyhexadec-9-enoate, not reported (65) | Leptolyngbya sp./Gulf of Thailand, Chumpon and Chonburi provinces | CC | Antibacterial activity Models: Vibrio harveyi and V. parahaemolyticus Assay: agar plate diffusion Concentrations: 0, 100, 125, 250, 350, 500, 650, 800 and 1000 µg/mL−1 MIC: 250–1000 µg/mL−1 for V. harveyi MIC: 350–1000 µg/mL−1 for V. parahaemolyticus PC: ampicillin (500 µg/mL−1) NC: penicillin (800 µg/mL−1) | [68] |
| Tiahuramide A, cyclic depsipeptide (66) | Lyngbya majuscule Harvey ex Gomont/Tiahura sector, Moorea Island in French Polynesia | TLC and HPLC | Antibacterial activity Assay: 96-well plates Model: Aeromonas salmonicida (MIC: 27 µM) Vibrio anguillarum (MIC: 33 µM) Shewanella baltica (MIC: >50 µM) Escherichia coli (MIC: 35 µM) Micrococcus luteus (MIC: 47 µM) | [69] |
| Tiahuramide B, cyclic depsipeptide (67) | Lyngbya majuscule Harvey ex Gomont/Tiahura sector, Moorea Island in French Polynesia | TLC and HPLC | Antibacterial activity Assay: 96-well plates Model: Aeromonas salmonicida (MIC: 9.4 µM) Vibrio anguillarum (MIC: 8.5 µM) Shewanella baltica (MIC: 22 µM) Escherichia coli (MIC: 12 µM) Micrococcus luteus (MIC: 29 µM) | [69] |
| Tiahuramide C, cyclic depsipeptide (68) | Lyngbya majuscule Harvey ex Gomont/Tiahura sector, Moorea Island in French Polynesia | TLC and HPLC | Antibacterial activity Assay: 96-well plates Model: Aeromonas salmonicida (MIC: 6.7 µM) Vibrio anguillarum (MIC: 7.4 µM) Shewanella baltica (MIC: 16 µM) Escherichia coli (MIC: 14 µM) Micrococcus luteus (MIC: 17 µM) | [69] |
3.7. Other Activities
| Compound Name/Class | Source/Place and Date | Separation Tools | Pharmacological Activity | Reference |
|---|---|---|---|---|
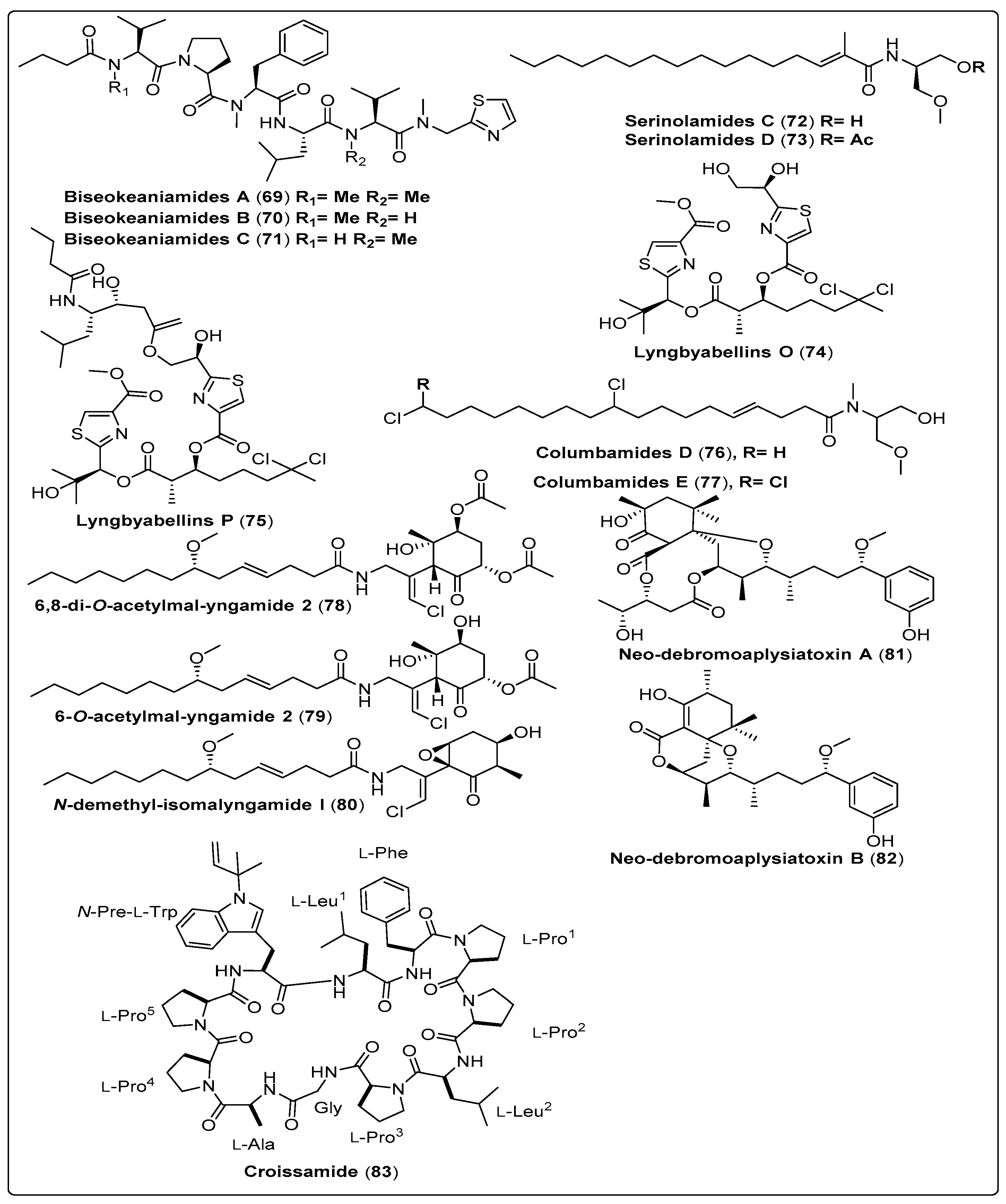
| Biseokeaniamide A, linear lipopeptide (69) | Okeania sp./Bise, Okinawa Prefecture, Japan (April 2015) | RP-HPLC and OSD-CC | Cell growth-inhibitory activity Model: HeLa and HL60 cells Assay: MTT assay IC50: 29 µM for HeLa cells IC50: 30 µM for HL60 cells PC: Tunicamycin Sterol O-acyltransferase inhibitory activity Model: CHO cells of African green monkey Assays: Cell-based assay IC50: 1.8 µM for SOAT1 PC: Purpactin A (IC50: 2.5 µM for SOAT1) IC50: 1.3 µM for SOAT2 PC: Purpactin A (IC50: 1.5 µM for SOAT2) Enzyme-based assay IC50: 1.8 µM for SOAT1 PC: Purpactin A (IC50: 0.9 µM for SOAT1) IC50: 9.6 µM for SOAT2 PC: Purpactin A (IC50: 1.8 µM for SOAT2) | [70] |
| Biseokeaniamide B, linear lipopeptide (70) | Okeania sp./Bise, Okinawa Prefecture, Japan (April 2015) | RP-HPLC and OSD-CC | Cell growth-inhibitory activity Model: HeLa and HL60 cells Assay: MTT assay IC50: 4.5 µM for HeLa cells IC50: 19 µM for HL60 cells PC: Tunicamycin Sterol O-acyltransferase inhibitory activity Model: CHO cells of African green monkey Assays: Cell-based assay IC50: 6.9 µM for SOAT1 PC: Purpactin A (IC50: 2.5 µM for SOAT1) IC50: 2.5 µM for SOAT2 PC: Purpactin A (IC50: 1.5 µM for SOAT2) Enzyme-based assay IC50: 6.8 µM for SOAT1 PC: Purpactin A (IC50: 0.9 µM for SOAT1) IC50: 9.9 µM for SOAT2 PC: Purpactin A (IC50: 1.8 µM for SOAT2) | [70] |
| Biseokeaniamide C, linear lipopeptides (71) | Okeania sp./Bise, Okinawa Prefecture, Japan (April 2015) | RP-HPLC and OSD-CC | Cell growth-inhibitory activities Model: HeLa and HL60 cells Assay: MTT assay IC50: 43 µM for HeLa cells IC50: >100 µM for HL60 cells PC: Tunicamycin Sterol O-acyltransferase inhibitory activity Model: CHO cells of African green monkey Assays: Cell-based assay IC50: >12 µM for SOAT1 PC: Purpactin A (IC50: 2.5 µM for SOAT1) IC50: 9.6 µM for SOAT2 PC: Purpactin A (IC50: 1.5 µM for SOAT2) Enzyme-based assay IC50: 11 µM for SOAT1 PC: Purpactin A (IC50: 0.9 µM for SOAT1) IC50: >32 µM for SOAT2 PC: Purpactin A (IC50: 1.8 µM for SOAT2) | [70] |
| Serinolamide C, fatty acid amide (72) | Okeania sp./Jeddah, Saudi Arabia (April 2015) | RP-HPLC | Antifouling activity Model: Amphibalanus amphitrite larvae EC50: Not reported Cytotoxic activity Model: Breast cancer cells (MCF7) GI50: Not reported | [71] |
| Serinolamide D, fatty acid amide (73) | Okeania sp./Jeddah, Saudi Arabia (April 2015) | RP-HPLC | Antifouling activity Model: Amphibalanus amphitrite larvae EC50: Not reported Cytotoxic activity Model: Breast cancer cells (MCF7) GI50: Not reported | [71] |
| Lyngbyabellin O, Not reported (74) | Okeania sp./Jeddah, Saudi Arabia (April 2015) | RP-HPLC | Antifouling activity Model: Amphibalanus amphitrite larvae EC50: 0.38 µM Cytotoxic activity Model: Breast cancer cells (MCF7) GI50: >160 µM | [71] |
