A Polyol-Polyol Super-Carbon-Chain Compound Containing Thirty-Six Carbon Stereocenters from the Dinoflagellate Amphidinium gibbosum: Absolute Configuration and Multi-Segment Modification


Abstract
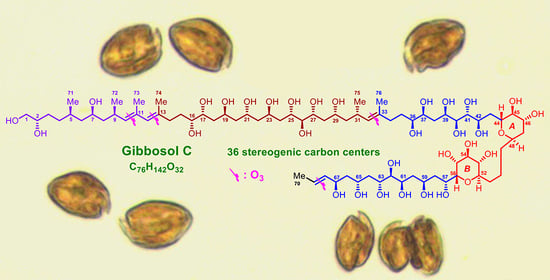
1. Introduction
2. Results and Discussion
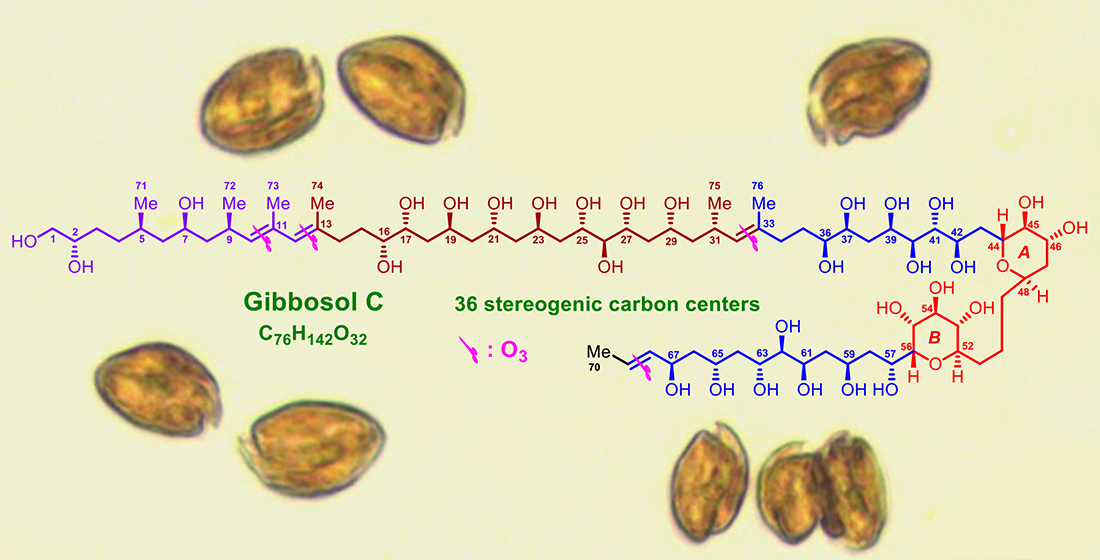
2.1. Planar Structure and Ozonolysis of Gibbosol C (1)
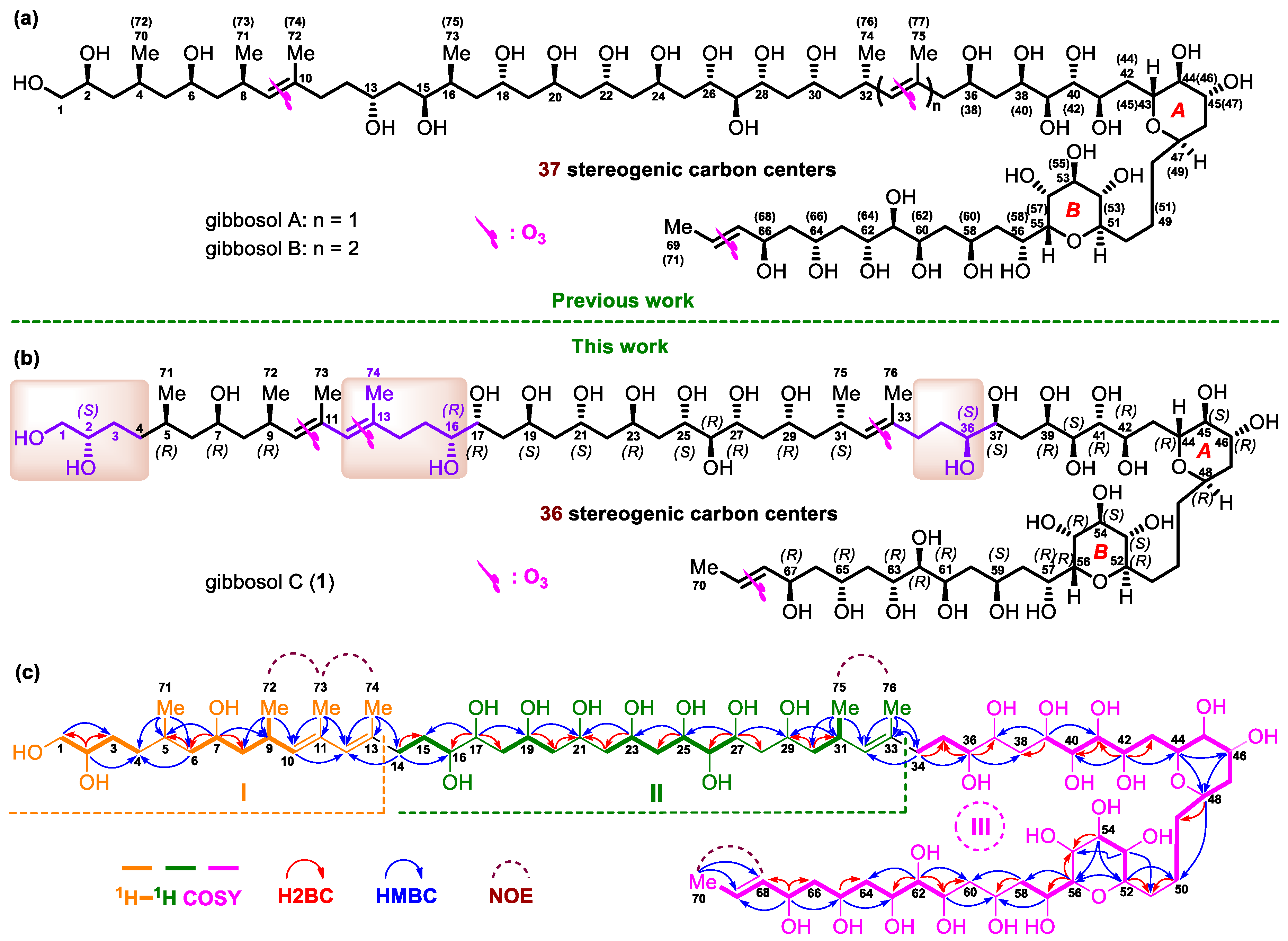
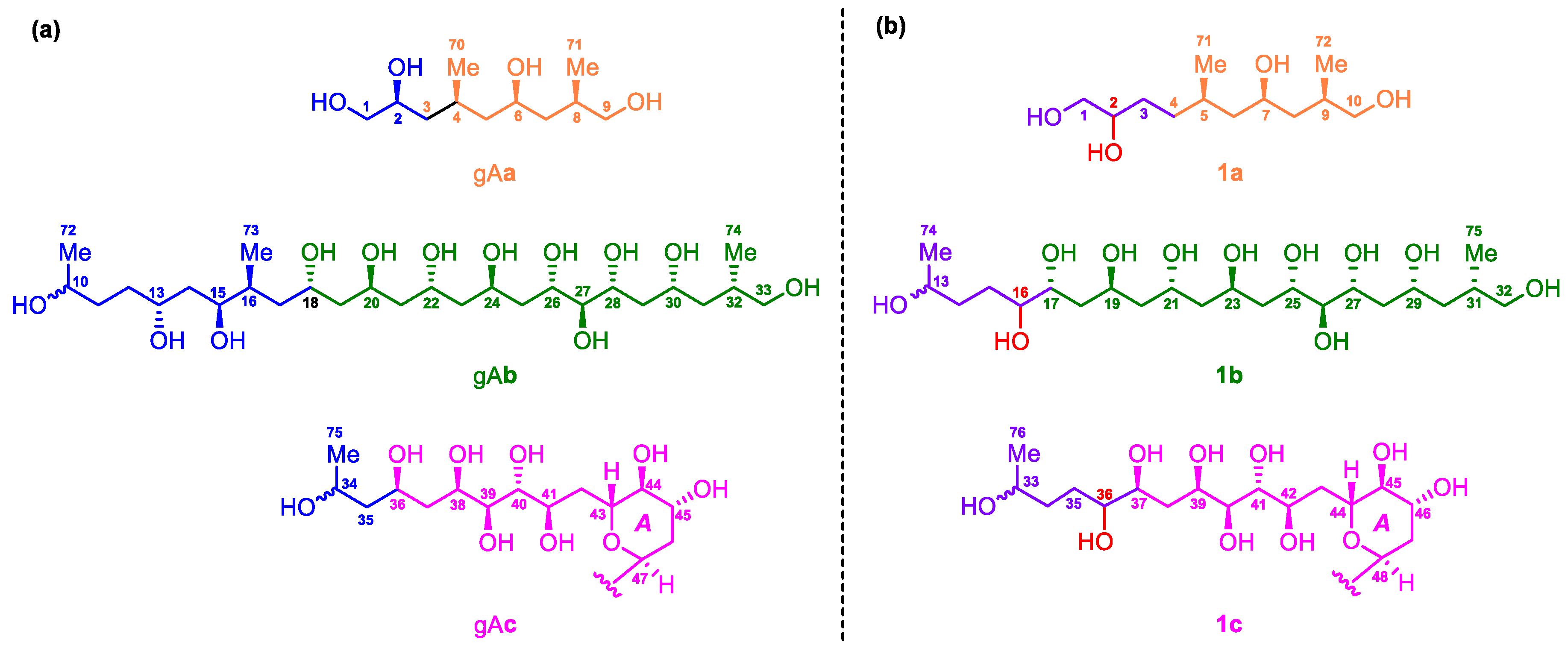
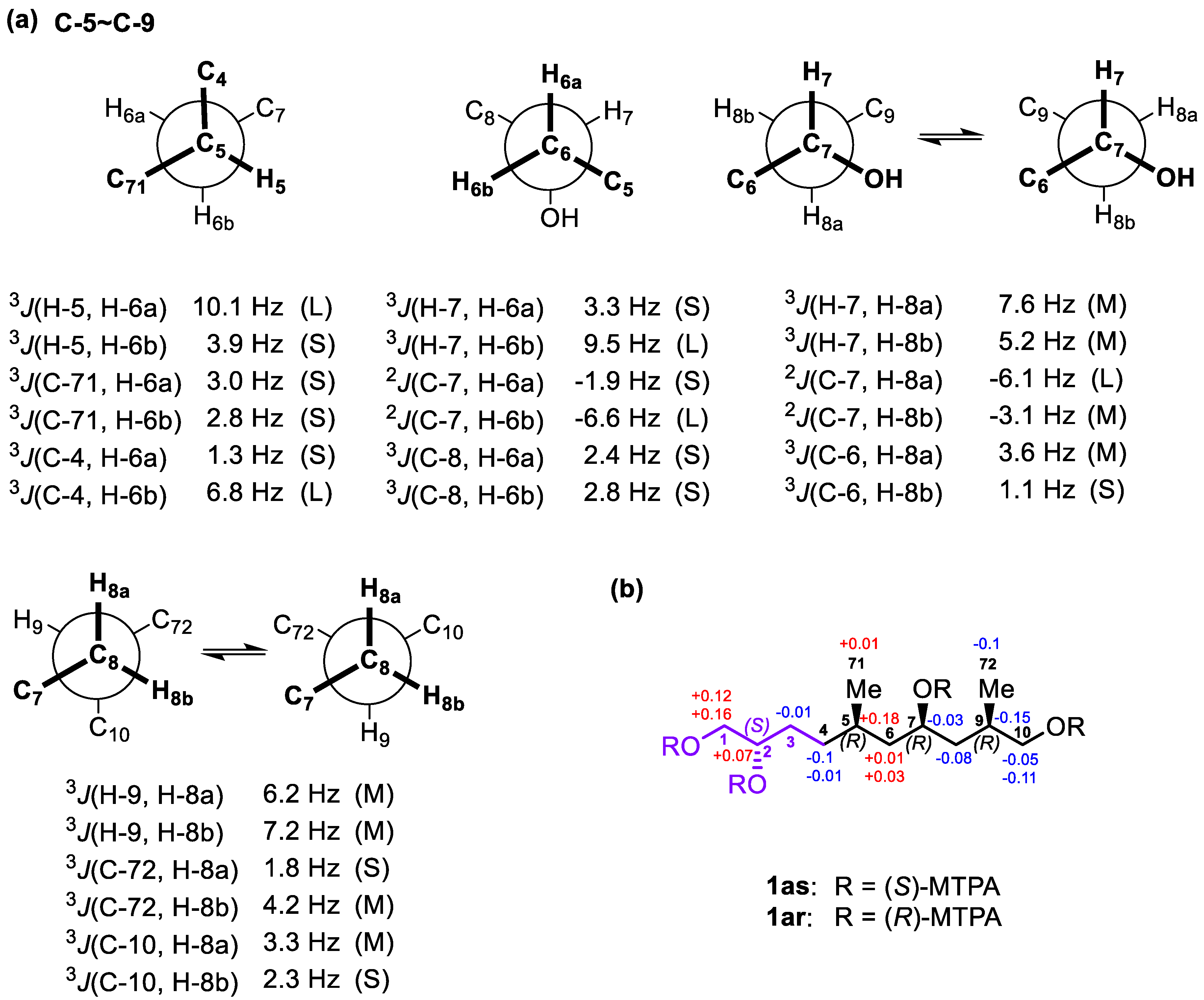
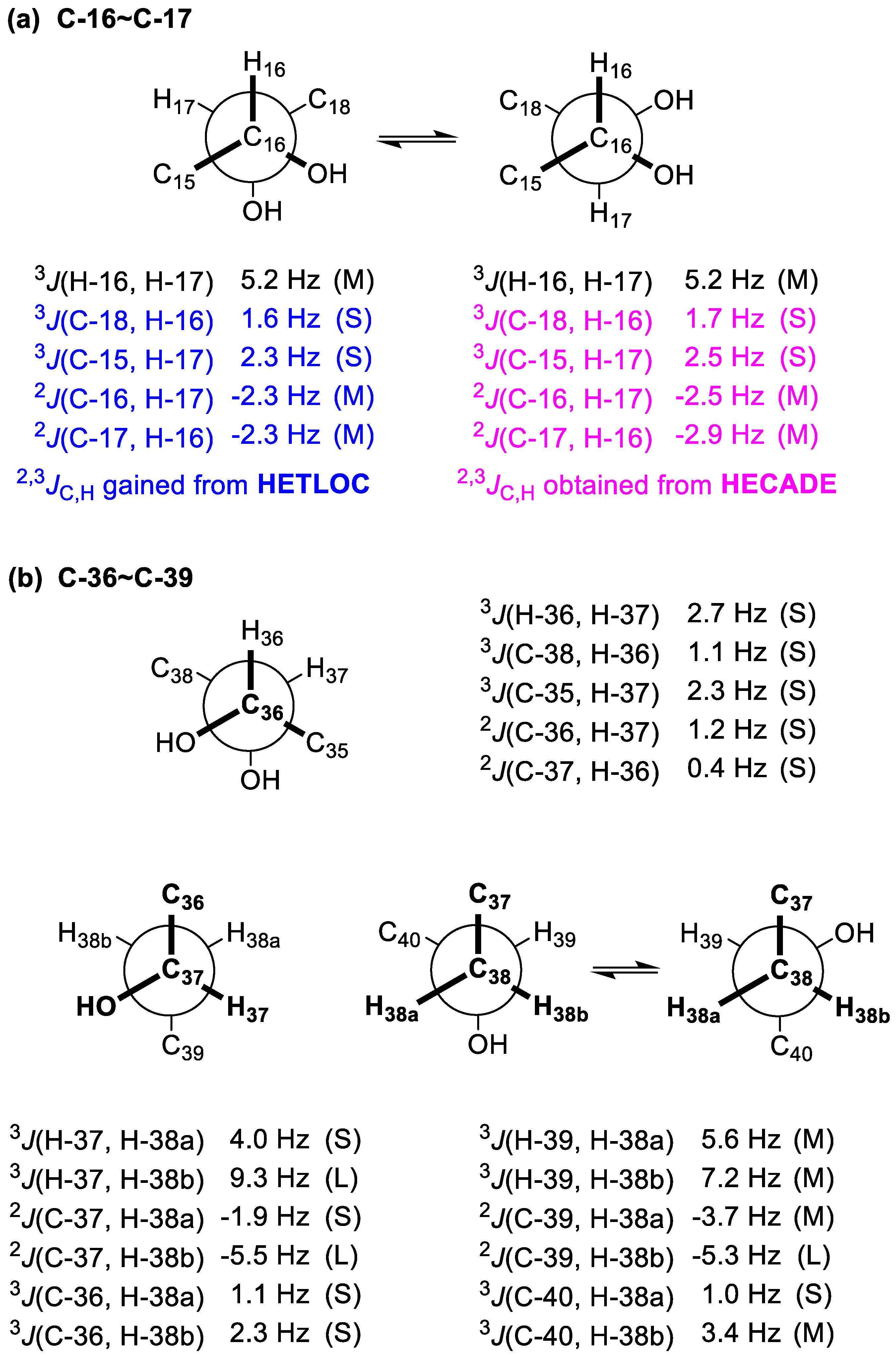
2.2. Relative and Absolute Configurations of Gibbosol C (1)
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Isolation of the Dinoflagellate and the Large-Scale Culture
3.3. Isolation of Gibbosol C (1)
3.4. Ozonolysis
3.5. Mosher’s MTPA Esters 1as and 1ar
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kobayashi, J.; Ishibashi, M. Bioactive metabolites of symbiotic marine microorganisms. Chem. Rev. 1993, 93, 1753–1769. [Google Scholar] [CrossRef]
- Kobayashi, J.; Kubota, T. Bioactive macrolides and polyketides from marine dinoflagellates of the genus Amphidinium. J. Nat. Prod. 2007, 70, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Uemura, D. Bioorganic studies on marine natural products-diverse chemical structures and bioactivities. Chem. Rec. 2006, 6, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Uemura, D. Marine huge molecules: The longest carbon chains in natural products. Chem. Rec. 2010, 10, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Murata, M.; Yasumoto, T.; Fujita, T.; Naoki, H. Amphidinol, a polyhydroxypolyene antifungal agent with an unprecedented structure, from a marine dinoflagellate, Ampbidnium klebsii. J. Am. Chem. Soc. 1991, 113, 9859–9861. [Google Scholar] [CrossRef]
