
Nornidulin, A New Inhibitor of Plasmodium falciparum Malate: Quinone Oxidoreductase (PfMQO) from Indonesian Aspergillus sp. BioMCC f.T.8501

 , , , , and
, , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
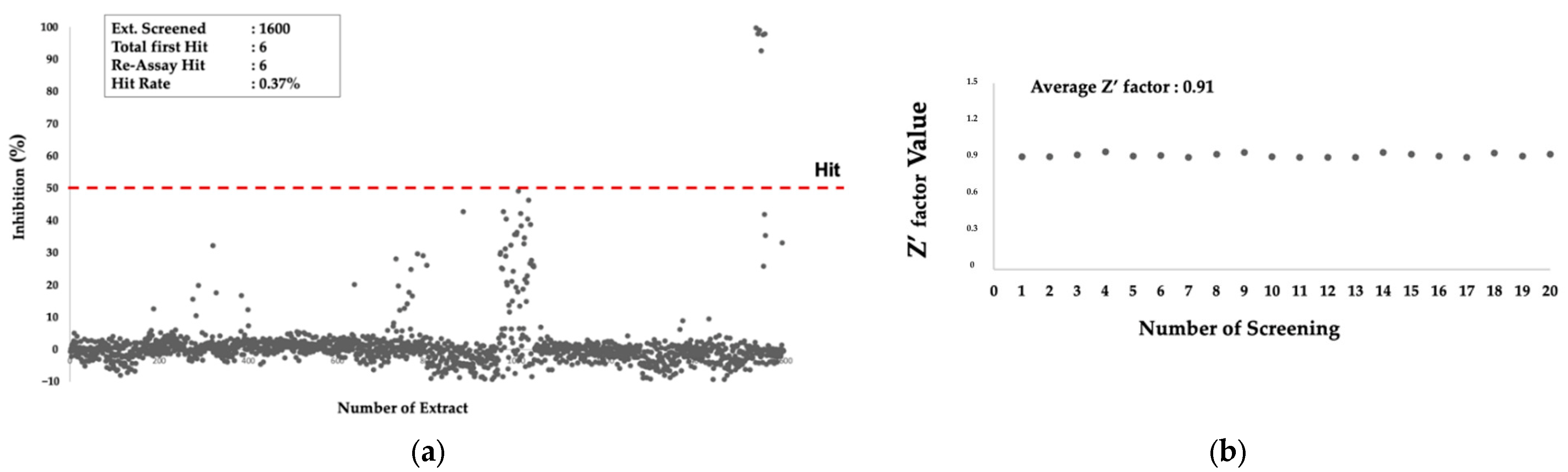
2.1. HTS to Obtain Active Microbial Extract against PfMQO
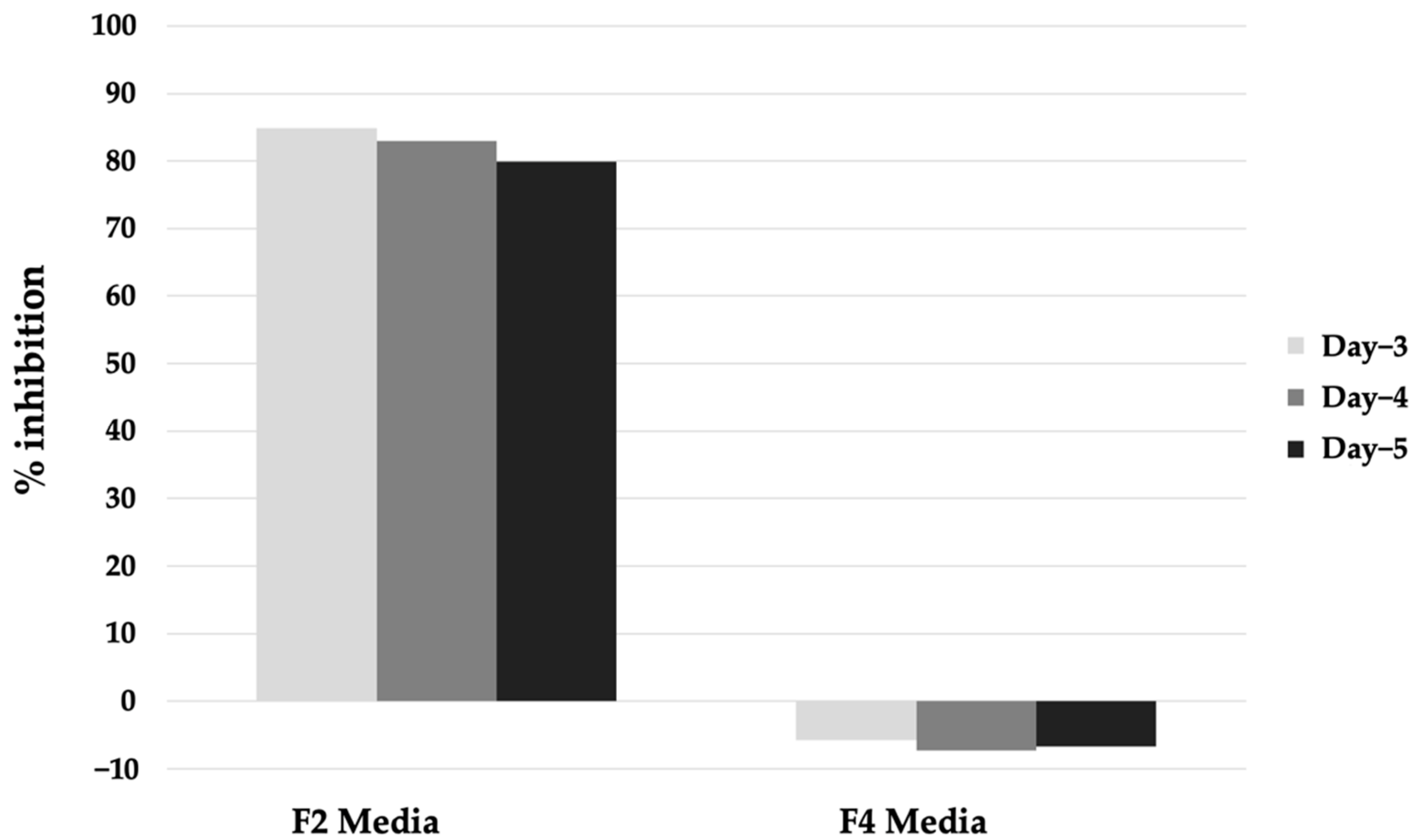
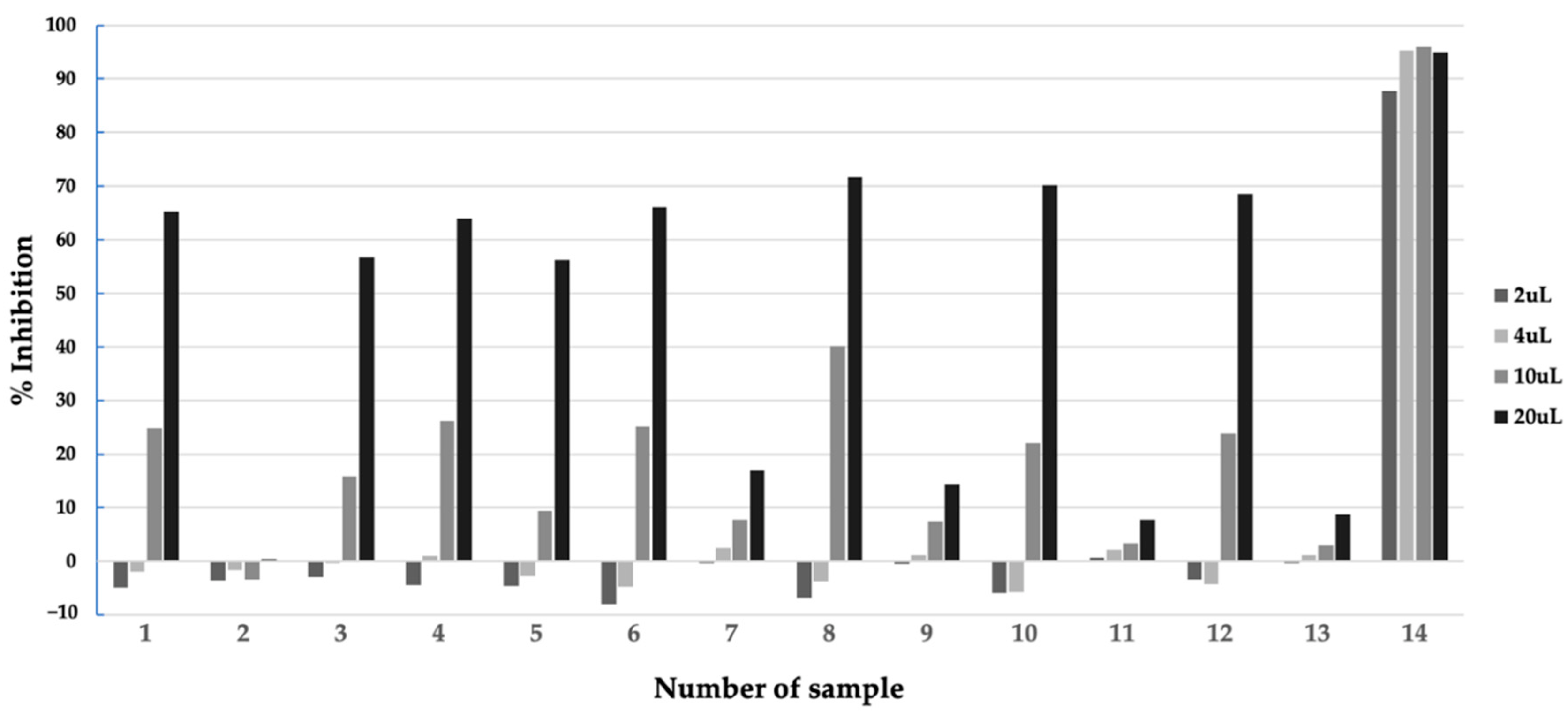
2.2. Preliminary Extraction Test
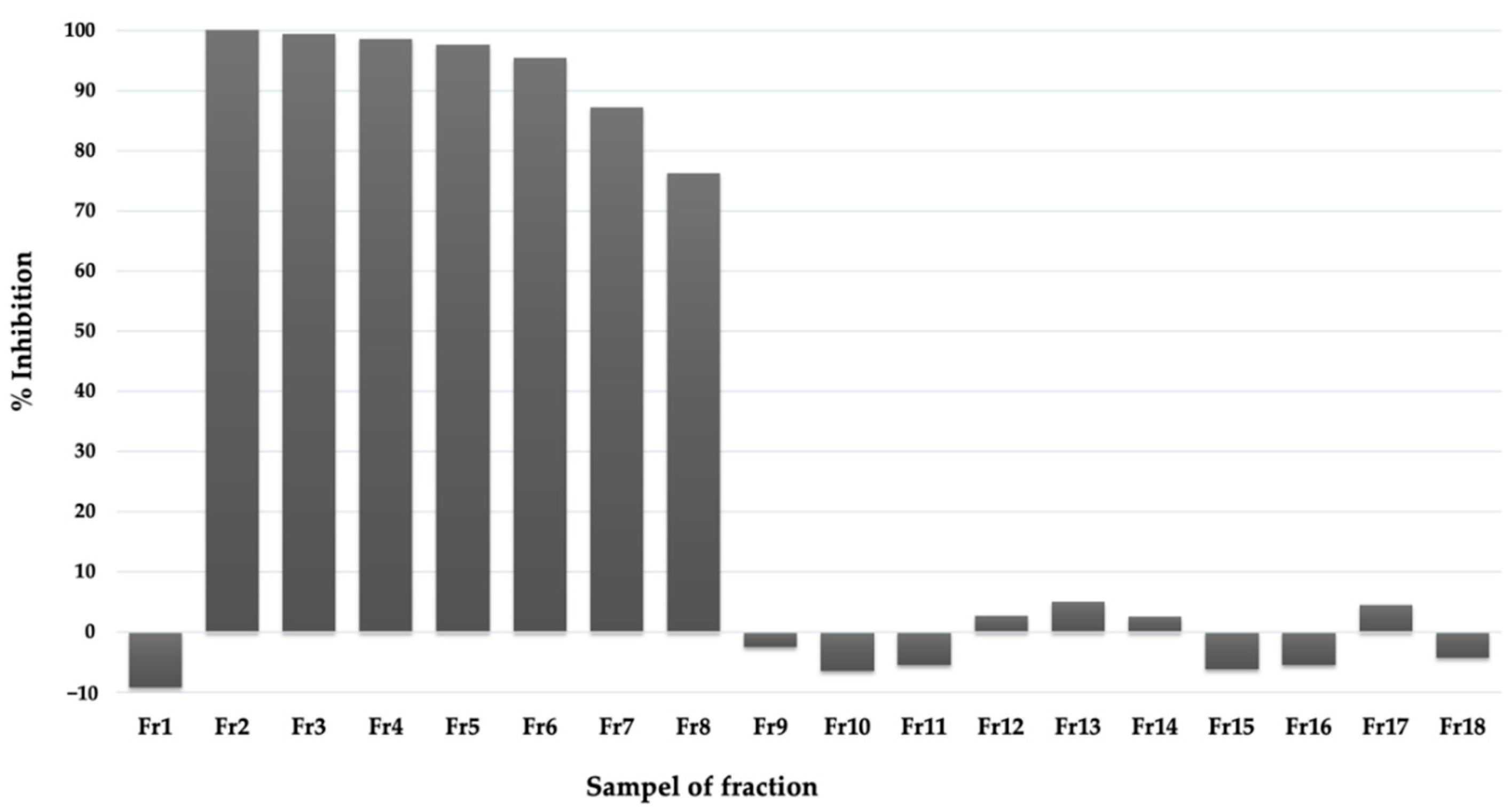
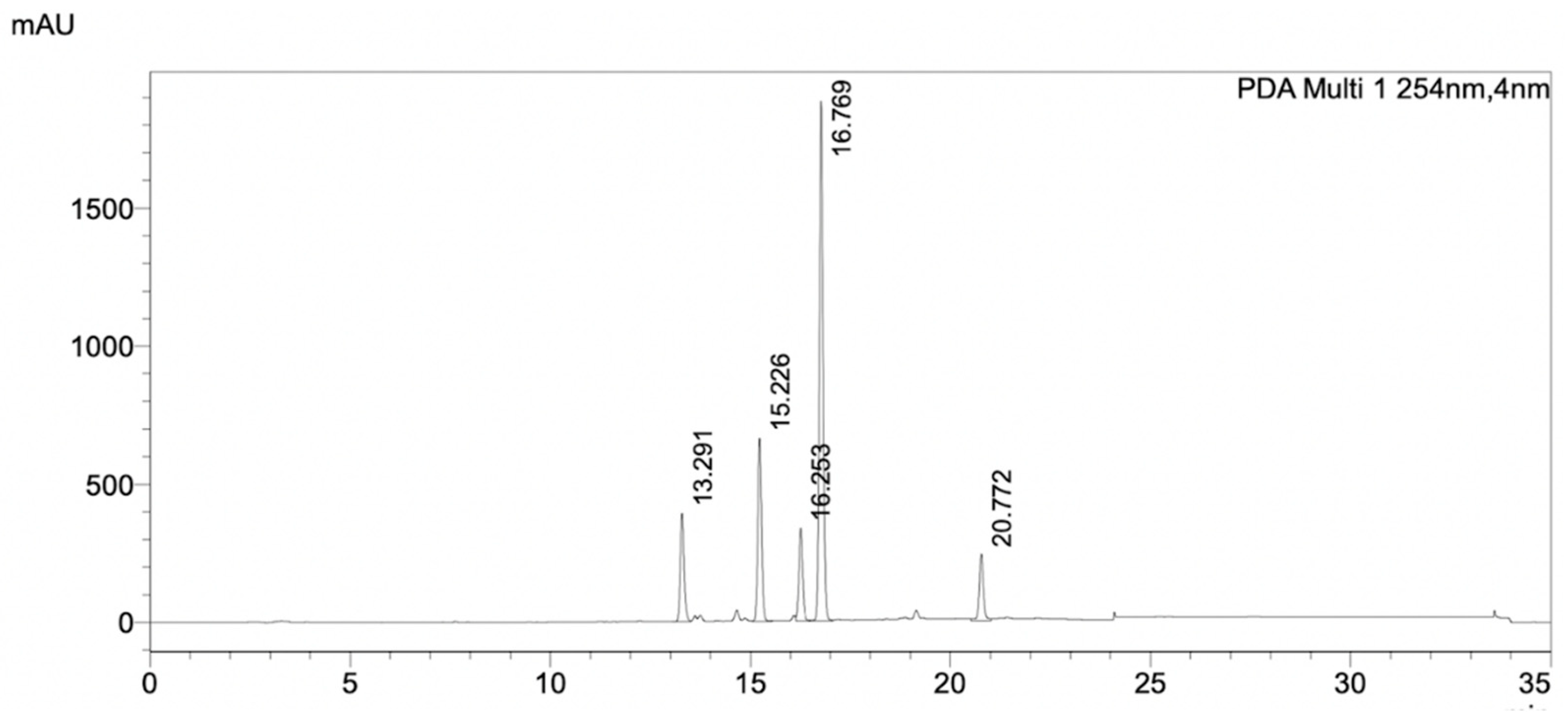
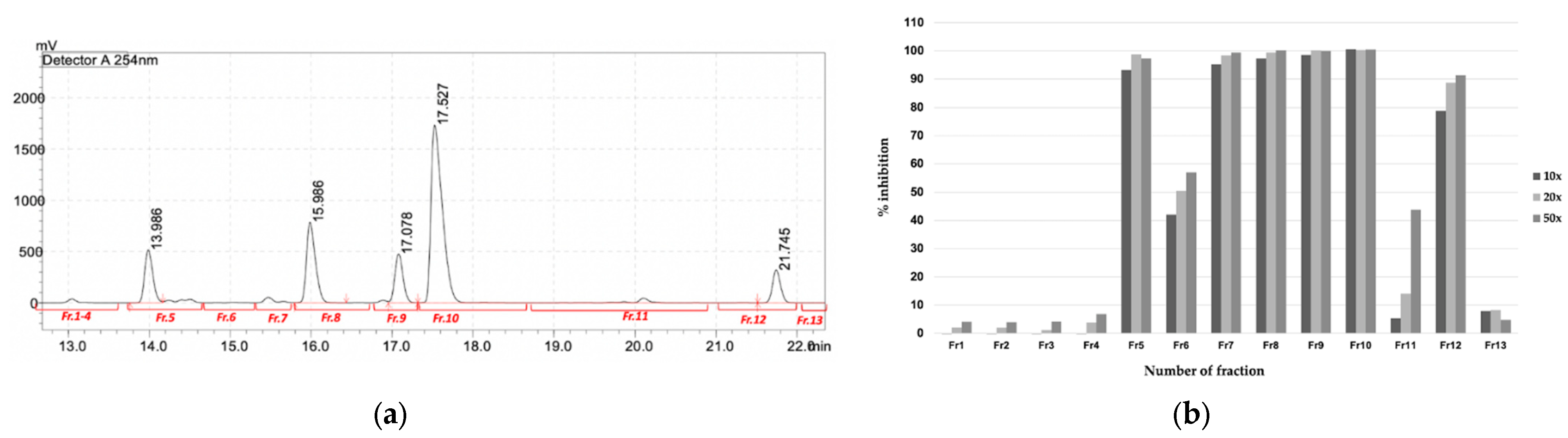
2.3. Purification and Identification of Bioactive Compounds
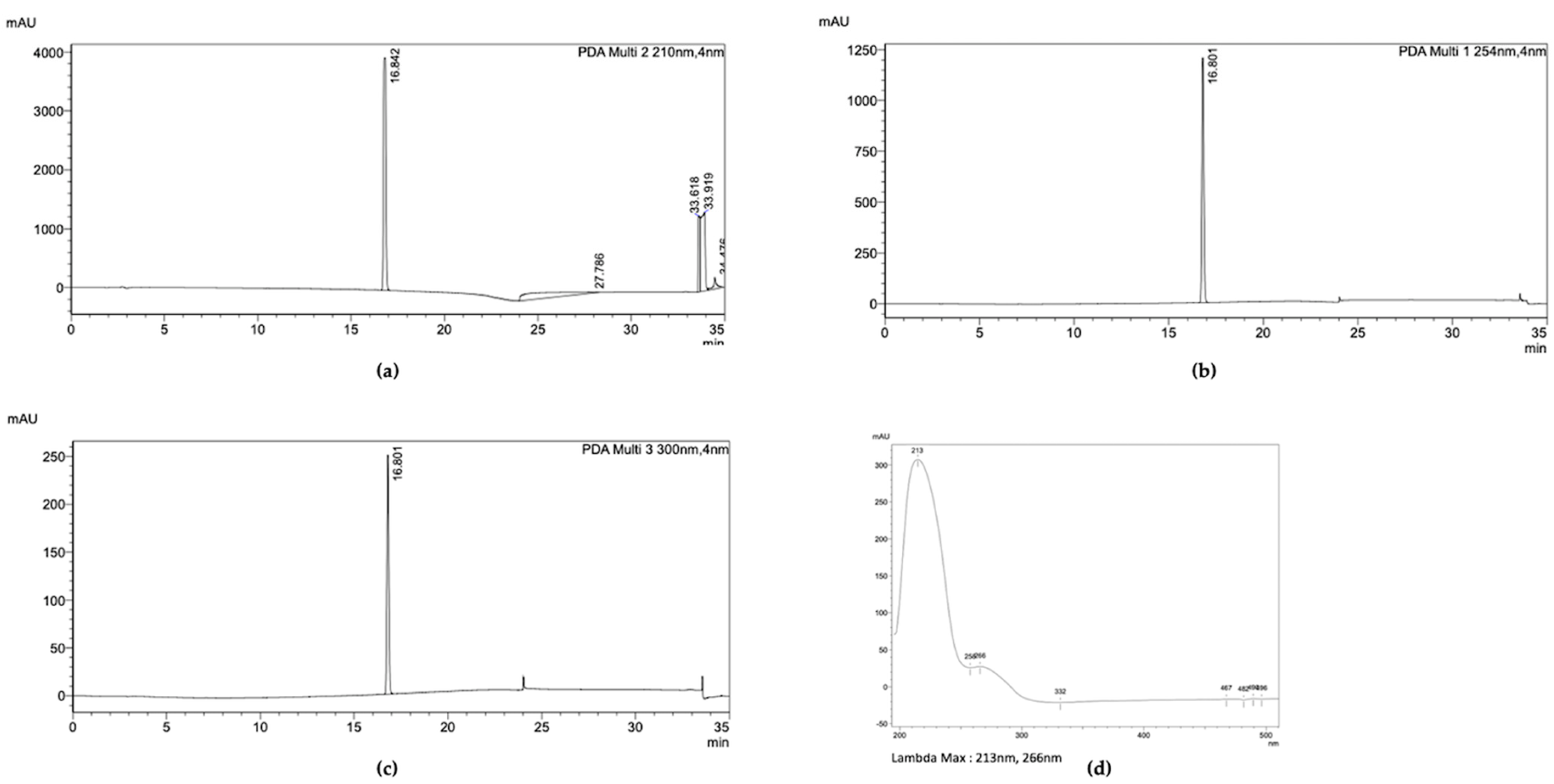
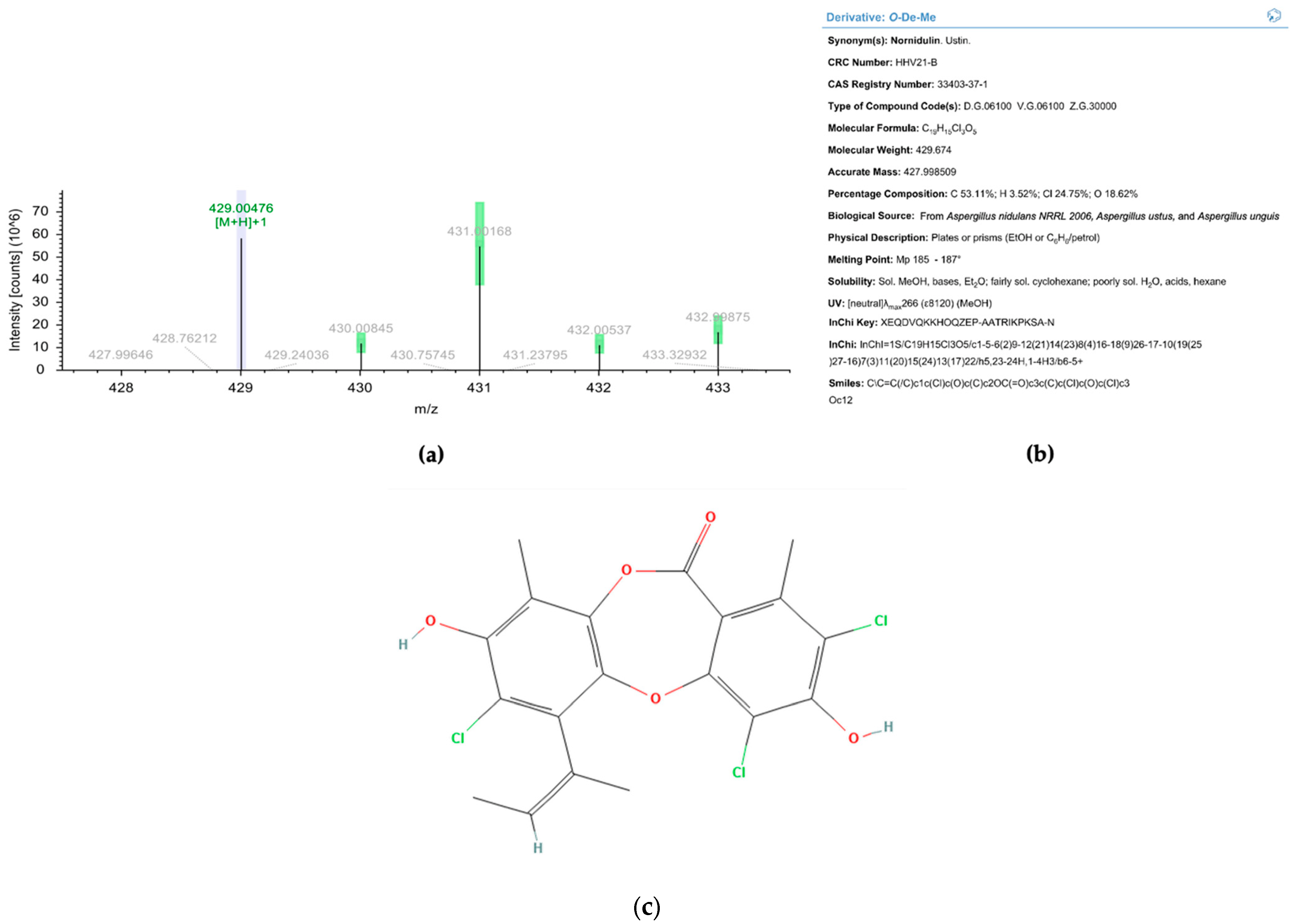
2.4. Characterization of the Purified Compound
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Microbial Extract Preparation
4.3. PfMQO Assay
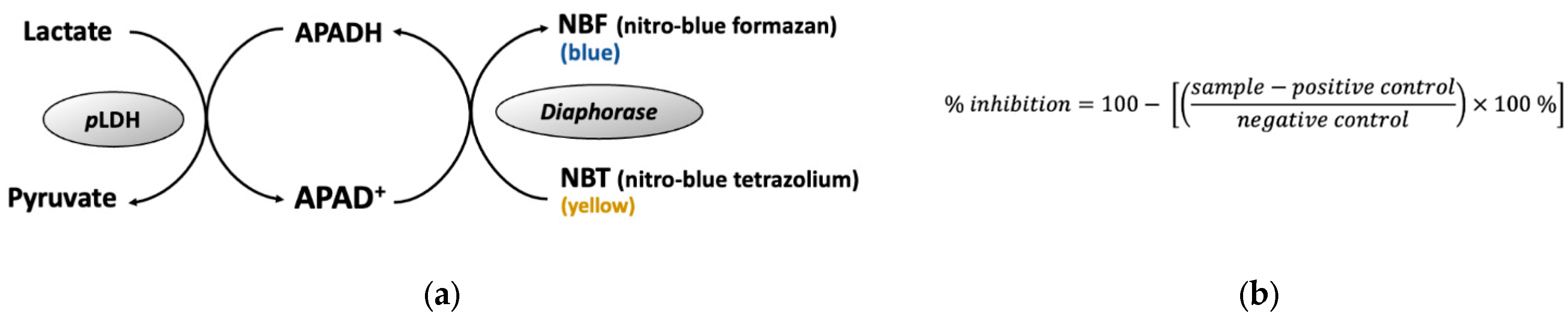
4.4. Lactate Dehydrogenase (pLDH) Assay
4.5. Preliminary Extraction Test
4.6. Thin Layer Chromatography (TLC)
4.7. Open Column Chromatography (OCC)