| Lyngbyabellin P, Not reported (75) | Okeania sp./Jeddah, Saudi Arabia (April 2015) | RP-HPLC | Antifouling activity Model: Amphibalanus amphitrite larvae EC50: 0.73 µM Cytotoxic activity Model: Breast cancer cells (MCF7) GI50: 9 mM | [71] |
| Columbamide D, chlorinated fatty acid amide (76) | Moorea bouillonii/Mantanani Island in Sabah, Malaysia | OSD-CC and HPLC | The biological activity could not be assessed because of highly cytotoxictrace amounts | [73] |
| Columbamide E, chlorinated fatty acid amide (77) | Moorea bouillonii/Mantanani Island in Sabah, Malaysia | OSD-CC and HPLC | The biological activity could not be assessed because of highly cytotoxic trace amounts | [73] |
| 6,8-di-O-acetylmal-yngamide 2, Malyngamide serie (78) | Moorea producens/Bise, Okinawa Prefecture, Japan (April 2016) | OSD-CC and HPLC | Stimulation of glucose uptake Model: L6 myotubes Concentration: (10–40 µM) PC: Nepodin NC: DMSO | [74] |
| 6-O-acetylmal-yngamide 2, Malyngamide series (79) | Moorea producens/Bise, Okinawa Prefecture, Japan (April 2016) | OSD-CC and HPLC | Stimulation of glucose uptake Model: L6 myotubes Concentration: (10–40 µM) PC: Nepodin NC: DMSO | [74] |
| N-demethyl-isomal-yngamide I, Malyngamide series (80) | Moorea producens/Bise, Okinawa Prefecture, Japan (April 2016) | OSD-CC and HPLC | Stimulation of glucose uptake Model: L6 myotubes Concentration: (10–40 µM) PC: Nepodin NC: DMSO | [74] |
| Neo-debromoaplysiatoxin A, polyketide (81) | Lyngbya sp./Hainan Island, China (November of 2016) | UPLC and HPLC | Blocking activity against Kv1.5 Model: Chinese hamster ovary IC50 = 6.94 ± 0.26 µM PC: Not reported | [72] |
| Neo-debromoaplysiatoxin B, polyketide (82) | Lyngbya sp./Hainan Island, China (November of 2016) | UPLC and HPLC | Blocking activity against Kv1.5 Model: Chinese hamster ovary IC50 = 0.30 ± 0.05 µM PC: Not reported | [72] |
| Croissamide, cyclic peptide (83) | Symploca sp./Minna Island, Okinawa | RP-HPLC and RP-CC | Did not show any significant activities for antimalarial activity, protease inhibitory activity and antibacterial activity | [75] |
4. Marine Cyanobacteria in Clinical Trials
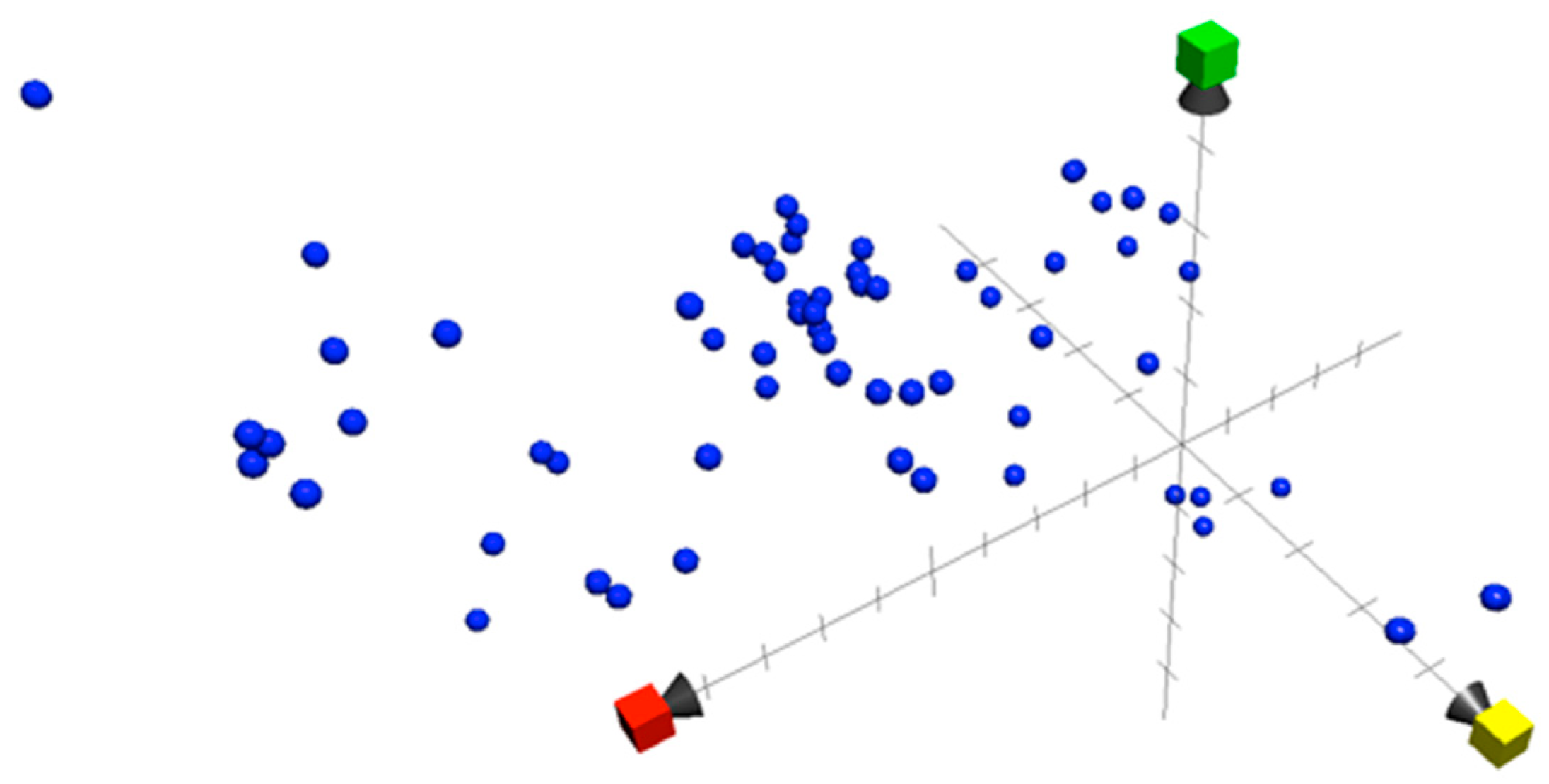
5. Chemical Property Space
6. Biological Properties
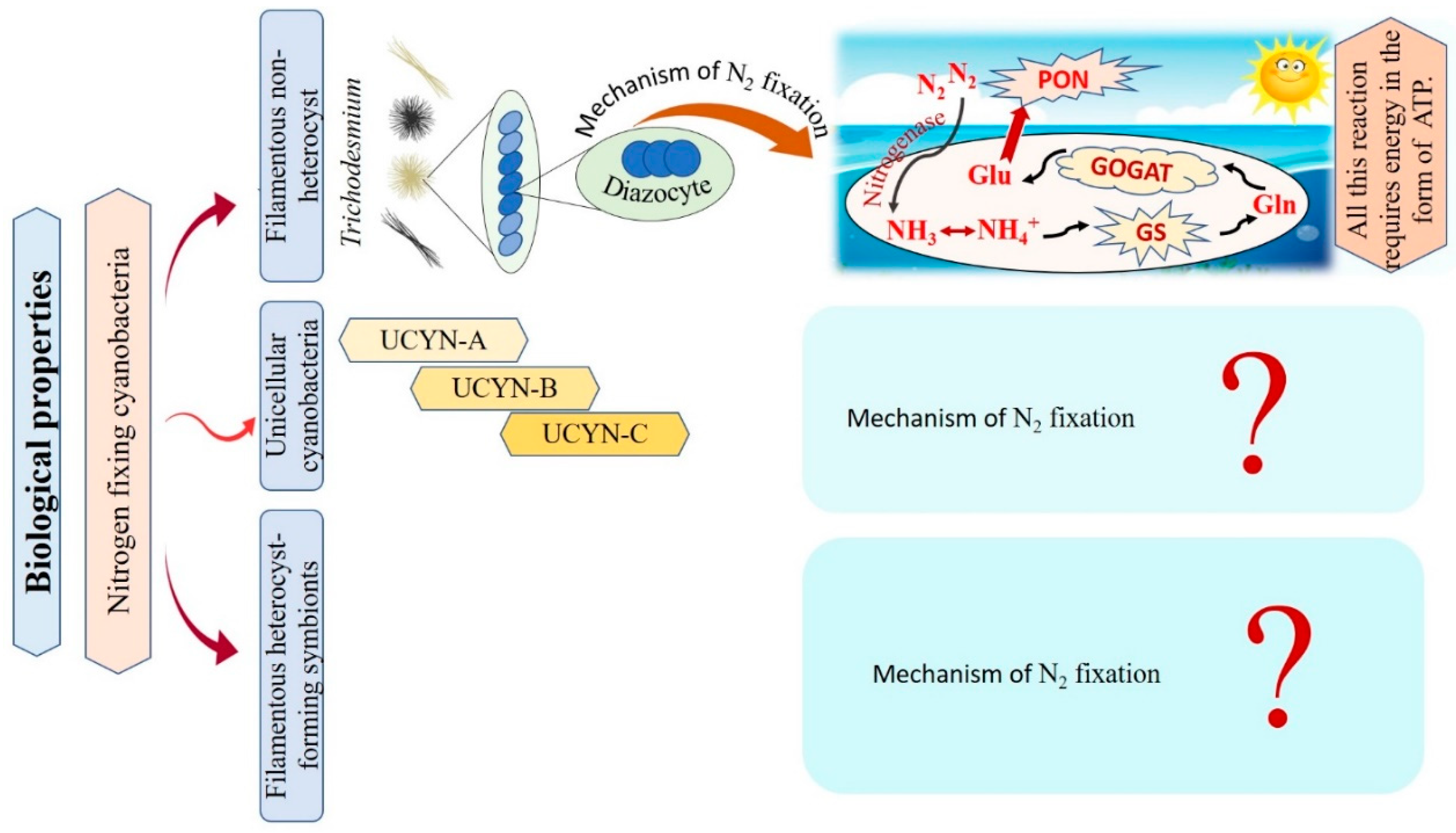
6.1. Mechanism of Marine Cyanobacteria in Nitrogen Fixation
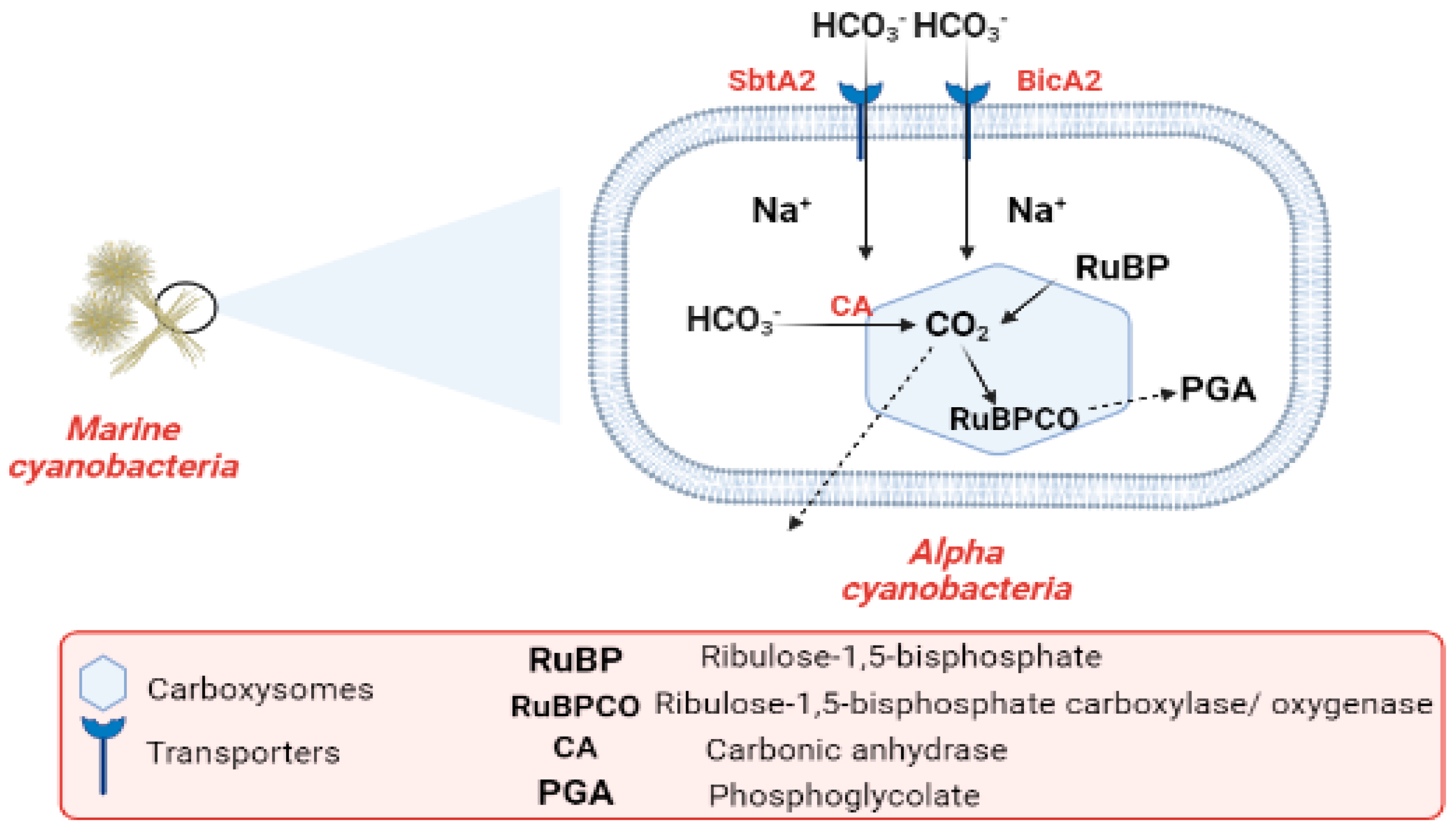
6.2. Mechanism of Marine Cyanobacteria in CO2 Fixation
7. Effect of Climate Changes on Marine Cyanobacteria
8. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, S.; Shi, N.; Huang, M.; Tan, X.; Yan, X.; Wang, A.; Huang, Y.; Ji, R.; Zhou, D.; Zhu, Y.-G.; et al. MoS2 Nanosheets–Cyanobacteria Interaction: Reprogrammed Carbon and Nitrogen Metabolism. ACS Nano 2021, 15, 16344–16356. [Google Scholar] [CrossRef]
- Azeez, R.; Dhanalakshmi, P.K.; Surenirakumar, K.; Nallamuthu, T. Growth and biochemical parameters of selective cultured cyanobacteria and exploiting antibacterial potency against human bacterial pathogens. Elixer. Appl. Bot. 2014, 72, 25537–25543. [Google Scholar]
- Altermann, W.; Kazmierczak, J.; Oren, A.; Wright, D.T. Cyanobacterial calcification and its rock-building potential during 3.5 billion years of Earth history. Geobiology 2006, 4, 147–166. [Google Scholar] [CrossRef]
- Bishoyi, A.K.; Sahoo, C.R.; Padhy, R.N. Recent progression of cyanobacteria and their pharmaceutical utility: An update. J. Biomol. Struct. Dyn. 2022, 41, 4219–4252. [Google Scholar] [CrossRef] [PubMed]
- Jirí, K.; Kastovský, J. Coincidences of structural and molecular characters in evolutionary lines of cyanobacteria. Algol. Stud. 2003, 109, 305–325. [Google Scholar] [CrossRef]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- Freeman, C.J.; Thacker, R.W. Complex interactions between marine sponges and their symbiotic microbial communities. Limnol. Oceanogr. 2011, 56, 1577–1586. [Google Scholar] [CrossRef]
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.T.; Egan, S. Tiny Microbes with a Big Impact: The Role of Cyanobacteria and Their Metabolites in Shaping Our Future. Mar. Drugs 2016, 14, 97. [Google Scholar] [CrossRef]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef]
- Zehr, J.P. Nitrogen fixation by marine cyanobacteria. Trends Microbiol. 2011, 19, 162–173. [Google Scholar] [CrossRef]
- Waterbury, J.B.; Watson, S.W.; Guillard, R.R.L.; Brand, L. Widespread occurrence of a unicellular, marine, planktonic, cyanobacterium. Nature 1979, 277, 293–294. [Google Scholar] [CrossRef]
- Sigman, D.M.; Boyle, E.A. Glacial/interglacial variations in atmospheric carbon dioxide. Nature 2000, 407, 859–869. [Google Scholar] [CrossRef]
