Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity



Abstract
1. Introduction
2. Results
2.1. Preparation of Sulfated Glucuronomannan Oligosaccharides
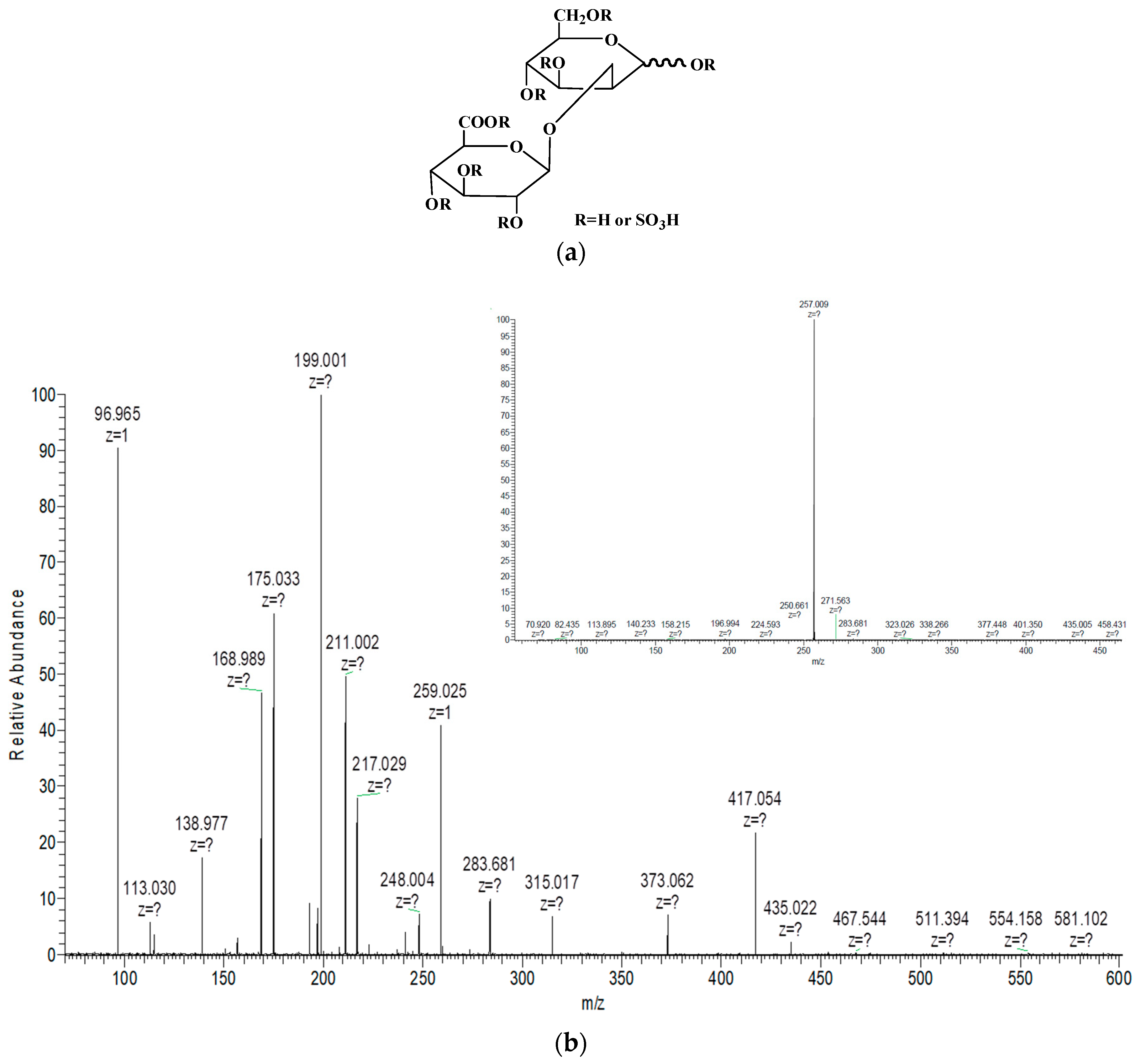
2.2. Structural Analysis of Sulfated Glucuronomannan Oligosaccharides by ESI-CID-MS/MS
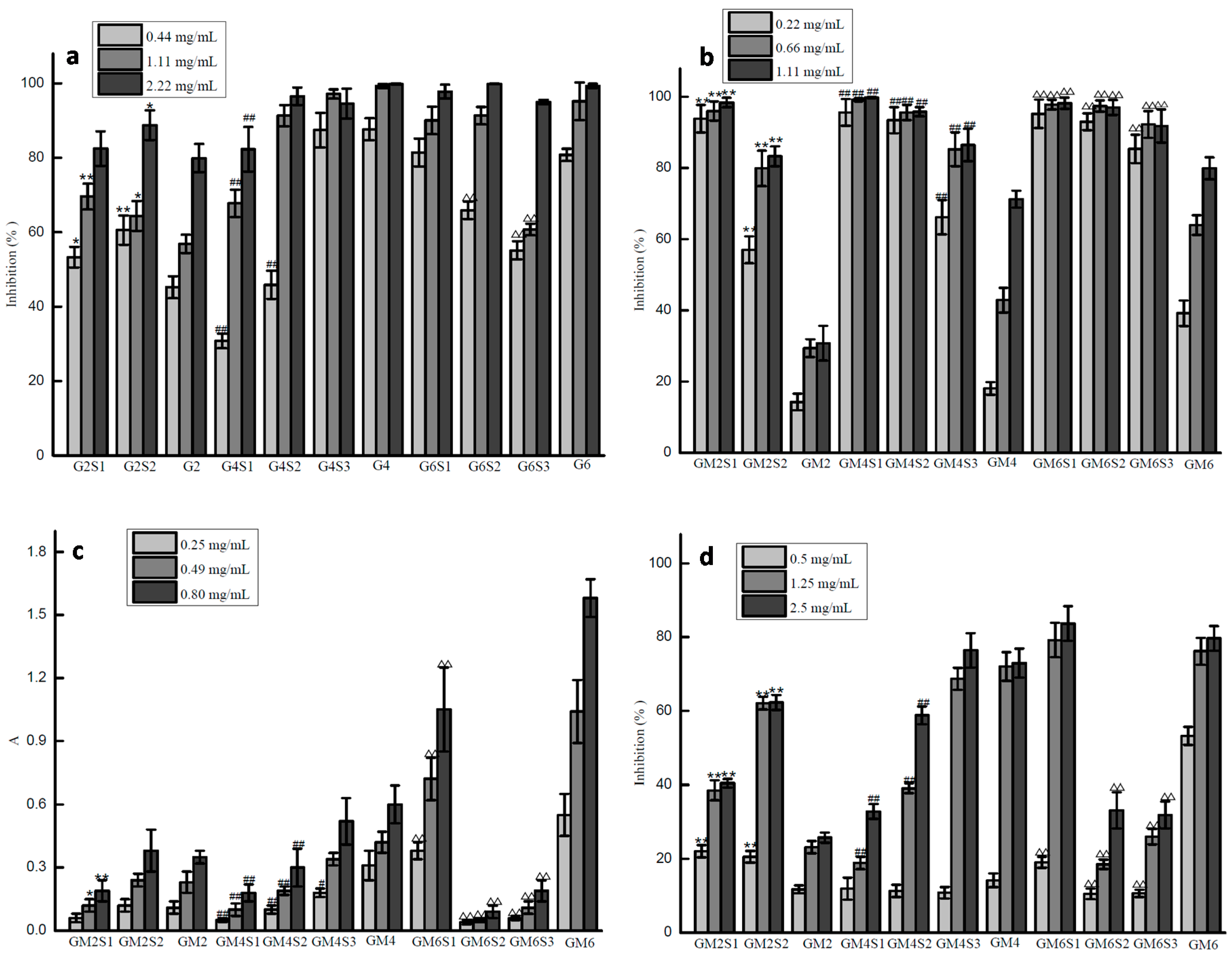
2.3. Antioxidant Activities of Glucuronomannan Oligosaccharides and Their Sulfated Fractions
3. Discussion
4. Materials and Methods
4.1. Preparation of Glucuronomannan Oligosaccharides
4.2. Preparation of Sulfated Glucuronomannan Oligosaccharides
4.3. MS Analysis of Oligosaccharides
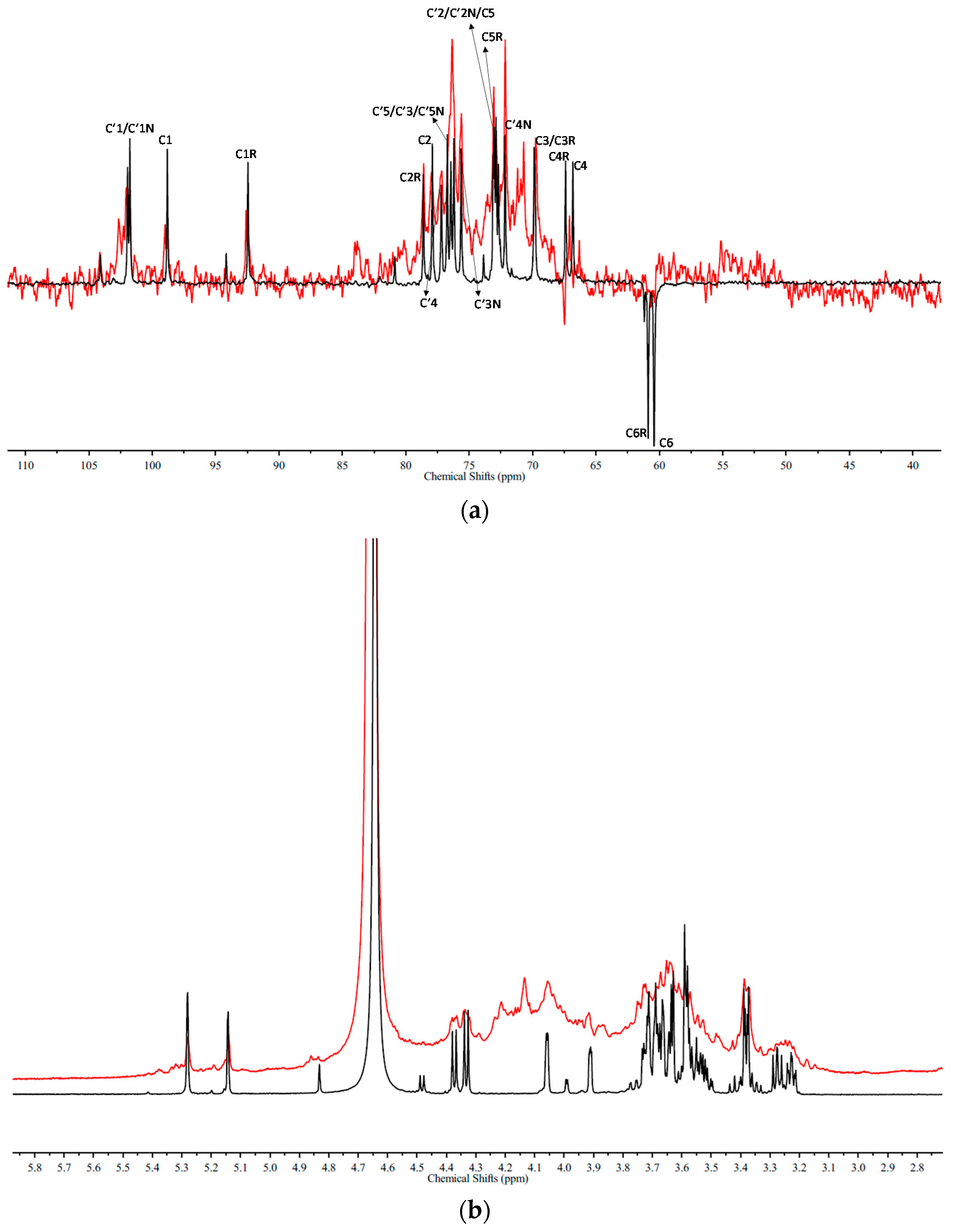
4.4. NMR Spectroscopy
4.5. Investigation of Antioxidant Activity
4.5.1. The Activity of Scavenging Hydroxyl Radicals
4.5.2. The Activity of Scavenging Superoxide Radicals
4.5.3. The Reducing Power Assay
4.5.4. DPPH Radical Scavenging Activity
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, R. Enzyme and microbial technology for synthesis of bioactive oligosaccharides: An update. Appl. Microbiol. Biotechnol. 2018, 102, 3017–3026. [Google Scholar] [CrossRef] [PubMed]
- McCranie, E.K.; Bachmann, B.O. Bioactive oligosaccharide natural products. Nat. Prod. Rep. 2014, 31, 1026–1042. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Wang, R.F.; Liu, B. An update on oligosaccharides and their esters from traditional Chinese medicines: Chemical structures and biological activities. Evid. Based Complement. Altern. Med. 2015, 2015, 512675. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Chang, P.; Li, X.; Gao, Z.; Sun, Y. The inhibitory effect of chitosan oligosaccharides on beta-site amyloid precursor protein cleaving enzyme 1 (BACE1) in HEK293 APPswe cells. Neurosci. Lett. 2018, 665, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Jeddou, K.B.; Bouaziz, F.; Helbert, C.B.; Nouri-Ellouz, O.; Maktouf, S.; Ellouz-Chaabouni, S.; Ellouz-Ghorbel, R. Structural, functional, and biological properties of potato peel oligosaccharides. Int. J. Biol. Macromol. 2018, 112, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Usov, A.I.; Bilan, M.I. Fucoidans-sulfatedd polysaccharides of brown algae. Russ. Chem. Rev. 2009, 78, 785–799. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H. Therapies from Fucoidan; Multifunctional Marine Polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Mourão, P.A.S. Structure, biology, evolution, and medical importance of sulfated fucans and galactans. Glycobiology 2008, 18, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Kelly, M.; Sanderson, C.J.; Nifantiev, N.E.; Usov, A.I. Further studies on the composition and structure of a fucoidan preparation from the brown alga Saccharina latissima. Carbohydr. Res. 2010, 345, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, J.; Ren, S.; Song, N.; Zhang, Q. Structural