4.8. Analytical and Semi-Preparative HPLC
4.9. LC-MS Analysis
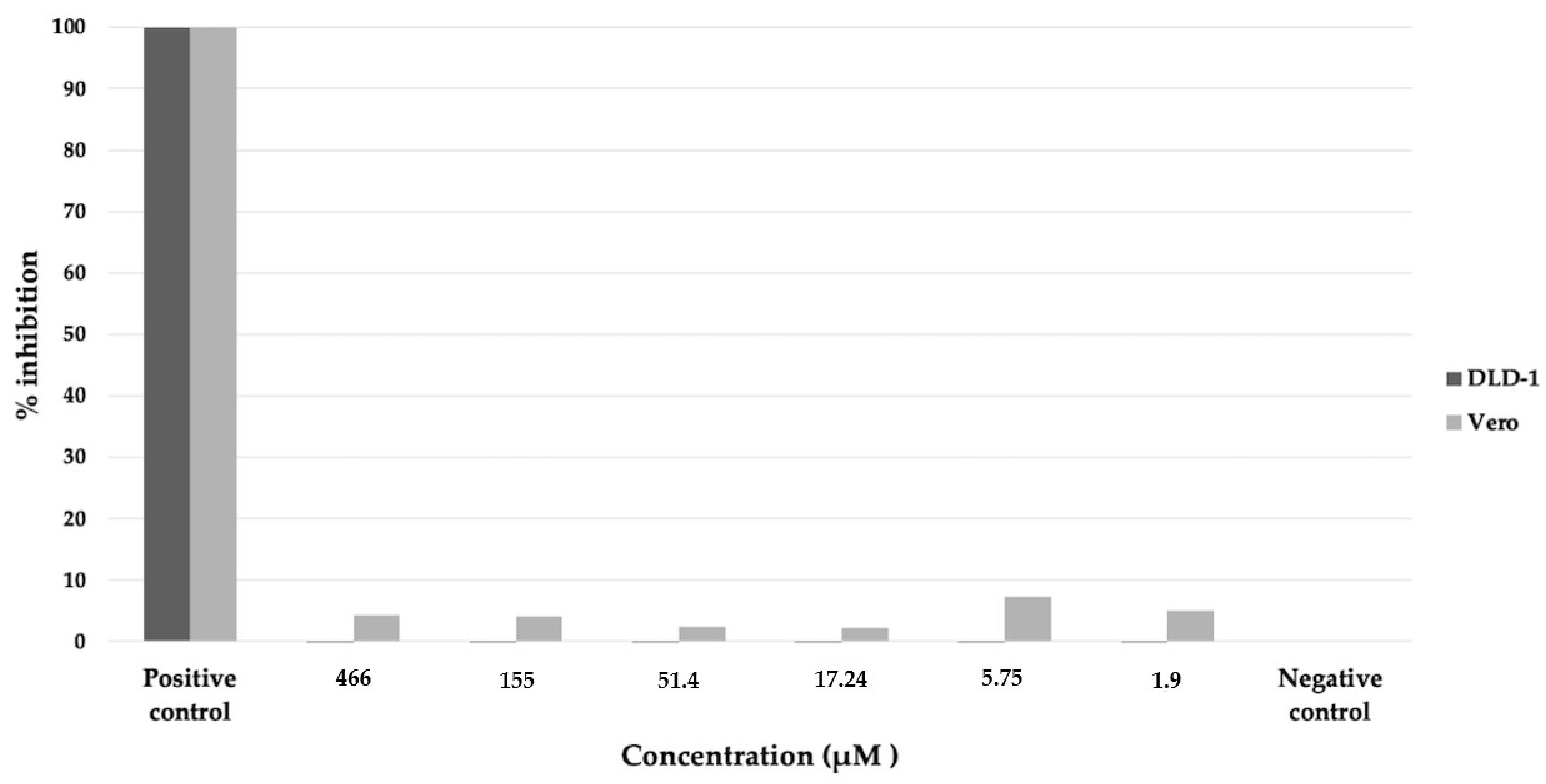
4.10. Cytotoxicity of Bioactive Compound on Mammalian Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report: 20 Years of Global Progress and Challenges; WHO: Geneva, Switzerland, 2020; ISBN 978-92-4-001579-1. [Google Scholar]
- Kumar, S.; Bhardwaj, T.R.; Prasad, D.N.; Singh, R.K. Drug targets for resistant malaria: Historic to future perspectives. Biomed. Pharmacother. 2018, 104, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.G.; Korsik, M.; Todd, M.H. The Past, Present and Future of Anti-Malarial Medicines. Malar. J. 2019, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report 2018; WHO: Geneva, Switzerland, 2018; ISBN 9789241565653. [Google Scholar]
- Van der Pluijm, R.W.; Tripura, R.; Hoglund, R.M.; Pyae Phyo, A.; Lek, D.; ul Islam, A.; Anvikar, A.R.; Satpathi, P.; Satpathi, S.; Behera, P.K.; et al. Triple artemisinin-based combination therapies versus artemisinin-based combination therapies for uncomplicated Plasmodium