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef]
- Ridley, C.P.; Bergquist, P.R.; Harper, M.K.; Faulkner, D.J.; Hooper, J.N.A.; Haygood, M.G. Speciation and Biosynthetic Variation in Four Dictyoceratid Sponges and Their Cyanobacterial Symbiont, Oscillatoria spongeliae. Chem. Biol. 2005, 12, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Hathaway, B.J.; Sudek, S.; Haygood, M.G.; Rosovitz, M.J.; Ravel, J.; Schmidt, E.W. Natural combinatorial peptide libraries in cyanobacterial symbionts of marine ascidians. Nat. Chem. Biol. 2006, 2, 729. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Harrigan, G.G.; Goetz, G.; Horgen, F.D. The cyanobacterial origin of potent anticancer agents originally isolated from sea hares. Curr. Med. Chem. 2002, 9, 1791–1806. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.M.T.D.; Herath, K.H.I.N.M.; Sanjeewa, K.K.A.; Jayawardena, T.U. Recent Reports on Bioactive Compounds from Marine Cyanobacteria in Relation to Human Health Applications. Life 2023, 13, 1411. [Google Scholar] [CrossRef]
- Kultschar, B.; Llewellyn, C. Secondary Metabolites in Cyanobacteria; Vijayakumar, R., Raja, S.S.S., Eds.; IntechOpen: Rijeka, Croatia, 2018; ISBN 978-1-78923-643-9. [Google Scholar]
- Uzair, B.; Tabassum, S.; Rasheed, M.; Rehman, S.F. Exploring Marine Cyanobacteria for Lead Compounds of Pharmaceutical Importance. Sci. World J. 2012, 2012, 179782. [Google Scholar] [CrossRef]
- Bhuyar, P.; Rahim, M.H.A.; Maniam, G.P.; Ramaraj, R.; Govindan, N. Exploration of bioactive compounds and antibacterial activity of marine blue-green microalgae (Oscillatoria sp.) isolated from coastal region of west Malaysia. SN Appl. Sci. 2020, 2, 1906. [Google Scholar] [CrossRef]
- Singh, U.; Singh, P.; Singh, A.K.; Laxmi; Kumar, D.; Tilak, R.; Shrivastava, S.K.; Asthana, R.K. Identification of antifungal and antibacterial biomolecules from a cyanobacterium, Arthrospira platensis. Algal Res. 2021, 54, 102215. [Google Scholar] [CrossRef]
- Mondal, A.; Bose, S.; Banerjee, S.; Patra, J.K.; Malik, J.; Mandal, S.K.; Kilpatrick, K.L.; Das, G.; Kerry, R.G.; Fimognari, C.; et al. Marine Cyanobacteria and Microalgae Metabolites—A Rich Source of Potential Anticancer Drugs. Mar. Drugs 2020, 18, 476. [Google Scholar] [CrossRef] [PubMed]
- Demirkiran, O.; Almaliti, J.; Leão, T.; Navarro, G.; Byrum, T.; Valeriote, F.A.; Gerwick, L.; Gerwick, W.H. Portobelamides A and B and Caciqueamide, Cytotoxic Peptidic Natural Products from a Caldora sp. Marine Cyanobacterium. J. Nat. Prod. 2021, 84, 2081–2093. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-C.; Strangman, W.K.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Thalassospiramides A and B, Immunosuppressive Peptides from the Marine Bacterium Thalassospira sp. Org. Lett. 2007, 9, 1525–1528. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Kokkaliari, S.; Ratnayake, R.; Sauvage, T.; dos Santos, L.A.H.; Luesch, H.; Paul, V.J. Anti-Inflammatory Dysidazirine Carboxylic Acid from the Marine Cyanobacterium Caldora sp. Collected from the Reefs of Fort Lauderdale, Florida. Molecules 2022, 27, 1717. [Google Scholar] [CrossRef] [PubMed]
- Renugadevi, K.; Valli Nachiyar, C.; Sowmiya, P.; Sunkar, S. Antioxidant activity of phycocyanin pigment extracted from marine filamentous cyanobacteria Geitlerinema sp. TRV57. Biocatal. Agric. Biotechnol. 2018, 16, 237–242. [Google Scholar] [CrossRef]
- Gutiérrez, M.; Suyama, T.L.; Engene, N.; Wingerd, J.S.; Matainaho, T.; Gerwick, W.H. Apratoxin D, a Potent Cytotoxic Cyclodepsipeptide from Papua New Guinea Collections of the Marine Cyanobacteria Lyngbya majuscula and Lyngbya sordida. J. Nat. Prod. 2008, 71, 1099–1103. [Google Scholar] [CrossRef]
- Almaliti, J.; Malloy, K.L.; Glukhov, E.; Spadafora, C.; Gutiérrez, M.; Gerwick, W.H. Dudawalamides A–D, Antiparasitic Cyclic Depsipeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2017, 80, 1827–1836. [Google Scholar] [CrossRef]
- Mishra, A.K.; Kaushik, M.S.; Tiwari, D.N. Chapter 8—Nitrogenase and Hydrogenase: Enzymes for Nitrogen Fixation and Hydrogen Production in Cyanobacteria; Mishra, A.K., Tiwari, D.N., Rai, A.N.B.T.-C., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 173–191. ISBN 978-0-12-814667-5. [Google Scholar]
- Berman-Frank, I.; Lundgren, P.; Falkowski, P. Nitrogen fixation and photosynthetic oxygen evolution in cyanobacteria. Res. Microbiol. 2003, 154, 157–164. [Google Scholar] [CrossRef]