- Murata, M.; Matsuoka, S.; Matsumori, N.; Paul, G.K.; Tachibana, K. Absolute configuration of amphidinol 3, the first complete structure determination from amphidinol homologues: Application of a new configuration analysis based on carbon-hydrogen spin-coupling constants. J. Am. Chem. Soc. 1999, 121, 870–871. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, D.; Guo, Y.; Wu, H.; Lin, L.; Wang, Z.; Ding, J.; Lin, Y. Lingshuiol, a novel polyhydroxyl compound with strongly cytotoxic activity from the marine dinoflagellate Amphidinium sp. Bioorg. Med. Chem. Lett. 2004, 14, 3117–3120. [Google Scholar]
- Huang, X.; Zhao, D.; Guo, Y.; Wu, H.; Trivellone, E.; Cimino, G. Lingshuiols A and B, two new polyhydroxy compounds from the Chinese marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 2004, 45, 5501–5504. [Google Scholar] [CrossRef]
- Washida, K.; Koyama, T.; Yamada, K.; Kita, M.; Uemura, D. Karatungiols A and B, two novel antimicrobial polyol compounds, from the symbiotic marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 2006, 47, 2521–2525. [Google Scholar] [CrossRef]
- Morsy, N.; Houdai, T.; Matsuoka, S.; Matsumori, N.; Adachi, S.; Oishi, T.; Murata, M.; Iwashita, T.; Fujita, T. Structures of new amphidinols with truncated polyhydroxyl chain and their membrane-permeabilizing activities. Bioorg. Med. Chem. 2006, 14, 6548–6554. [Google Scholar] [CrossRef]
- Huang, S.; Kuo, C.; Lin, Y.; Chen, Y.; Lu, C. Carteraol E, a potent polyhydroxyl ichthyotoxin from the dinoflagellate Amphidinium carterae. Tetrahedron Lett. 2009, 50, 2512–2515. [Google Scholar] [CrossRef]
- Hanif, N.; Ohno, O.; Kitamura, M.; Yamada, K.; Uemura, D. Symbiopolyol, a VCAM-1 inhibitor from a symbiotic dinoflagellate of the jellyfish Mastigias papua. J. Nat. Prod. 2010, 73, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Inuzuka, T.; Yamamoto, Y.; Yamada, K.; Uemura, D. Amdigenol A, a long carbon-backbone polyol compound, produced by the marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 2012, 53, 239–242. [Google Scholar] [CrossRef]