falciparum malaria: A multicentre, open-label, randomised clinical trial. Lancet 2020, 395, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Kokkonda, S.; El Mazouni, F.; White, K.L.; White, J.; Shackleford, D.M.; Lafuente-Monasterio, M.J.; Rowland, P.; Manjalanagara, K.; Joseph, J.T.; Garcia-Pérez, A.; et al. Isoxazolopyrimidine-Based Inhibitors of Plasmodium falciparum Dihydroorotate Dehydrogenase with Antimalarial Activity. ACS Omega 2018, 3, 9227–9240. [Google Scholar] [CrossRef] [PubMed]
- Llanos-Cuentas, A.; Casapia, M.; Chuquiyauri, R.; Hinojosa, J.C.; Kerr, N.; Rosario, M.; Toovey, S.; Arch, R.H.; Phillips, M.A.; Rozenberg, F.D.; et al. Antimalarial activity of single-dose DSM265, a novel plasmodium dihydroorotate dehydrogenase inhibitor, in patients with uncomplicated Plasmodium falciparum or Plasmodium vivax malaria infection: A proof-of-concept, open-label, phase 2a study. Lancet Infect. Dis. 2018, 18, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Miyazaki, Y.; Inaoka, D.K.; Hartuti, E.D.; Watanabe, Y.