- Stal, L.J. Nitrogen Fixation in Cyanobacteria. eLS 2015, 1–9. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.-Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Fekry, M.I.; Al-Hammady, M.A.; Khalil, M.N.; El-Seedi, H.R.; Meyer, A.; Porzel, A.; Westphal, H.; Wessjohann, L.A. Cytotoxic Effects of Sarcophyton sp. Soft Corals—Is There a Correlation to Their NMR Fingerprints? Mar. Drugs 2017, 15, 211. [Google Scholar] [CrossRef] [PubMed]
- Yosri, N.; Khalifa, S.A.M.; Guo, Z.; Xu, B.; Zou, X.; El-Seedi, H.R. Marine organisms: Pioneer natural sources of polysaccharides/proteins for green synthesis of nanoparticles and their potential applications. Int. J. Biol. Macromol. 2021, 193, 1767–1798. [Google Scholar] [CrossRef] [PubMed]
- Sergeev, V.N.; Gerasimenko, L.M.; Zavarzin, G.A. The Proterozoic History and Present State of Cyanobacteria. Microbiology 2002, 71, 623–637. [Google Scholar] [CrossRef]
- Golubic, S.; Seong-Joo, L. Early cyanobacterial fossil record: Preservation, palaeoenvironments and identification. Eur. J. Phycol. 1999, 34, 339–348. [Google Scholar] [CrossRef]
- Golubic, S.; Hofmann, H.J. Comparison of Holocene and mid-Precambrian Entophysalidaceae (Cyanophyta) in stromatolitic algal mats: Cell division and degradation. J. Paleontol. 1976, 50, 1074–1082. [Google Scholar]
- Hofmann, H.J. Precambrian Microflora, Belcher Islands, Canada: Significance and Systematics. J. Paleontol. 1976, 50, 1040–1073. [Google Scholar]
- Lanier, W.P. Interstitial and peloid microfossils from the 2.0 Ga Gunflint Formation: Implications for the paleoecology of the Gunflint Stromatolites. Precambrian Res. 1989, 45, 291–318. [Google Scholar] [CrossRef]
- Sharma, M.; Shukla, Y. The evolution and distribution of life in the Precambrian eon-global perspective and the Indian record. J. Biosci. 2009, 34, 765–776. [Google Scholar] [CrossRef]
- Sharma, M. Palaeobiology of Mesoproterozoic Salkhan Limestone, Semri Group, Rohtas, Bihar, India: Systematics and significance. J. Earth Syst. Sci. 2006, 115, 67–98. [Google Scholar] [CrossRef]
- Sharma, M.; Sergeev, V.N. Genesis of carbonate precipitate patterns and associated microfossils in Mesoproterozoic formations of India and Russia—A comparative study. Precambrian Res. 2004, 134, 317–347. [Google Scholar] [CrossRef]
- Knoll, A.H.; Sergeev, V.N. Taphonomic and evolutionary changes across the Mesoproterozoic-Neoproterozoic transition. Neues Jahrb. Geol. Palaontol. Abh. 1995, 195, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Castenholz, R.W. Species usage, concept, and evolution in the cyanobacteria (blue green algae). J. Phycol. 1992, 28, 737–745. [Google Scholar] [CrossRef]
- Komárek, J. Review of the cyanobacterial genera implying planktic species after recent taxonomic revisions according to polyphasic methods: State as of 2014. Hydrobiologia 2016, 764, 259–270. [Google Scholar] [CrossRef]
- Komárek, J. Recent changes (2008) in cyanobacteria taxonomy based on a combination of molecular background with phenotype and ecological consequences (genus and species concept). Hydrobiologia 2010, 639, 245–259. [Google Scholar] [CrossRef]
- Naman, C.B.; Rattan, R.; Nikoulina, S.E.; Lee, J.; Miller, B.W.; Moss, N.A.; Armstrong, L.; Boudreau, P.D.; Debonsi, H.M.; Valeriote, F.A.; et al. Integrating Molecular Networking and Biological Assays to Target the Isolation of a Cytotoxic Cyclic Octapeptide, Samoamide A, from an American Samoan Marine Cyanobacterium. J. Nat. Prod. 2017, 80, 625–633. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Kudo, T.; Yamano, A.; Sumimoto, S.; Iwasaki, A.; Suenaga, K.; Teruya, T. Odobromoamide, a terminal alkynyl bromide-containing cyclodepsipeptide from the marine cyanobacterium Okeania sp. Bull. Chem. Soc. Jpn. 2017, 90, 436–440. [Google Scholar] [CrossRef]
- Tao, Y.; Li, P.; Zhang, D.; Glukhov, E.; Gerwick, L.; Zhang, C.; Murray, T.F.; Gerwick, W.H. Samholides, Swinholide-Related Metabolites from a Marine Cyanobacterium cf. Phormidium sp. J. Org. Chem. 2018, 83, 3034–3046. [Google Scholar] [CrossRef]
- Nakamura, F.; Maejima, H.; Kawamura, M.; Arai, D.; Okino, T.; Zhao, M.; Ye, T.; Lee, J.; Chang, Y.-T.; Fusetani, N.; et al. Kakeromamide A, a new cyclic pentapeptide inducing astrocyte differentiation isolated from the marine cyanobacterium Moorea bouillonii. Bioorg. Med. Chem. Lett. 2018, 28, 2206–2209. [Google Scholar] [CrossRef]
- Yu, H.-B.; Glukhov, E.; Li, Y.; Iwasaki, A.; Gerwick, L.; Dorrestein, P.C.; Jiao, B.-H.; Gerwick, W.H. Cytotoxic Microcolin Lipopeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2019, 82, 2608–2619. [Google Scholar] [CrossRef]