- Inuzuka, T.; Yamada, K.; Uemura, D. Amdigenols E and G, long carbon-chain polyol compounds, isolated from the marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 2014, 55, 6319–6323. [Google Scholar] [CrossRef]
- Satake, M.; Cornelio, K.; Hanashima, S.; Malabed, R.; Murata, M.; Matsumori, N.; Zhang, H.; Hayashi, F.; Mori, S.; Kim, J.S.; et al. Structures of the largest amphidinol homologues from the dinoflagellate Amphidinium carterae and structure-activity relationships. J. Nat. Prod. 2017, 80, 2883–2888. [Google Scholar] [CrossRef] [PubMed]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a new cytotoxic and antifungal amphidinol from the dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Deeds, J.R.; Satake, M.; Ribeiro, A.A.; Place, A.R.; Wright, J.L.C. Isolation and characterization of karlotoxin 1, a new amphipathic toxin from Karlodinium veneficum. Tetrahedron Lett. 2008, 49, 6457–6461. [Google Scholar] [CrossRef]
- Peng, J.; Place, A.R.; Yoshida, W.; Anklin, C.; Hamann, M.T. Structure and absolute configuration of karlotoxin-2, an ichthyotoxin from the marine dinoflagellate Karlodinium veneficum. J. Am. Chem. Soc. 2010, 132, 3277–3279. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Deeds, J.R.; Tatters, A.O.; Place, A.R.; Tomas, C.R.; Wright, J.L.C. Structure and relative potency of several karlotoxins from Karlodinium veneficum. J. Nat. Prod. 2010, 73, 1360–1365. [Google Scholar] [CrossRef]
- Waters, A.L.; Oh, J.; Place, A.R.; Hamann, M.T. Stereochemical studies of the karlotoxin class using NMR spectroscopy and DP4 chemical-shift analysis: Insights into their mechanism of action. Angew. Chem. Int. Ed. 2015, 54, 1–7. [Google Scholar] [CrossRef]