-I.I.; Shiba, T.; Harada, S.; Saimoto, H.; Burrows, J.N.; Benito, F.J.G.; et al. Identification of Plasmodium falciparum Mitochondrial Malate: Quinone Oxidoreductase Inhibitors from the Pathogen Box. Genes 2019, 10, 471. [Google Scholar] [CrossRef] [PubMed]
- Flannery, E.L.; Chatterjee, A.K.; Winzeler, E.A. Antimalarial drug discovery—Approaches and progress towards new medicines. Nat. Rev. Microbiol. 2017, 15, 572. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Ganesan, S.M.; Dass, S.; Morrisey, J.M.; Pou, S.; Nilsen, A.; Riscoe, M.K.; Mather, M.W.; Vaidya, A.B. Mitochondrial type II NADH dehydrogenase of Plasmodium falciparum (PfNDH2) is dispensable in the asexual blood stages. PLoS ONE 2019, 14, e0214023. [Google Scholar] [CrossRef] [PubMed]
- Hartuti, E.D.; Inaoka, D.K.; Komatsuya, K.; Miyazaki, Y.; Miller, R.J.; Xinying, W.; Sadikin, M.; Prabandari, E.E.; Waluyo, D.; Kuroda, M.; et al. Biochemical studies of membrane bound Plasmodium falciparum mitochondrial L-malate:quinone oxidoreductase, a potential drug target. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Waluyo, D.; Prabandari, E.E.; Pramisandi, A.; Hidayati, D.N.; Chrisnayanti, E.; Puspitasari, D.J.; Dewi, D.; Suryani; Kristiningrum; Oktaviani, A.N.; et al. Exploring natural microbial resources for the discovery of anti-malarial compounds. Parasitol. Int. 2021, 85, 102432. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, A.; Iwatsuki, M.; Namatame, M.; Nishihara-Tsukashima, A.; Sunazuka, T.; Takahashi, Y.; Ömura, S.; Otoguro, K. Borrelidin, a potent antimalarial: Stage-specific inhibition profile of synchronized cultures of Plasmodium falciparum. J. Antibiot. 2011, 64, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Sadorn, K.; Saepua, S.; Bunbamrung, N.; Boonyuen, N.; Komwijit, S.; Rachtawee, P.; Pittayakhajonwut, P. Diphenyl ethers and depsidones from the endophytic fungus Aspergillus unguis BCC54176. Tetrahedron 2022, 105, 132612. [Google Scholar] [CrossRef]
- Pramisandi, A.; Dobashi, K.; Mori, M.; Nonaka, K.; Matsumoto, A.; Tokiwa, T.; Higo, M.; Kristiningrum; Amalia, E.; Nurkanto, A.; et al. Microbial inhibitors active against plasmodium falciparum dihydroorotate dehydrogenase derived from an indonesian soil fungus, talaromyces pinophilus biomcc-f.T.3979. J. Gen. Appl. Microbiol. 2020, 66, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Dean, F.M.; Roberts, J.C.; Robertson, A. The chemistry of fungi. Part XXII. Nidulin and nornidulin (“ustin”): Chlorine-containing metabolic products of Aspergillus nidulans. J. Chem. Soc. 1954, 1432–1439. [Google Scholar] [CrossRef]
- Beach, W.F.; Richards, J.H. The Structure and Biosynthesis of Nidulin. J. Org. Chem. 1963, 28, 2746–2751. [Google Scholar] [CrossRef]
- Yang, W.C.; Bao, H.Y.; Liu, Y.Y.; Nie, Y.Y.; Yang, J.M.; Hong, P.Z.; Zhang, Y. Depsidone derivatives and a cyclopeptide produced by marine fungus aspergillus unguis under chemical induction and by its plasma induced mutant. Molecules 2018, 23, 2245. [Google Scholar] [CrossRef] [PubMed]