- Choi, H.; Engene, N.; Byrum, T.; Hwang, S.; Oh, D.-C.; Gerwick, W.H. Dragocins A−D, Structurally Intriguing Cytotoxic Metabolites from a Panamanian Marine Cyanobacterium. Org. Lett. 2019, 21, 266–270. [Google Scholar] [CrossRef]
- Nagai, H.; Sato, S.; Iida, K.; Hayashi, K.; Kawaguchi, M.; Uchida, H.; Satake, M. Oscillatoxin I: A New Aplysiatoxin Derivative, from a Marine Cyanobacterium. Toxins 2019, 11, 366. [Google Scholar] [CrossRef]
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Omura, S.; Suenaga, K. Isolation and total synthesis of hoshinolactam, an antitrypanosomal lactam from a marine cyanobacterium. Org. Lett. 2017, 19, 890–893. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Iwasaki, A.; Sezawa, D.; Fujimura, H.; Nozaki, T.; Saito-Nakano, Y.; Suenaga, K.; Teruya, T. Isolation and Total Synthesis of Mabuniamide, a Lipopeptide from an Okeania sp. Marine Cyanobacterium. J. Nat. Prod. 2019, 82, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Sweeney-Jones, A.M.; Gagaring, K.; Antonova-Koch, J.; Zhou, H.; Mojib, N.; Soapi, K.; Skolnick, J.; McNamara, C.W.; Kubanek, J. Antimalarial Peptide and Polyketide Natural Products from the Fijian Marine Cyanobacterium Moorea producens. Mar. Drugs 2020, 18, 167. [Google Scholar] [CrossRef]
- Kurisawa, N.; Iwasaki, A.; Jeelani, G.; Nozaki, T.; Suenaga, K. Iheyamides A–C, Antitrypanosomal Linear Peptides Isolated from a Marine Dapis sp. Cyanobacterium. J. Nat. Prod. 2020, 83, 1684–1690. [Google Scholar] [CrossRef]
- Ebihara, A.; Iwasaki, A.; Miura, Y.; Jeelani, G.; Nozaki, T.; Suenaga, K. Isolation and Total Synthesis of Bromoiesol sulfates, Antitrypanosomal arylethers from a Salileptolyngbya sp. Marine Cyanobacterium. J. Org. Chem. 2021, 86, 11763–11770. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Iwasaki, A.; Kurisawa, N.; Suzuki, R.; Jeelani, G.; Matsubara, T.; Sato, T.; Nozaki, T.; Suenaga, K. Motobamide, an Antitrypanosomal Cyclic Peptide from a Leptolyngbya sp. Marine Cyanobacterium. J. Nat. Prod. 2021, 84, 1649–1655. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Law, B.K.; Paul, V.J.; Luesch, H. Grassystatins D–F, Potent Aspartic Protease Inhibitors from Marine Cyanobacteria as Potential Antimetastatic Agents Targeting Invasive Breast Cancer. J. Nat. Prod. 2017, 80, 2969–2986. [Google Scholar] [CrossRef]
- Gallegos, D.A.; Saurí, J.; Cohen, R.D.; Wan, X.; Videau, P.; Vallota-Eastman, A.O.; Shaala, L.A.; Youssef, D.T.A.; Williamson, R.T.; Martin, G.E.; et al. Jizanpeptins, Cyanobacterial Protease Inhibitors from a Symploca sp. Cyanobacterium Collected in the Red Sea. J. Nat. Prod. 2018, 81, 1417–1425. [Google Scholar] [CrossRef]
- Keller, L.; Canuto, K.M.; Liu, C.; Suzuki, B.M.; Almaliti, J.; Sikandar, A.; Naman, C.B.; Glukhov, E.; Luo, D.; Duggan, B.M.; et al. Tutuilamides A–C: Vinyl-Chloride-Containing Cyclodepsipeptides from Marine Cyanobacteria with Potent Elastase Inhibitory Properties. ACS Chem. Biol. 2020, 15, 751–757. [Google Scholar] [CrossRef]
- Luo, D.; Putra, M.; Ye, T.; Paul, V.; Luesch, H. Isolation, Structure Elucidation and Biological Evaluation of Lagunamide D: A New Cytotoxic Macrocyclic Depsipeptide from Marine Cyanobacteria. Mar. Drugs 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.Y.; Ong, J.F.; Goh, H.C.; Coffill, C.R.; Tan, L.T. Benderamide A, a Cyclic Depsipeptide from a Singapore Collection of Marine Cyanobacterium cf Lyngbya sp. Mar. Drugs 2018, 16, 409. [Google Scholar] [CrossRef] [PubMed]
- Phyo, M.Y.; Goh, T.M.; Goh, J.X.; Tan, L.T. Trikoramides B–D, Bioactive Cyanobactins from the Marine Cyanobacterium Symploca hydnoides. Mar. Drugs 2021, 19, 548. [Google Scholar] [CrossRef] [PubMed]
- Phyo, M.Y.; Katermeran, N.P.; Goh, J.X.; Tan, L.T. Trikoveramides A-C, cyclic depsipeptides from the marine cyanobacterium Symploca hydnoides. Phytochemistry 2021, 190, 112879. [Google Scholar] [CrossRef] [PubMed]
- Maneechote, N.; Yingyongnarongkul, B.; Suksamran, A.; Lumyong, S. Inhibition of Vibrio spp. by 2-Hydroxyethyl-11-hydroxyhexadec-9-enoate of Marine Cyanobacterium Leptolyngbya sp. LT19. Aquac. Res. 2017, 48, 2088–2095. [Google Scholar] [CrossRef]
- Levert, A.; Alvariño, R.; Bornancin, L.; Abou Mansour, E.; Burja, A.M.; Genevière, A.-M.; Bonnard, I.; Alonso, E.; Botana, L.; Banaigs, B. Structures and Activities of Tiahuramides A–C, Cyclic Depsipeptides from a Tahitian Collection of the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2018, 81, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Tadenuma, T.; Sumimoto, S.; Ohshiro, T.; Ozaki, K.; Kobayashi, K.; Teruya, T.; Tomoda, H.; Suenaga, K. Biseokeaniamides A, B, and C, sterol O-acyltransferase inhibitors from an Okeania sp. marine cyanobacterium. J. Nat. Prod. 2017, 80, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Petitbois, J.G.; Casalme, L.O.; Lopez, J.A.V.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Umezawa, T.; Matsuda, F.; et al. Serinolamides and Lyngbyabellins from an Okeania sp. Cyanobacterium Collected from the Red Sea. J. Nat. Prod. 2017, 80, 2708–2715. [Google Scholar] [CrossRef]