analysis of a heteropolysaccharide from Saccharina japonica by electrospray mass spectrometry in tandem with collision-induced dissociation tandem mass spectrometry (ESI-CID-MS/MS). Mar. Drugs 2012, 10, 2138–2152. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.D.; Lv, Y.J.; Liu, X.X.; Zhao, X.L.; Jiao, G.L.; Tai, W.J.; Wang, P.P.; Zhao, X.; Cai, C.; Yu, G.L. Structural Study of Sulfated Fuco-Oligosaccharide Branched Glucuronomannan from Kjellmaniella crassifolia by ESI-CID-MS/MS. J. Carbohydr. Chem. 2015, 34, 303–317. [Google Scholar] [CrossRef]
- Haroun-Bouhedja, F.; Ellouali, M.; Sinquin, C.; Boisson-Vidal, C. Relationship between Sulfate Groups and Biological Activities of Fucans. Thromb. Res. 2010, 100, 453–459. [Google Scholar] [CrossRef]
- Karmakar, P.; Pujol, C.A.; Damonte, E.B.; Ghosh, T.; Ray, B. Polysaccharides from Padina tetrastromatica: Structural features, chemical modification and antiviral activity. Carbohydr. Polym. 2010, 80, 513–520. [Google Scholar] [CrossRef]
- Mestechkina, N.M.; Shcherbukhin, V.D. Sulfated Polysaccharides and Their Anticoagulant Activity:A Review. Appl. Biochem. Microbiol. 2010, 46, 291–298. [Google Scholar] [CrossRef]