- Cai, P.; He, S.; Zhou, C.; Place, A.R.; Had, S.; Ding, L.; Chen, H.; Jiang, Y.; Guo, C.; Xu, Y.; et al. Two new karlotoxins found in Karlodinium veneficum (strain GM2) from the East China Sea. Harm. Algae 2016, 58, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.A.; Binzer, S.B.; Hoeck, C.; Meier, S.; de Medeiros, L.S.; Andersen, N.G.; Place, A.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Karmitoxin: An amine-containing polyhydroxy-polyene toxin from the marine dinoflagellate Karlodinium armiger. J. Nat. Prod. 2017, 80, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Espiritu, R.A.; Matsumori, N.; Tsuda, M.; Murata, M. Direct and stereospecific interaction of amphidinol 3 with sterol in lipid bilayers. Biochemistry 2014, 53, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.S.; Yoon, E.Y.; Jeong, E.J.; Park, J.; Kim, E.-H.; Rho, J.-R. Determination of the absolute configuration of polyhydroxy compound ostreol B isolated from the dinoflagellate Ostreopsis cf. ovate. J. Org. Chem. 2018, 83, 194–202. [Google Scholar] [CrossRef]
- Li, W.S.; Yan, R.J.; Yu, Y.; Shi, Z.; Mándi, A.; Shen, L.; Kurtán, T.; Wu, J. Determination of the absolute configuration of super-carbon-chain compounds by a combined chemical, spectroscopic, and computational approach: Gibbosols A and B. Angew. Chem. Int. Ed. 2020, 59, 13028–13036. [Google Scholar] [CrossRef]
- Hamamoto, Y.; Tachibana, K.; Holland, P.T.; Shi, F.; Beuzenberg, V.; Itoh, Y.; Satake, M. Brevisulcenal-F: A polycyclic ether toxin associated with massive fish-kills in New Zealand. J. Am. Chem. Soc. 2012, 134, 4963–4968. [Google Scholar] [CrossRef]
- Matsumori, N.; Kaneno, D.; Murata, M.; Nakamura, H.; Tachibana, K. Stereochemical determination of acyclic structures based on carbon-proton spin-coupling constants. A method of configuration analysis for natural products. J. Org. Chem. 1999, 64, 866–876. [Google Scholar] [CrossRef]