- Han, B.-N.; Liang, T.-T.; Keen, L.J.; Fan, T.-T.; Zhang, X.-D.; Xu, L.; Zhao, Q.; Wang, S.-P.; Lin, H.-W. Two Marine Cyanobacterial Aplysiatoxin Polyketides, Neo-debromoaplysiatoxin A and B, with K+ Channel Inhibition Activity. Org. Lett. 2018, 20, 578–581. [Google Scholar] [CrossRef]
- Lopez, J.A.V.; Petitbois, J.G.; Vairappan, C.S.; Umezawa, T.; Matsuda, F.; Okino, T. Columbamides D and E: Chlorinated Fatty Acid Amides from the Marine Cyanobacterium Moorea bouillonii Collected in Malaysia. Org. Lett. 2017, 19, 4231–4234. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, K.; Yamano, A.; Ozaki, K.; Sumimoto, S.; Iwasaki, A.; Suenaga, K.; Teruya, T. Three new malyngamides from the marine cyanobacterium Moorea producens. Mar. Drugs 2017, 15, 367. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Iwasaki, A.; Sumimoto, S.; Sano, T.; Hitomi, Y.; Ohno, O.; Suenaga, K. Croissamide, a proline-rich cyclic peptide with an N-prenylated tryptophan from a marine cyanobacterium Symploca sp. Tetrahedron Lett. 2018, 59, 3806–3809. [Google Scholar] [CrossRef]
- Huang, I.-S.; Zimba, P. V Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [CrossRef]
- Rosén, J.; Lövgren, A.; Kogej, T.; Muresan, S.; Gottfries, J.; Backlund, A. ChemGPS-NPWeb: Chemical space navigation online. J. Comput. Aided. Mol. Des. 2009, 23, 253–259. [Google Scholar] [CrossRef]
- Alajlani, M.M.; Backlund, A. Evaluating Antimycobacterial Screening Schemes Using Chemical Global Positioning System-Natural Product Analysis. Molecules 2020, 25, 945. [Google Scholar] [CrossRef]
- Alajlani, M.M. The Chemical Property Position of Bedaquiline Construed by a Chemical Global Positioning System-Natural Product. Molecules 2022, 27, 753. [Google Scholar] [CrossRef]
- Alajlani, A.M.B. Predicting the mechanism of action of antituberculosis agents using chemical global positioning system—Natural product. Planta Med. 2016, 82, P47. [Google Scholar] [CrossRef]
- Díez, B.; Bergman, B.; El-Shehawy, R. Marine diazotrophic cyanobacteria: Out of the blue. Plant Biotechnol. 2008, 25, 221–225. [Google Scholar] [CrossRef]
- Capone, D.G.; Burns, J.A.; Montoya, J.P.; Subramaniam, A.; Mahaffey, C.; Gunderson, T.; Michaels, A.F.; Carpenter, E.J. Nitrogen fixation by Trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical North Atlantic Ocean. Glob. Biogeochem. Cycles 2005, 19, 1–17. [Google Scholar] [CrossRef]
- Breitbarth, E.; Oschlies, A.; LaRoche, J. Physiological constraints on the global distribution of Trichodesmium—Effect of temperature on diazotrophy. Biogeosciences 2007, 4, 53–61. [Google Scholar] [CrossRef]
- El-Shehawy, R.; Lugomela, C.; Ernst, A.; Bergman, B. Diurnal expression of hetR and diazocyte development in the filamentous non-heterocystous cyanobacterium Trichodesmium erythraeum. Microbiology 2003, 149, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Wannicke, N.; BP, K.; Voss, M. Release of fixed N2 and C as dissolved compounds by Trichodesmium erythreum and Nodularia spumigena under the influence of high light and high nutrient (P). Aquat. Microb. Ecol. 2009, 57, 175–189. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Capone, D.G. The nitrogen physiology of the marine N2-fixing cyanobacteria Trichodesmium spp. Trends Plant Sci. 2000, 5, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kranz, S.A.; Eichner, M.; Rost, B. Interactions between CCM and N2 fixation in Trichodesmium. Photosynth. Res. 2011, 109, 73–84. [Google Scholar] [CrossRef]
- Zehr, J.P.; Waterbury, J.B.; Turner, P.J.; Montoya, J.P.; Omoregie, E.; Steward, G.F.; Hansen, A.; Karl, D.M. Unicellular cyanobacteria fix N2 in the subtropical North Pacific Ocean. Nature 2001, 412, 635–638. [Google Scholar] [CrossRef]
- Zehr, J.P.; Mellon, M.T.; Zani, S. New Nitrogen-Fixing Microorganisms Detected in Oligotrophic Oceans by Amplification of Nitrogenase (nifH) Genes. Appl. Environ. Microbiol. 1998, 64, 3444–3450. [Google Scholar] [CrossRef]
- Zehr, J.P.; Bench, S.R.; Carter, B.J.; Hewson, I.; Niazi, F.; Shi, T.; Tripp, H.J.; Affourtit, J.P. Globally distributed uncultivated oceanic N2-fixing cyanobacteria lack oxygenic photosystem II. Science 2008, 322, 1110–1112. [Google Scholar] [CrossRef]