- Jin, W.H.; Zhang, W.J.; Liang, H.Z.; Zhang, Q.B. The Structure-Activity Relationship between Marine Algae Polysaccharides and Anti-Complement Activity. Mar. Drugs 2016, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Teruya, T.; Konishi, T.; Uechi, S.; Tamaki, H.; Tako, M. Anti-proliferative activity of oversulfated fucoidan from commercially cultured Cladosiphon okamuranus TOKIDA in U937 cells. Int. J. Biol. Macromol. 2007, 41, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.-M.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–156. [Google Scholar] [CrossRef]
- Saad, O.M.; Leary, J.A. Delineating Mechanisms of Dissociation for Isomeric Heparin Disaccharides Using Isotope Labeling and Ion Trap Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2004, 15, 1274–1286. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Shon, M.-Y.; Kim, T.-H.; Sung, N.-J. Antioxidants and free radical scavenging activity of Phellinus baumii (Phellinus of Hymenochaetaceae) extracts. Food Chem. 2003, 82, 593–597. [Google Scholar] [CrossRef]
- Zhu, X.; Lee, H.G.; Casadesus, G.; Avila, J.; Drew, K.; Perry, G.; Smith, M.A. Oxidative imbalance in Alzheimer’s disease. Mol. Neurobiol. 2005, 31, 205–217. [Google Scholar] [CrossRef]
- Wade, C.R.; Jackson, P.G.; Highton, J.; van Rij, A.M. Lipid peroxidation and malondialdehyde in the synovial fluid and plasma of patients with rheumatoid arthritis. Clin. Chim. Acta 1987, 164, 245–250. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, W.; Liu, G.; Yao, J.; Shan, T.; Sun, C.; Zhang, Q. The structure-activity relationship between polysaccharides from Sargassum thunbergii and anti-tumor activity. Int. J. Biol. Macromol. 