- Bifulco, G.; Dambruoso, P.; Gomez-Paloma, L.; Riccio, R. Determination of relative configuration in organic compounds by NMR spectroscopy and computational methods. Chem. Rev. 2007, 107, 3744–3779. [Google Scholar] [CrossRef]
- Ohtani, I.; Kusumi, T.; Kashman, Y.; Kakisawa, H. High-field FT NMR application of Mosher’s method. The absolute configurations of marine terpenoids. J. Am. Chem. Soc. 1991, 113, 4092–4096. [Google Scholar] [CrossRef]
- De Riccardis, F.; Minale, L.; Riccio, R.; Giovannitti, B.; Iorrizi, M.; Debitus, C. Phosphated and sulphated marine polyhydroxylated steroids from the starfish Tremaster novaecaledonìae. Gazz. Chim. Ital. 1993, 123, 79–86. [Google Scholar]
- Finamore, E.; Minale, L.; Riccio, R.; Rinaldo, G.; Zollo, F. Novel marine polyhydroxylated steroids from the starfish Myxoderma platyacanthum. J. Org. Chem. 1991, 56, 1146–1153. [Google Scholar] [CrossRef]
- Higashibayashi, S.; Czechtizky, W.; Kobayashi, Y.; Kishi, Y. Universal NMR databases for contiguous polyols. J. Am. Chem. Soc. 2003, 125, 14379–14393. [Google Scholar] [CrossRef] [PubMed]
- Seike, H.; Ghosh, I.; Kishi, Y. Attempts to assemble a universal NMR database without synthesis of NMR database compounds. Org. Lett. 2006, 8, 3861–3864. [Google Scholar] [CrossRef] [PubMed]
- Preiss, D.J.; Sattar, N. Vascular cell adhesion molecule-1: A viable therapeutic target for atherosclerosis? Int. J. Clin. Pract. 2007, 61, 697–701. [Google Scholar] [CrossRef]
- Jiao, H.L.; Zhang, Q.; Lin, Y.B.; Gao, Y.; Zhang, P. The ovotransferrin-derived peptide IRW attenuates lipopolysaccharide-induced inflammatory responses. BioMed Res. Int. 2019, 8676410. [Google Scholar] [CrossRef]