- Ten, L.; Domingos, S.; Martins, S.; Lima, L.M. De Secondary Metabolites Diversity of Aspergillus unguis and Their Bioactivities: A Potential Target to Be Explored. Biomolecules 2022, 12, 1820. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Van Anh, C.; Kwon, J.H.; Kang, J.S.; Lee, H.S.; Heo, C.S.; Shin, H.J. Antibacterial and Cytotoxic Phenolic Polyketides from Two Marine-Derived Fungal Strains of Aspergillus unguis. Pharmaceuticals 2022, 15, 74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A Simple Statistical Parameter for Use in Evaluation and Validation of High Throughput Screening Assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cahyono, A.W.; Fitri, L.E.; Winarsih, S.; Prabandari, E.E.; Waluyo, D.; Pramisandi, A.; Chrisnayanti, E.; Dewi, D.; Siska, E.; Nurlaila, N.; et al. Nornidulin, A New Inhibitor of Plasmodium falciparum Malate: Quinone Oxidoreductase (PfMQO) from Indonesian Aspergillus sp. BioMCC f.T.8501. Pharmaceuticals 2023, 16, 268. https://doi.org/10.3390/ph16020268
Cahyono AW, Fitri LE, Winarsih S, Prabandari EE, Waluyo D, Pramisandi A, Chrisnayanti E, Dewi D, Siska E, Nurlaila N, et al. Nornidulin, A New Inhibitor of Plasmodium falciparum Malate: Quinone Oxidoreductase (PfMQO) from Indonesian Aspergillus sp. BioMCC f.T.8501. Pharmaceuticals. 2023; 16(2):268. https://doi.org/10.3390/ph16020268
Chicago/Turabian StyleCahyono, Alfian Wika, Loeki Enggar Fitri, Sri Winarsih, Erwahyuni Endang Prabandari, Danang Waluyo, Amila Pramisandi, Evita Chrisnayanti, Diana Dewi, Eka Siska, Nurlaila Nurlaila, and et al. 2023. "Nornidulin, A New Inhibitor of Plasmodium falciparum Malate: Quinone Oxidoreductase (PfMQO) from Indonesian Aspergillus sp. BioMCC f.T.8501" Pharmaceuticals 16, no. 2: 268. https://doi.org/10.3390/ph16020268
APA StyleCahyono, A. W., Fitri, L. E., Winarsih, S., Prabandari, E. E., Waluyo, D., Pramisandi, A., Chrisnayanti, E., Dewi, D., Siska, E., Nurlaila, N., Nugroho, N. B., Nozaki, T., & Suciati, S. (2023). Nornidulin, A New Inhibitor of Plasmodium falciparum Malate: Quinone Oxidoreductase (PfMQO) from Indonesian Aspergillus sp. BioMCC f.T.8501. Pharmaceuticals, 16(2), 268. https://doi.org/10.3390/ph16020268