2017, 105, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, H.Y. Antioxidant Activity of Various Tea Extracts in Relation to Their Antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Chemical Compositions |
|---|---|
| G2S1 | GlcAMan(SO3H)3–6 |
| G2S2 | GlcAMan(SO3H)1–3 |
| G2 | GlcAMan |
| G4S1 | GlcA2Man2(SO3H)8–11 |
| G4S2 | GlcA2Man2(SO3H)5–9 |
| G4S3 | GlcA2Man2(SO3H)1–5 |
| G4 | GlcA2Man2 |
| G6S1 | GlcA3Man3(SO3H)8–15 |
| G6S2 | GlcA3Man3(SO3H)4–10 |
| G6S3 | GlcA3Man3(SO3H)1–6 |
| G6 | GlcA3Man3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W.; Ren, L.; Liu, B.; Zhang, Q.; Zhong, W. Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity. Mar. Drugs 2018, 16, 291. https://doi.org/10.3390/md16090291
Jin W, Ren L, Liu B, Zhang Q, Zhong W. Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity. Marine Drugs. 2018; 16(9):291. https://doi.org/10.3390/md16090291
Chicago/Turabian StyleJin, Weihua, Langlang Ren, Bing Liu, Quanbin Zhang, and Weihong Zhong. 2018. "Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity" Marine Drugs 16, no. 9: 291. https://doi.org/10.3390/md16090291
APA StyleJin, W., Ren, L., Liu, B., Zhang, Q., & Zhong, W. (2018). Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity. Marine Drugs, 16(9), 291. https://doi.org/10.3390/md16090291