- Park, J.-G.; Ryu, S.Y.; Jung, I.-H.; Lee, Y.-H.; Kang, K.J.; Lee, M.-R.; Lee, M.-N.; Sonn, S.K.; Lee, J.H.; Lee, H.; et al. Evaluation of VCAM-1 antibodies as therapeutic agent for atherosclerosis in apolipoprotein E-deficient mice. Atherosclerosis 2013, 226, 356–363. [Google Scholar] [CrossRef]
- Ling, S.; Nheu, L.; Komesaroff, P.A. Cell adhesion molecules as pharmaceutical target in atherosclerosis. Mini-Rev. Med. Chem. 2012, 12, 175–183. [Google Scholar] [CrossRef]
- Cutignano, A.; Nuzzo, G.; Sardo, A.; Fontana, A. The missing piece in biosynthesis of amphidinols: First evidence of glycolate as a starter unit in new polyketides from Amphidinium carterae. Mar Drugs 2017, 15, 157. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Satake, M.; Wright, J.L.C. Polyketide biosynthesis in dinoflagellates: What makes it different? Nat. Prod. Rep. 2014, 31, 1101–1137. [Google Scholar] [CrossRef]
- Meng, Y.; Van Wagoner, R.M.; Misner, I.; Tomas, C.; Wright, J.L.C. Structure and biosynthesis of amphidinol 17, a hemolytic compound from Amphidinium carterae. J. Nat. Prod. 2010, 73, 409–415. [Google Scholar] [CrossRef]
- Kellman, R.; Stüken, A.; Orr, R.J.S.; Svendsen, H.M.; Jakobsen, K.S. Biosynthesis and molecular genetics of polyketides in marine dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | δH (J in Hz) | δC, Type |
|---|---|---|
| 1a | 3.41, m | 67.4, CH2 |
| 1b | 3.45, m | |
| 2 | 3.53, m | 73.6, CH |
| 3a | 1.39, m | 31.9, CH2 |
| 3b | 1.48, m | |
| 4a | 1.32, m | 34.8, CH2 |
| 4b | 1.32, m | |
| 5 | 1.65, m f | 30.5, CH |
| 6a | 1.12, m | 47.1, CH2 |
| 6b | 1.46, m | |
| 7 | 3.62, m | 68.5, CH |
| 8a | 1.28, m | 47.4, CH2 |
| 8b | 1.43, m a | |
| 9 | 2.74, m | 30.6, CH |
| 10 | 4.92, br d (9.8) i | 136.2, CH |
| 11 | 133.3, qC | |
| 12 | 5.67, br s | 130.3, CH |
| 13 | 136.3, qC | |
| 14a | 2.08, m | 37.7, CH2 |