- Prechtl, J.; Kneip, C.; Lockhart, P.; Wenderoth, K.; Maier, U.-G. Intracellular Spheroid Bodies of Rhopalodia gibba Have Nitrogen-Fixing Apparatus of Cyanobacterial Origin. Mol. Biol. Evol. 2004, 21, 1477–1481. [Google Scholar] [CrossRef]
- Zehr, J.P.; Montoya, J.P.; Jenkins, B.D.; Hewson, I.; Mondragon, E.; Short, C.M.; Church, M.J.; Hansen, A.; Karl, D.M. Experiments linking nitrogenase gene expression to nitrogen fixation in the North Pacific subtropical gyre. Limnol. Oceanogr. 2007, 52, 169–183. [Google Scholar] [CrossRef]
- Stenegren, M.; Caputo, A.; Berg, C.; Bonnet, S.; Foster, R.A. Distribution and drivers of symbiotic and free-living diazotrophic cyanobacteria in the western tropical South Pacific. Biogeosciences 2018, 15, 1559–1578. [Google Scholar] [CrossRef]
- Thompson, A.W.; Zehr, J.P. Cellular interactions: Lessons from the nitrogen-fixing cyanobacteria. J. Phycol. 2013, 49, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Sohm, J.A.; Webb, E.A.; Capone, D.G. Emerging patterns of marine nitrogen fixation. Nat. Rev. Microbiol. 2011, 9, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Shiozaki, T.; Ijichi, M.; Kodama, T.; Takeda, S.; Furuya, K. Heterotrophic bacteria as major nitrogen fixers in the euphotic zone of the Indian Ocean. Glob. Biogeochem. Cycles 2014, 28, 1096–1110. [Google Scholar] [CrossRef]
- Dron, A.; Rabouille, S.; Claquin, P.; Le Roy, B.; Talec, A.; Sciandra, A. Light–dark (12:12) cycle of carbon and nitrogen metabolism in Crocosphaera watsonii WH8501: Relation to the cell cycle. Environ. Microbiol. 2012, 14, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Compaoré, J.; Stal, L.J. Oxygen and the light–dark cycle of nitrogenase activity in two unicellular cyanobacteria. Environ. Microbiol. 2010, 12, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Turk-Kubo, K.A.; Frank, I.E.; Hogan, M.E.; Desnues, A.; Bonnet, S.; Zehr, J.P. Diazotroph community succession during the VAHINE mesocosm experiment (New Caledonia lagoon). Biogeosciences 2015, 12, 7435–7452. [Google Scholar] [CrossRef]
- Taniuchi, Y.; Chen, Y.L.; Chen, H.-Y.; Tsai, M.-L.; Ohki, K. Isolation and characterization of the unicellular diazotrophic cyanobacterium Group C TW3 from the tropical western Pacific Ocean. Environ. Microbiol. 2012, 14, 641–654. [Google Scholar] [CrossRef]
- Reddy, K.J.; Haskell, J.B.; Sherman, D.M.; Sherman, L.A. Unicellular, aerobic nitrogen-fixing cyanobacteria of the genus Cyanothece. J. Bacteriol. 1993, 175, 1284–1292. [Google Scholar] [CrossRef]
- Andreeva, N.A.; Melnikov, V.V.; Snarskaya, D.D. The Role of Cyanobacteria in Marine Ecosystems. Russ. J. Mar. Biol. 2020, 46, 154–165. [Google Scholar] [CrossRef]
- Foster, R.A.; Subramaniam, A.; Zehr, J.P. Distribution and activity of diazotrophs in the Eastern Equatorial Atlantic. Environ. Microbiol. 2009, 11, 741–750. [Google Scholar] [CrossRef]
- Mishra, U.; Pabbi, S. Cyanobacteria: A potential biofertilizer for rice. Resonance 2004, 9, 6–10. [Google Scholar] [CrossRef]
- Karlson, A.M.L.; Duberg, J.; Motwani, N.H.; Hogfors, H.; Klawonn, I.; Ploug, H.; Barthel Svedén, J.; Garbaras, A.; Sundelin, B.; Hajdu, S.; et al. Nitrogen fixation by cyanobacteria stimulates production in Baltic food webs. Ambio 2015, 44, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.N.; Verma, I.; Kumar, M. Chapter 23—Cyanobacteria: Potential Source of Biofertilizer and Synthesizer of Metallic Nanoparticles; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 351–367. ISBN 978-0-12-819311-2. [Google Scholar]
- Levitan, O.; Rosenberg, G.; Setlik, I.; Setlikova, E.; Grigel, J.; Klepetar, J.; Prasil, O.; Berman-Frank, I. Elevated CO2 enhances nitrogen fixation and growth in the marine cyanobacterium Trichodesmium. Glob. Chang. Biol. 2007, 13, 531–538. [Google Scholar] [CrossRef]
- Case, A.E.; Atsumi, S. Cyanobacterial chemical production. J. Biotechnol. 2016, 231, 106–114. [Google Scholar] [CrossRef]
- Zhang, A.; Carroll, A.L.; Atsumi, S. Carbon recycling by cyanobacteria: Improving CO2 fixation through chemical production. FEMS Microbiol. Lett. 2017, 364, fnx165. [Google Scholar] [CrossRef] [PubMed]
- Rae, B.D.; Long, B.M.; Badger, M.R.; Price, G.D. Functions, Compositions, and Evolution of the Two Types of Carboxysomes: Polyhedral Microcompartments That Facilitate CO2 Fixation in Cyanobacteria and Some Proteobacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 357–379. [Google Scholar] [CrossRef] [PubMed]