| 14b | 2.26, m | |
| 15a | 1.47, m | 32.3, CH2 |
| 15b | 1.74, m | |
| 16 | 3.42, m | 75.81, CH |
| 17 | 3.70, m | 72.8, CH |
| 18a | 1.50, m | 41.2, CH2 |
| 18b | 1.67, m e | |
| 19 | 4.10, m | 66.5, CH |
| 20a | 1.58, m b | 46.9, CH2 |
| 20b | 1.58, m b | |
| 21 | 4.09, m j | 66.4, CH |
| 22a | 1.58, m b | 46.9, CH2 |
| 22b | 1.58, m b | |
| 23 | 4.12, m | 66.3, CH |
| 24a | 1.54, m | 41.5, CH2 |
| 24b | 1.79, m | |
| 25 | 3.88, m | 70.5, CH |
| 26 | 3.37 t, (6.3) | 78.8, CH |
| 27 | 3.83, m | 73.1, CH |
| 28a | 1.58, m b | 41.0, CH2 |
| 28b | 1.80, m | |
| 29 | 3.78, m | 70.2, CH |
| 30a | 1.35, m | 46.8, CH2 |
| 30b | 1.43, m a | |
| 31 | 2.69, m | 30.1, CH |
| 32 | 4.92, br d (9.8) i | 132.1, CH |
| 33 | 135.5, CH | |
| 34a | 2.04, m | 37.1, CH2 |
| 34b | 2.19, m | |
| 35a | 1.55, m c | 32.44, CH2 |
| 35b | 1.69, m | |
| 36 | 3.44, m | 74.7, CH |
| 37 | 3.69, m | 73.5, CH |
| 38a | 1.77, m g | 37.4, CH2 |
| 38b | 1.81, m | |
| 39 | 4.13, m | 70.36, CH |
| 40 | 3.46, m | 74.6, CH |
| 41 | 3.69, m | 75.7, CH |
| 42 | 4.04, m | 70.06, CH |
| 43a | 1.77, m g | 34.9, CH2 |
| 43b | 1.98, m | |
| 44 | 3.67, m | 70.9, CH |
| 45 | 3.03, t (8.4) | 77.6, CH |
| 46 | 3.75, m | 70.4, CH |
| 47a | 1.70, m | 37.6, CH2 |
| 47b | 1.88, m | |
| 48 | 3.93, m | 73.2, CH |
| 49a | 1.38, m | 32.4, CH2 |
| 49b | 1.91, m | |
| 50a | 1.53, m | 23.4, CH2 |
| 50b | 1.53, m | |
| 51a | 1.52, m d | 32.8, CH2 |
| 51b | 1.75, m | |
| 52 | 3.41, m | 76.5, CH |
| 53 | 3.14, t (7.7) | 75.2, CH |
| 54 | 3.72, t (7.7) | 75.1, CH |
| 55 | 3.78, m | 73.8, CH |
| 56 | 3.60, dd (9.1, 4.9) | 75.76, CH |
| 57 | 4.39, m | 67.7, CH |
| 58a | 1.52, m d | 42.1, CH2 |
| 58b | 1.85, m h | |
| 59 | 4.09, m j | 67.1, CH |
| 60a | 1.67, m e | 42.7, CH2 |
| 60b | 1.85, m h | |
| 61 | 4.09, m | 69.7, CH |
| 62 | 3.24, dd (7.7, 1.4) | 77.1, CH |