- Yeates, T.O.; Kerfeld, C.A.; Heinhorst, S.; Cannon, G.C.; Shively, J.M. Protein-based organelles in bacteria: Carboxysomes and related microcompartments. Nat. Rev. Microbiol. 2008, 6, 681–691. [Google Scholar] [CrossRef]
- Turmo, A.; Gonzalez-Esquer, C.R.; Kerfeld, C.A. Carboxysomes: Metabolic modules for CO2 fixation. FEMS Microbiol. Lett. 2017, 364, fnx176. [Google Scholar] [CrossRef]
- Cannon, G.C.; Bradburne, C.E.; Aldrich, H.C.; Baker, S.H.; Heinhorst, S.; Jessup, M. Shively Microcompartments in Prokaryotes: Carboxysomes and Related Polyhedra. Appl. Environ. Microbiol. 2001, 67, 5351–5361. [Google Scholar] [CrossRef]
- Price, G.D.; Pengelly, J.J.L.; Forster, B.; Du, J.; Whitney, S.M.; von Caemmerer, S.; Badger, M.R.; Howitt, S.M.; Evans, J.R. The cyanobacterial CCM as a source of genes for improving photosynthetic CO2 fixation in crop species. J. Exp. Bot. 2013, 64, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Silvarrey Barruffa, A.; Sposito, V.; Faggian, R. Climate change and cyanobacteria harmful algae blooms: Adaptation practices for developing countries. Mar. Freshw. Res. 2021, 72, 1722–1734. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt tolerance of the harmful cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
















| Intervention or Treatment/Form | Condition or/Disease | Type of the Study/Clinical Trials Phase/No. of Participants | Dose | Administration Route/Effects | References ClinicalTrials.GovIdentifier |
|---|---|---|---|---|---|
| Apomivir®/Capsule | Influenza | Interventional/Phase 2/196 participants | Apomivir®: (120 mg b.i.d.) 1 capsule twice daily Placebo: 1 capsule twice daily | Orally after meal/Apomivir® has been proven to have excellent broad-spectrum antiviral ability, especially for seasonal influenza viruses (Influenza virus A and B) | (https://clinicaltrials.gov (accessed on 15 April 2023), NCT01677689) |
| Dietary Supplement: Water-soluble Extract of the cyanobacteria Aphanizomenon Flos-aquae (AFA)/Capsules | Congestive heart failure, chronic | Observational/Not reported/15 participants | 2 capsules 3 time a day | Oral/The improvement in patient’s quality of life after 12 months of trial admission | (https://clinicaltrials.gov accessed on 15 April 2023), NCT04515537) |
| Phycocyanin (Phycocare®) Other: Placebo/Not reported | Metastatic gastric cancer | Interventional/Not Applicable/110 participants | Phycocare: Every day for 9 months (except days of chemotherapy: no Phycocare) Placebo: Every day for 9 months (except days of chemotherapy = no Placebo) | Not reported/phycocyanin may give protection against oxaliplatin-induced neuropathy | (https://clinicaltrials.gov accessed on 15 April 2023), NCT05025826) |
| Dietary Supplement: Immulina TM Dietary Supplement: Placebo/Capsules | Post-acute COVID-19 syndrome | Interventional/Not Applicable/120 participants | Immulina TM: (200 mg per capsule) 2–200 mg capsules given by mouth in the morning and 2–200 mg capsules given by mouth in the evening for 8 weeks durationPlacebo: As the same | Oral/Not reported | (https://clinicaltrials.gov accessed on 15 April 2023), NCT05524532) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Seedi, H.R.; El-Mallah, M.F.; Yosri, N.; Alajlani, M.; Zhao, C.; Mehmood, M.A.; Du, M.; Ullah, H.; Daglia, M.; Guo, Z.; et al. Review of Marine Cyanobacteria and the Aspects Related to Their Roles: Chemical, Biological Properties, Nitrogen Fixation and Climate Change. Mar. Drugs 2023, 21, 439. https://doi.org/10.3390/md21080439
El-Seedi HR, El-Mallah MF, Yosri N, Alajlani M, Zhao C, Mehmood MA, Du M, Ullah H, Daglia M, Guo Z, et al. Review of Marine Cyanobacteria and the Aspects Related to Their Roles: Chemical, Biological Properties, Nitrogen Fixation and Climate Change. Marine Drugs. 2023; 21(8):439. https://doi.org/10.3390/md21080439
Chicago/Turabian StyleEl-Seedi, Hesham R., Mohamed F. El-Mallah, Nermeen Yosri, Muaaz Alajlani, Chao Zhao, Muhammad A. Mehmood, Ming Du, Hammad Ullah, Maria Daglia, Zhiming Guo, and et al. 2023. "Review of Marine Cyanobacteria and the Aspects Related to Their Roles: Chemical, Biological Properties, Nitrogen Fixation and Climate Change" Marine Drugs 21, no. 8: 439. https://doi.org/10.3390/md21080439
APA StyleEl-Seedi, H. R., El-Mallah, M. F., Yosri, N., Alajlani, M., Zhao, C., Mehmood, M. A., Du, M., Ullah, H., Daglia, M., Guo, Z., Khalifa, S. A. M., & Shou, Q. (2023). Review of Marine Cyanobacteria and the Aspects Related to Their Roles: Chemical, Biological Properties, Nitrogen Fixation and Climate Change. Marine Drugs, 21(8), 439. https://doi.org/10.3390/md21080439