| 63 | 3.82, m | 71.9, CH |
| 64a | 1.59, m | 42.2, CH2 |
| 64b | 1.94, m | |
| 65 | 4.08, m j | 68.7, CH |
| 66a | 1.55, m c | 45.7, CH2 |
| 66b | 1.65, m f | |
| 67 | 4.26, m | 70.1, CH |
| 68 | 5.51, ddq (14.7, 6.3, 1.4) | 135.9, CH |
| 69 | 5.66, m | 126.4, CH |
| 70 | 1.68, br d (6.3) | 17.9, CH3 |
| 71 | 0.88, d (6.3) | 19.7, CH3 |
| 72 | 0.96, d (6.3) | 22.2, CH3 |
| 73 | 1.72, br s | 17.6, CH3 |
| 74 | 1.76, br s | 18.1, CH3 |
| 75 | 0.94, d (7.0) | 22.4, CH3 |
| 76 | 1.68, br s | 16.7, CH3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.-S.; Luo, Z.; Zhu, Y.-L.; Yu, Y.; Wu, J.; Shen, L. A Polyol-Polyol Super-Carbon-Chain Compound Containing Thirty-Six Carbon Stereocenters from the Dinoflagellate Amphidinium gibbosum: Absolute Configuration and Multi-Segment Modification. Mar. Drugs 2020, 18, 590. https://doi.org/10.3390/md18120590
Li W-S, Luo Z, Zhu Y-L, Yu Y, Wu J, Shen L. A Polyol-Polyol Super-Carbon-Chain Compound Containing Thirty-Six Carbon Stereocenters from the Dinoflagellate Amphidinium gibbosum: Absolute Configuration and Multi-Segment Modification. Marine Drugs. 2020; 18(12):590. https://doi.org/10.3390/md18120590
Chicago/Turabian StyleLi, Wan-Shan, Zeng Luo, Yan-Lan Zhu, Yi Yu, Jun Wu, and Li Shen. 2020. "A Polyol-Polyol Super-Carbon-Chain Compound Containing Thirty-Six Carbon Stereocenters from the Dinoflagellate Amphidinium gibbosum: Absolute Configuration and Multi-Segment Modification" Marine Drugs 18, no. 12: 590. https://doi.org/10.3390/md18120590
APA StyleLi, W.-S., Luo, Z., Zhu, Y.-L., Yu, Y., Wu, J., & Shen, L. (2020). A Polyol-Polyol Super-Carbon-Chain Compound Containing Thirty-Six Carbon Stereocenters from the Dinoflagellate Amphidinium gibbosum: Absolute Configuration and Multi-Segment Modification. Marine Drugs, 18(12), 590. https://doi.org/10.3390/md18120590